Cloning, Characterization and Expression Analysis of the Phosphate Starvation Response Gene, ClPHR1, from Chinese Fir

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Pi Treatment and Growth Conditions
2.2. RNA Extraction and Synthesis of First-Strand cDNA
2.3. Intermediate Conservative Region of ClPHR1Gene and the Full-Length cDNA of ClPHR1 Gene
2.4. Bioinformatics Analysis
2.5. Construction of ClPHR1 Transient Expression Vector and Subcellular Localization
2.6. Expression of the ClPHR1 Gene in Roots, Stems and Leaves of Chinese Fir Seedlings
2.7. Transformation of Arabidopsis and Identification of Transgenic Plants
2.8. Treatment of ClPHR1 Transgenic and Wild-type Arabidopsis and Expression Analysis of Phosphorous Starvation Response Genes
3. Results
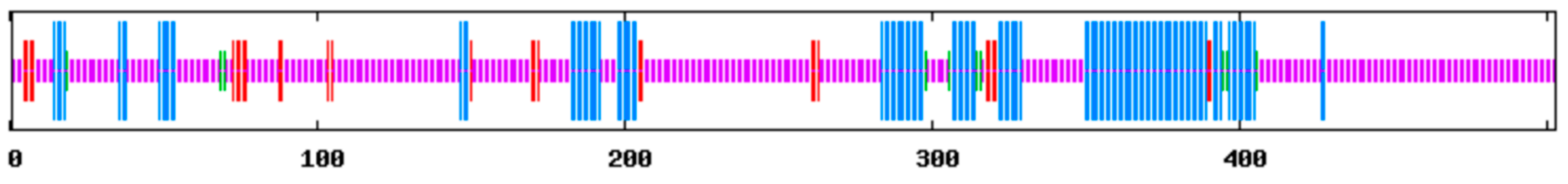
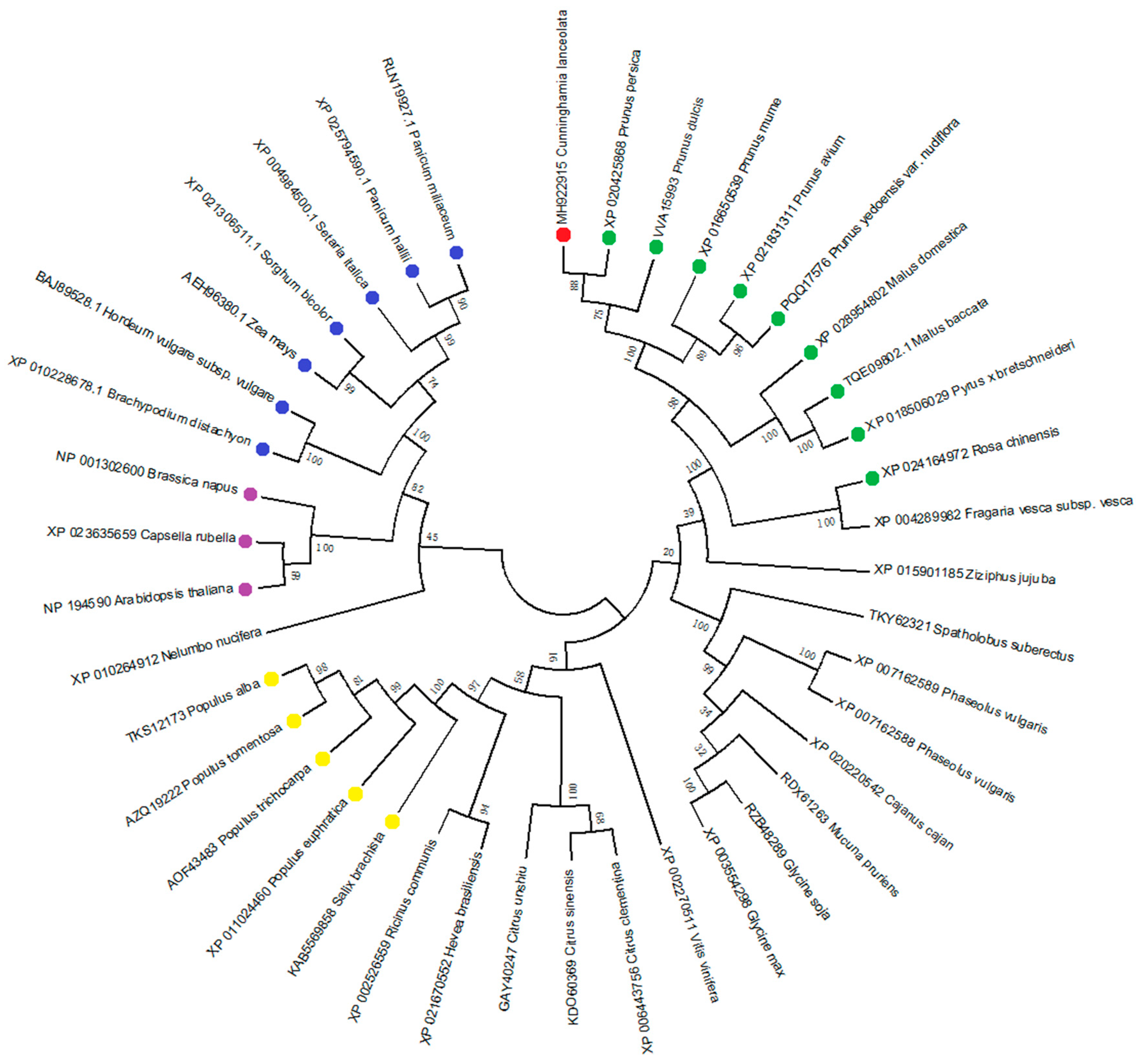
3.1. Cloning and Sequence Analysis of ClPHR1
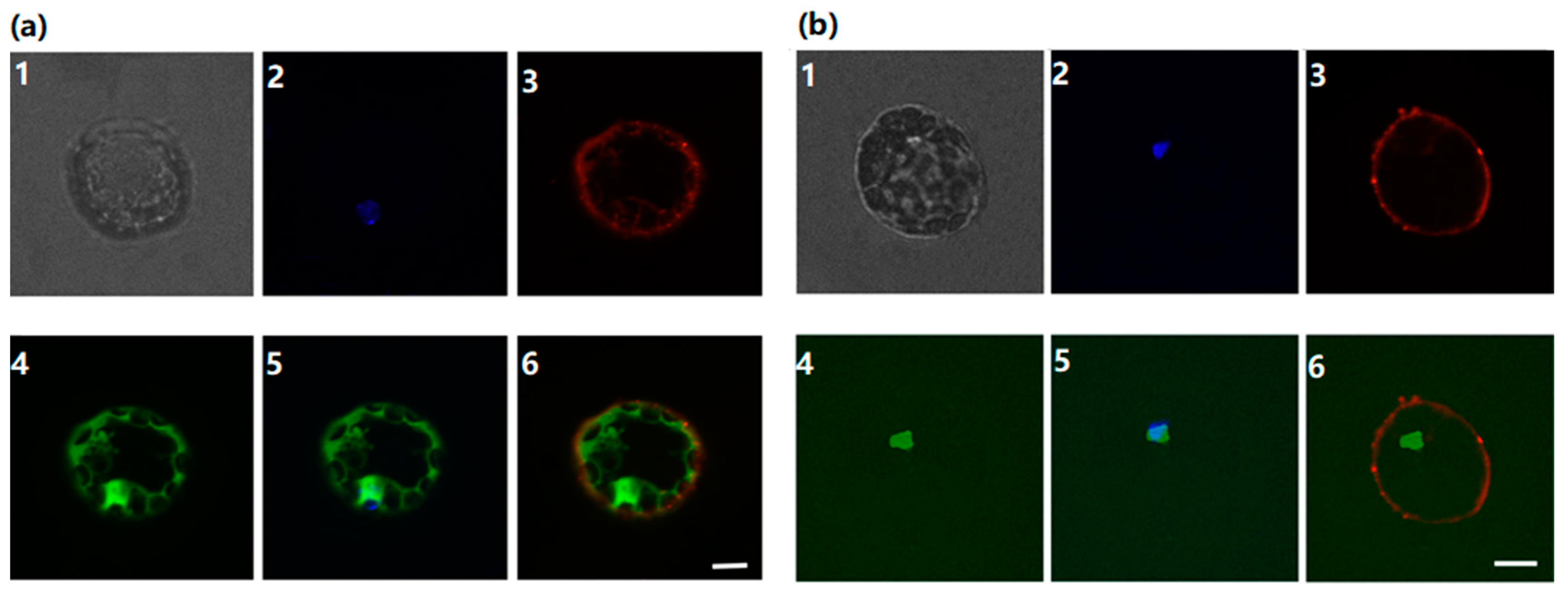
3.2. Subcellular Localization of ClPHR1 Gene
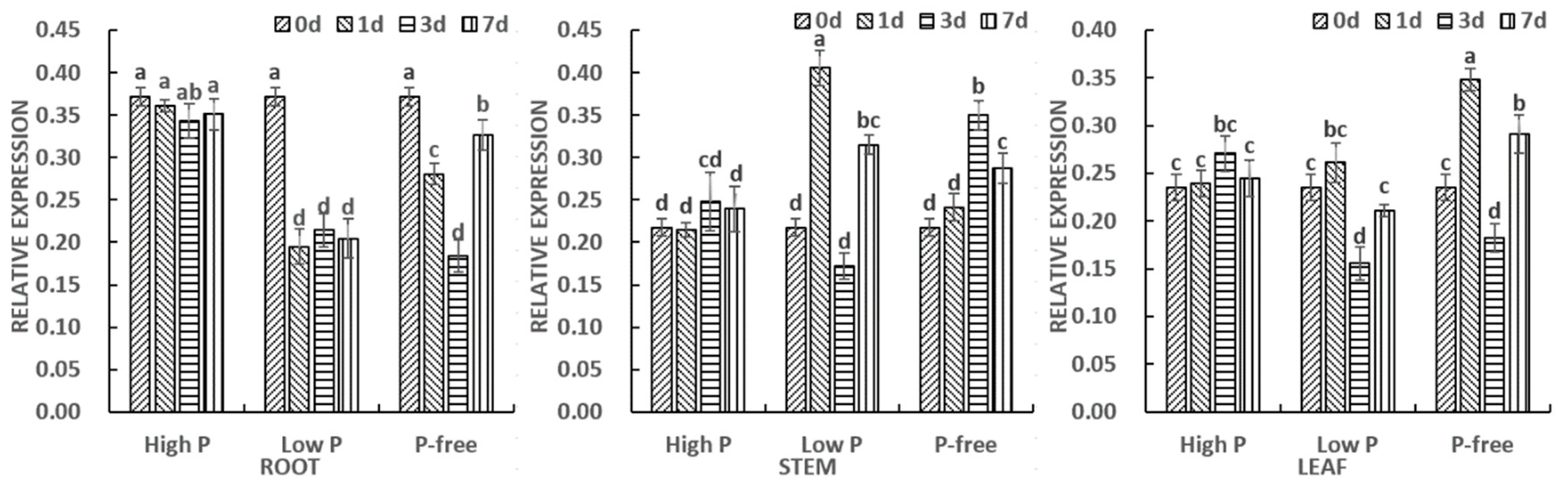
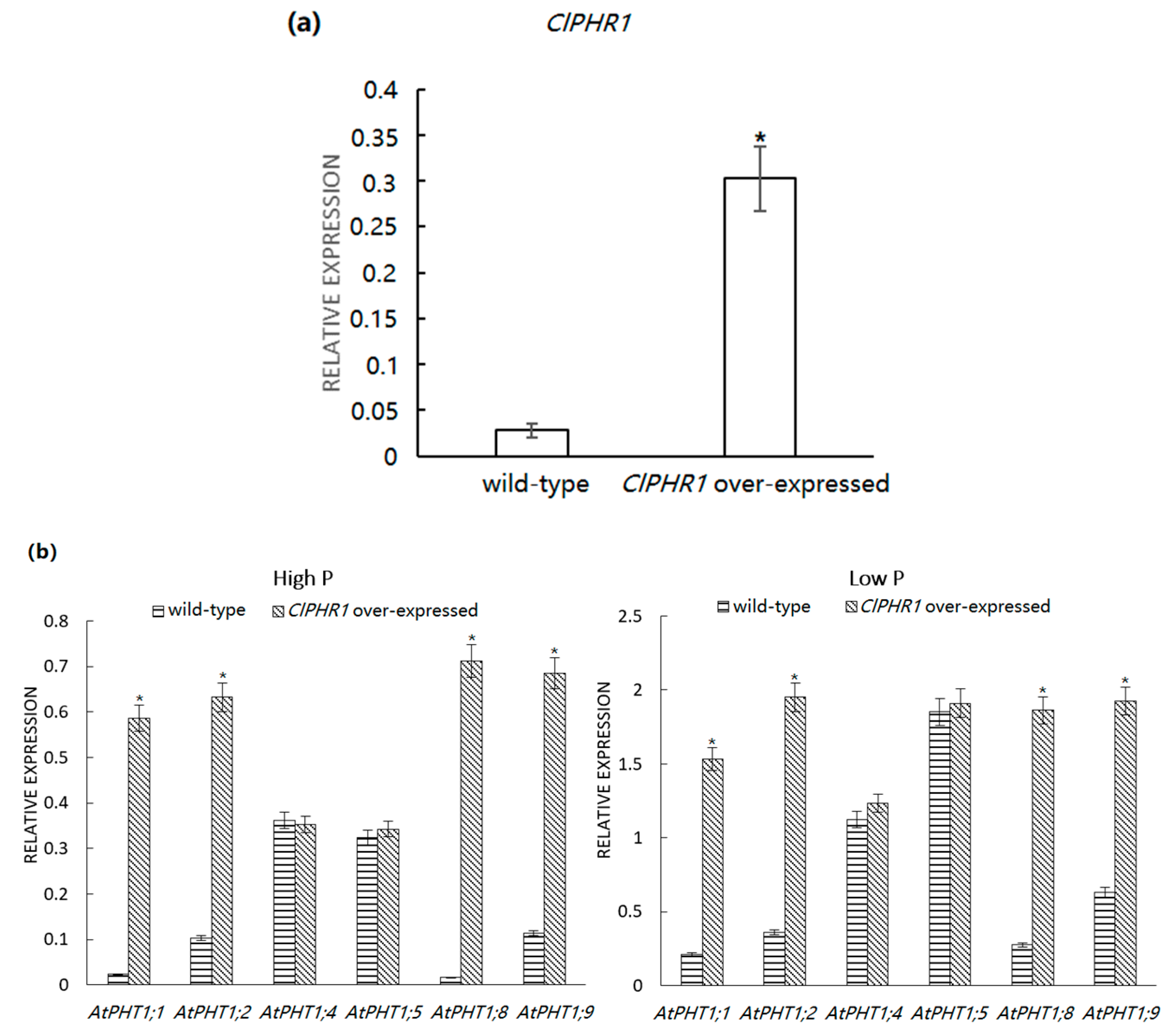
3.3. Expression Analysis of ClPHR1
3.4. Expression of Pi Transporter in ClPHR1-Overexpressing Transgenic Plants
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Bian, L.M.; Zheng, R.H.; Su, S.D.; Lin, H.Z.; Xiao, H.; Wu, H.X.; Shi, J. Spatial analysis increases efficiency of progeny testing of Chinese fir. J. For. Res. 2017, 28, 445–452. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.Q.; Chhin, S.; Fu, L.Y.; Lu, L.L.; Duan, A.G.; Zhang, J.G. Climate-sensitive tree height–diameter allometry for Chinese fir in southern China. For. Int. J. For. Res. 2019, 92, 167–176. [Google Scholar] [CrossRef]
- Wang, C.Q.; Xue, L.; Dong, Y.H.; Hou, L.Y.; Wei, Y.H.; Jiao, R.Z. Responses of soil microbial community structure to stand densities of Chinese fir plantations. J. For. Res. 2019, 24, 162–167. [Google Scholar] [CrossRef]
- Ma, X.Q.; Heal, K.V.; Liu, A.Q.; Jarvis, P.G. Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China. For. Ecol. Manag. 2007, 243, 61–74. [Google Scholar] [CrossRef]
- Lynch, J.P. Roots of the second green revolution. Aust. J. Bot. 2007, 55, 493–512. [Google Scholar] [CrossRef]
- Niu, Y.F.; Chai, R.S.; Jin, G.L.; Wang, H.; Tang, C.X.; Zhang, Y.S. Responses of root architecture development to low phosphorus availability: A review. Ann. Bot. 2013, 112, 391–408. [Google Scholar] [CrossRef] [PubMed]
- Farooq, T.H.; Wu, W.; Tigabu, M. Growth, Biomass Production and Root Development of Chinese fir in Relation to Initial Planting Density. Forests 2019, 10, 236. [Google Scholar] [CrossRef] [Green Version]
- Wang, C.Q.; Xue, L.; Dong, Y.H.; Hou, L.Y.; Wei, Y.H.; Chen, J.Q.; Jiao, R.Z. The development of Chinese fir plantations undergo significant changes in soil microbial metabolic function and enzyme activities. J. For. Res. 2019, 24, 261–265. [Google Scholar] [CrossRef]
- Li, L.; Luke, M.M.; Chen, F.S.; Wang, H.M.; Ma, Z.Q.; Guo, D.L. Different responses of absorptive roots and arbuscular mycorrhizal fungi to fertilization provide diverse nutrient acquisition strategies in Chinese fir. For. Ecol. Manag. 2019, 433, 64–72. [Google Scholar] [CrossRef]
- Bumb, B.L.; Baanante, C.A. The Role of Fertilizer in Sustaining Food Security and Protecting the Environment to 2020; Food, agriculture and the environment discussion paper 17; International Food Policy Research Institute Research Institute: Washington, DC, USA, 1996. [Google Scholar]
- Runge-Metzger, A. Closing the Cycle: Obstacles to Efficient P Management for Improved Global Food Security. In Phosphorus in the Global Environment: Transfers, Cycles and Management; Tiessen, H., Ed.; John Wiley and Sons Ltd.: Chichester, UK, 1995; pp. 27–42. [Google Scholar]
- Chen, Y.H.; Nguyen, T.H.N.; Qin, J.J.; Jiao, Y.; Li, Z.L.I.; Ding, S.; Lu, Y.; Liu, Q.F.; Luo, Z.B. Phosphorus assimilation of Chinese fir from two provenances during acclimation to changing phosphorus availability. Environ. Exp. Bot. 2018, 153, 21–34. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, L.Y.; Chen, S.Q.; Chen, Z.J.; Fan, Y.H. Study on Modification of Fast-growing Chinese Fir Stumpage. IOP Conf. Ser. Earth Environ. Sci. 2018, 108. [Google Scholar] [CrossRef]
- Zheng, H.Q.; Hu, D.H.; Wei, R.P.; Yan, S.; Wang, R.H. Chinese Fir Breeding in the High-Throughput Sequencing Era: Insights from SNPs. Forests 2019, 10, 681. [Google Scholar] [CrossRef] [Green Version]
- Plaxton, W.C.; Tran, H.T. FOCUS ISSUE ON PHOSPHORUS PLANT PHYSIOLOGY: Metabolic Adaptations of Phosphate-Starved Plants. Plant Cell 2011, 156, 1006–1015. [Google Scholar]
- Wang, F.; Deng, M.J.; Xu, J.M.; Zhu, X.L.; Mao, C.Z. Molecular mechanisms of phosphate transport and signaling in higher plants. Semin. Cell Dev. Biol. 2018, 74, 114–122. [Google Scholar] [CrossRef] [PubMed]
- Devaiah, B.N.; Karthikeyan, A.S.; Raghothama, K.G. WRKY75 transcription factor is a modulator of phosphate acquisition and root development in Arabidopsis(Article). Plant Physiol. 2007, 143, 1789–1801. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rishmawi, L.; Pesch, M.; Juengst, C.; Schauss, A.; Schrader, A.; Huelskamp, M. Non-cell-autonomous regulation of root hair patterning genes by WRKY75 in Arabidopsis thaliana. Plant Physiol. 2014, 165, 186–195. [Google Scholar] [CrossRef] [Green Version]
- Devaiah, B.N.; Nagarajan, V.K.; Raghothama, K.G. Phosphate homeostasis and root development in Arabidopsis are synchronized by the zinc finger transcription factor ZAT6(Article). Plant Physiol. 2007, 145, 147–159. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Nguyen, X.C.; Kim, K.E.; Han, H.J.; Yoo, J.; Lee, K.; Kim, M.C.; Yun, D.; Chung, W.S. Phosphorylation of the zinc finger transcriptional regulator ZAT6 by MPK6 regulates Arabidopsis seed germination under salt and osmotic stress. Biochem. Biophys. Res. Commun. 2013, 430, 1054–1059. [Google Scholar] [CrossRef]
- Devaiah, B.N.; Madhuvanthi, R.; Karthikeyan, A.S.; Raghothama, K.G. Phosphate Starvation Responses and Gibberellic Acid Biosynthesis Are Regulated by the MYB62 Transcription Factor in Arabidopsis. Mol. Plant 2009, 2, 43–58. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.; Ma, L.G.; Hou, X.L.; Wang, M.Y.; Wu, Y.R.; Liu, F.Y.; Wang, D.X. Phosphate starvation triggers distinct alterations of genome expression in Arabidopsis roots and leaves. Plant Physiol. 2003, 132, 1260–1271. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.H.; Jenkins, G.I.; Nimmo, H.G. Identification of an F-box protein that negatively regulates P(i) starvation responses. Plant Cell Physiol. 2008, 49, 1902–1906. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bournier, M.; Tissot, N.; Mari, S.; Boucherez, J.; Lacombe, E.; Briat, J.F.; Gaymard, F. Arabidopsis Ferritin 1 (AtFer1) Gene Regulation by the Phosphate Starvation Response 1 (AtPHR1) Transcription Factor Reveals a Direct Molecular Link between Iron and Phosphate Homeostasis. J. Biol. Chem. 2013, 288, 22670–22680. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.H.; Wang, Y.J.; Wu, B.; Kong, Y.B.; Li, W.L.; Chang, W.S.; Zhang, C.Y. GmPHR1, a Novel Homolog of the AtPHR1 Transcription Factor, Plays a Role in Plant Tolerance to Phosphate Starvation. Agric. Sci. China 2014, 13, 2584–2593. [Google Scholar] [CrossRef] [Green Version]
- Chiou, T.J.; Lin, S.I. Signaling Network in Sensing Phosphate Availability in Plants. Annu. Rev. Plant Biol. 2010, 62, 185–206. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Finnegan, P.M. Regulation of phosphate starvation responses in higher plants. Ann. Bot. 2010, 105, 513–526. [Google Scholar] [CrossRef] [PubMed]
- Rubio, V.; Linhares, F.; Solano, R.; Martín, A.C.; Iglesias, J.; Leyva, A.; Paz-Ares, J. A conserved MYB transcription factor involved in phosphate starvation signaling both in vascular plants and in unicellular algae. Genes Dev. 2001, 15, 2122–2133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nilsson, L.; MÜller, R.; Nielsen, T.H. Increased expression of the MYB-related transcription factor, PHR1, leads to enhanced phosphate uptake in Arabidopsis thaliana(Article). Plant Cell Environ. 2007, 30, 1499–1512. [Google Scholar] [CrossRef]
- Bari, R.; Pant, B.D.; Stitt, M.; Scheible, W.R. PHO2, microRNA399, and PHR1 define a phosphate-signaling pathway in plants(Article). Plant Physiol. 2006, 141, 988–999. [Google Scholar] [CrossRef] [Green Version]
- Tesfaye, M.; Liu, J.Q.; Allan, D.L.; Vance, C.P. FOCUS ISSUE ON LEGUME BIOLOGY: Genomic and Genetic Control of Phosphate Stress in Legumes. Plant Cell 2007, 144, 594–603. [Google Scholar]
- Motte, H.; Beeckman, T. PHR1 Balances between Nutrition and Immunity in Plants (Short Survey). Dev. Cell 2017, 41, 5–7. [Google Scholar] [CrossRef] [Green Version]
- Wu, P.F.; Ma, X.Q.; Chen, Y.L.; Lin, W.J.; Huang, S.Y.; Liu, L.Q. Comparison of phosphorus use efficiency among clonal test plantations of Chinese fir. J. Fujian Agric. For. Univ. 2012, 41, 40–45. [Google Scholar]
- Maas, C.; Werr, W. Mechanism and optimized conditions for PEG mediated DNA transfection into plant protoplasts. Plant Cell Rep. 1989, 8, 148–151. [Google Scholar] [CrossRef]
- Chen, R.; Chen, W.; Tigabu, M.; Zhong, W.; Li, Y.; Ma, X.; Li, M. Screening and Evaluation of Stable Reference Genesfor Quantitative Real-Time Polymerase ChainReaction (qRT-PCR) Analysis in Chinese Fir Rootsunder Water, Phosphorus, and Nitrogen Stresses. Forests 2019, 10, 1087. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Clough, S.J.; Bent, A.F. Floral dip: A simplified method forAgrobacterium-mediated transformation ofArabidopsis thaliana. Plant J. Cell Mol. Biol. 1998, 16, 735–743. [Google Scholar] [CrossRef] [Green Version]
- Kyte, J.; Doolittle, R.F. A Simple Method for Displaying the Hydropathic Character of a Protein. J. Mol. Biol. 1982, 157, 105–132. [Google Scholar] [CrossRef] [Green Version]
- Thomas, B.E.A. Regulation of Phosphate Starvation Response in Arabidopsis. Master’s Thesis, Texas A&M University, College Station, TX, USA, 2006. [Google Scholar]
- Dolores, G.A.; Odilón, O.R.J.; Lenin, Y.V.; Luis, C.T.; Luis, H.E. Adaptation to Phosphate Scarcity: Tips from Arabidopsis Roots. Trends Plant Sci. 2018, 23, 721–730. [Google Scholar]
- Osorio, M.B.B.; Ng, S.; Berkowitz, O.; Clercq, I.D.; Mao, C.; Shou, H.; Whelan, J.; Jost, R. SPX4 Acts on PHR1-Dependent and -Independent Regulation of Shoot Phosphorus Status in Arabidopsis. Plant Physiol. 2019, 181, 332–352. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, P.; Wang, X.M. Role of OsPHR2 on phosphorus homeostasis and root hairs development in rice (Oryza sativa L.). Plant Signal. Behav. 2008, 3, 674–675. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.H.; Bai, J.R.; Liu, H.M.; Sun, Y.; Shi, X.Y.; Ren, Z.Q. Overexpression of a Maize Transcription Factor ZmPHR1 Improves Shoot Inorganic Phosphate Content and Growth of Arabidopsis under Low-Phosphate Conditions. Plant Mol. Biol. Report. 2013, 31, 665–677. [Google Scholar] [CrossRef]
- Ren, F.; Guo, Q.Q.; Chang, L.L.; Chen, L.; Zhao, C.Z.; Zhong, H.; Li, X.B. Brassica napus PHR1 Gene Encoding a MYB-Like Protein Functions in Response to Phosphate Starvation. PLoS ONE 2012, 7, e44005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghothama, K.G. Phosphate acquisition. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999, 50, 665–693. [Google Scholar] [CrossRef] [PubMed]
- Li, L.H.; Qiu, X.H.; Li, X.H.; Wang, S.P.; Lian, X.M. The expression profile of genes in rice roots under low phosphorus stress. Sci. China Ser. C Life Sci. 2009, 52, 1055–1064. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Ma, X.; Tigabu, M.; Chen, W.; Liu, A.Q.; Odén, P.C. Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can. J. Res. 2011, 41, 228–234. [Google Scholar] [CrossRef]
- Chen, F.J.; Liu, Q.; Vogel, J.P.; Wu, J.J. Agrobacterium-Mediated Transformation of Brachypodiumdistachyon. Curr. Protoc. Plant Biol. 2019, 4, e20088. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Arabidopsis TAIR Gene Identifier | Primer Sequence (5′ to 3′) |
|---|---|---|
| Pht1;1 | AT5G43350 | F: AACGCCTCCTCAAGTTGACTAC |
| R: TGTTCTTGGCAACCAAAGCG | ||
| Pht1;2 | AT5G43370 | F: TTAGCGCACAACGGAAAGAC |
| R: ACGCATCGGTAAAGAAACCC | ||
| Pht1;4 | AT2G38940 | F: TTGGCTTGGATTTGGCATCG |
| R: AAAACCGCAGAGACAAAGGC | ||
| Pht1;5 | AT2G32830 | F: TGCGACGACGTTTGTTGTTC |
| R: ACCCAAACGCTCCAACAATC | ||
| Pht1;8 | AT1G20860 | F: TCTTGGTGGCGTTTGCATTG |
| R: TTGCCCGTCAACAATCTGTG | ||
| Pht1;9 | AT1G76430 | F: AGTTTGGTGCGATTGTTGGC |
| R: ACGCGATTCTCACACGTTTC |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, W.; Chen, R.; Zhang, Y.; Li, J.; Tigabu, M.; Ma, X.; Li, M. Cloning, Characterization and Expression Analysis of the Phosphate Starvation Response Gene, ClPHR1, from Chinese Fir. Forests 2020, 11, 104. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010104
Chen W, Chen R, Zhang Y, Li J, Tigabu M, Ma X, Li M. Cloning, Characterization and Expression Analysis of the Phosphate Starvation Response Gene, ClPHR1, from Chinese Fir. Forests. 2020; 11(1):104. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010104
Chicago/Turabian StyleChen, Wanting, Ranhong Chen, Ying Zhang, Jiaoyang Li, Mulualem Tigabu, Xiangqing Ma, and Ming Li. 2020. "Cloning, Characterization and Expression Analysis of the Phosphate Starvation Response Gene, ClPHR1, from Chinese Fir" Forests 11, no. 1: 104. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010104