A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum

 , , , , , , ,
, , , , , , , 
Abstract
:1. Introduction
2. Material and Methods
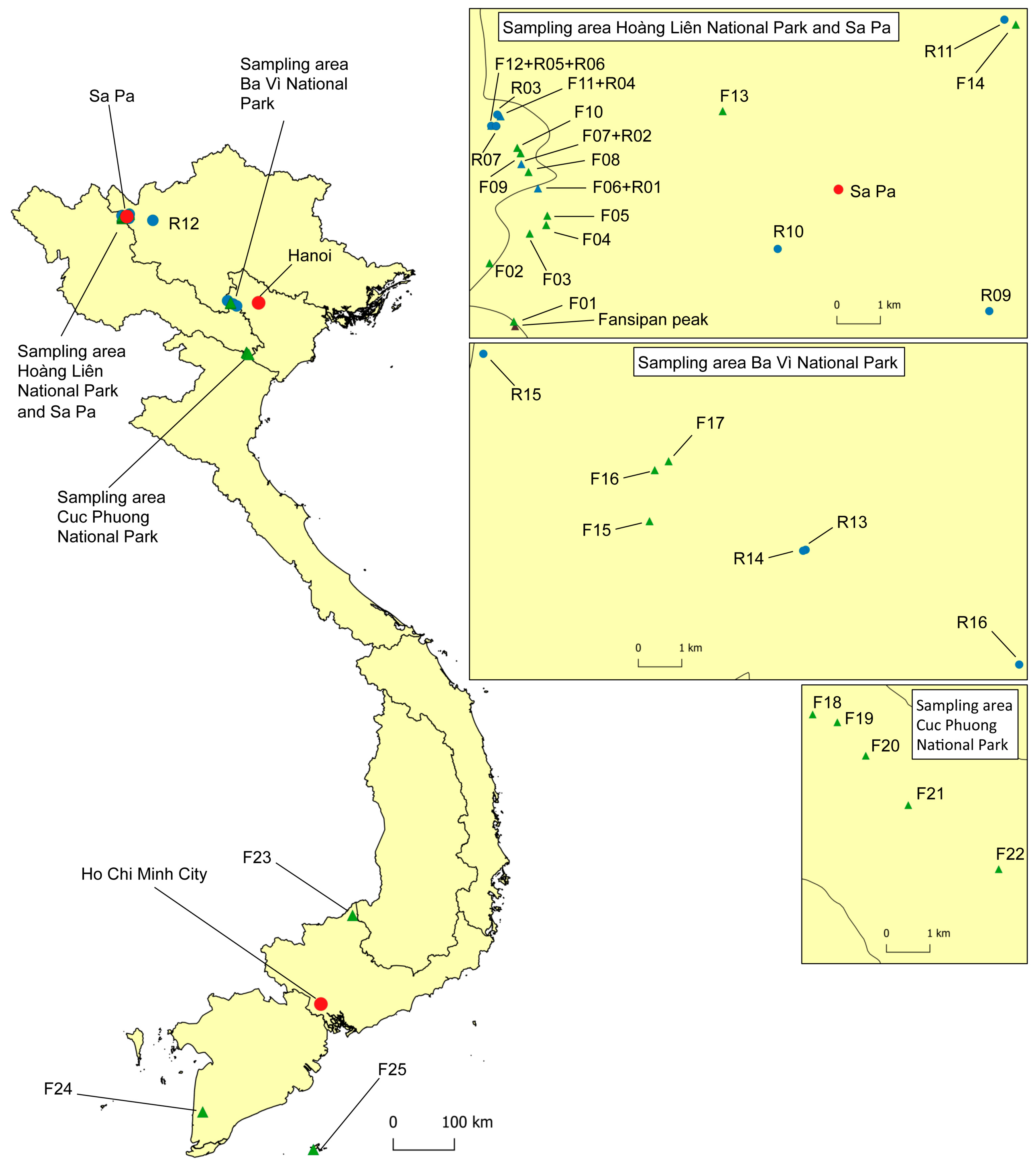
2.1. Sampling and Phytophthora Isolation
2.2. Molecular Identification of Isolates
2.3. Classical Identification of Isolates
3. Results
3.1. Phytophthora Diversity in Natural and Semi-Natural Forest Stands
3.2. Phytophthora Diversity in Natural Forest Streams and Rivers
3.3. Association between Phytophthora Presence in the Rhizosphere and Disease Symptoms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Brasier, C.M.; Robredo, F.; Ferraz, J.F.P. Evidence for Phytophthora cinnamomi involvement in Iberian oak decline. Plant Pathol. 1993, 42, 140–145. [Google Scholar] [CrossRef]
- Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide; APS Press: Saint Paul, MN, USA, 1996. [Google Scholar]
- Hansen, E.M.; Goheen, D.J.; Jules, E.S.; Ullian, B. Managing Port–Orford–Cedar and the introduced pathogen Phytophthora lateralis. Plant Dis. 2000, 84, 4–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; Blaschke, H.; Osswald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Jung, T.; Vettraino, A.M.; Cech, T.L.; Vannini, A. The impact of invasive Phytophthora species on European forests. Phytophthora: A Global Perspective; Lamour, K., Ed.; CABI: Wallingford, UK, 2013; pp. 146–158. ISBN 978-1-78064-093-8. [Google Scholar]
- Jung, T.; Pérez–Sierra, A.; Durán, A.; Horta Jung, M.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Persoonia 2018, 40, 182–220. [Google Scholar] [CrossRef] [Green Version]
- Rizzo, D.M.; Garbelotto, M.; Davidson, J.M.; Slaugter, G.W. Phytophthora ramorum as the cause of extensive mortality of Quercus spp. and Lithocarpus densiflorus in California. Plant Dis. 2002, 86, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Hardham, A.R. Phytophthora cinnamomi. Mol. Plant Pathol. 2005, 6, 589–604. [Google Scholar] [CrossRef]
- Cahill, D.M.; Rookes, J.E.; Wilson, B.A.; Gibson, L.; Mcdougall, K.L. Turner Review No. 17. Phytophthora cinnamomi and Australia’s biodiversity: Impacts, predictions and progress towards control. Aust. J. Bot. 2008, 56, 279–310. [Google Scholar] [CrossRef]
- Jung, T.; Chang, T.T.; Bakonyi, J.; Seress, D.; Pérez-Sierra, A.; Yang, X.; Hong, C.; Scanu, B.; Fu, C.H.; Hsueh, K.-L.; et al. Diversity of Phytophthora species in natural ecosystems of Taiwan and association with disease symptoms. Plant Pathol. 2017, 66, 194–211. [Google Scholar] [CrossRef]
- Jung, T.; Durán, A.; Sanfuentes von Stowasser, E.; Schena, L.; Mosca, S.; Fajardo, S.; González, M.; Navarro Ortega, A.D.; Bakonyi, J.; Seress, D.; et al. Diversity of Phytophthora species in Valdivian rainforests and association with severe dieback symptoms. Forest Pathol. 2018, 48, e12443. [Google Scholar] [CrossRef]
- Zeng, H.-C.; Ho, H.-H.; Zheng, F.-C. A survey of Phytophthora species on Hainan Island of South China. J. Phytopathol. 2009, 157, 33–39. [Google Scholar] [CrossRef]
- Brasier, C.M.; Vettraino, A.M.; Chang, T.T.; Vannini, A. Phytophthora lateralis discovered in an old growth Chamaecyparis forest in Taiwan. Plant Pathol. 2010, 59, 595–603. [Google Scholar] [CrossRef]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef] [PubMed]
- Vettraino, A.M.; Brasier, C.M.; Brown, A.V.; Vannini, A. Phytophthora himalsilva sp. nov. an unusually phenotypically variable species from a remote forest in Nepal. Fungal Biol. 2011, 115, 275–287. [Google Scholar] [CrossRef] [PubMed]
- Huai, W.X.; Tian, G.; Hansen, E.M.; Zhao, W.X.; Goheen, E.M.; Grünwald, N.J.; Cheng, C. Identification of Phytophthora species baited and isolated from forest soil and streams in northwestern Yunnan province, China. Forest Pathol. 2013, 43, 87–103. [Google Scholar] [CrossRef]
- Hüberli, D.; Hardy, G.E.S.T.J.; White, D.; Williams, N.; Burgess, T.I. Fishing for Phytophthora from Western Australia’ s waterways: A distribution and diversity survey. Australas. Plant Pathol. 2013, 42, 251–260. [Google Scholar] [CrossRef]
- Oh, E.; Gryzenhout, M.; Wingfield, B.D.; Wingfield, M.J.; Burgess, T.I. Surveys of soil and water reveal a goldmine of Phytophthora diversity in South African natural ecosystems. IMA Fungus 2013, 4, 123–131. [Google Scholar] [CrossRef] [PubMed]
- Burgess, T.I.; White, D.; McDougall, K.M.; Garnas, J.; Dunstan, W.A.; Català, S.; Carnegie, A.J.; Worboys, S.; Cahill, D.; Vettraino, A.M.; et al. Distribution and diversity of Phytophthora across Australia. Pac. Conserv. Biol. 2017, 23, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Hansen, E.M.; Winton, L.; Oßwald, W.; Delatour, C. Three new species of Phytophthora from European oak forests. Mycol. Res. 2002, 106, 397–411. [Google Scholar] [CrossRef]
- Jung, T.; Stukely, M.J.C.; Hardy, G.E.S.J.; White, D.; Paap, T.; Dunstan, W.A.; Burgess, T.I. Multiple new Phytophthora species from ITS Clade 6 associated with natural ecosystems in Australia: Evolutionary and ecological implications. Persoonia 2011, 26, 13–39. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Horta Jung, M.; Scanu, B.; Seress, D.; Kovács, D.M.; Maia, C.; Pérez-Sierra, A.; Chang, T.-T.; Chandelier, A.; Heungens, A.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Persoonia 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Horta Jung, M.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, T.I.; Webster, J.L.; Ciampini, J.A.; White, D.; Hardy, G.E.S.J.; Stukely, M.J.C. Re-evaluation of Phytophthora species isolated during 30 years of vegetation health surveys in Western Australia using molecular techniques. Plant Dis. 2009, 93, 215–223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Burgess, T.I.; Simamora, A.V.; White, D.; Wiliams, B.; Schwager, M.; Stukely, M.J.C.; Hardy, G.E.S.J. New species from Phytophthora Clade 6a: Evidence for recent radiation. Persoonia 2018, 41, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Brasier, C. Phytophthora biodiversity: How many Phytophthora species are there? In Phytophthoras in Forests and Natural Ecosystems: Fourth Meeting of the International Union of Forest Research Organizations (IUFRO) Working Party S07.02.09, USDA Forest Service; Goheen, E.M., Frankel, S.J., Eds.; Pacific Southwest Research Station: Albany, NY, USA, 2009. [Google Scholar]
- Kroon, L.P.; Brouwer, H.; de Cock, A.W.; Govers, F. The genus Phytophthora anno 2012. Phytopathology 2012, 102, 348–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Tyler, B.M.; Hong, C. An expanded phylogeny for the genus Phytophthora. IMA Fungus 2017, 8, 355–384. [Google Scholar] [CrossRef] [Green Version]
- Cooke, D.E.L.; Drenth, A.; Duncan, J.M.; Wagels, G.; Brasier, C.M. A molecular phylogeny of Phytophthora and related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Ko, W.H.; Chang, H.S.; Su, H.J. Isolates from Phytophthora cinnamomi from Taiwan as evidence for an Asian origin of the species. Trans. Br. Mycol. Soc. 1978, 71, 496–499. [Google Scholar] [CrossRef]
- Ko, W.H.; Wang, S.Y.; Ann, P.J. The possible origin and relation of Phytophthora katsurae and P. heveae, discovered in a protected natural forest in Taiwan. Bot. Stud. 2006, 47, 273–277. [Google Scholar]
- Chang, T.T.; Wang, W.W.; Wang, W.Y. Use of random amplified polymorphic DNA markers for the detection of genetic variation in Phytophthora cinnamomi in Taiwan. Bot. Bull. Acad. Sin. 1996, 37, 165–171. Available online: https://ejournal.sinica.edu.tw/bbas/content/1996/3/bot373-01.html (accessed on 16 December 2019).
- Ho, H.H.; Lu, J.Y. A synopsis of the occurrence and pathogenicity of Phytophthora species in mainland China. Mycopathologia 1997, 138, 143–161. [Google Scholar] [CrossRef]
- Drenth, A.; Guest, D.I. Diversity and management of Phytophthora in Southeast Asia; ACIAR Monograph 114; Australian Centre for International Agricultural Research: Canberra, Australia, 2004; p. 238. ISBN 1-86320-405-9.
- Ann, P.J.; Wong, I.T.; Tsai, J.N. New records of Phytophthora diseases of aromatic crops in Taiwan. Plant Pathol. Bull. 2010, 19, 53–68. [Google Scholar]
- Chang-Fu, H.; Chung-Fu, S. Introduction to the flora of Taiwan, 1: Geography, Geology, Climate, and Soils. Flora of Taiwan Second Edition. Editorial Committee of the Flora of Taiwan Second Edition. 1994. Available online: http://tai2.ntu.edu.tw/ebook/ebookpage.php?volume=1&book=Fl.%20Taiwan%202nd%20edit.&page=1 (accessed on 11 January 2020).
- Chang-Fu, H.; Chung-Fu, S.; Kuoh-Cheng, Y. Introduction to the Flora of Taiwan, 3: Floristics, Phytogeography, and Vegetation. Flora of Taiwan Second Edition. Editorial Committee of the Flora of Taiwan Second Edition. 1994. Available online: http://tai2.ntu.edu.tw/ebook/ebookpage.php?book=Fl.%20Taiwan%202nd%20edit.&volume=1&page=7 (accessed on 11 January 2020).
- Chung-Fu, S. Introduction to the flora of Taiwan, 2: Geotectonic Evolution, Paleogeography, and the Origin of the Flora. Flora of Taiwan Second Edition. Editorial Committee of the Flora of Taiwan Second Edition. 1994. Available online: http://tai2.ntu.edu.tw/ebook/ebookpage.php?book=Fl.%20Taiwan%202nd%20edit.&volume=1&page=3 (accessed on 11 January 2020).
- Gower, D.J.; Johnson, K.G.; Richardson, J.E.; Rosen, B.R.; Rüber, L.; Williams, S.T. Biotic Evolution and Environmental Change in Southeast Asia; The Systematics Association Special Volume 82; Cambridge University Press: New York, NY, USA, 2012; ISBN 13- 978-1107001305. [Google Scholar]
- Averyanov, L.V.; Loc, P.K.; Hiep, N.T.; Harder, D.K. Phytogeographic review of Vietnam and adjacent areas of Eastern Indochina. Komarovia 2003, 3, 1–83. [Google Scholar]
- Wurster, C.M.; Bird, M.I.; Bull, I.D.; Creed, F.; Bryant, C.; Dungait, J.A.J.; Paz, V. Forest contraction in north equatorial Southeast Asia during the last glacial maximum. Proc. Natl. Acad. Sci. USA 2010, 107, 15508–15511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T. Beech decline in Central Europe driven by the interaction between Phytophthora infections and climatic extremes. Forest Pathol. 2009, 39, 73–94. [Google Scholar] [CrossRef]
- Jung, T.; Burgess, T.I. Re-evaluation of Phytophthora citricola isolates from multiple woody hosts in Europe and North America reveals a new species, Phytophthora plurivora sp. nov. Persoonia 2009, 22, 95–110. [Google Scholar] [CrossRef] [Green Version]
- Hansen, E.M.; Reeser, P.W.; Sutton, W. Phytophthora beyond agriculture. Annu. Rev. Phytopathol. 2012, 50, 359–378. [Google Scholar] [CrossRef]
- Scanu, B.; Hunter, G.C.; Linaldeddu, B.T.; Franceschini, A.; Maddau, L.; Jung, T.; Denman, S. A taxonomic re-evaluation reveals that Phytophthora cinnamomi and P. cinnamomi var. parvispora are separate species. Forest Pathol. 2014, 44, 1–20. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. ISBN 0123721806. [Google Scholar]
- Kroon, L.P.N.M.; Bakker, F.T.; van den Bosch, G.B.M.; Bonants, P.J.M.; Flier, W.G. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef]
- Martin, F.N.; Tooley, P.W. Phylogenetic relationships among Phytophthora species inferred from sequence analysis of mitochondrially encoded cytochrome oxidase I and II genes. Mycologia 2003, 95, 269–284. [Google Scholar] [CrossRef]
- Aragaki, M.; Uchida, J.Y. Morphological distinctions between Phytophthora capsici and P. tropicalis sp. nov. Mycologia 2001, 93, 137–145. [Google Scholar] [CrossRef]
- Ilieva, E.; Man In’t Veld, W.A.; Veenbaas-Rijks, W.; Pieters, R. Phytophthora multivesiculata, a new species causing rot in Cymbidium. Eur. J. Plant Pathol. 1998, 104, 677–684. [Google Scholar] [CrossRef]
- Jung, T.; Scanu, B.; Bakonyi, J.; Seress, D.; Kovács, G.M.; Durán, A.; Sanfuentes von Stowasser, E.; Schena, L.; Mosca, S.; Thu, P.Q.; et al. Nothophytophthora gen. nov., a new sister genus of Phytophthora from natural and semi-natural ecosystems. Persoonia 2017, 39, 143–174. [Google Scholar] [CrossRef] [Green Version]
- Werres, S.; Marwitz, R.; Man In’t Veld, W.A.M.; Bonants, P.J.M.; De Weerd, M.; Themann, K.; Ilieva, E.; Baayen, R.P. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and Viburnum. Mycol. Res. 2001, 105, 1155–1165. [Google Scholar] [CrossRef]
- Brasier, C.M.; Cooke, D.E.L.; Duncan, J.M.; Hansen, E.M. Multiple new phenotypic taxa from trees and riparian ecosystems in Phytophthora gonapodyides–P. megasperma ITS Clade 6, which tend to be high-temperature tolerant and either inbreeding or sterile. Mycol. Res. 2003, 107, 277–290. [Google Scholar] [CrossRef]
- Safaiefarahani, B.; Mostowfizadeh-Ghalamfarsa, R.; Hardy, G.E.S.J.; Burgess, T.I. Re-evaluation of the Phytophthora cryptogea species complex and the description of a new species, Phytophthora pseudocryptogea sp. nov. Mycol. Prog. 2015, 14, 108. [Google Scholar] [CrossRef]
- Jung, T.; Nechwatal, J. Phytophthora gallica sp. nov., a new species from rhizosphere soil of declining oak and reed stands in France and Germany. Mycol. Res. 2008, 112, 1195–1205. [Google Scholar] [CrossRef]
- Yang, X.; Hong, C. Phytophthora virginiana sp. nov., a high-temperature tolerant species from irrigation water in Virginia. Mycotaxon 2013, 126, 167–176. [Google Scholar] [CrossRef]
- Balci, Y.; Balci, S.; Eggers, J.; MacDonald, W.L.; Juzwik, J.; Long, R.P.; Gottschalk, K.W. Phytophthora spp. associated with forest soils in eastern and north-central U.S. oak ecosystems. Plant Dis. 2007, 91, 705–710. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Horta Jung, M.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and distribution of Phytophthora species in protected natural areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Milenković, I.; Keča, N.; Karadžić, D.; Radulović, Z.; Nowakowska, J.A.; Oszako, T.; Sikora, K.; Corcobado, T.; Jung, T. Isolation and pathogenicity of Phytophthora species from poplar plantations in Serbia. Forests 2018, 9, 330. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Aguín Casal, O.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2016, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, S.K.; Zhou, Y.; Lamour, K. Oomycetes baited from streams in Tennessee 2010–2012. Mycologia 2013, 105, 1516–1523. [Google Scholar] [CrossRef]
- Brasier, C.M. Evolutionary Biology of Phytophthora. I. Genetic system, sexuality and variation. Annu. Rev. Phytopathol. 1992, 30, 153–171. [Google Scholar] [CrossRef]
- Català, S.; Peréz-Sierra, A.; Abad-Campos, P. The use of genus-specific amplicon pyrosequencing to assess Phytophthora species diversity using eDNA from soil and water in Northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, N.J.; Garbelotto, M.; Goss, E.M.; Heungens, K.; Prospero, S. Emergence of the sudden oak death pathogen Phytophthora ramorum. Trends Microbiol. 2012, 20, 131–138. [Google Scholar] [CrossRef]
- Van Poucke, K.; Franceschini, S.; Webber, J.F.; Vercauteren, A.; Turner, J.A.; McCracken, A.R.; Heungens, K.; Brasier, C.M. Discovery of a fourth evolutionary lineage of Phytophthora ramorum: EU2. Fungal Biol. 2012, 116, 1178–1191. [Google Scholar] [CrossRef]
- Arentz, F.; Simpson, J.A. Distribution of Phytophthora cinnamomi in Papua New Guinea and notes on its origin. Trans. Br. Mycol. Soc. 1986, 87, 289–295. [Google Scholar] [CrossRef]
- Brasier, C.M.; Scott, J.K. European oak declines and global warming: A theoretical assessment with special reference to the activity of Phytophthora cinnamomi. Bull. OEPP EPPO Bull. 1994, 24, 221–234. [Google Scholar] [CrossRef]
- Brasier, C.M. Phytophthora cinnamomi and oak decline in southern Europe. Environmental constraints including climate change. Ann. Des. Sci. For. 1996, 53, 347–358. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; Scott, J.K.; McDougall, K.L.; Stukely, M.J.C.; Crane, C.; Dunstan, W.A.; Brigg, F.; Andjic, V.; White, D.; Rudman, T.; et al. Current and projected global distribution of Phytophthora cinnamomi, one of the world’s worst plant pathogens. Glob. Chang. Biol. 2017, 23, 1661–1674. [Google Scholar] [CrossRef] [Green Version]
- Shearer, B.L.; Crane, C.E.; Cochrane, A. Quantification of the susceptibility of the native flora of the South-West Botanical Province, Western Australia, to Phytophthora cinnamomi. Aust. J. Bot. 2004, 52, 435–443. [Google Scholar] [CrossRef]
- Hardham, A.R.; Blackman, L.M. Phytophthora cinnamomi. Mol. Plant Pathol. 2018, 19, 260–285. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zentmyer, G.A. Phytophthora Cinnamomi and the Diseases it Causes; Monograph No. 10; The American Phytopathological Society: Saint Paul, MN, USA, 1980; ISBN 0890540306. [Google Scholar]
- Linde, C.; Drenth, A.; Kemp, G.H.J.; Wingfield, M.J.; Von Broembsen, S.L. Population structure of Phytophthora cinnamomi in South Africa. Phytopathology 1997, 87, 822–827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dobrowolski, M.P.; Tommerup, I.C.; Blakeman, H.D.; O’Brien, P.A. Non-Mendelian inheritance revealed in a genetic analysis of sexual progeny of Phytophthora cinnamomi with microsatellite markers. Fungal Genet. Biol. 2002, 35, 197–212. [Google Scholar] [CrossRef] [PubMed]
- Dobrowolski, M.P.; Tommerup, I.C.; Shearer, B.L.; O’Brien, P.A. Three clonal lineages of Phytophthora cinnamomi in Australia revealed by microsatellites. Phytopathology 2003, 93, 695–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; Colquhoun, I.J.; Hardy, G.E.S.J. New insights into the survival strategy of the invasive soilborne pathogen Phytophthora cinnamomi in different natural ecosystems in Western Australia. Forest Pathol. 2013, 43, 266–288. [Google Scholar] [CrossRef]
- Bezuidenhout, C.M.; Denman, S.; Kirk, S.A.; Botha, W.J.; Mostert, L.; McLeod, A. Phytophthora taxa associated with cultivated Agathosma, with emphasis on the P. citricola complex and P. capensis sp. nov. Persoonia 2010, 25, 32–49. [Google Scholar] [CrossRef]
- Nagel, J.H.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J.; Hardy, G.E.S.J.; Stukely, M.; Burgess, T.I. Characterization of Phytophthora hybrids from ITS clade 6 associated with riparian ecosystems in South Africa and Australia. Fungal Biol. 2013, 117, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I. Molecular characterization of natural hybrids formed between five related indigenous Clade 6 Phytophthora species. PLoS ONE 2015, 10, e0134225. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Maia, C.; Horta Jung, M. Host range testing of known and novel Phytophthora species from Asia, Central and South America among major forest tree species from Europe. Unpublished work. 2020. [Google Scholar]
- Eschen, R.; Douma, J.C.; Grégoire, J.-C.; Mayer, F.; Rigaux, L.; Potting, R.P.J. A risk categorisation and analysis of the geographic and temporal dynamics of the European import of plants for planting. Biol. Invasions 2017, 19, 3243–3257. [Google Scholar] [CrossRef]
- Rahman, M.Z.; Uematsu, S.; Takeuchi, T.; Shirai, K.; Ishiguro, Y.; Suga, H.; Kageyama, K. Two new species, Phytophthora nagaii sp. nov. and P. fragariaefolia sp. nov., causing serious diseases on rose and strawberry plants, respectively, in Japan. J. Gen. Plant Pathol. 2014, 80, 348–365. [Google Scholar] [CrossRef]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; De Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gaitniekis, T.; et al. Biogeographic patterns and determinants of invasion by alien forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liebhold, A.M.; Brockerhoff, E.G.; Garrett, L.J.; Parke, J.L.; Britton, K.O. Live plant imports: The major pathway for forest insect and pathogen invasions of the US. Front. Ecol. Environ. 2012, 10, 135–143. [Google Scholar] [CrossRef]
- Eschen, R.; Rigaux, L.; Sukovata, L.; Vettraino, A.M.; Marzano, M.; Grégoire, J.-C. Phytosanitary inspection of woody plants for planting at European Union entry points: A practical enquiry. Biol. Invasions 2015, 17, 2403–2413. [Google Scholar] [CrossRef] [Green Version]
- Eschen, R.; Britton, K.; Brockerhoff, E.; Burgess, T.; Dalley, V.; Epanchin-Niell, R.; Gupta, K.; Hardy, G.; Huang, Y.; Kenis, M.; et al. International variation in phytosanitary legislation and regulations governing importation of plants for planting. Environ. Sci. Policy 2015, 51, 228–237. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site no. | GPS Coordinates | Altitude (m a.s.l) | Location | Geological Substrate | Vegetation | Sampled Tree Species (no. of Phytophthora-Positive/Sampled Trees) | Phytophthora and Nothophytophthora spp. (no. of Positive Samples) a,b |
|---|---|---|---|---|---|---|---|
| F01 | N22 18.466 E103 46.480 | 2903 | Fansipan, Hoàng Liên National Park (NP) | Triassic schists and sandstones | Subalpine Rhododendron scrub | Rhododendron spp. (0/3) | - |
| F02 | N22 19.194 E103 46.177 | 2636 | Fansipan, Hoàng Liên NP | Triassic schists and sandstones | Upper montane Rhododendron (‘Elfin’) cloud forest | Rhododendron arboreum, mix from 3 trees with dieback (DB) (1/1) | ATT1 (1), CIN A1 (1) |
| F03 | N22 19.563 E103 46.679 | 2337 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | Quercus glauca, DB (2/2) | CIN A2 (2) |
| F04 | N22 19.670 E103 46.885 | 2242 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | Meliosma henryi (1/1) | CAS (1) |
| Betula alnoides & Elaeocarpus japonicus (1/1) | ATT1 (1), CIN A2 (1) c | ||||||
| Castanopsis acuminatissima, mix from 2 trees, DB (1/1) | ATT1 (1), CIN A2 (1) | ||||||
| C. acuminatissima with DB & Acer campbellii (1/1) | VIE (1) | ||||||
| F05 | N22 19.786 E103 46.899 | 2249 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | Neolitsea poilanei, DB (3/3) | ATT1 (3), CAS (1), CIN A2 (3) |
| C. acuminatissima mix from 3 trees, DB (1/1) | ATT (1), CIN A2 (1) | ||||||
| Illicium griffithii & C. acuminatissima, DB (1/1) | CAS (1) | ||||||
| F06 | N22 20.127 E103 46.782 | 2083 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | C. acuminatissima, DB (2/2) | CIN A2 (2), GRE (1), CAS (1) d |
| M. henryi & A. campbellii (1/1) | GRE (1) | ||||||
| M. henryi & Neolitsea merilliana (1/1) | CIN A2 (1), MUV1 (1) | ||||||
| F07 | N22 20.430 E103 46.574 | 2010 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | Illicium tsaii & Rhododendron sinofalconeri (1/1) | CAS (1) |
| C. acuminatissima, DB, necrotic root lesion (1/1) | CIN A2 (1) | ||||||
| F08 | N22 20.331 E103 46.664 | 2066 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | Casearia annamensis (1/1) | CIN A2 (1) |
| Acer oblongum, mix from 2 trees (0/1) | - e | ||||||
| F09 | N22 20.565 E103 46.565 | 2010 | Hoàng Liên NP | Triassic schists and sandstones | Montane evergreen cloud forest | C. acuminatissima (0/1) | - e |
| Q. glauca (0/2) | - | ||||||
| F10 | N22 20.632 E103 46.523 | 2022 | Hoàng Liên NP | Triassic schists and sandstones | Montane, evergreen cloud forest | Neolitsea polycarpa, mix from 3 trees, DB (1/1) | CIN A2 (1) |
| N. polycarpa, Symplocos pseudobarberina & Beilschmiedia roxburghiana (1/1) | CIN A2 (1) | ||||||
| F11 | N22 21.026 E103 46.315 | 1910 | Hoàng Liên NP | Triassic schists and sandstones | Montane, evergreen broadleaved forest | A. oblongum & Symplocos dryophila (2/2) | ATT (1), ATT2 (1), CIN A1 (2), CIN A2 (1), CIN A2ho (1) e,f |
| C. acuminatissima DB, Ilex leseeneri & Eurya annamensis (1/1) | CIN A2 (1), CAS (1) | ||||||
| R. arboreum (1/1) g | CHL (1), RAM A1 (1) | ||||||
| F12 | N22 20.909 E103 46.199 | 1895 | Hoàng Liên NP | Triassic schists and sandstones | Montane, evergreen broadleaved forest | Acer oliverianum, Eryobotrya cavaleriei & Symplocos quillaminii (1/1) | CIN A1 (1) |
| Q. glauca (1/1) | CIN A1 (1) | ||||||
| F13 | N22 21.090 E103 49.092 | 1717 | Xin Chài mountain | Triassic schists and sandstones | Montane Alnus forest on steep loamy slope | Alnus nepalensis (2/3) | CIT VII (1), MEA1 (1), ×TRO2 (1), VIE e |
| F14 | N22 22.168 E103 52.758 | 1367 | Sau Chua mountain | Triassic schists and sandstones | Montane Chamaecyparis-Quercus forest | Chamaecyparis hodginsii (7/9) | CAS (7) c,e |
| F15 | N21 3.699 E105 21.733 | 1108 | Ba Vì National Park (NP) | Triassic schists and sandstones and porphyrites | Suptropical humid evergreen forest | Castanopsis chinensis (2/2) | ATT3 (1), CAS (1), CIN A1 (1) e |
| C. chinensis & Beilschmiedia fordii (1/1) | CAS (1), HEV (1), PAR (1) d | ||||||
| Dysoxylum juglans, DB (1/1) | ATT3 (1) d | ||||||
| Eberhardtia tonkinensis, Antidesma sp. & Jasminum sp. (0/1) | - d | ||||||
| Eurya japonica & Nephelium lappaceum (1/1) | CAS (1) e | ||||||
| Lithocarpus bacgangensis (1/1) | CAS (1) d | ||||||
| Lithocarpus pseudosundaicus (0/1) | - | ||||||
| Machilus bonii (1/1) | CIN A1 (1) e | ||||||
| Magnolia annamensis (1/1) | CIN A2 (1) e | ||||||
| Q. glauca, mix from 3 trees (1/1) | ATT3 (1), CIN A1 (1), HEV (1), PAR (1) e | ||||||
| Vernicia montana & Antidesma sp. (1/1) | ATT3 (1), CIN A1 (1) d | ||||||
| F16 | N21 04.455 E105 21.810 | 807 | Ba Vì NP | Triassic schists and sandstones and porphyrites | Suptropical humid evergreen forest | Caryodaphnosis baviensis (0/2) | - e |
| Lithocarpus bacgangensis (1/1) | HEV (1) e | ||||||
| Meliosma arnottiana (1/1) | CIT IX (1) e | ||||||
| Phoebe petelotii, Machilus thunbergii & Claoxylon indicum (1/1) | CIN A2 (1) e | ||||||
| F17 | N21 04.587 E105 22.016 | 713 | Ba Vì NP | Triassic schists and sandstones and porphyrites | Suptropical humid evergreen forest | Alsodaphne velutina & Litsea brevipetiolata (1/1) | - e |
| Bischofia javanica & Litsea monocephala (0/1) | - e | ||||||
| C. chinensis (1/1) | ATT3 (1), CIN A2 (1) d | ||||||
| Castanopsis tonkinensis (1/1) | ATT3 (1) e | ||||||
| Q. glauca (0/1) | - e | ||||||
| F18 | N20 20.876 E105 35.793 | 392 | Cuc Phuong National Park (NP) | Triassic limestones | Tropical evergreen lowland rainforest | C. baviensis & Litsea robusta (0/1) | - e |
| Dracontomelum duppereanum, mix from 2 trees (0/1) | - e | ||||||
| Saraca dives, mix from 2 trees (1/1) | HEV (1) e | ||||||
| F19 | N20 20.779 E105 36.099 | 356 | Cuc Phuong NP | Triassic limestones | Tropical evergreen lowland rainforest | Allophylus cobbe, mix from 2 trees (0/1) | - e |
| D. duppereanum & S. dives (0/2) | - e,h | ||||||
| F20 | N20 20.366 E105 36.452 | 318 | Cuc Phuong NP | Triassic limestones | Tropical evergreen lowland rainforest | A. cobbe, Ficus sp., Merremia boisiana & Homalium sp. (1/1) | MEA2 (1) e,i |
| S. dives (0/2) | - e | ||||||
| F21 | N20 19.755 E105 36.979 | 267 | Cuc Phuong NP | Triassic limestones | Tropical evergreen lowland rainforest | Anogeissus acuminata (0/1) | - d or e |
| A. acuminata & Taxotrophis macrophylla (1/2) | CIT X (1) e,j | ||||||
| F22 | N20 18.963 E105 38.101 | 264 | Cuc Phuong NP | Triassic limestones | Tropical evergreen lowland rainforest | C. baviensis (0/1) | - e |
| C. baviensis & S. dives (0/1) | - e,j | ||||||
| S. dives, mix from 2 trees (0/1) | - e | ||||||
| F23 | N12 06.326 E107 09.396 | 417 | Bù Gia Mập National Park | Quaternary alluvial sediments | Tropical evergreen lowland rainforest | Dipterocarpus alatus, Ailanthus triphysa, Hopea odorata & Dalbergia oliveri (1/1) | HEV (1) e |
| F24 | N9 13.645 E104 57.330 | 4 | U Minh Hạ National Park | Quaternary peat | Tropical lowland peat forest | Melaleuca cajuputi (0/3) | - k |
| F25 | N8 40.621 E106 34.836 | 55 | Côn Đảo National Park, Côn Lôn island | Rhyolite and diorite | Tropical evergreen lowland rainforest | Chukrasia tabularis (0/1) | - e,k |
| A. triphysa, C. tabularis (1/1) | CIT XI e,l | ||||||
| Leucaena leucocephala, Canarium album & Hopea odorata (1/1) | BOT2 e,j | ||||||
| H. odorata, C. album, D. alatus (1/1) | HEV l, m |
| Site no. | GPS Coordinates | Altitude (m a.s.l) | River; Province | Location of Catchment and Vegetation | Sampling Method a | Phytophthora and Nothophytophthora spp. b,c |
|---|---|---|---|---|---|---|
| R01 | N22 20.127 E103 46.782 | 2083 | Forest stream 1; Lào Cai | Hoàng Liên NP; subalpine and montane Rhododendron scrub and forests, montane broadleaved forests | Baiting raft Fallen leaves/flowers | CAP, ×HET A1, ×HET A1ho, MUV1, RAM A1 d CIT VII, RAM A1, SYL2, VIE |
| R02 | N22 20.440 E103 46.576 | 2007 | Forest stream 2; Lào Cai | Hoàng Liên NP; subalpine and montane Rhododendron scrub and forests, montane broadleaved forests | Baiting raft Fallen leaves/flowers | RAM A1, SYL2 GAL1, GAL2, MUV1, RAM A1, SYL2, VIE e,f,g,h |
| R03 | N22 21.046 E103 46.273 | 1913 | Forest stream 3, tributary of forest stream 5; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | CHL, CIT VII, RAM A1, RAM A2, SYL2 e |
| R04 | N22 21.029 E103 46.317 | 1904 | Forest stream 4, tributary of forest stream 5; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | CHL, ×HET A1, RAM A1, RAM A2, SYL2, SYL3 i |
| R05 | N22 20.906 E103 46.197 | 1895 | Forest stream 5, Gold river, downstream of R03, R04, R06-R08; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | CHL, CIT VII, ×HET A1, RAM A1, SYL2 e |
| R06 | N22 20.911 E103 46.199 | 1896 | Forest stream 6, tributary of forest stream 5; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | CHL, ×HET A1 h,i |
| R07 | N22 20.902 E103 46.261 | 1912 | Forest stream 7, tributary of the Gold river; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | ×HET A1, RAM A1 |
| R08 | N22 20.904 E103 46.259 | 1911 | Forest stream 5, Gold river; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft | CIT VII, SYL2, SYL3 |
| R09 | N22 18.597 E103 52.426 | 1013 | Muong Hoa River; Lào Cai | Hoàng Liên NP; subalpine and montane Rhododendron scrub and forests, montane broadleaved forests, rice fields | Baiting raft | KEL, PSC, ×KUN |
| R10 | N22 19.372 E103 49.780 | 1193 | Forest stream 9, Cat Cat River; Lào Cai | Hoàng Liên NP; montane broadleaved forests | Baiting raft Fallen leaves/flowers | CAP, CIT VII, CIT VIII, SYL1, SYL3 e CHL, PSC, QUI, SYL 1, SYL3, RAM A1, VIE f,j,k |
| R11 | N22 22.230 E103 52.615 | 1308 | Forest stream 8; tributary of Ngòi Duôi River; Lào Cai | Sau Chua mountain; Chamaecyparis hodginsii forest F24; broadleaved mountain forests and Cunninghamia lanceolata plantations | Baiting raft Fallen leaves/flowers | BIT, CIT VII, CIT IX, MAC, QUI, SYL3 CIT IX, KEL, RAM A1 l |
| R12 | N22 16.787 E104 13.394 | 63 | Red River (Sông Hồng); Lào Cai | Large catchment in N-Vietnam and Yunnan; subalpine and montane Rhododendron scrub and forests, montane broadleaved forests, forest plantations, rice fields, horticulture | Baiting raft | ×KUN, ×PER4, ×VIR |
| R13 | N21 03.275 E105 24.050 | 59 | Stream 9; Hanoi | Ba Vi NP; subtropical evergreen forests, rice fields | Baiting raft | ×INS, ×GRE3, ×KUN, ×PER4, ×VIR e |
| R14 | N21 3.261 E105 24.012 | 60 | Stream 10, tributary of stream 9; Hanoi | Ba Vi NP; subtropical evergreen forests, rice fields | Baiting raft | ×KUN, ×PER 4 i |
| R15 | N21 06.177 E105 19.267 | 26 | Black River (Sông Đà); Hanoi | Large catchment in N-Vietnam and Yunnan; subalpine and montane Rhododendron scrub and forests, montane broadleaved forests, subtropical evergreen forests, forest plantations, rice fields, horticulture | Baiting raft | DRE A1, ×VIR l,m |
| R16 | N21 01.576 E105 27.218 | 26 | Stream 11; Hanoi | Forest plantations, rice fields, horticulture | Baiting raft | ×PER4, ×VIR n |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, M.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum. Forests 2020, 11, 93. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010093
Jung T, Scanu B, Brasier CM, Webber J, Milenković I, Corcobado T, Tomšovský M, Pánek M, Bakonyi J, Maia C, et al. A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum. Forests. 2020; 11(1):93. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010093
Chicago/Turabian StyleJung, Thomas, Bruno Scanu, Clive M. Brasier, Joan Webber, Ivan Milenković, Tamara Corcobado, Michal Tomšovský, Matěj Pánek, József Bakonyi, Cristiana Maia, and et al. 2020. "A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum" Forests 11, no. 1: 93. https://0-doi-org.brum.beds.ac.uk/10.3390/f11010093