Root Traits and Biomechanical Properties of Three Tropical Pioneer Tree Species for Forest Restoration in Landslide Areas

Abstract
:1. Introduction
2. Materials and Methods
2.1. Seed Collection
2.2. Seedling Preparation
2.3. Growth Performance and Root System Architecture Investigation
2.4. Vertical Pullout Test
2.5. Root Tensile and Young’s Modulus Tests
2.6. Data Analysis and Statistics
3. Results
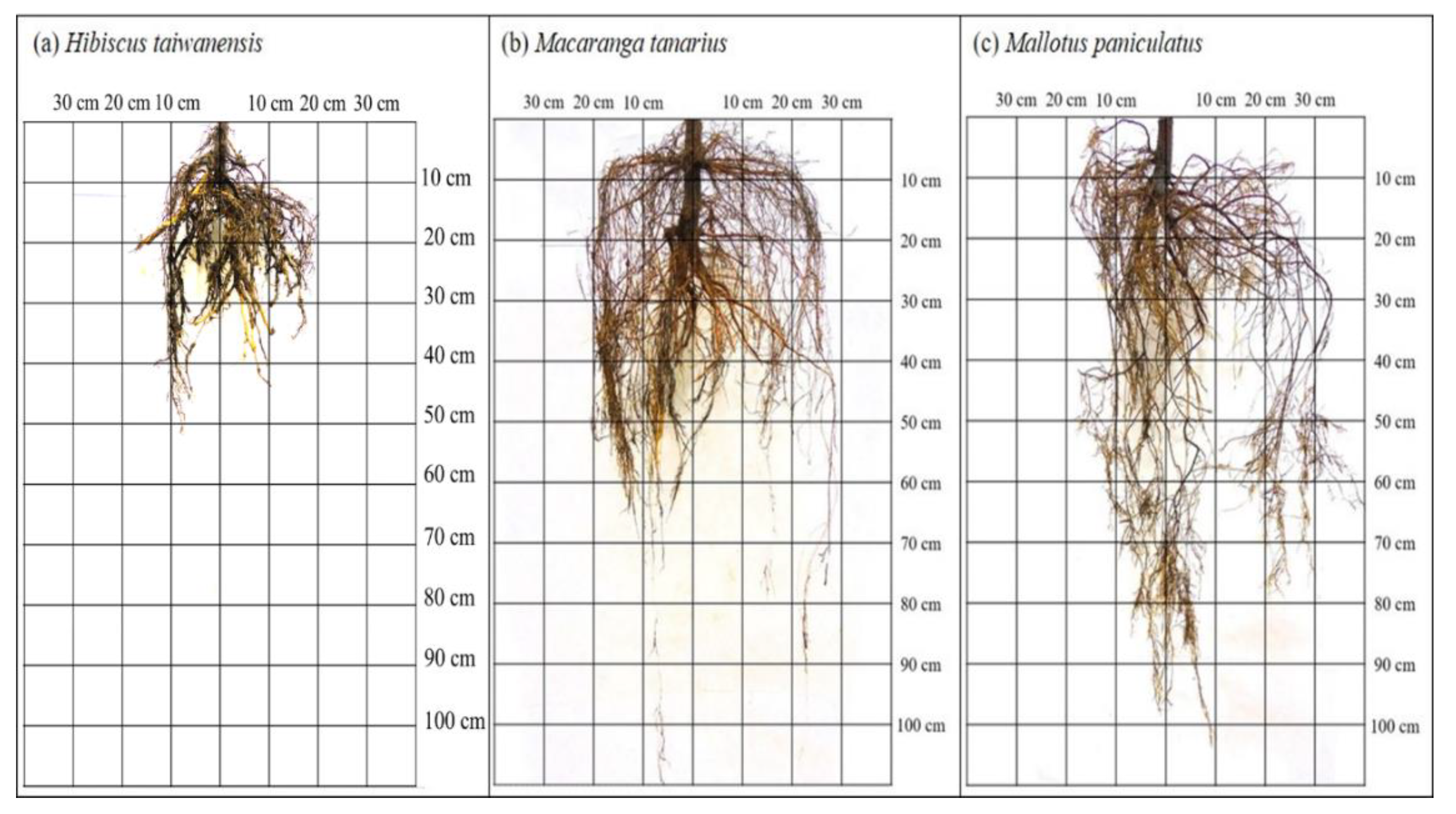
3.1. Root System Architecture
3.2. Seedling Growth Performance
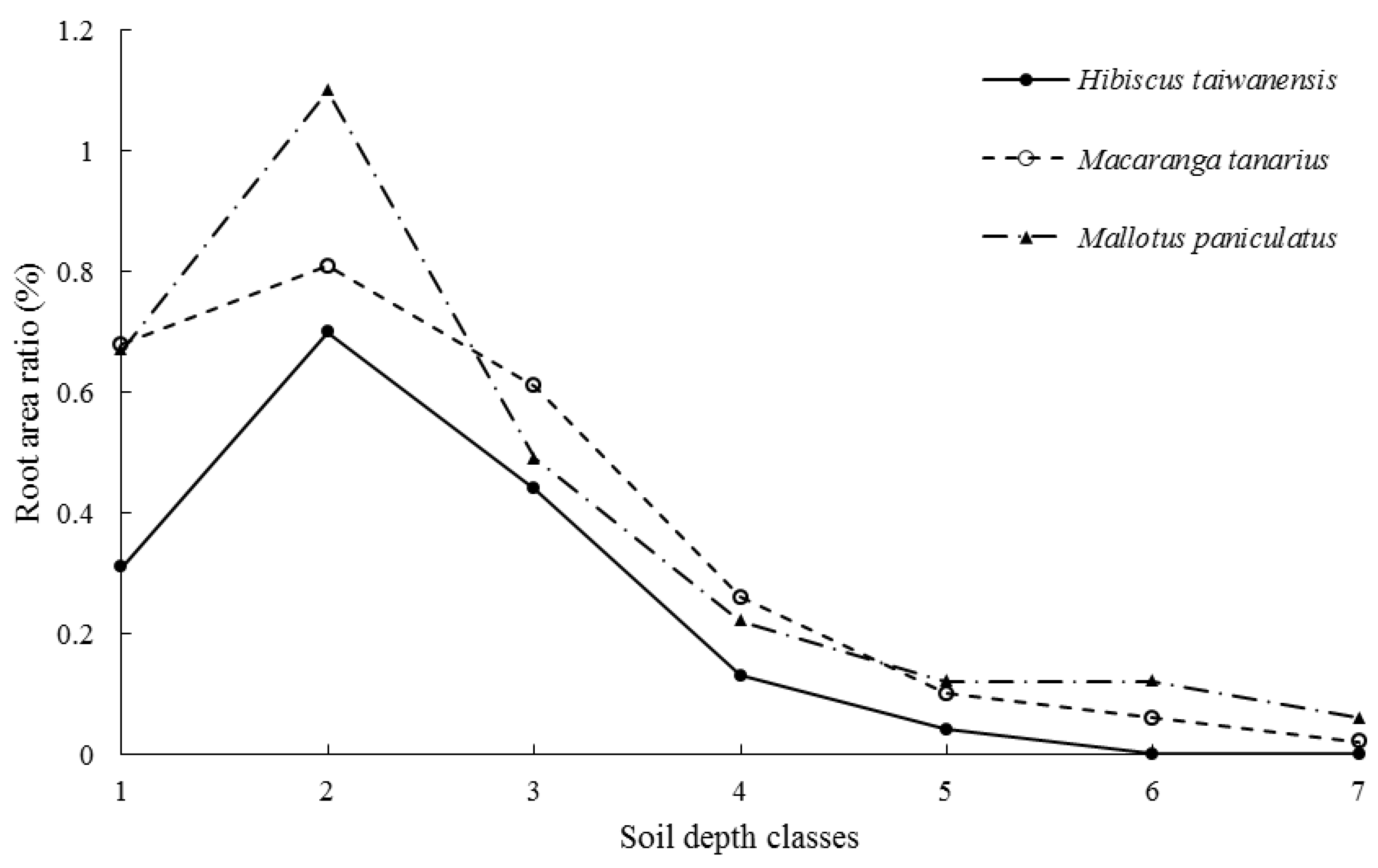
3.3. Root Traits
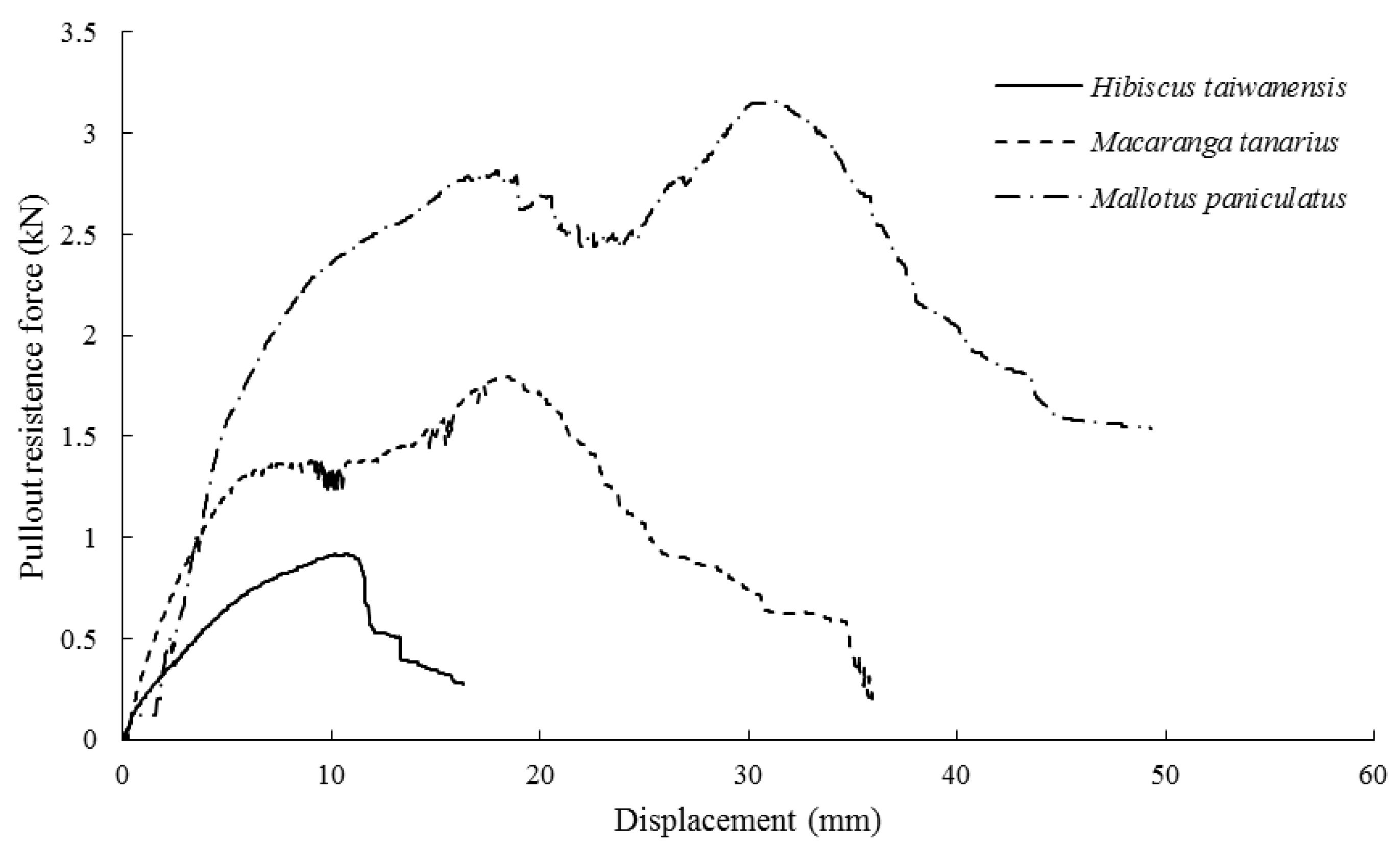
3.4. Root Anchorage Capability
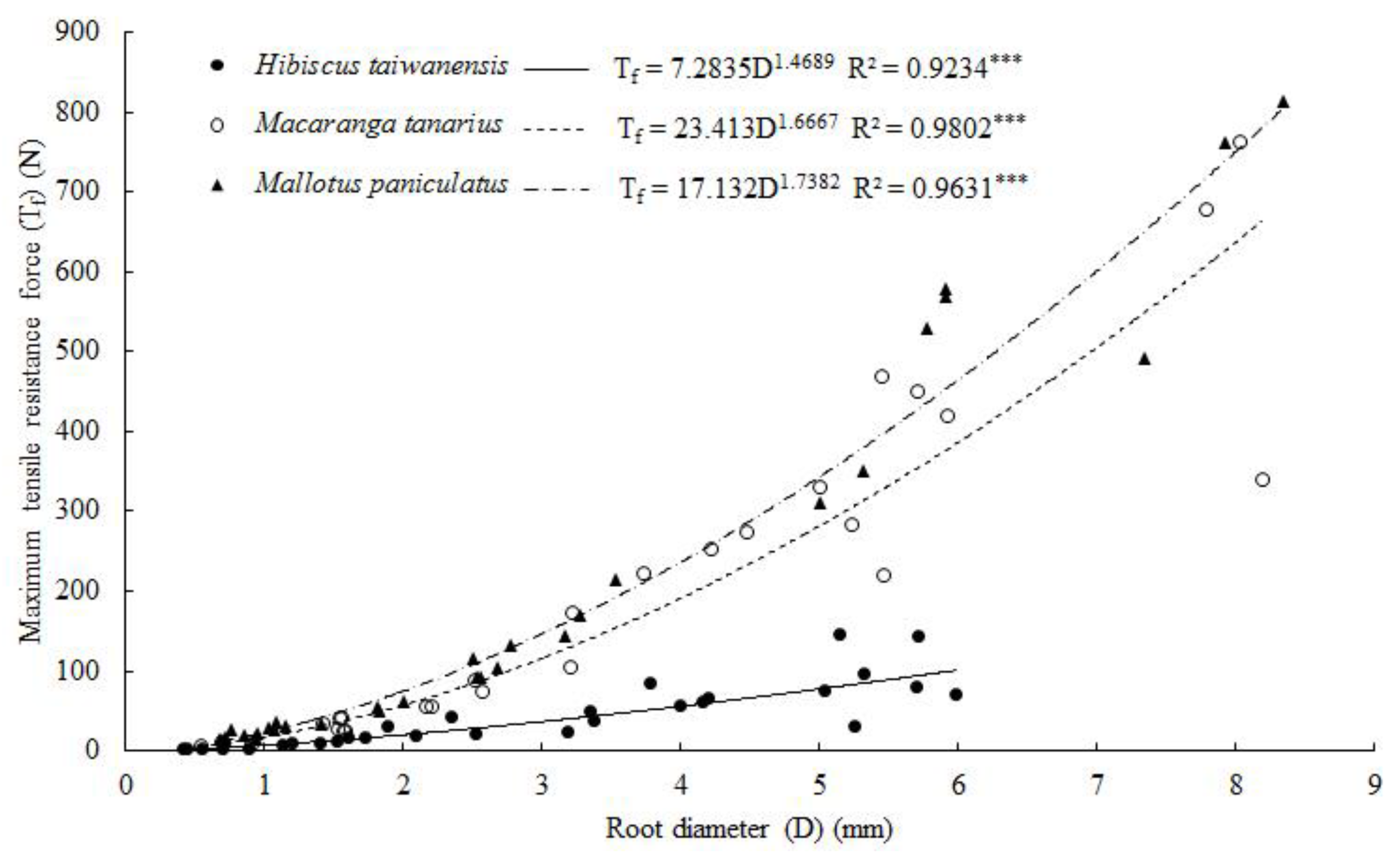
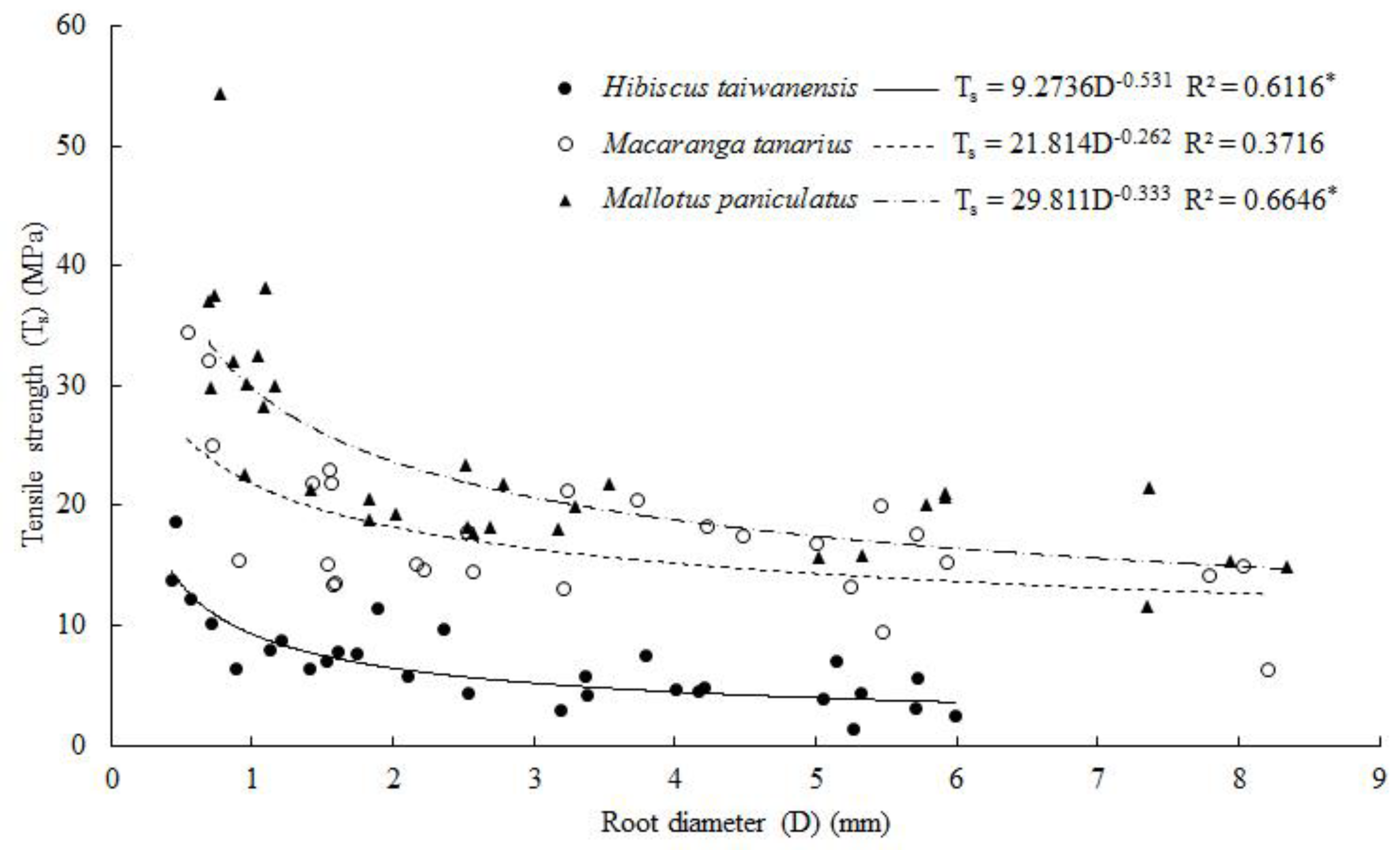
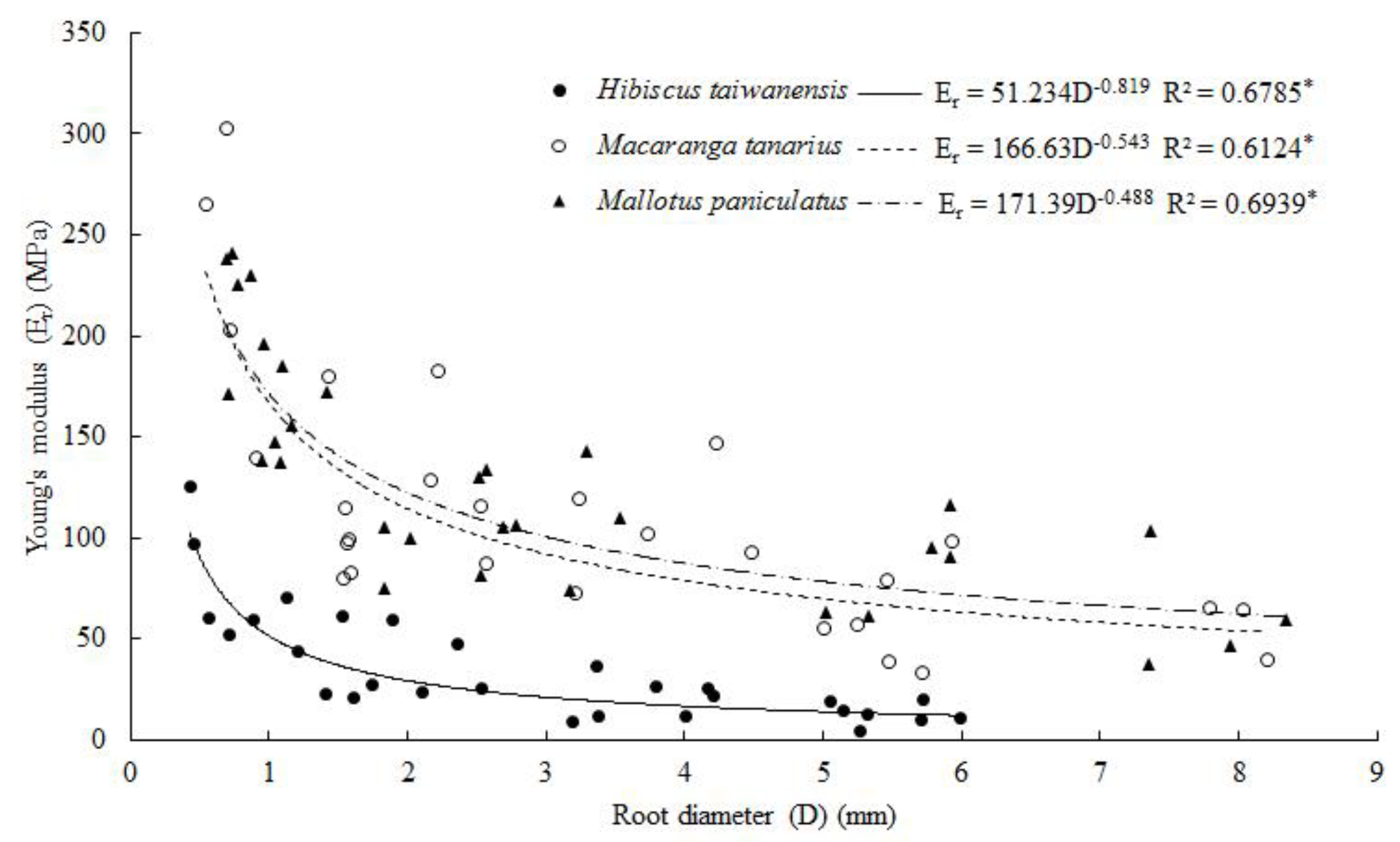
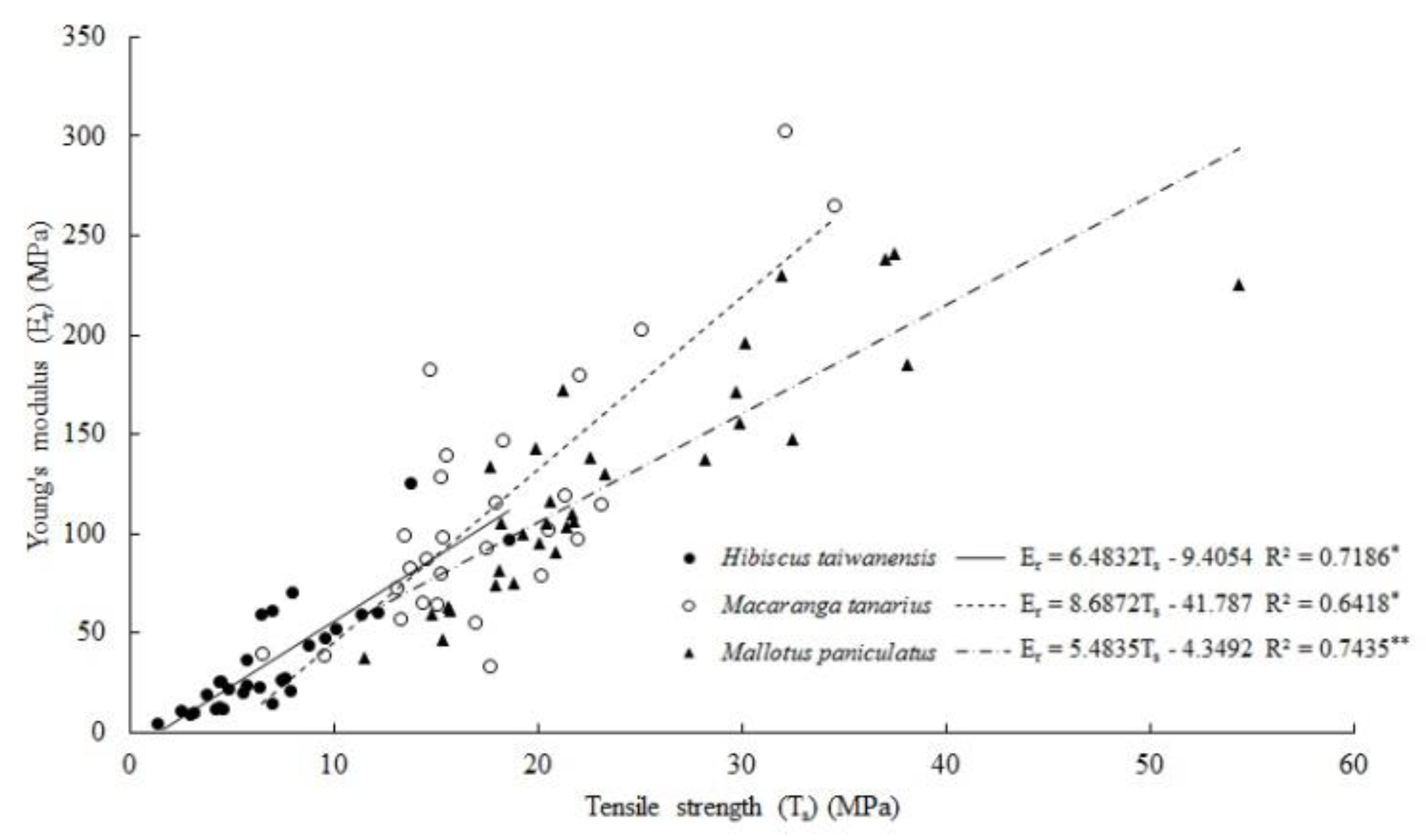
3.5. Root Tensile Strength and Young’s Modulus
4. Discussion
4.1. Root System Architecture
4.2. Seedling Growth Performance
4.3. Root Traits
4.4. Relationship between Root Morphological Traits and Anchorage Capability
4.5. Root Tensile Resistance, Tensile Strength and Young’s Modulus
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflict of Interests
References
- Chang, J.C. Natural hazards in Taiwan. GeoJournal 1996, 38, 251–257. [Google Scholar] [CrossRef]
- Tsou, C.Y.; Feng, Z.Y.; Chigira, M. Catastrophic landslide induced by Typhoon Morakot, Shiaolin, Taiwan. Geomorphol. 2011, 127, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T.; Tsai, S.M. The nitrogen-fixing Frankia significantly increases growth, uprooting resistance and root tensile strength of Alnus formosana. Afr. J. Biotechnol. 2018, 17, 213–225. [Google Scholar]
- Polster, D.F. Successional reclamation in Western Canada: New light on an old subject. In Proceedings of the America Society of Mining and Reclamation, Calgary, AB, Canada, 27–31 August 1989; pp. 333–338. [Google Scholar] [CrossRef]
- Ngugi, M.R.; Neldner, V.J.; Doley, D.; Kusy, B.; Moore, D.; Richter, C. Soil moisture dynamics and restoration of self-sustaining native vegetation ecosystem on an open-cut coal mine. Restor. Ecol. 2015, 23, 615–624. [Google Scholar] [CrossRef]
- Huang, H.B. Soil and water conservation design. In Soil and Water Conservation Engineering; Huang, H.B., Ed.; Wu-Nan Culture Enterprise: Taipei, Taiwan, 2014; pp. 71–91. [Google Scholar]
- Lin, S.H. Vegetation and slope stability. In Vegetation Engineering of Slopeland; Lin, S.H., Ed.; Wu-Nan Culture Enterprise: Taipei, Taiwan, 2016; pp. 105–120. [Google Scholar]
- Chou, F.S.; Lin, W.C.; Chen, Y.H.; Liao, C.K. Monitoring the vegetation dynamics of early succession following a landslide on Shanping forest road. Taiwan J. For. Sci. 2015, 30, 217–228. [Google Scholar]
- Chang, C.E. Malvaceae. In Flora of Taiwan, 2nd ed.; Editorial Committee of Flora of Taiwan, Ed.; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 1993; pp. 742–744. [Google Scholar]
- Hsieh, C.F. Euphorbiaceae. In Flora of Taiwan; Editorial Committee of Flora of Taiwan: Taipei, Taiwan, 1993; pp. 484–487. [Google Scholar]
- Lin, S.H.; Cheng, Y.J.; Chen, J.N. Vegetation types and habitat characteristics of landslide areas. For. Res. Newslett. 2011, 18, 40–45. [Google Scholar]
- Galias, D.C.; Cuevas, V.C. The regenerating forest of Magbukún Aceta in Morong, Bataan, Philippines: A biological hotspot for protection and conservation. Philipp. J. Syst. Biol. 2018, 12, 77–102. [Google Scholar]
- Chen, T.L.; Wey, T.H.; Lee, J.; Lu, F.Y. Comparison of the natural and artificial regeneration in landslide areas caused by an earthquake in the Chaoling region, Central Taiwan. J. Chin. Soil Water Conserv. 2009, 40, 49–65. [Google Scholar]
- Chen, Y.C.; Wu, C.F.; Lin, S.H. Mechanisms of forest restoration in landslide treatment areas. Sustainability 2014, 6, 6766–6780. [Google Scholar] [CrossRef] [Green Version]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots withstanding their environment: Exploiting root system architecture responses to abiotic stress to improve crop tolerance. Front. Plant Sci. 2016, 7. [Google Scholar] [CrossRef] [Green Version]
- Yen, C.P. Tree root patterns and erosion control. In International Workshop on Soil Erosion and its Counter-Measures; Jantawat, S., Ed.; Soil Conservation Society of Thailand: Bangkok, Thailand, 1987; pp. 92–111. [Google Scholar]
- Saifuddin, M.; Osman, N.; Rahman, M.M.; Boyce, A.N. Soil reinforcement capability of two legume species from plant morphological traits and mechanical properties. Curr. Sci. 2015, 108, 1340–1347. [Google Scholar]
- Stokes, A.; Atger, C.; Bengough, A.G.; Fourcaud, T.; Sidle, R.C. Desirable plant root traits for protecting natural and engineered slopes against landslides. Plant Soil 2009, 324, 1–30. [Google Scholar] [CrossRef]
- Lin, D.G.; Wang, S.H.; Chen, W.H.; Chou, J.C. Comparisons of pull-out resistance and shear strength increment of soil/root system for plants commonly used in soil and water conservation: India-charcoal trema, cotton rose, and subcostate crape myrtle. J. Chin. Soil Water Conserv. 2016, 47, 84–94. [Google Scholar]
- Avani, N.; Lateh, H.; Bibalani, G.H. Study of root distribution of Macaranga tanarius (L.) Müll. Arg. (Parasol leaf tree) on East-West highway slope, Malaysia. J. BioSci. Biotechnol. 2013, 2, 195–200. [Google Scholar]
- Lateh, H.; Avani, N.; Bibalani, G. Tensile strength and root distribution of Acacia mangium and Macaranga tanarius at spatial variation (Case study: East-West highway, Malaysia). Int. J. Biosci. 2015, 6, 18–28. [Google Scholar]
- Böhm, W. Excavation methods. In Methods of Studying Root Systems; Billings, W.D., Colley, F., Lange, O.L., Olson, J.S., Eds.; Springer: Heidelberg, Germany, 1979; pp. 5–18. [Google Scholar]
- Bischetti, G.B.; Chiaradia, E.A.; Simonato, T.; Speziali, B.; Vitali, B.; Vullo, P.; Zocco, A. Root strength and root area ratio of forest species in Lombardy (Northern Italy). Plant Soil 2005, 278, 11–22. [Google Scholar] [CrossRef]
- Bouma, T.J.; Nielsen, K.L.; Koutstaal, K. Sample preparation and scanning protocol for computerized analysis of root length and diameter. Plant Soil 2000, 218, 185–196. [Google Scholar] [CrossRef]
- Pang, W.; Crow, W.T.; Luc, J.E.; McSorley, R.; Giblin-Davis, R.M.; Kruse, J.K. Comparison of water displacement and WinRHIZO software for plant root parameter assessment. Plant Dis. 2011, 95, 1308–1310. [Google Scholar] [CrossRef] [Green Version]
- Rose, L. Pitfalls in root trait calculations: How ignoring diameter heterogeneity can lead to overestimation of functional traits. Front. Plant Sci. 2017, 8, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Burylo, M.; Dutoit, T.; Rey, F. Species traits as practical tools for ecological restoration of Marly eroded lands. Restor. Ecol. 2014, 22, 633–640. [Google Scholar] [CrossRef]
- Gould, I.J.; Quinton, J.N.; Weigelt, A.; De Deyn, G.B.; Bardgett, R.D. Plant diversity and root traits benefit physical properties key to soil function in grasslands. Ecol. Lett. 2016, 19, 1140–1149. [Google Scholar] [CrossRef]
- Hales, T.C.; Miniat, C.F. Soil moisture causes dynamic adjustments to root reinforcement that reduce slope stability. Earth Surf. Process. Landf. 2017, 42, 803–813. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.T.; Yen, L.Z.; Lee, M.J. Wind affects the growth, root anchorage and tensile strength of Australian pine (Casuarina equisetifolia) seedlings. J. Forest Res. 2019, 24, 219–229. [Google Scholar] [CrossRef]
- Cofie, P.; Koolen, A.J. Test speed and other factors affecting the measurements of tree root properties used in soil reinforcement models. Soil Tillage Res. 2001, 63, 51–56. [Google Scholar] [CrossRef]
- Genet, M.; Kokutse, N.; Stokes, A.; Fourcaud, T.; Caim, X.; Ji, J.; Mickovski, S.B. Root reinforcement in plantations of Cryptomeria japonica D. Don: Effect of tree age and stand structure on slope stability. For. Ecol. Manag. 2008, 256, 1517–1526. [Google Scholar]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Biomechanics of nodal, seminal and lateral roots of barley: Effects of diameter, waterlogging and mechanical impedance. Plant Soil 2013, 370, 407–418. [Google Scholar]
- Loades, K.W.; Bengough, A.G.; Bransby, M.F.; Hallett, P.D. Effect of root age on biomechanics of seminal and nodal roots of barley (Hordeum vulgare L.) in contrasting soil environments. Plant Soil 2015, 395, 253–261. [Google Scholar] [CrossRef] [Green Version]
- De Baets, S.; Poeson, J.; Reubens, B.; Wemans, K.; Baerdemaeker, J.D.; Muys, B. Root tensile strength and root distribution of typical Mediterranean plant species and their contribution to soil shear strength. Plant Soil 2008, 305, 207–226. [Google Scholar] [CrossRef]
- Burylo, M.; Hudek, C.; Rey, F. Soil reinforcement by the roots of six dominant species on eroded mountainous marly slopes (Southern Alps, France). Catena 2011, 84, 70–78. [Google Scholar] [CrossRef]
- Chimungu, J.G.; Loades, K.W.; Lynch, J.P. Root anatomical phenes predict root penetration ability and biomechanical properties in maize (Zea mays). J. Exp. Bot. 2015, 66, 3151–3162. [Google Scholar] [CrossRef] [Green Version]
- Gray, D.H.; Sotir, R.B. Role of vegetation in the stability of slopes. In Biotechnical and Soil Bioengineering Slope Stabilization; Gray, D.H., Sotir, R.B., Eds.; John Wiley & Sons: New York, NY, USA, 1996; pp. 54–105. [Google Scholar]
- Boldrin, D.; Leung, A.K.; Bengough, A.G. Effects of root dehydration on biomechanical properties of woody roots of Ulex europaeus. Plant Soil 2018, 431, 347–369. [Google Scholar] [CrossRef] [Green Version]
- Danjon, F.; Reubens, B. Assessing and analyzing 3D architecture of woody root systems, a review of methods and applications in tree and soil stability, resource acquisition and allocation. Plant Soil 2008, 303, 1–34. [Google Scholar] [CrossRef]
- Ghestem, M.; Veylon, G.; Bernard, A.; Stokes, A. Influence of plant roots system morphology and architectural traits on soil shear resistance. Plant Soil 2014, 377, 43–61. [Google Scholar] [CrossRef]
- Fan, C.C.; Chen, Y.W. The effect of root architecture on the shearing resistance of root-permeated soils. Ecol. Eng. 2010, 36, 813–826. [Google Scholar] [CrossRef]
- Saifuddin, M.; Normaniza, O. Rooting characteristics of some tropical plants for slope protection. J. Trop. Forest Sci. 2016, 28, 469–478. [Google Scholar]
- Forde, B.G. Nitrogen signalling pathways shaping root systemarchitecture: An update. Curr. Opin. Plant Biol. 2014, 21, 30–36. [Google Scholar] [CrossRef]
- Kiba, T.; Krapp, A. Plant nitrogen acquisition under low availability: Regulation of uptake and root architecture. Plant Cell Physiol. 2016, 57, 707–714. [Google Scholar] [CrossRef] [Green Version]
- Jobbágy, E.G.; Jackson, R.B. The distribution of soil nutrients with depth: Global patterns and the imprint of plants. Biogeochemistry 2001, 53, 51–77. [Google Scholar] [CrossRef]
- February, E.C.; Higgins, S.I. The distribution of tree and grass roots in savannas in relation to soil nitrogen and water. S. Afr. J. Bot. 2010, 76, 517–523. [Google Scholar] [CrossRef] [Green Version]
- Wasson, A.; Richards, R.; Chatrath, R.; Misra, S.; Prasad, S.S.; Rebetzke, G.; Kirkegaard, J.; Christopher, J.; Watt, M. Traits and selection strategies to improve root systems and water uptake in water-limited wheat crops. J. Exp. Bot. 2012, 63, 3485–3498. [Google Scholar] [CrossRef] [Green Version]
- Pang, C.C.; Ma, X.K.K.; Lo, J.P.L.; Hung, T.T.H.; Hau, B.C.H. Vegetation succession on landslides in Hong Kong: Plant regeneration, survivorship and constraints to restoration. Glob. Ecol. Conserv. 2018, 15, e00428. [Google Scholar] [CrossRef]
- Davis, A.S.; Jacobs, D.F. Quantifying root system quality of nursery seedlings and relationship to outplanting performance. New For. 2005, 30, 295–311. [Google Scholar] [CrossRef]
- Morrissey, R.C.; Jacobs, D.F.; Davis, A.S.; Rathfon, R.A. Survival and competitiveness of Quercus rubra regeneration associated with planting stocktype and harvest opening intensity. New For. 2010, 40, 273–287. [Google Scholar] [CrossRef]
- Grossnickle, S.C. Why seedlings survive: Influence of plant attributes. New For. 2010, 43, 711–738. [Google Scholar] [CrossRef]
- Jackson, R.B.; Mooney, H.A.; Schulze, E.-D. A global budget for fine root biomass, surface area, and nutrient contents. Proc. Natl. Acad. Sci. USA 1997, 94, 7362–7366. [Google Scholar] [CrossRef] [Green Version]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root traits contributing to plant productivity under drought. Front. Plant Sci. 2013, 4, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Kramer-Walter, K.R.; Bellingham, P.J.; Millar, T.R.; Smissen, R.D.; Richardson, S.J.; Laughlin, D.C. Root traits are multidimensional: Specific root length is independent from root tissue density and the plant economic spectrum. J. Ecol. 2016, 104, 1299–1310. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Roumet, C.; Buisson, E.; Dutoit, T. Linking plant morphological traits to uprooting resistance in eroded marly lands (Southern Alps, France). Plant Soil 2009, 324, 31–42. [Google Scholar] [CrossRef]
- De Baets, S.; Poesen, J.; Knapen, A.; Barberá, G.G.; Navarro, J.A. Root characteristics of representative Mediterranean plant species and their erosion-reducing potential during concentrated runoff. Plant Soil 2007, 294, 169–183. [Google Scholar] [CrossRef]
- Burylo, M.; Rey, F.; Mathys, N.; Dutoit, T. Plant root traits affecting the resistance of soils to concentrated flow erosion. Earth Surf. Proc. Land. 2012, 37, 1463–1470. [Google Scholar] [CrossRef]
- Katuwal, S.; Vermang, J.; Cornelis, W.M.; Gabriels, D.; Moldrup, P.; De Jonge, L.W. Effect of root density on erosion and erodibility of a loamy soil under simulated rain. Soil Sci. 2013, 178, 29–36. [Google Scholar] [CrossRef]
- Jones, B.; Ljung, K. Subterranean space exploration: The development of root system architecture. Curr. Opin. Plant Biol. 2012, 15, 97–102. [Google Scholar] [CrossRef]
- Ola, A.; Dodd, I.C.; Quinton, J.N. Can we manipulate root system architecture to control erosion? Soil 2015, 1, 603–612. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Jia, Z.; Gu, L.; Gao, J. Vertical and lateral uprooting resistance of Salix matsudana Koidz in a riparian area. For. Chron. 2013, 89, 162–168. [Google Scholar] [CrossRef]
- Lee, J.T.; Tsai, S.M.; Lee, M.J. Uprooting resistance of two tropical tree species for sand dune stabilization. Afr. J. Agric. Res. 2017, 12, 3214–3220. [Google Scholar]
- Edmaier, K.; Crouzy, B.; Ennos, R.; Burlando, P.; Perona, P. Influence of root characteristics and soil variables on the uprooting mechanics of Avena sativa and Medicago sativa seedlings. Earth Surf. Process. Landf. 2014, 39, 1354–1364. [Google Scholar] [CrossRef]
- Yang, M.; Défossez, P.; Danjon, F.; Fourcaud, T. Analyzing key factors of roots and soil contributing to tree anchorage of Pinus species. Trees 2017, 32, 703–712. [Google Scholar]
- Leung, F.T.Y.; Yan, R.W.M.; Hau, B.C.H.; Tham, L.G. Mechanical pull-out capacity and root reinforcement of four native tree and shrub species on ecological rehabilitation of roadside slopes in Hong Kong. J. Trop. For. Sci. 2018, 30, 25–38. [Google Scholar]
- Lin, D.G.; Liu, W.T.; Lin, S.H. Estimating the effect of shear strength increment due to root on the stability of Makino bamboo forest slopeland. J. GeoEng. 2011, 6, 73–88. [Google Scholar]
- Tardío, G.; Mickovski, S.B. Method for synchronisation of soil and root behaviour for assessment of stability of vegetated slopes. Ecol. Eng. 2015, 82, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Genet, M.; Stokes, A.; Salin, F.; Mickovski, S.B. The influence of cellulose content on tensile strength in tree roots. Plant Soil 2005, 278, 1–9. [Google Scholar] [CrossRef]
- Tosi, M. Root tensile strength relationships and their slope stability implications of three shrub species in the Northern Apennines (Italy). Geomorphology 2007, 87, 268–283. [Google Scholar] [CrossRef]
- Sanchez-Castillo, L.; Kubota, T.; Cantu-Silva, I.; Yañez-Diaz, M.; Pequeño-Ledezma, M. Comparisons of the root mechanical properties of three native Mexican tree species for soil bioengineering practices. Bot. Sci. 2017, 95, 259–269. [Google Scholar] [CrossRef] [Green Version]
- Capilleri, P.P.; Motta, E.; Raciti, E. Experimental study on native plant root tensile strength for slope stabilization. Procedia Eng. 2016, 158, 116–121. [Google Scholar] [CrossRef]
- Saifuddin, M.; Normaniza, O. Evaluation of hydro-mechanical properties and root architecture of plants for soil reinforcement. Curr. Sci. 2014, 107, 845–852. [Google Scholar]
- Yang, Y.; Chen, L.; Li, N.; Zhang, Q. Effect of root moisture content and diameter on root tensile properties. PLoS ONE 2016, 11, e0151791. [Google Scholar]
- Zhang, C.B.; Chen, L.H.; Jiang, J. Why fine tree roots are stronger than thicker roots: The role of cellulose and lignin in relation to slope stability. Geomorphology 2014, 206, 196–202. [Google Scholar] [CrossRef]
- Hathaway, R.L.; Penny, D. Root strength in some Populus and Salix clones. N. Z. J. Bot. 1975, 13, 333–344. [Google Scholar] [CrossRef] [Green Version]
- Liang, T.; Bengough, A.G.; Knappett, J.A.; Wood, D.M.; Loades, K.W.; Hallett, P.D.; Boldrin, D.; Leung, A.K.; Meijer, G.J. Scaling of the reinforcement of soil slopes by living plants in a geotechnical centrifuge. Ecol. Eng. 2017, 109, 207–227. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Properties | Soil |
|---|---|
| pH (water) | 7.98 |
| Conductivity (dS m−1) | 0.13 |
| Organic carbon (g kg−1) | 0.07 |
| Total nitrogen (%) | 0.03 |
| Phosphorus (mg kg−1) | 21 |
| Potassium (mg kg−1) | 152 |
| Calcium (mg kg−1) | 3491 |
| Magnesium (mg kg−1) | 320 |
| Zn (ppm) | 4.3 |
| Mn (ppm) | 163 |
| Fe (ppm) | 343 |
| Cu (ppm) | 1.8 |
| Cd (ppm) | 0.01 |
| Cr (ppm) | 0.74 |
| Ni (ppm) | 1.5 |
| Pb (ppm) | 4.4 |
| Species | Root Area Ratio (%) | ||||||
|---|---|---|---|---|---|---|---|
| 0–10 cm | 10–20 cm | 20–30 cm | 30–40 cm | 40–50 cm | 50–60 cm | 60–70 cm | |
| H. t. | 0.31 ± 0.04b | 0.70 ± 0.11a | 0.44 ± 0.14a | 0.13 ± 0.071a | 0.04±0.037a | - | - |
| M. t. | 0.68 ± 0.05a | 0.81 ± 0.13a | 0.61 ± 0.09a | 0.26±0.055a | 0.1 ± 0.028a | 0.06 ± 0.017b | 0.02 ± 0.006a |
| M. p. | 0.67 ± 0.08a | 1.10 ± 0.15a | 0.49 ± 0.06a | 0.22±0.028a | 0.12 ± 0.02a | 0.12 ± 0.28a | 0.06 ± 0.03a |
| Growth Parameters | H. taiwanensis | M. tanarius | M. paniculatus | ANOVA |
|---|---|---|---|---|
| H (cm) | 142.6 ±10.2b | 138.1 ± 15.3b | 161.1 ± 13.9a | 4.388* |
| RCD (mm) | 16.7 ± 0.9b | 25.0 ± 1.9a | 24.1 ± 2.3a | 44.892*** |
| TL (cm) | 44.3 ± 10.1b | 83.1 ± 18.1a | 78.4 ± 15.2a | 9.362** |
| RT | 5391 ± 2426b | 10780 ± 1944a | 7905 ± 1811a | 18.614*** |
| TRL (cm) | 3950.7 ± 166.2b | 7474.2 ± 137.9a | 6447.4 ± 133.0a | 18.796*** |
| RB (g) | 57.6 ± 16.4b | 96.3 ± 9.4a | 104.8 ± 11.4a | 44.846*** |
| SB (g) | 59.5 ± 11.8b | 58.3 ± 6.9b | 146.0 ± 32.4a | 17.621*** |
| Root Traits | H. taiwanensis | M. tanarius | M. paniculatus | ANOVA |
|---|---|---|---|---|
| RD (kg m−3) | 1.07 ± 0.30b | 1.78 ± 0.17a | 1.94 ± 0.21a | 15.39** |
| RLD (km m−3) | 0.73 ± 0.31b | 1.38 ± 0.26a | 1.19 ± 0.21a | 6.54* |
| RM (g m−3) | 1067.4 ± 302.9b | 1782.4 ± 174.6a | 1939.8 ± 210.8a | 15.56** |
| RSA (cm2) | 2995.3 ± 959.4b | 5391.4 ± 786.8a | 5277.4 ± 325.4a | 5.23* |
| RTD (g cm−3) | 0.15 ± 0.03b | 0.29 ± 0.04a | 0.29 ± 0.02a | 30.56*** |
| RV (cm3) | 377.5 ± 76.8a | 332.5 ± 58.5a | 360.0 ± 46.9a | 0.54 ns |
| SRL (m g−1) | 0.69 ± 0.26a | 0.77 ± 0.07a | 0.62 ± 0.13a | 0.76 ns |
| Biomechanical Properties | H. taiwanensis | M. tanarius | M. paniculatus | ANOVA |
|---|---|---|---|---|
| Maximum Pullout Resistance Force (kN) | 1.02±0.33b | 1.53±0.64b | 2.96±0.58a | 24.936*** |
| Morphological traits | Species | Regression equation | R2 | p |
|---|---|---|---|---|
| RCD (mm) | H. taiwanensis M. tanarius M. paniculatus | Pr = 0.3215RCD − 4.3803 Pr = 0.3135RCD − 6.2987 Pr = 0.2212RCD − 2.3782 | 0.7427* 0.8413** 0.7559* | 0.031 0.004 0.011 |
| TL (cm) | H. taiwanensis M. tanarius M. paniculatus | Pr = 0.007TL + 0.691 Pr = -0.001TL + 1.641 Pr = -0.004TL + 3.216 | 0.023 0.027 0.026 | 0.746 0.954 0.73 |
| RT | H. taiwanensis M. tanarius M. paniculatus | Pr = 0.00004064RT + 0.784 Pr = 0.0001RT − 0.32 Pr = -0.00003811RT + 3.261 | 0.054 0.17 0.01 | 0.617 0.358 0.835 |
| RB (g) | H. taiwanensis M. tanarius M. paniculatus | Pr = 0.0236RB−0.4553 Pr = 0.0715RB−5.6277 Pr = 0.0646RB−3.8727 | 0.8276** 0.8270** 0.8959** | 0.004 0.005 0.001 |
| SB (g) | H. taiwanensis M. tanarius M. paniculatus | Pr = 0.0301SB−0.8055 Pr = 0.0121SB−0.6939 Pr =0.0179SB+0.2498 | 0.7122* 0.6969* 0.7251* | 0.017 0.019 0.015 |
| Species | Regression Equation | R2 |
|---|---|---|
| H. taiwanensi | Pr = 0.341RCD + 0.014TL − 5.33 | 0.833* |
| M. tanarius | Pr = 0.320RCD + 0.006TL − 6.94 | 0.855* |
| M. paniculatus | Pr = 0.246RCD − 0.009TL − 2.35 | 0.907** |
| Parameters | H. taiwanensis | M. tanarius | M. paniculatus | ANOVA |
|---|---|---|---|---|
| Root diameter (mm) | 2.93 ± 1.83a | 3.47 ± 2.33a | 3.11 ± 2.41a | 0.448ns |
| Tensile resistance force (N) | 42.59 ± 7.46b | 197.04 ± 39.45a | 213.41 ± 46.98a | 6.612*** |
| Tensile strength (MPa) | 6.89 ± 3.71c | 17.77 ± 5.98b | 23.89 ± 8.93a | 50.764*** |
| Root Diameter (mm) | Tensile Strength (MPa) | ||
|---|---|---|---|
| H. taiwanensis | M. tanarius | M. paniculatus | |
| 0-1 | 12.21 ± 4.5b | 26.78 ± 8.53a | 34.78 ± 10.01a |
| 1-2 | 8.13 ± 1.63c | 18.21 ± 4.57b | 27.06 ± 7.14a |
| 2-5 | 5.41 ± 1.89b | 16.98 ± 2.81a | 19.82 ± 2.04a |
| 5-10 | 3.97 ± 1.9b | 14.27 ± 4.18a | 17.37 ± 3.49a |
| Biomechanical Properties | Hibiscus taiwanensis | Macaranga tanarius | Mallotus paniculatus | ANOVA |
|---|---|---|---|---|
| Young’s modulus (MPa) | 25.27 ± 8.35b | 112.6 ± 64.9a | 127.15 ± 56.8a | 26.346*** |
| Root Diameter (mm) | Young’s Modulus (MPa) | ||
|---|---|---|---|
| H. taiwanensis | M. tanarius | M. paniculatus | |
| 0–1 | 78.44 ± 31.42b | 227.91 ± 71.72a | 205.55 ± 39.06a |
| 1–2 | 43.47 ± 20.25b | 109.31 ± 37.08a | 139.60 ± 38.11a |
| 2–5 | 23.60 ± 11.83b | 116.78 ± 33.59a | 109.06 ± 23.02a |
| 5–10 | 12.91 ± 5.41b | 59.35 ± 20.8a | 74.62 ± 27.33a |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, J.-T.; Chu, M.-Y.; Lin, Y.-S.; Kung, K.-N.; Lin, W.-C.; Lee, M.-J. Root Traits and Biomechanical Properties of Three Tropical Pioneer Tree Species for Forest Restoration in Landslide Areas. Forests 2020, 11, 179. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020179
Lee J-T, Chu M-Y, Lin Y-S, Kung K-N, Lin W-C, Lee M-J. Root Traits and Biomechanical Properties of Three Tropical Pioneer Tree Species for Forest Restoration in Landslide Areas. Forests. 2020; 11(2):179. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020179
Chicago/Turabian StyleLee, Jung-Tai, Ming-Yang Chu, Yu-Syuan Lin, Kuan-Ning Kung, Wen-Chi Lin, and Ming-Jen Lee. 2020. "Root Traits and Biomechanical Properties of Three Tropical Pioneer Tree Species for Forest Restoration in Landslide Areas" Forests 11, no. 2: 179. https://0-doi-org.brum.beds.ac.uk/10.3390/f11020179