Plasticity of Root Traits under Competition for a Nutrient-Rich Patch Depends on Tree Species and Possesses a Large Congruency between Intra- and Interspecific Situations



Abstract
:1. Introduction
2. Material and Methods
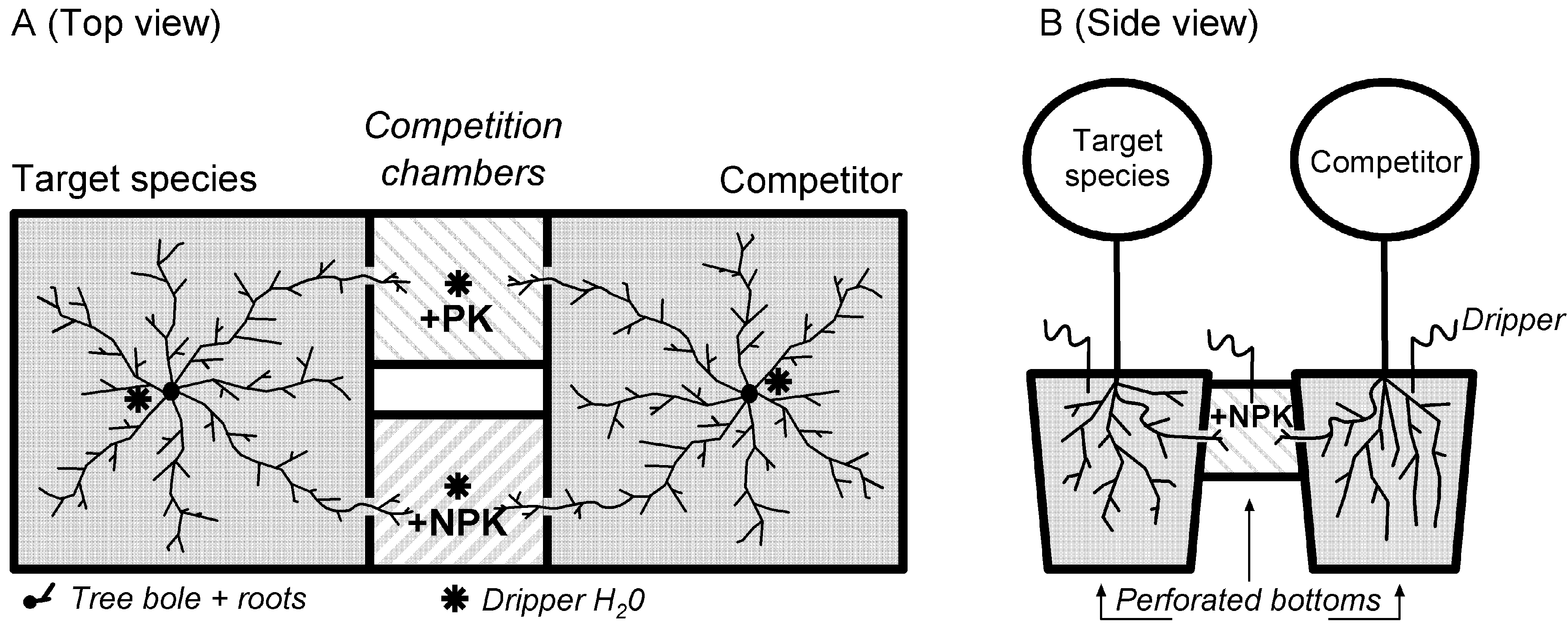
2.1. Experiment Set-Up
2.2. Harvesting of NPK Fertilized Competition Chambers
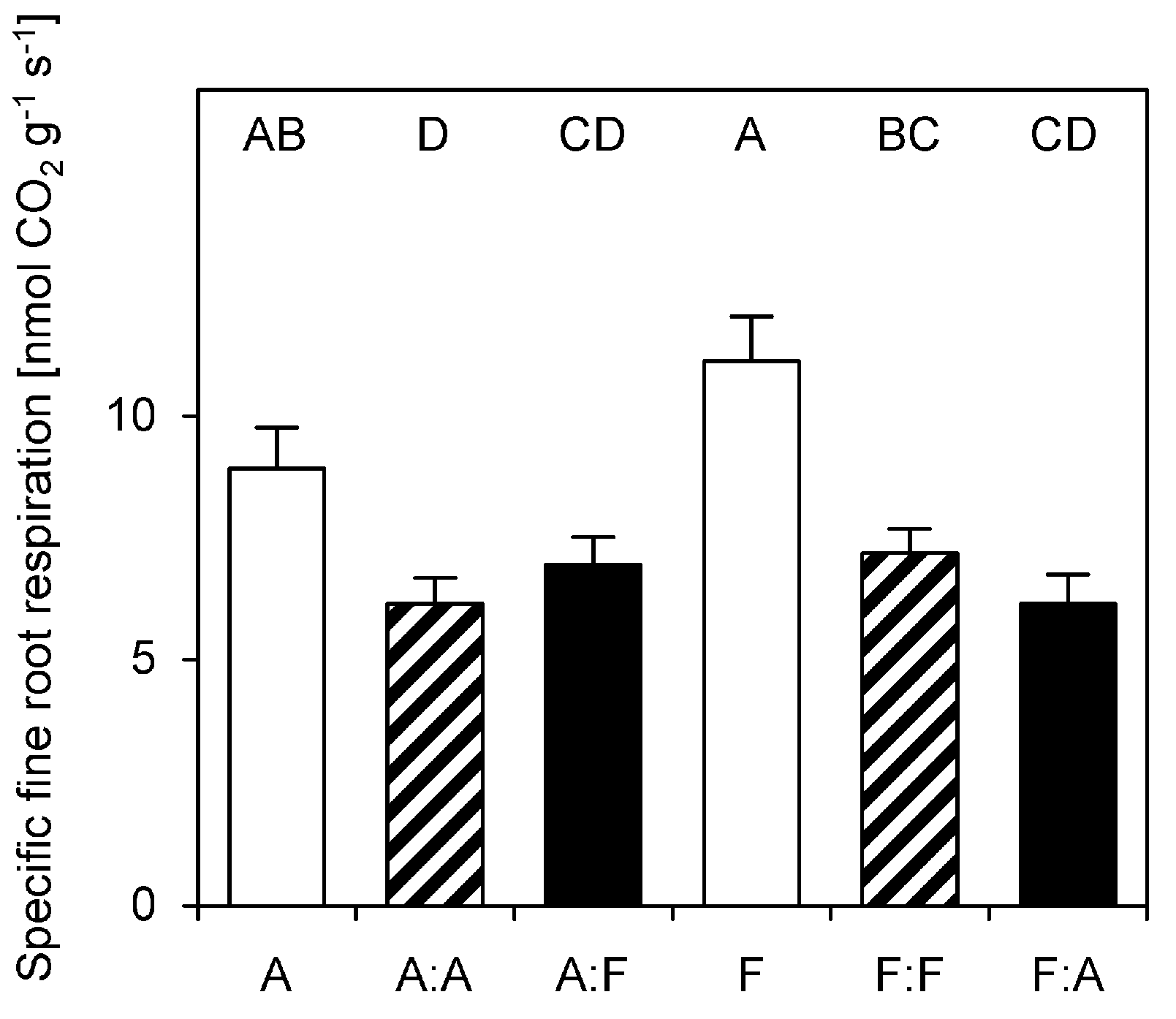
2.3. Specific Fine Root Respiration
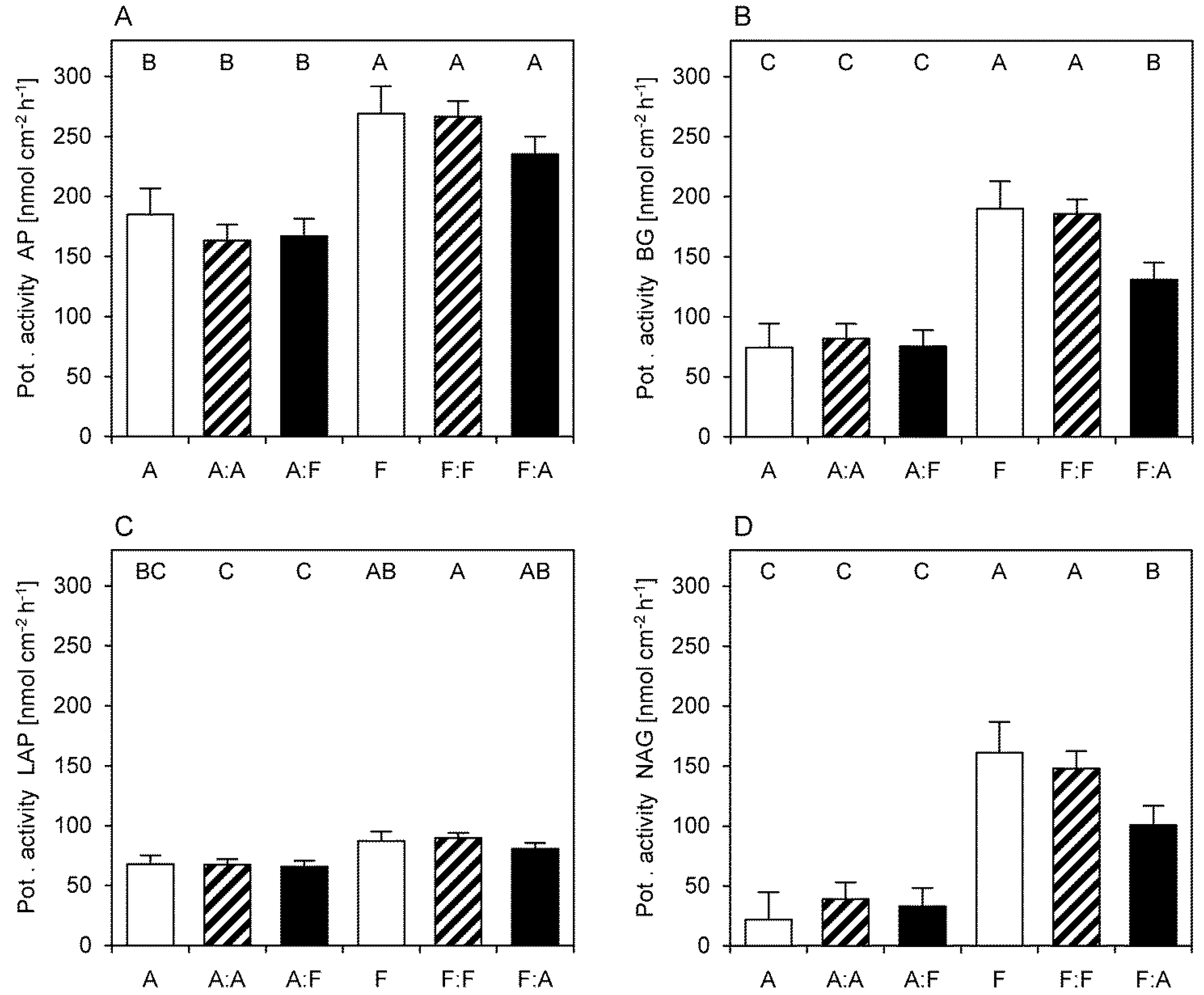
2.4. Potential Enzymatic Activity
2.5. Root Morphology, Biomass and Root Competition Intensity
2.6. Root and Soil Chemical Analysis
2.7. Root Trait Plasticity
2.8. Statistical Analysis
3. Results
3.1. Root Biomass and Competition Intensity
3.2. Fine Root Respiration
3.3. Potential Extracellular Enzymatic Activities
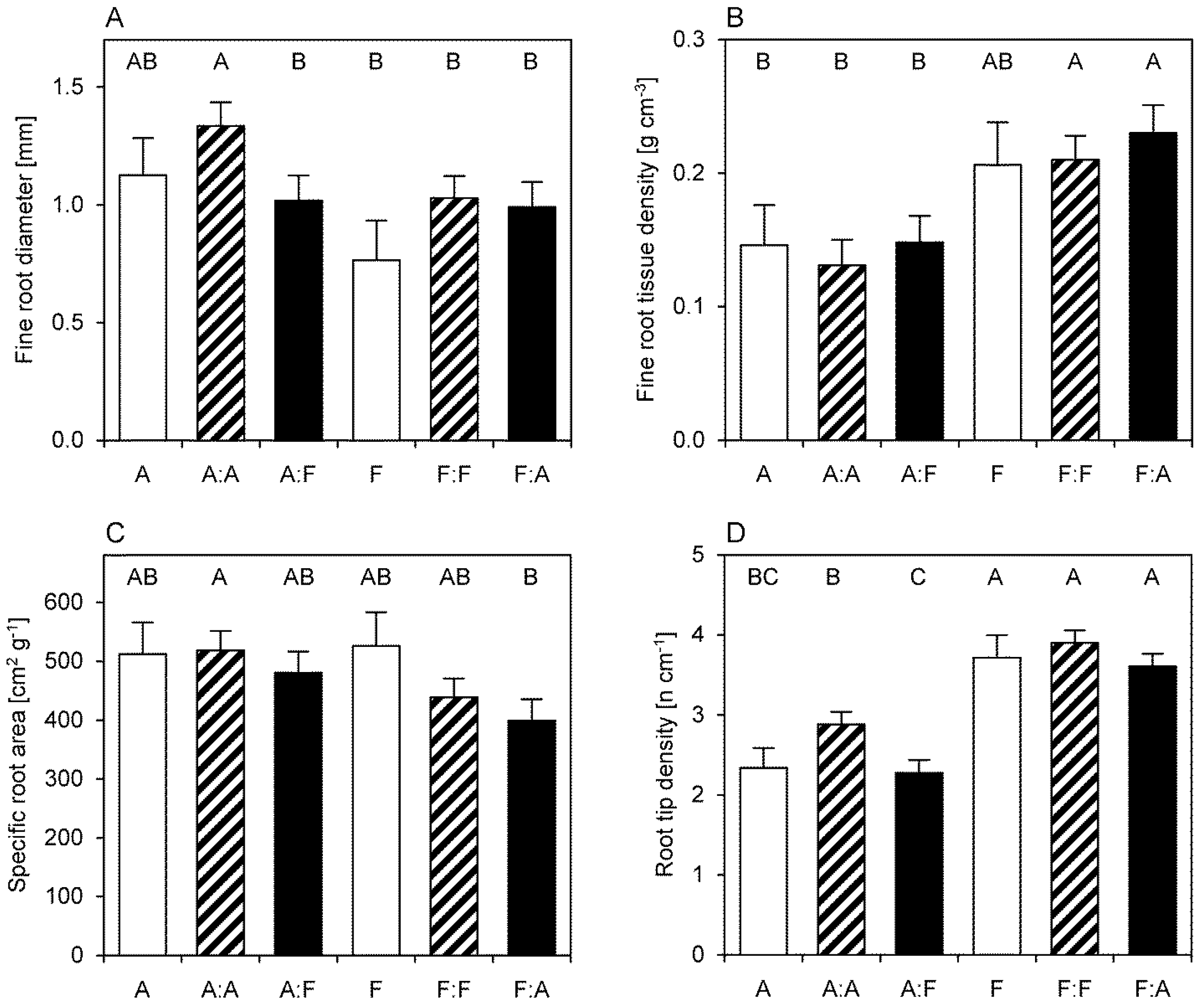
3.4. Fine Root Morphology
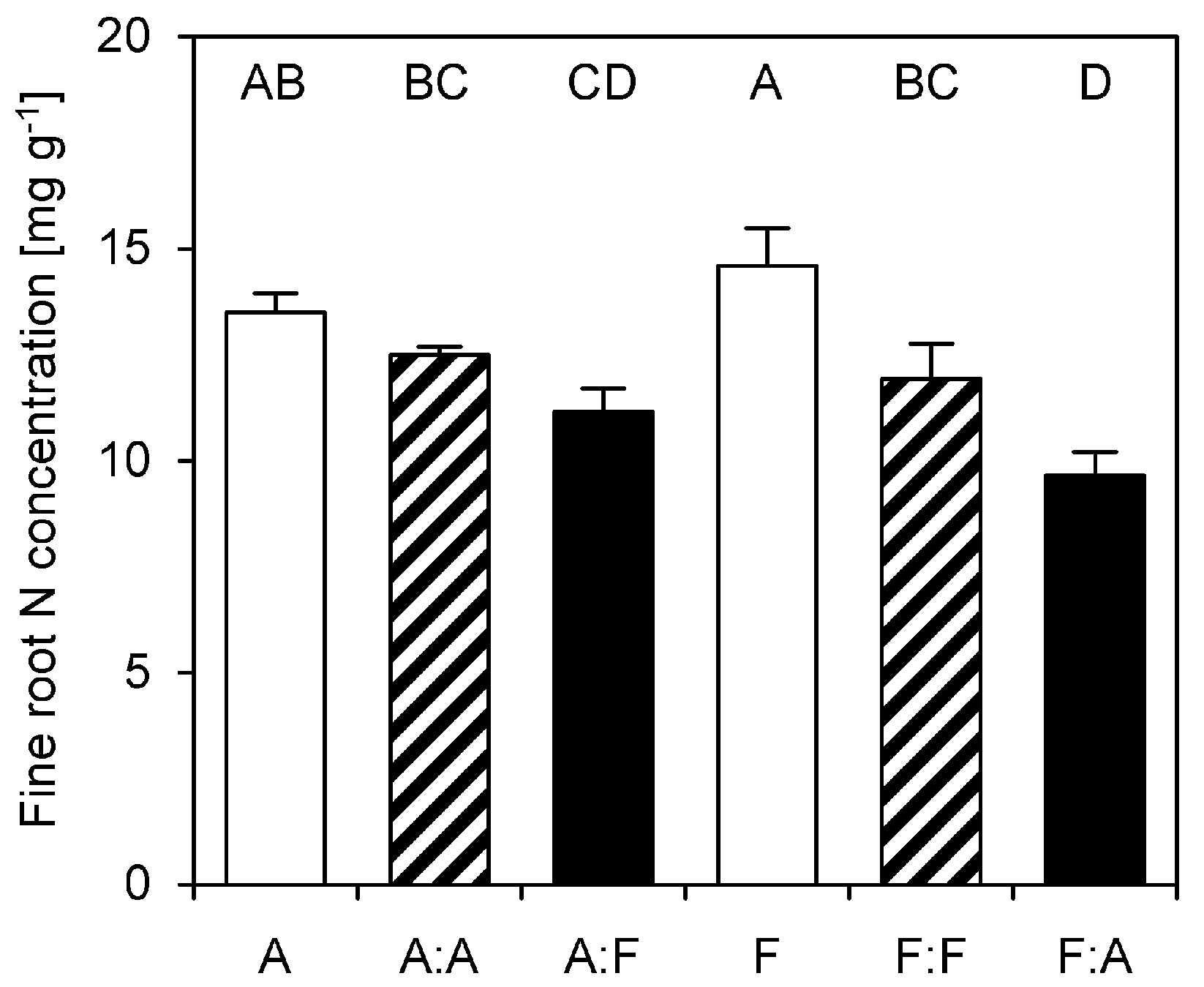
3.5. Fine Root N Concentrations
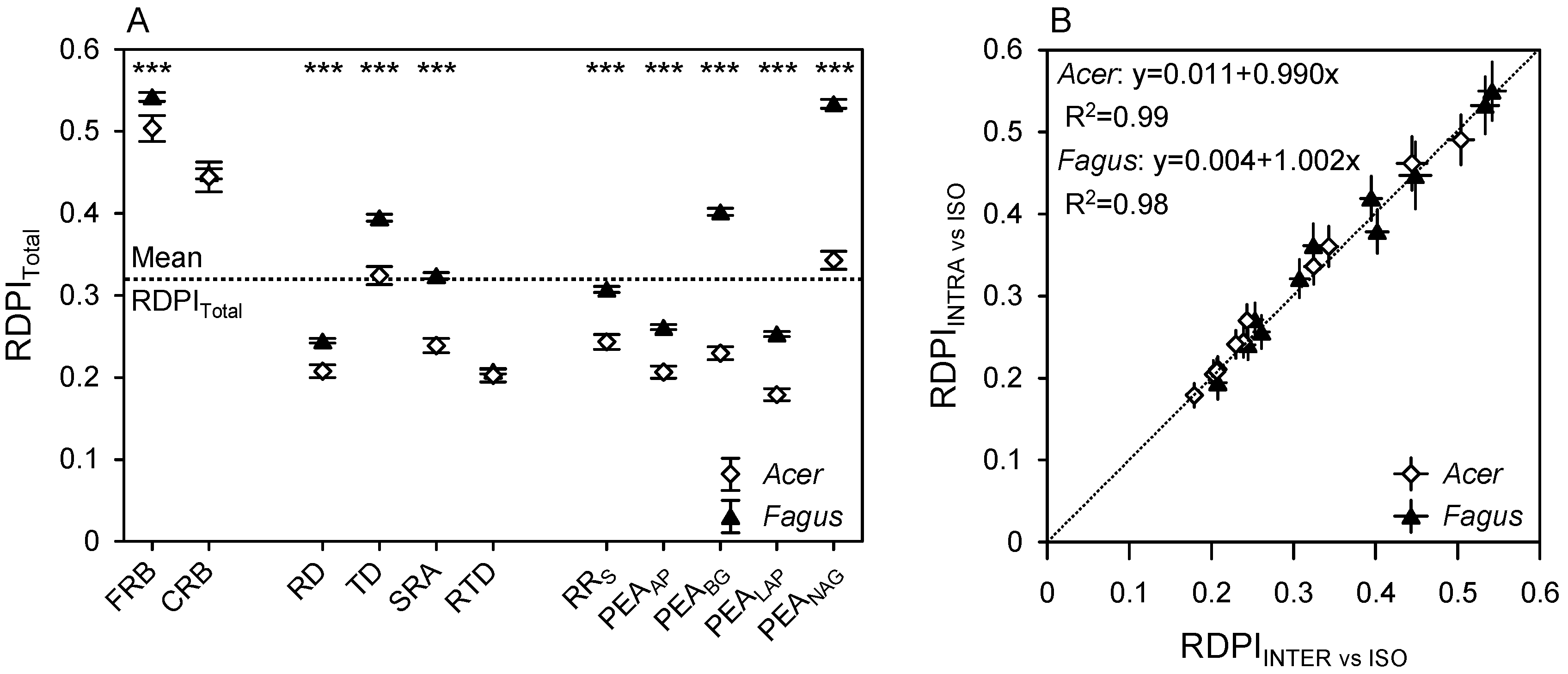
3.6. Plasticity Index
4. Discussion
4.1. Influence of Competition for a Nutrient-Rich Soil Spot on Fine Root Foraging Behaviour, Root Nitrogen Status and Root Trait Characteristics
4.2. Species-Specific Plasticity of Functional Root Traits (under Intra- and Interspecific Competition)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Coomes, D.A.; Grubb, P.J. Impacts of root competition in forests and woodlands: A theoretical framework and review of experiments. Ecol. Monogr. 2000, 70, 171–207. [Google Scholar] [CrossRef]
- Leuschner, C.; Hertel, D.; Coners, H.; Büttner, V. Root competition between beech and oak: A hypothesis. Oecologia 2001, 126, 276–284. [Google Scholar] [CrossRef] [PubMed]
- Rajaniemi, T.K. Evidence for size asymmetry of belowground competition. Basic Appl. Ecol. 2003, 4, 239–247. [Google Scholar] [CrossRef]
- Weiss, L.; Schalow, L.; Jeltsch, F.; Geissler, K. Experimental evidence for root competition effects on community evenness in one of two phytometer species. J. Plant Ecol. 2019, 12, 281–291. [Google Scholar] [CrossRef]
- Donald, C. The interaction of competition for light and for nutrients. Aust. J. Agric. Res. 1958, 9, 421–435. [Google Scholar] [CrossRef]
- Casper, B.B.; Jackson, R.B. Plant competition underground. Annu. Rev. Ecol. Syst. 1997, 28, 545–570. [Google Scholar] [CrossRef] [Green Version]
- Coomes, D.A.; Allen, R.B. Effects of size, competition and altitude on tree growth. J. Ecol. 2007, 95, 1084–1097. [Google Scholar] [CrossRef]
- De Kroon, H.; Mommer, L.; Nishiwaki, A. Root competition: Towards a mechanistic understanding. In Root Ecology; de Kroon, H., Visser, E.J.W., Eds.; Springer: Berlin, Germany, 2003; Volume 168, pp. 215–235. [Google Scholar]
- De Kroon, H.; Hendriks, M.; van Ruijven, J.; Ravenek, J.; Padilla, F.M.; Jongejans, E.; Visser, E.J.W.; Mommer, L. Root responses to nutrients and soil biota: Drivers of species coexistence and ecosystem productivity. J. Ecol. 2012, 100, 6–15. [Google Scholar] [CrossRef]
- Trinder, C.J.; Brooker, R.W.; Robinson, D. Plant ecology’s guilty little secret: Understanding the dynamics of plant competition. Funct. Ecol. 2013, 27, 918–929. [Google Scholar] [CrossRef]
- Rewald, B.; Razaq, M.; Lixue, Y.; Li, J.; Khan, F.; Jie, Z. Root order-based traits of manchurian walnut & larch and their plasticity under interspecific competition. Sci. Rep. 2018, 8, 9815. [Google Scholar]
- Fowler, N. The role of competition in plant communities in arid and semiarid regions. Annu. Rev. Ecol. Syst. 1986, 17, 89–110. [Google Scholar] [CrossRef]
- Wang, P.; Stieglitz, T.; Zhou, D.W.; Cahill, J.F., Jr. Are competitive effect and response two sides of the same coin, or fundamentally different? Funct. Ecol. 2010, 24, 196–207. [Google Scholar] [CrossRef]
- Chesson, P. Mechanisms of maintenance of species diversity. Annu. Rev. Ecol. Syst. 2000, 31, 343–366. [Google Scholar] [CrossRef] [Green Version]
- Rust, S.; Savill, P.S. The root systems of Fraxinus excelsior and Fagus sylvatica and their competitive relationships. Forestry 2000, 73, 499–508. [Google Scholar] [CrossRef]
- Büttner, V.; Leuschner, C. Spatial and temporal patterns of fine-root abundance in a mixed oak beech forest. For. Ecol. Manag. 1994, 70, 11–21. [Google Scholar] [CrossRef]
- Rewald, B.; Leuschner, C. Belowground competition in a broad-leaved temperate mixed forest: Pattern analysis and experiments in a four-species stand. Eur. J. For. Res. 2009, 128, 387–398. [Google Scholar] [CrossRef] [Green Version]
- Chave, J.; Coomes, D.; Jansen, S.; Lewis, S.L.; Swenson, N.G.; Zanne, A.E. Towards a worldwide wood economics spectrum. Ecol. Lett. 2009, 12, 351–366. [Google Scholar] [CrossRef]
- Díaz, S.; Kattge, J.; Cornelissen, J.H.C.; Wright, I.J.; Lavorel, S.; Dray, S.; Reu, B.; Kleyer, M.; Wirth, C.; Prentice, I.C. The global spectrum of plant form and function. Nature 2016, 529, 167–171. [Google Scholar] [CrossRef]
- Bardgett, R.D.; Mommer, L.; De Vries, F.T. Going underground: Root traits as drivers of ecosystem processes. Trends Ecol. Evol. 2014, 29, 692–699. [Google Scholar] [CrossRef]
- Reich, P.B. The world-wide ‘fast–slow’ plant economics spectrum: A traits manifesto. J. Ecol. 2014, 102, 275–301. [Google Scholar] [CrossRef]
- Kunstler, G.; Falster, D.; Coomes, D.A.; Hui, F.; Kooyman, R.M.; Laughlin, D.C.; Poorter, L.; Vanderwel, M.; Vieilledent, G.; Wright, S.J.; et al. Plant functional traits have globally consistent effects on competition. Nature 2015, 529, 204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Forey, E.; Langlois, E.; Lapa, G.; Korboulewsky, N.; Robson, T.M.; Aubert, M. Tree species richness induces strong intraspecific variability of beech (Fagus sylvatica) leaf traits and alleviates edaphic stress. Eur. J. For. Res. 2016, 135, 707–717. [Google Scholar] [CrossRef]
- Laliberté, E. Below-ground frontiers in trait-based plant ecology. New Phytol. 2017, 213, 1597–1603. [Google Scholar] [CrossRef] [PubMed]
- Kirfel, K.; Heinze, S.; Hertel, D.; Leuschner, C. Effects of bedrock type and soil chemistry on the fine roots of european beech—A study on the belowground plasticity of trees. For. Ecol. Manag. 2019, 444, 256–268. [Google Scholar] [CrossRef]
- Freschet, G.T.; Pagès, L.; Iversen, C.M.; Comas, L.H.; Rewald, B.; Roumet, C.; Klimešová, J.; Zadworny, M.; Poorter, H.; Postma, J.A.; et al. A starting guide to root ecology: Strengthening ecological concepts and standardizing root classification, sampling, processing and trait measurements. New Phytol. 2020. under review. [Google Scholar]
- Ostonen, I.; Püttsepp, Ü.; Biel, C.; Alberton, O.; Bakker, M.R.; Lõhmus, K.; Majdi, H.; Metcalfe, D.; Olsthoorn, A.F.M.; Pronk, A.; et al. Specific root length as an indicator of environmental change. Plant Biosyst. 2007, 141, 426–442. [Google Scholar] [CrossRef]
- De la Riva, E.G.; Marañón, T.; Pérez-Ramos, I.M.; Navarro-Fernández, C.M.; Olmo, M.; Villar, R. Root traits across environmental gradients in mediterranean woody communities: Are they aligned along the root economics spectrum? Plant Soil 2018, 424, 35–48. [Google Scholar] [CrossRef]
- Beyer, F.; Hertel, D.; Jung, K.; Fender, A.-C.; Leuschner, C. Competition effects on fine root survival of Fagus sylvatica and Fraxinus excelsior. For. Ecol. Manag. 2013, 302, 14–22. [Google Scholar] [CrossRef]
- Li, X.; Rennenberg, H.; Simon, J. Competition for nitrogen between Fagus sylvatica and Acer pseudoplatanus seedlings depends on soil nitrogen availability. Front. Plant Sci. 2015, 6, 302. [Google Scholar] [CrossRef] [Green Version]
- Mayfield, M.M.; Levine, J.M. Opposing effects of competitive exclusion on the phylogenetic structure of communities. Ecol. Lett. 2010, 13, 1085–1093. [Google Scholar] [CrossRef]
- Callaway, R.M.; Pennings, S.C.; Richards, C.L. Phenotypic plasticity and interactions among plants. Ecology 2003, 84, 1115–1128. [Google Scholar] [CrossRef] [Green Version]
- Violle, C.; Enquist, B.J.; McGill, B.J.; Jiang, L.; Albert, C.H.; Hulshof, C.; Jung, V.; Messier, J. The return of the variance: Intraspecific variability in community ecology. Trends Ecol. Evol. 2012, 27, 244–252. [Google Scholar] [CrossRef] [PubMed]
- De Bello, F.; Carmona, C.P.; Mason, N.W.; Sebastià, M.T.; Lepš, J. Which trait dissimilarity for functional diversity: Trait means or trait overlap? J. Veg. Sci. 2013, 24, 807–819. [Google Scholar] [CrossRef]
- Schmid, I.; Kazda, M. Root distribution of Norway spruce in monospecific and mixed stands on different soils. For. Ecol. Manag. 2002, 159, 37–47. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.J.W.; van Ruijven, J.; Kuyper, T.W.; Mohren, G.M.J.; Sterck, F.J. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef] [Green Version]
- Kong, D.; Wang, J.; Wu, H.; Valverde-Barrantes, O.J.; Wang, R.; Zeng, H.; Kardol, P.; Zhang, H.; Feng, Y. Nonlinearity of root trait relationships and the root economics spectrum. Nat. Commun. 2019, 10, 2203. [Google Scholar] [CrossRef] [Green Version]
- Farrior, C.E. Theory predicts plants grow roots to compete with only their closest neighbours. Proc. R. Soc. B Biol. Sci. 2019, 286, 20191129. [Google Scholar] [CrossRef] [Green Version]
- Faillace, C.A.; Caplan, J.S.; Grabosky, J.C.; Morin, P.J. Beneath it all: Size, not origin, predicts belowground competitive ability in exotic and native shrubs. J. Torrey Bot. Soc. 2018, 145, 30–40. [Google Scholar] [CrossRef]
- Valverde-Barrantes, O.J.; Smemo, K.A.; Feinstein, L.M.; Kershner, M.W.; Blackwood, C.B. Aggregated and complementary: Symmetric proliferation, overyielding, and mass effects explain fine-root biomass in soil patches in a diverse temperate deciduous forest landscape. New Phytol. 2015, 205, 731–742. [Google Scholar] [CrossRef] [Green Version]
- Nicholson, B.A.; Jones, M.D. Early-successional ectomycorrhizal fungi effectively support extracellular enzyme activities and seedling nitrogen accumulation in mature forests. Mycorrhiza 2017, 27, 247–260. [Google Scholar] [CrossRef]
- Paul, C.; Brandl, S.; Friedrich, S.; Falk, W.; Härtl, F.; Knoke, T. Climate change and mixed forests: How do altered survival probabilities impact economically desirable species proportions of Norway spruce and European beech? Ann. For. Sci. 2019, 76, 14. [Google Scholar] [CrossRef] [Green Version]
- Silva Pedro, M.; Rammer, W.; Seidl, R. Tree species diversity mitigates disturbance impacts on the forest carbon cycle. Oecologia 2015, 177, 619–630. [Google Scholar] [CrossRef] [PubMed]
- Petritan, A.M.; Von Lüpke, B.; Petritan, I.C. Effects of shade on growth and mortality of maple (Acer pseudoplatanus), ash (Fraxinus excelsior) and beech (Fagus sylvatica) saplings. For. Int. J. For. Res. 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Tinya, F.; Márialigeti, S.; Bidló, A.; Ódor, P. Environmental drivers of the forest regeneration in temperate mixed forests. For. Ecol. Manag. 2019, 433, 720–728. [Google Scholar] [CrossRef] [Green Version]
- Rewald, B.; Ammer, C.; Hartmann, H.; Malyshev, A.V.; Meier, I.C. Editorial: Woody plants and forest ecosystems in a complex world—Ecological interactions and physiological functioning above and below ground. Front. Plant Sci. 2020. under review. [Google Scholar] [CrossRef]
- Ellenberg, H.; Leuschner, C. Vegetation Mitteleuropas Mit den Alpen, 6th ed.; Ulmer: Stuttgart, Germany, 2010; p. 1357. (In German) [Google Scholar]
- Collet, C.; Fournier, M.; Ningre, F.; Hounzandji, A.P.-I.; Constant, T. Growth and posture control strategies in Fagus sylvatica and Acer pseudoplatanus saplings in response to canopy disturbance. Ann. Bot. 2011, 107, 1345–1353. [Google Scholar] [CrossRef] [Green Version]
- Brundrett, M.C. Mycorrhizal associations and other means of nutrition of vascular plants: Understanding the global diversity of host plants by resolving conflicting information and developing reliable means of diagnosis. Plant Soil 2009, 320, 37–77. [Google Scholar] [CrossRef]
- Phillips, R.P.; Brzostek, E.; Midgley, M.G. The mycorrhizal-associated nutrient economy: A new framework for predicting carbon-nutrient couplings in temperate forests. New Phytol. 2013, 199, 41–51. [Google Scholar] [CrossRef]
- Lindahl, B.D.; Tunlid, A. Ectomycorrhizal fungi-potential organic matter decomposers, yet not saprotrophs. New Phytol. 2015, 205, 1443–1447. [Google Scholar] [CrossRef] [PubMed]
- Read, D.; Perez-Moreno, J. Mycorrhizas and nutrient cycling in ecosystems—A journey towards relevance? New Phytol. 2003, 157, 475–492. [Google Scholar] [CrossRef]
- Simon, J.; Li, X.; Rennenberg, H. Competition for nitrogen between european beech and sycamore maple shifts in favour of beech with decreasing light availability. Tree Physiol. 2014, 34, 49–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hommel, R.; Siegwolf, R.; Zavadlav, S.; Arend, M.; Schaub, M.; Galiano, L.; Haeni, M.; Kayler, Z.E.; Gessler, A. Impact of interspecific competition and drought on the allocation of new assimilates in trees. Plant Biol. 2016, 18, 785–796. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.; Dannenmann, M.; Pena, R.; Gessler, A.; Rennenberg, H. Nitrogen nutrition of beech forests in a changing climate: Importance of plant-soil-microbe water, carbon, and nitrogen interactions. Plant Soil 2017, 418, 89–114. [Google Scholar] [CrossRef]
- Lang, C.; Seven, J.; Polle, A. Host preferences and differential contributions of deciduous tree species shape mycorrhizal species richness in a mixed central european forest. Mycorrhiza 2011, 21, 297–308. [Google Scholar] [CrossRef] [Green Version]
- Madsen, P.; Larsen, J.B. Natural regeneration of beech (Fagus sylvatica L.) with respect to canopy density, soil moisture and soil carbon content. For. Ecol. Manag. 1997, 97, 95–105. [Google Scholar] [CrossRef]
- Poorter, H.; Bühler, J.; van Dusschoten, D.; Climent, J.; Postma, J.A. Pot size matters: A meta-analysis of the effects of rooting volume on plant growth. Funct. Plant Biol. 2012, 39, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Pritsch, K.; Garbaye, J. Enzyme secretion by ecm fungi and exploitation of mineral nutrients from soil organic matter. Ann. For. Sci. 2011, 68, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Otgonsuren, B.; Rewald, B.; Godbold, D.L.; Göransson, H. Ectomycorrhizal inoculation of populus nigra modifies the response of absorptive root respiration and root surface enzyme activity to salinity stress. Flora Morphol. Distrib. Funct. Ecol. Plants 2016, 224, 123–129. [Google Scholar] [CrossRef]
- Wilson, S.D.; Keddy, P.A. Measuring diffuse competition along an environmental gradient—Results from a shoreline plant community. Am. Nat. 1986, 127, 862–869. [Google Scholar] [CrossRef]
- Grace, J.B. On the measurement of plant competition intensity. Ecology 1995, 76, 305–308. [Google Scholar] [CrossRef]
- Valladares, F.; Sanchez-Gomez, D.; Zavala, M.A. Quantitative estimation of phenotypic plasticity: Bridging the gap between the evolutionary concept and its ecological applications. J. Ecol. 2006, 94, 1103–1116. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2019. [Google Scholar]
- Ameztegui, A. Plasticity: An R Package to Determine Several Plasticity Indices. 2017. GitHub: GitHub repository. Available online: https://github.com/ameztegui/Plasticity (accessed on 15 April 2019).
- Moran, M.D. Arguments for rejecting the sequential bonferroni in ecological studies. Oikos 2003, 100, 403–405. [Google Scholar] [CrossRef]
- Kubisch, P.; Hertel, D.; Leuschner, C. Fine root productivity and turnover of ectomycorrhizal and arbuscular mycorrhizal tree species in a temperate broad-leaved mixed forest. Front. Plant Sci. 2016, 7, 301. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bolte, A.; Villanueva, I. Interspecific competition impacts on the morphology and distribution of fine roots in european beech (Fagus sylvatica L.) and norway spruce (Picea abies (L.) karst.). Eur. J. For. Res. 2006, 125, 15–26. [Google Scholar] [CrossRef]
- Zwetsloot, M.J.; Goebel, M.; Paya, A.; Grams, T.E.E.; Bauerle, T.L. Specific spatio-temporal dynamics of absorptive fine roots in response to neighbor species identity in a mixed beech-spruce forest. Tree Physiol. 2019, in press. [Google Scholar] [CrossRef]
- Meinen, C.; Hertel, D.; Leuschner, C. Biomass and morphology of fine roots in temperate broad-leaved forests differing in tree species diversity: Is there evidence of below-ground overyielding? Oecologia 2009, 161, 99–111. [Google Scholar] [CrossRef] [Green Version]
- Rewald, B.; Leuschner, C. Does root competition asymmetry increase with water availability? Plant Ecol. Divers. 2009, 2, 255–264. [Google Scholar] [CrossRef]
- Dudley, S.A.; Murphy, G.P.; File, A.L. Kin recognition and competition in plants. Funct. Ecol. 2013, 27, 898–906. [Google Scholar] [CrossRef]
- Hertel, D.; Leuschner, C. The in situ root chamber: A novel tool for the experimental analysis of root competition in forest soils. Pedobiologia 2006, 50, 217–224. [Google Scholar] [CrossRef]
- Fotelli, M.N.; Rudolph, P.; Rennenberg, H.; Geáler, A. Irradiance and temperature affect the competitive interference of blackberry on the physiology of european beech seedlings. New Phytol. 2005, 165, 453–462. [Google Scholar] [CrossRef]
- Simon, J.; Waldhecker, P.; Brüggemann, N.; Rennenberg, H. Competition for nitrogen sources between european beech (Fagus sylvatica) and sycamore maple (Acer pseudoplatanus) seedlings. Plant Biol. 2010, 12, 453–458. [Google Scholar] [CrossRef] [PubMed]
- Fotelli, M.N.; Rennenberg, H.; Geáler, A. Effects of drought on the competitive interference of an early successional species (Rubus fruticosus) on Fagus sylvatica L. Seedlings: N-15 uptake and partitioning, responses of amino acids and other n compounds. Plant Biol. 2002, 4, 311–320. [Google Scholar] [CrossRef]
- Rewald, B.; Rechenmacher, A.; Godbold, D.L. It’s complicated: Intra-root system variability of respiration and morphological traits in four deciduous tree species. Plant Physiol. 2014, 166, 736–745. [Google Scholar] [CrossRef] [Green Version]
- Meier, I.C.; Angert, A.; Falik, O.; Shelef, O.; Rachmilevitch, S. Increased root oxygen uptake in pea plants responding to non-self neighbors. Planta 2013, 238, 577–586. [Google Scholar] [CrossRef] [PubMed]
- Sandén, H.; Mayer, M.; Stark, S.; Sandén, T.; Nilsson, L.O.; Jepsen, J.U.; Wäli, P.R.; Rewald, B. Moth outbreaks reduce decomposition in subarctic forest soils. Ecosystems 2019, in press. [Google Scholar]
- Cheeke, T.E.; Phillips, R.P.; Brzostek, E.R.; Rosling, A.; Bever, J.D.; Fransson, P. Dominant mycorrhizal association of trees alters carbon and nutrient cycling by selecting for microbial groups with distinct enzyme function. New Phytol. 2017, 214, 432–442. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Yan, L.; Korpelainen, H.; Niinemets, Ü.; Li, C. Plant-plant interactions and N fertilization shape soil bacterial and fungal communities. Soil Biol. Biochem. 2019, 128, 127–138. [Google Scholar] [CrossRef]
- Curt, T.; Coll, L.; Prévosto, B.; Balandier, P.; Kunstler, G. Plasticity in growth, biomass allocation and root morphology in beech seedlings as induced by irradiance and herbaceous competition. Ann. For. Sci. 2005, 62, 51–60. [Google Scholar] [CrossRef] [Green Version]
- Bloom, A.J.; Chapin, F.S., ІІІ; Mooney, H.A. Resource limitation in plants—An economic analogy. Annu. Rev. Ecol. Syst. 1985, 16, 363–392. [Google Scholar] [CrossRef]
- Bauhus, J.; Messier, C. Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of Eastern Canada. Can. J. For. Res. 1999, 29, 260–273. [Google Scholar] [CrossRef]
- Wang, W.; Wang, Y.; Hoch, G.; Wang, Z.; Gu, J. Linkage of root morphology to anatomy with increasing nitrogen availability in six temperate tree species. Plant Soil 2018, 425, 189–200. [Google Scholar] [CrossRef]
- Lei, P.; Scherer-Lorenzen, M.; Bauhus, J. Belowground facilitation and competition in young tree species mixtures. For. Ecol. Manag. 2012, 265, 191–200. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B.; et al. Redefining fine roots improves understanding of belowground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Iversen, C.M.; McCormack, M.L.; Powell, A.S.; Blackwood, C.B.; Freschet, G.T.; Kattge, J.; Roumet, C.; Stover, D.B.; Soudzilovskaia, N.A.; Valverde-Barrantes, O.J. A global fine-root ecology database to address below-ground challenges in plant ecology. New Phytol. 2017, 215, 15–26. [Google Scholar] [CrossRef] [Green Version]
- Kattge, J.; DÍAz, S.; Lavorel, S.; Prentice, I.C.; Leadley, P.; BÖNisch, G.; Garnier, E.; Westoby, M.; Reich, P.B.; Wright, I.J.; et al. Try—A global database of plant traits. Glob. Chang. Biol. 2011, 17, 2905–2935. [Google Scholar] [CrossRef]
- Petriţan, A.M.; von Lüpke, B.; Petriţan, I.C. Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur. J. For. Res. 2009, 128, 61–74. [Google Scholar] [CrossRef] [Green Version]
- Keel, S.G.; Campbell, C.D.; Högberg, M.N.; Richter, A.; Wild, B.; Zhou, X.; Hurry, V.; Linder, S.; Näsholm, T.; Högberg, P. Allocation of carbon to fine root compounds and their residence times in a boreal forest depend on root size class and season. New Phytol. 2012, 194, 972–981. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Root Competition § | Target Species’ Root | Belowground Competitor | Treatment (Abbrev.) | Realized Replication (n) |
|---|---|---|---|---|
| Isolation (ISO; no competition) | Acer | none | A | 18 |
| Fagus | none | F | 16 | |
| Intraspecific competition (INTRA) | Acer | Acer | A:A | 48 |
| Fagus | Fagus | F:F | 52 | |
| Interspecific competition (INTER) | Acer | Fagus | A:F | 40 * |
| Fagus | Acer | F:A | 39 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lak, Z.A.; Sandén, H.; Mayer, M.; Godbold, D.L.; Rewald, B. Plasticity of Root Traits under Competition for a Nutrient-Rich Patch Depends on Tree Species and Possesses a Large Congruency between Intra- and Interspecific Situations. Forests 2020, 11, 528. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050528
Lak ZA, Sandén H, Mayer M, Godbold DL, Rewald B. Plasticity of Root Traits under Competition for a Nutrient-Rich Patch Depends on Tree Species and Possesses a Large Congruency between Intra- and Interspecific Situations. Forests. 2020; 11(5):528. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050528
Chicago/Turabian StyleLak, Zana A., Hans Sandén, Mathias Mayer, Douglas L. Godbold, and Boris Rewald. 2020. "Plasticity of Root Traits under Competition for a Nutrient-Rich Patch Depends on Tree Species and Possesses a Large Congruency between Intra- and Interspecific Situations" Forests 11, no. 5: 528. https://0-doi-org.brum.beds.ac.uk/10.3390/f11050528