Biomass Allocation to Resource Acquisition Compartments Is Affected by Tree Density Manipulation in European Beech after Three Decades


Abstract
:1. Introduction
2. Material and Methods
2.1. Site and Stand Conditions
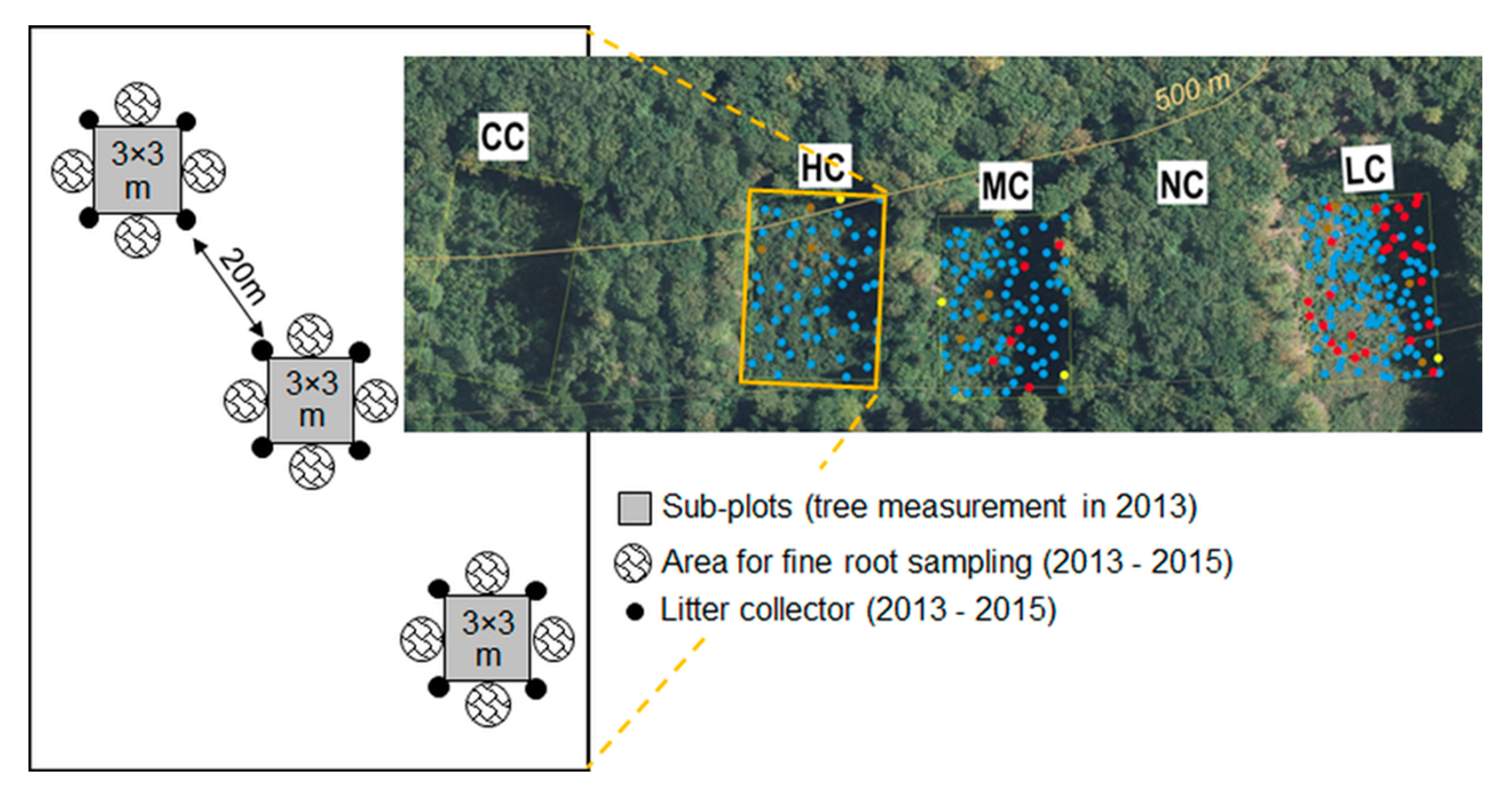
2.2. Plot and Subplot Design
2.3. Tree Measurements
2.4. Fine Root Sampling and Processing
2.5. Foliage Sampling and Processing
2.6. Calculations and Statistical Analyses
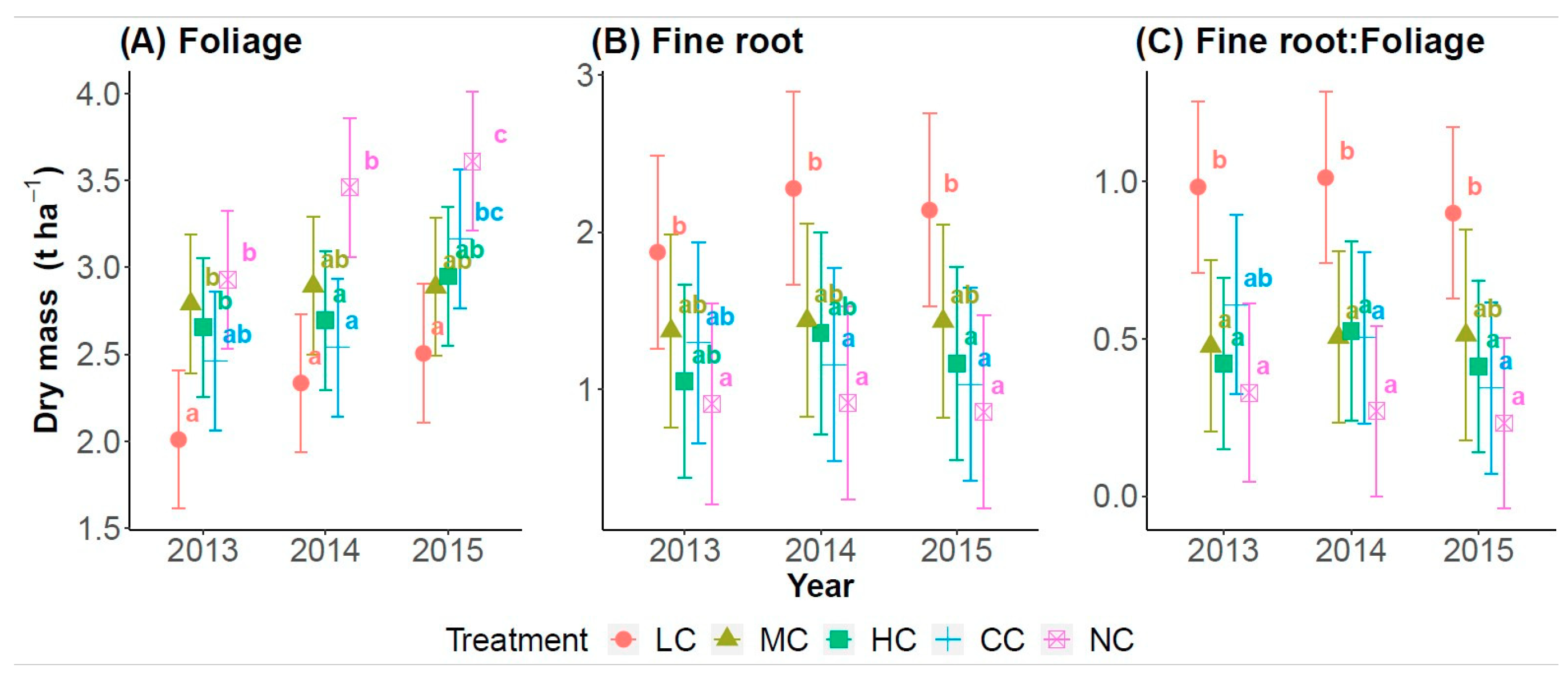
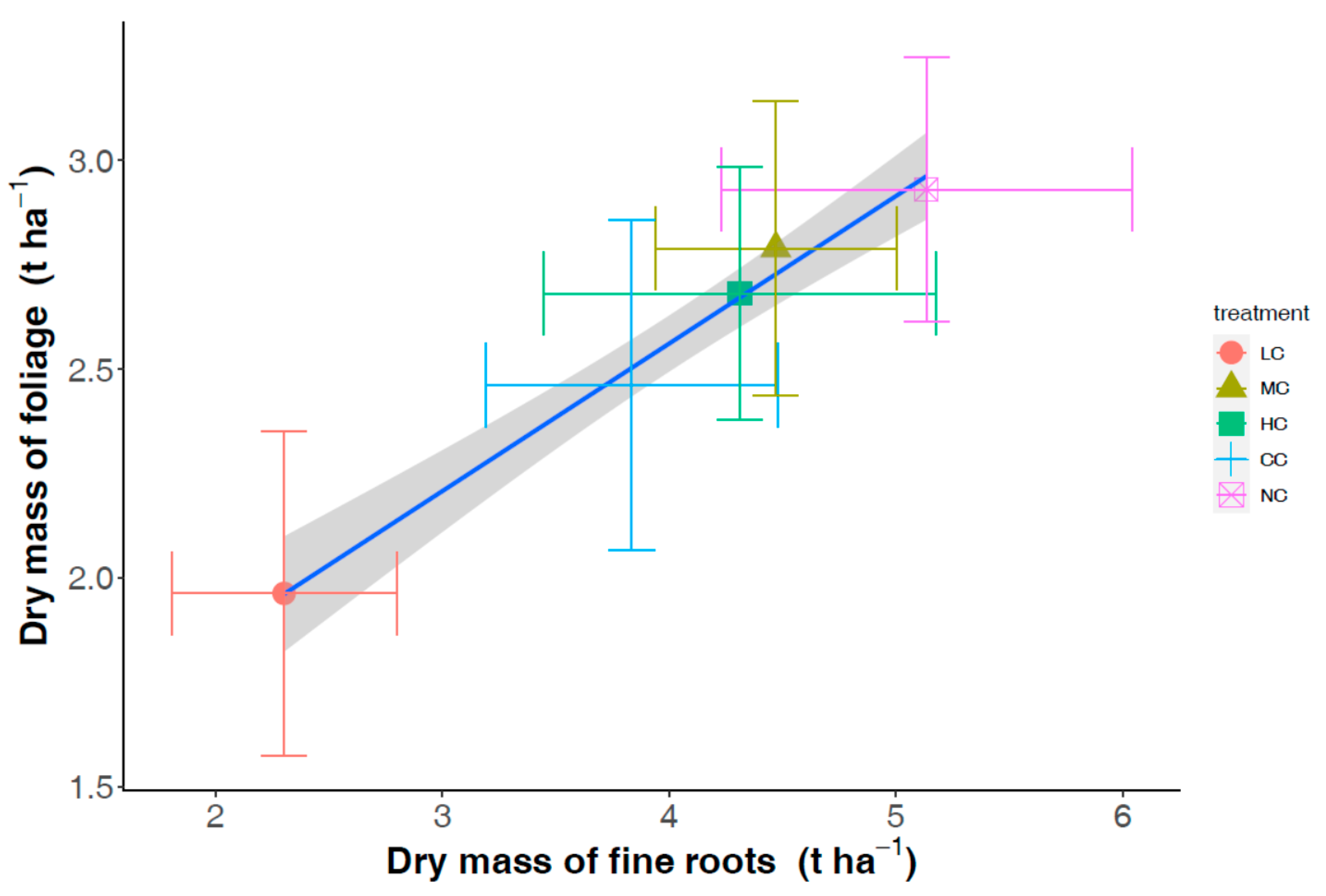
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- De Rigo, D.; Caudullo, G.; Houston Durrant, T.; Mauri, A. The European Atlas of Forest Tree Species: Modelling, Data and Information on Forest Tree Species. Eur. Atlas For. Tree Species 2016. [Google Scholar] [CrossRef]
- Ministry of Agriculture. Green Report. Report on the Status of Forestry in the Slovak Republic; Ministry of Agriculture of the Slovak Republic: Staré Mesto, Slovakia, 2019.
- Barna, M.; Kulfan, J.; Bublinec, E. Beech and Beech Ecosystems of Slovakia; Veda: Bratislava, Slovakia, 2011. [Google Scholar]
- Hlásny, T.; Mátyás, C.; Seidl, R.; Kulla, L.; Merganičová, K.; Trombik, J.; Dobor, L.; Barcza, Z.; Konôpka, B. Climate Change Increases the Drought Risk in Central European Forests: What Are the Options for Adaptation? Cent. Eur. For. J. 2014, 60, 5–18. [Google Scholar] [CrossRef] [Green Version]
- Lindner, M.M.; Maroschek, S.; Netherer, A.; Kremer, A.; Barbati, J.; Garcia-Gonzalo, R.; Seidl, S.; Delzon, P.; Corona, P.; Kolström, M.; et al. Climate Change Impacts, Adaptive Capacity, and Vulnerability of European Forest Ecosystems. For. Ecol. Manag. 2010, 259, 698–709. [Google Scholar] [CrossRef]
- Condés, S.; Sterba, H.; Aguirre, A.; Bielak, K.; Bravo-Oviedo, A.; Coll, L.; Pach, M.; Pretzsch, H.; Vallet, P.; Del Río, M. Estimation and Uncertainty of the Mixing Effects on Scots Pine—European Beech Productivity from National Forest Inventories Data. Forests 2018, 9, 518. [Google Scholar] [CrossRef] [Green Version]
- Petritan, A.M.; Von Lüpke, B.; Petritan, I.C. Effects of Shade on Growth and Mortality of Maple (Acer Pseudoplatanus), Ash (Fraxinus Excelsior) and Beech (Fagus Sylvatica) Saplings. Forestry 2007, 80, 397–412. [Google Scholar] [CrossRef]
- Jarcuska, B.; Barna, M. Influence of Light Availability on Height Growth of Naturally Regenerated Beech with Different Growth Histories. Austrian J. For. Sci. 2011, 128, 53–65. [Google Scholar]
- Man, R.; Kayahara, G.J.; Rice, J.A.; MacDonald, G.B. Eleven-Year Responses of a Boreal Mixedwood Stand to Partial Harvesting: Light, Vegetation, and Regeneration Dynamics. For. Ecol. Manag. 2008, 255, 697–706. [Google Scholar] [CrossRef]
- Kreutzweiser, D.P.; Hazlett, P.W.; Gunn, J.M. Logging Impacts on the Biogeochemistry of Boreal Forest Soils and Nutrient Export to Aquatic Systems: A Review. Environ. Rev. 2008, 16, 157–179. [Google Scholar] [CrossRef]
- Wilson, S.D.; Tilman, D. Plant Competition and Resource Availability in Response to Disturbance and Fertilization. Ecology 1993, 74, 599–611. [Google Scholar] [CrossRef]
- Brunner, I.; Bakker, M.R.; Björk, R.G.; Hirano, Y.; Lukac, M.; Aranda, X.; Børja, I.; Eldhuset, T.D.; Helmisaari, H.S.; Jourdan, C.; et al. Fine-Root Turnover Rates of European Forests Revisited: An Analysis of Data from Sequential Coring and Ingrowth Cores. Plant Soil 2013, 362, 357–372. [Google Scholar] [CrossRef]
- Schulze, E.-D.; Chapin, E. Plant Specialization to Environments of Different Resource Availability. In Potentials and Limitations of Ecosystem Analysis; Springer: Berlin/Heidelberg, Germany, 1987; pp. 120–148. [Google Scholar]
- Konôpka, B.; Pajtík, J.; Šebeň, V.; Surový, P.; Merganičová, K. Biomass Allocation into Woody Parts and Foliage in Young Common Aspen (Populus Tremula L.)—Trees and a Stand-Level Study in the Western Carpathians. Forests 2020, 11, 464. [Google Scholar] [CrossRef] [Green Version]
- Fleck, S.; Čater, M. Manual on Methods and Criteria for Harmonized Sampling, Assessment, Monitoring and Analysis of the Effects of Air Pollution on Forests; Federal Research Centre for Forestry and Forest Products: Braunschweig, Germany, 2016. [Google Scholar]
- Lukac, M. Fine Root Turnover. In Measuring Roots; Springer: Berlin/Heidelberg, Germany, 2012; pp. 363–373. [Google Scholar]
- Van Do, T.; Sato, T.; Saito, S.; Kozan, O. Fine-Root Production and Litterfall: Main Contributions to Net Primary Production in an Old-Growth Evergreen Broad-Leaved Forest in Southwestern Japan. Ecol. Res. 2015, 30, 921–930. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Chen, J.M.; Yang, X.; Fan, W.; Li, M.; He, L. Influence of Site Index on the Relationship between Forest Net Primary Productivity and Stand Age. PLoS ONE 2017, 12. [Google Scholar] [CrossRef] [Green Version]
- Kukla, J.; Bublinec, E.; Schieber, B.; Kellerová, D.; Bičárová, S.; Janík, R. Immission-Load-Related Dynamics of Sulphur in Precipitation and in Lysimetric Solutions Penetrating through Beech Ecosystems. Folia Oecol. 2017, 44, 96–106. [Google Scholar] [CrossRef] [Green Version]
- Kukla, J.; Kontriš, J.; Kontrišová, O.; Gregor, J.; Mihálik, A. Causes of floristical differentiation of Dentario bulbiferae-Fagetum (Zlatnik 1935) Hartmann 1953 and Carici pilosae-Fagetum Oberd. Ekol. Bratisl. 1998, 17, 177–186. [Google Scholar]
- Smith, D.M.; Larsen, B.C.; Kelty, M.J.; Ashton, P.M.S. The Practice of Silvicul-Ture: Applied Forest Ecology, 9th ed.; John Wileyand Sons: Hoboken, NJ, USA, 1997; p. 537. [Google Scholar]
- Barna, M.; Sedmák, R.; Marušák, R. Response of European beech radial growth to shelterwood cutting. Folia Oecol. 2010, 37, 125–136. [Google Scholar]
- Barna, M.; Schieber, B.; Cicák, A. Effect of post-cutting changes in site conditions on the morphology and phenology of naturally regenerated beech seedlings (Fagus sylvatica L.). Pol. J. Ecol. 2009, 57, 461–472. [Google Scholar]
- Reineke, L.H. Perfecting a stand-density index for even-aged forests. J. Agric. Res. 1933, 46, 627–638. [Google Scholar]
- Sterba, H. The precision of species proportion by area when estimated by angle counts and yield tables. Forestry 1998, 71, 25–32. [Google Scholar] [CrossRef] [Green Version]
- Koulelis, P.P.; Daskalakou, E.N.; Ioannidis, K.E. Impact of Regional Climatic Conditions on Tree Growth on Mainland Greece. Folia Oecol. 2019, 46, 127–136. [Google Scholar] [CrossRef] [Green Version]
- Barna, M.; Bosela, M. Tree Species Diversity Change in Natural Regeneration of a Beech Forest under Different Management. For. Ecol. Manag. 2015, 342, 93–102. [Google Scholar] [CrossRef]
- Vogt, K.A.; Vogt, D.J.; Bloomfield, J. Analysis of Some Direct and Indirect Methods for Estimating Root Biomass and Production of Forests at an Ecosystem Level. In Root Demographics and Their Efficiencies in Sustainable Agriculture, Grasslands and Forest Ecosystems; Springer: Berlin/Heidelberg, Germany, 1998; pp. 687–720. [Google Scholar]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using Lme4. arXiv 2014, arXiv:1406.5823. [Google Scholar]
- Team, R.D. R: A Language and Environment for Statistical Computing. Vienna Austria R Found. Stat. Comput. 2004. Available online: https://www.R-project.org/ (accessed on 25 October 2019).
- Satterthwaite, F.E. An Approximate Distribution of Estimates of Variance Components. Biom. Bull. 1946, 2, 110–114. [Google Scholar] [CrossRef]
- Russell, L. Emmeans: Estimated Marginal Means, Aka Least-Squares Means. R Package Version 2018, 1, 3. [Google Scholar]
- Becker, R. The New S Language; CRC Press: Boca Raton, FL, USA, 2018. [Google Scholar]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer: Berlin/Heidelberg, Germany, 2016. [Google Scholar]
- Konôpka, B.; Lukac, M. Moderate Drought Alters Biomass and Depth Distribution of Fine Roots in Norway Spruce. For. Pathol. 2013, 43, 115–123. [Google Scholar] [CrossRef]
- Jian, S.; Zhao, C.; Fang, S.; Yu, K. The Distribution of Fine Root Length Density for Six Artificial Afforestation Tree Species in Loess Plateau of Northwest China. For. Syst. 2015, 24, 3. [Google Scholar] [CrossRef] [Green Version]
- Boncina, A.; Kadunc, A.; Robic, D. Effects of Selective Thinning on Growth and Development of Beech (Fagus Sylvatica L.) Forest Stands in South-Eastern Slovenia. Ann. For. Sci. 2007, 64, 47–57. [Google Scholar] [CrossRef] [Green Version]
- Pajtík, J.; Konôpka, B.; Marušák, R. Above-Ground Net Primary Productivity in Young Stands of Beech and Spruce. Cent. Eur. For. J. 2013, 59, 154–162. [Google Scholar] [CrossRef] [Green Version]
- Tateno, R.; Hishi, T.; Takeda, H. Above-and Belowground Biomass and Net Primary Production in a Cool-Temperate Deciduous Forest in Relation to Topographical Changes in Soil Nitrogen. For. Ecol. Manag. 2004, 193, 297–306. [Google Scholar] [CrossRef]
- Finér, L.; Helmisaari, H.-S.; Lõhmus, K.; Majdi, H.; Brunner, I.; Børja, I.; Eldhuset, T.; Godbold, D.; Grebenc, T.; Konôpka, B. Variation in Fine Root Biomass of Three European Tree Species: Beech (Fagus Sylvatica L.), Norway Spruce (Picea Abies L. Karst.), and Scots Pine (Pinus Sylvestris L.). Plant Biosyst. 2007, 141, 394–405. [Google Scholar] [CrossRef]
- Neumann, M.; Godbold, D.L.; Hirano, Y.; Finér, L. Improving Models of Fine Root Carbon Stocks and Fluxes in European Forests. J. Ecol. 2020, 108, 496–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, P.; Kozlowski, T.T. Physiology of Woody Plants; Academic Press: Cambridge, MA, USA, 1979. [Google Scholar]
- Sagnard, F.; Pichot, C.; Dreyfus, P.; Jordano, P.; Fady, B. Modelling Seed Dispersal to Predict Seedling Recruitment: Recolonization Dynamics in a Plantation Forest. Ecol. Modell. 2007, 203, 464–474. [Google Scholar] [CrossRef]
- Wagner, S.; Collet, C.; Madsen, P.; Nakashizuka, T.; Nyland, R.D.; Sagheb-Talebi, K. Beech Regeneration Research: From Ecological to Silvicultural Aspects. For. Ecol. Manag. 2010, 259, 2172–2182. [Google Scholar] [CrossRef]
- Zhang, L.; Dong, L.; Liu, Q.; Liu, Z. Spatial Patterns and Interspecific Associations During Natural Regeneration in Three Types of Secondary Forest in the Central Part of the Greater Khingan Mountains, Heilongjiang Province, China. Forests 2020, 11, 152. [Google Scholar] [CrossRef] [Green Version]
- Feldmann, E.; Glatthorn, J.; Ammer, C.; Leuschner, C. Regeneration Dynamics Following the Formation of Understory Gaps in a Slovakian Beech Virgin Forest. Forests 2020, 11, 585. [Google Scholar] [CrossRef]
- Barna, M. The Effects of Cutting Regimes on Natural Regeneration in Submountain Beech Forests: Species Diversity and Abundance. J. For. Sci. 2008, 54, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Bakker, M.R.; Turpault, M.-P.; Huet, S.; Nys, C. Root Distribution of Fagus Sylvatica in a Chronosequence in Western France. J. For. Res. 2008, 13, 176–184. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plot | Silvicultural System | First Harvest (1989) | Second Harvest (2004) | Third Harvest (2009) | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Red. BA | BA | RD | Red. BA | BA | RD | Red. BA | BA | RD | ||
| NC | control plot (no cut) | 0 | 41 | 90 | 0 | 45 | 100 | 0 | 49 | 100 |
| LC | shelterwood (light cut) | 24 | 29 | 70 | 43 | 21 | 50 | 100 | 0 | 0 |
| MC | shelterwood (medium cut) | 44 | 19 | 50 | 53 | 13 | 30 | 100 | 0 | 0 |
| HC | shelterwood (heavy cut) | 68 | 14 | 30 | 100 | 0 | 0 | 0 | 0 | |
| CC | clear cut | 100 | 0 | 0 | 0 | 0 | 0 | |||
| Plot | Silvicultural System | Growth Stage | Density (Stems ha−1) | Height (m) | Beech | |
|---|---|---|---|---|---|---|
| Mean | 25–75% | (%) | ||||
| NC | control plot (no cut) | mature | 600 | 29.1 | 21.3–32.5 | 94.4 |
| seedling | 42,600 | 0.3 | 0.1–0.3 | 79.3 | ||
| LC | shelterwood (light cut) | thicket | 41,900 | 2.2 | 1.4–3.0 | 77.3 |
| MC | shelterwood (medium cut) | thicket | 58,100 | 2.9 | 1.0–3.9 | 87.7 |
| HC | shelterwood (heavy cut) | thicket | 22,200 | 5.1 | 3.0–7.2 | 60.7 |
| CC | clear cut | thicket | 11,100 | 7.5 | 4.0–10.3 | 40.3 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Konôpka, B.; Barna, M.; Bosela, M.; Lukac, M. Biomass Allocation to Resource Acquisition Compartments Is Affected by Tree Density Manipulation in European Beech after Three Decades. Forests 2020, 11, 940. https://0-doi-org.brum.beds.ac.uk/10.3390/f11090940
Konôpka B, Barna M, Bosela M, Lukac M. Biomass Allocation to Resource Acquisition Compartments Is Affected by Tree Density Manipulation in European Beech after Three Decades. Forests. 2020; 11(9):940. https://0-doi-org.brum.beds.ac.uk/10.3390/f11090940
Chicago/Turabian StyleKonôpka, Bohdan, Milan Barna, Michal Bosela, and Martin Lukac. 2020. "Biomass Allocation to Resource Acquisition Compartments Is Affected by Tree Density Manipulation in European Beech after Three Decades" Forests 11, no. 9: 940. https://0-doi-org.brum.beds.ac.uk/10.3390/f11090940