Assessment of SITE for CO2 and Energy Fluxes Simulations in a Seasonally Dry Tropical Forest (Caatinga Ecosystem)

 , ,
, ,  , ,
, ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Description of the Experimental Area
2.2. Micrometeorological Measurements
2.3. Data Processing and Post Processing
2.4. Energy Balance
2.5. Net Ecosystem Exchange
2.6. Description of SITE Model and Site Specific Biophysical Parameters
3. Results and Discussion
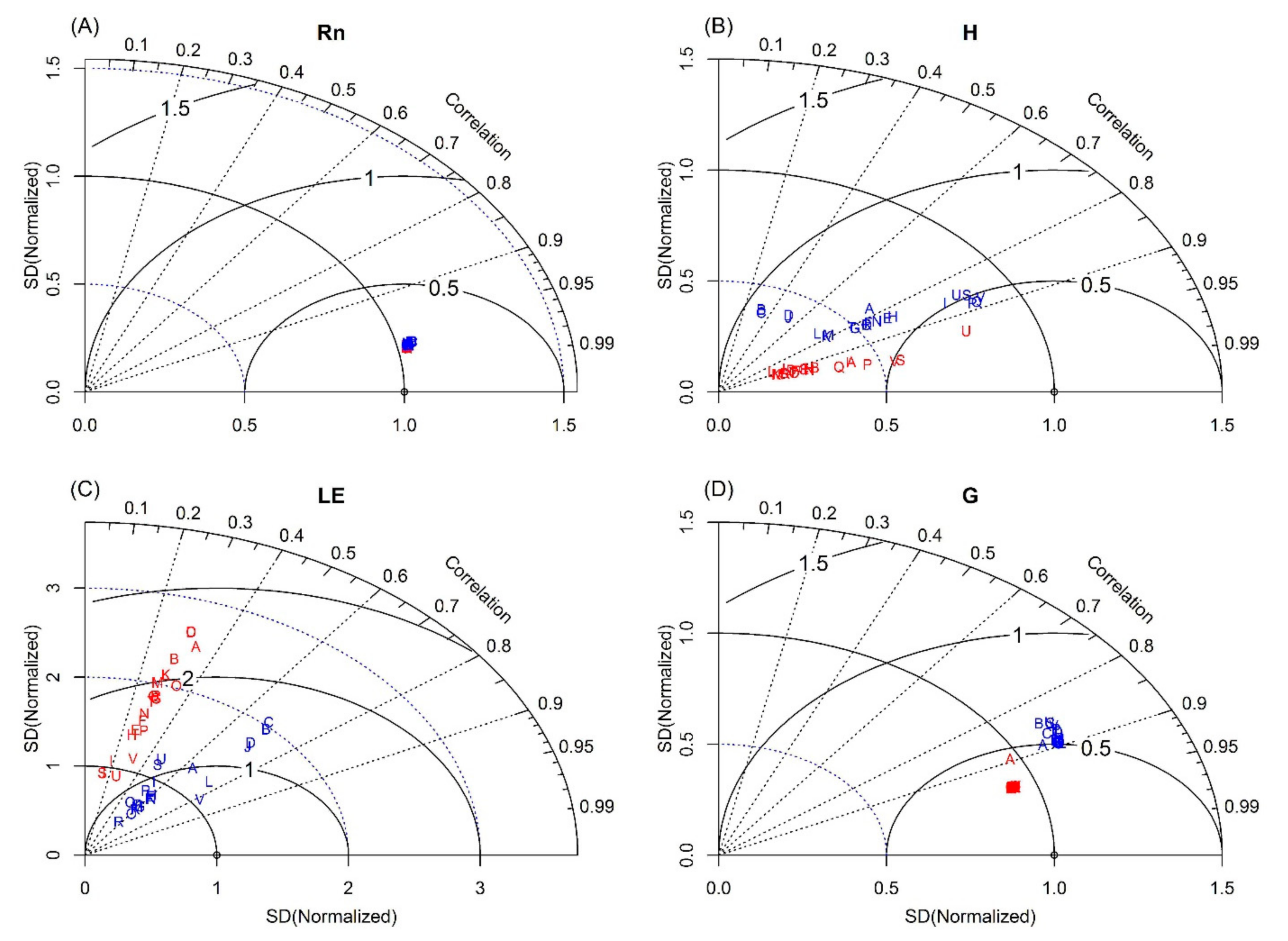
3.1. Calibration Test
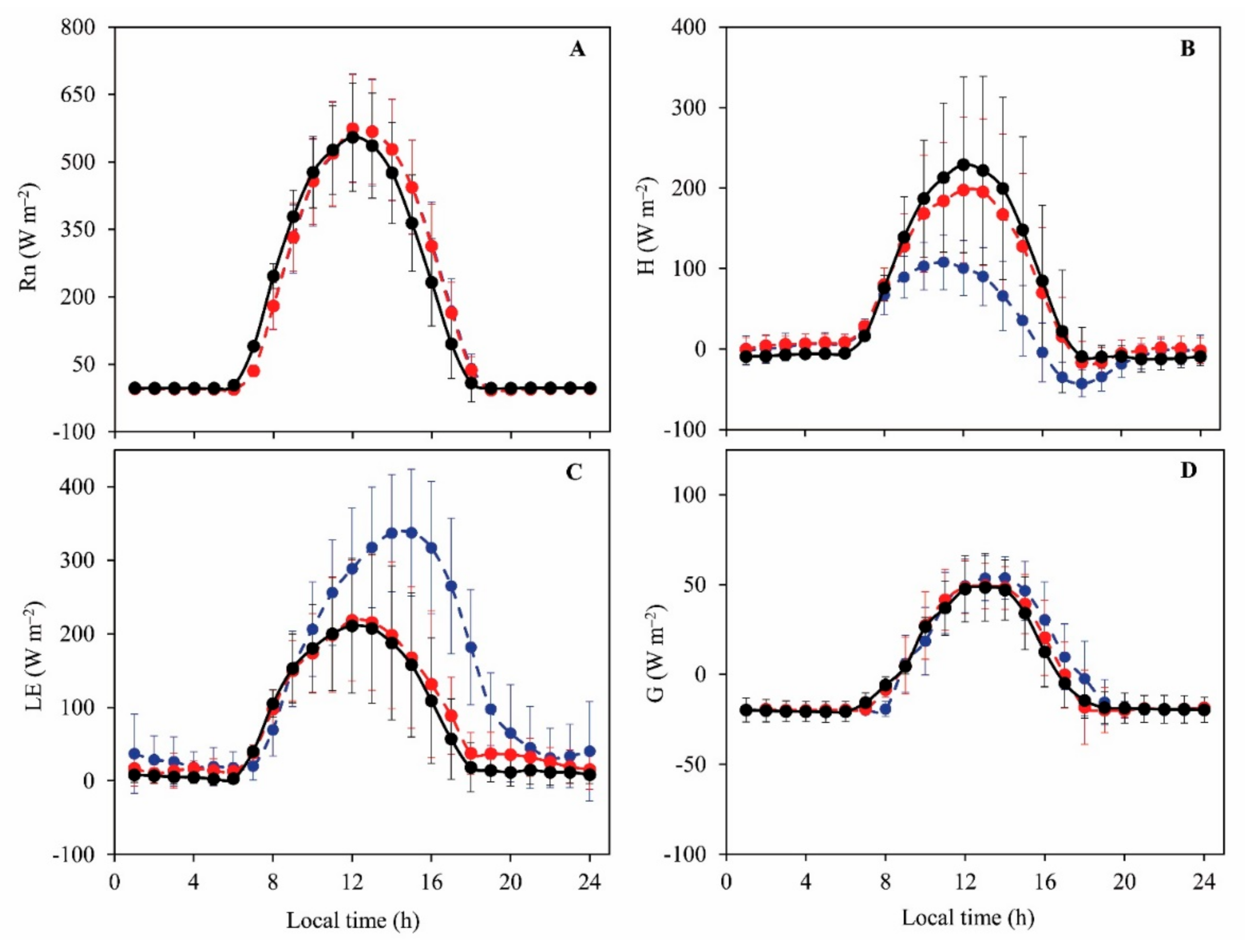
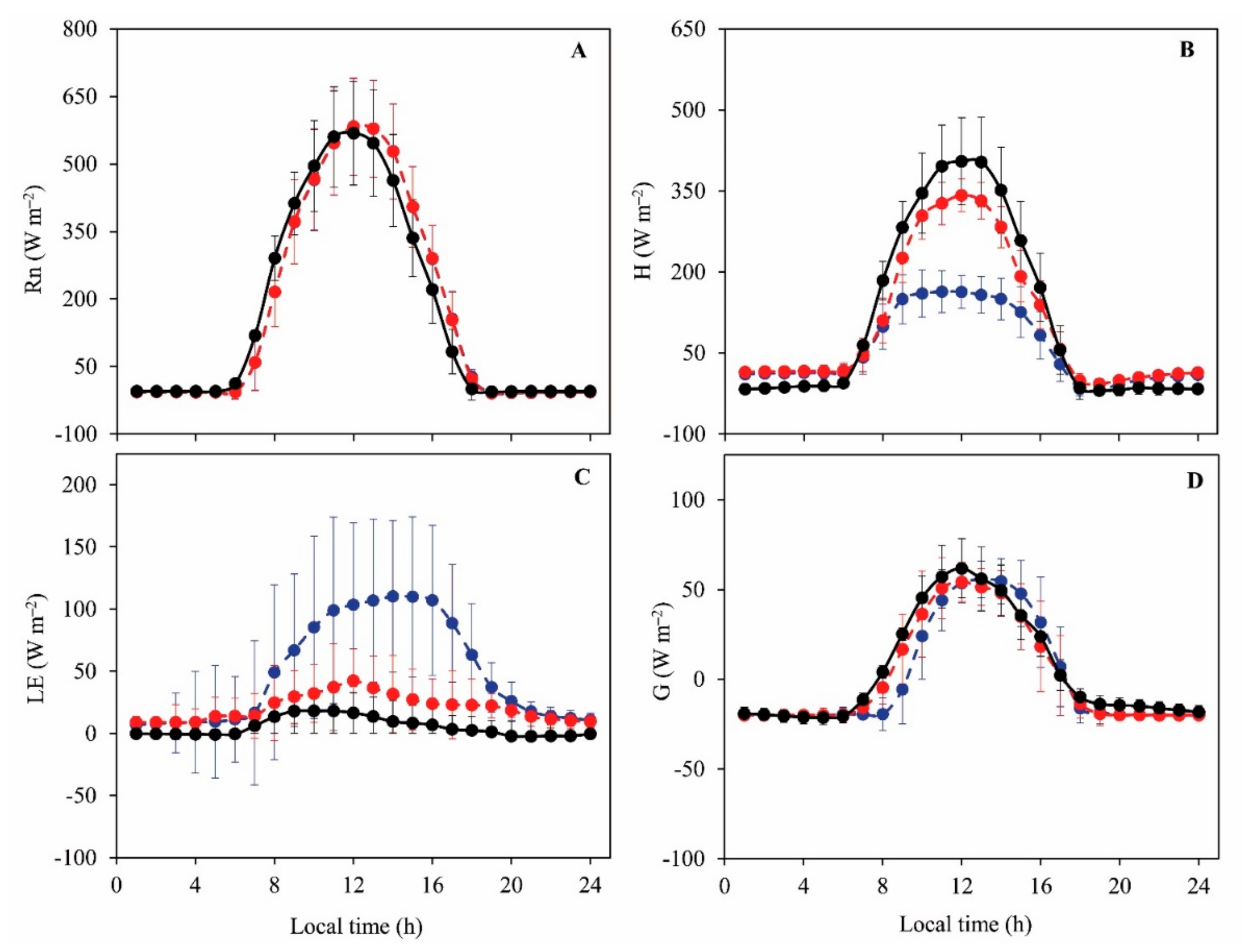
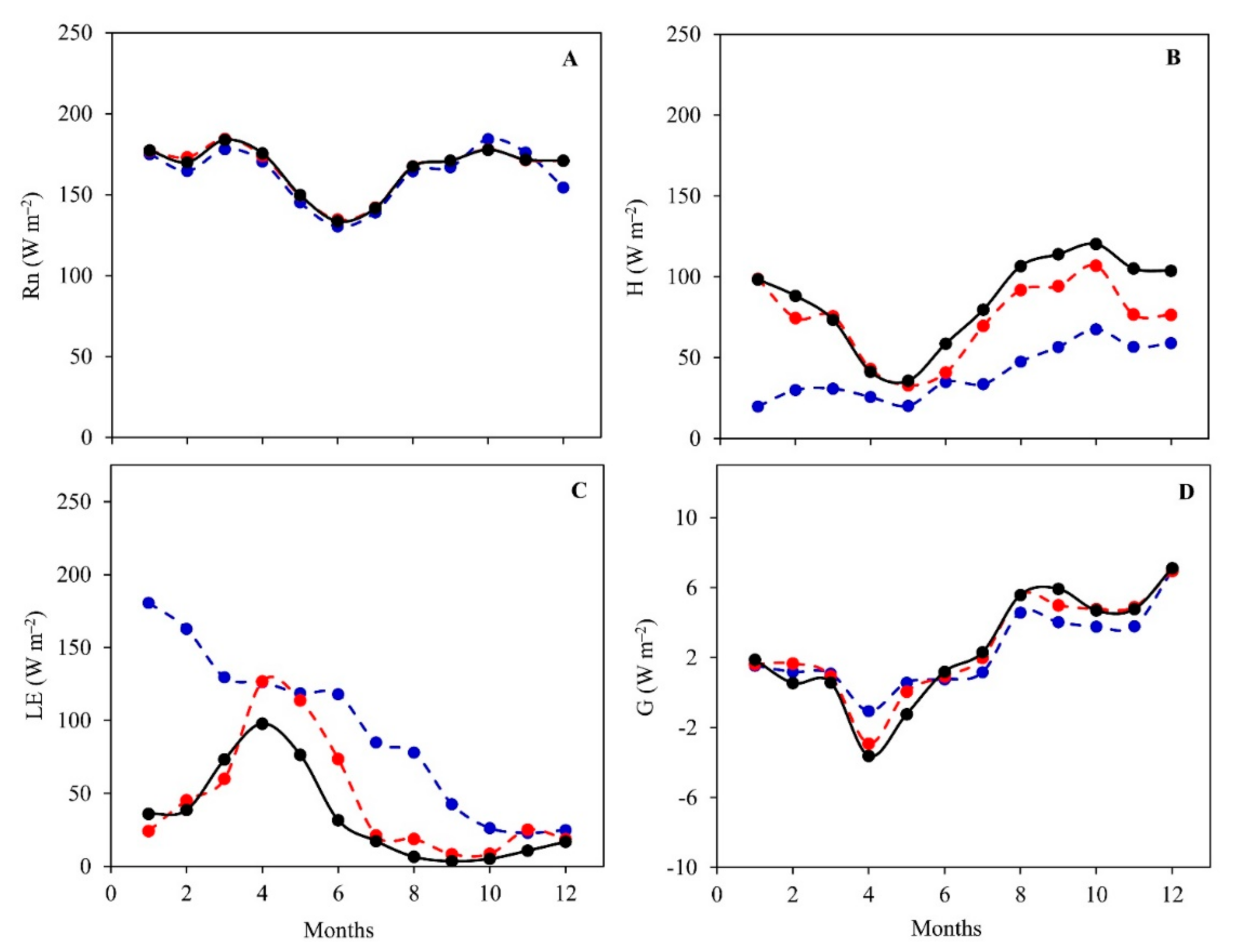
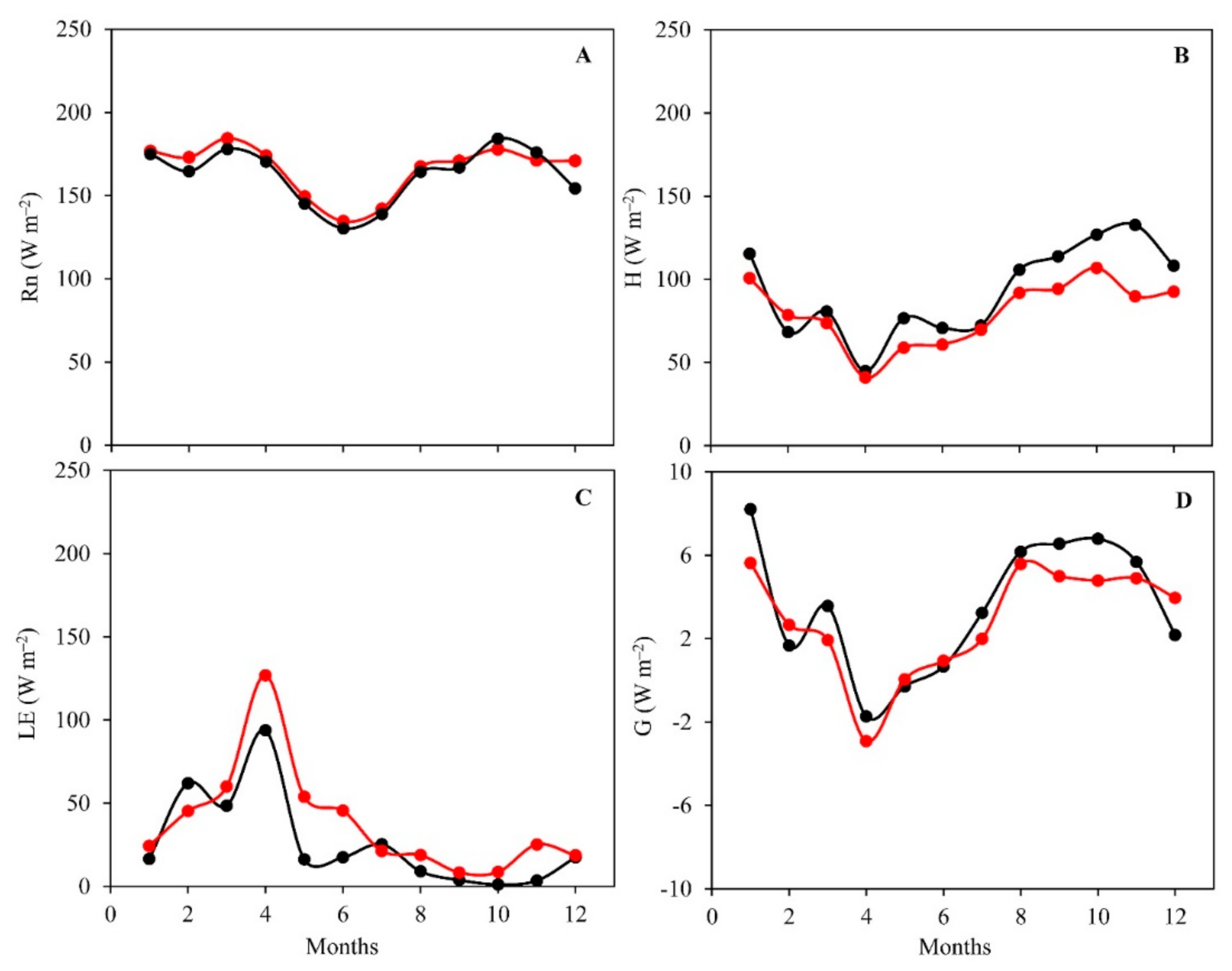
3.2. Daily Variations and Seasonal Dynamics of Simulated Energy Fluxes
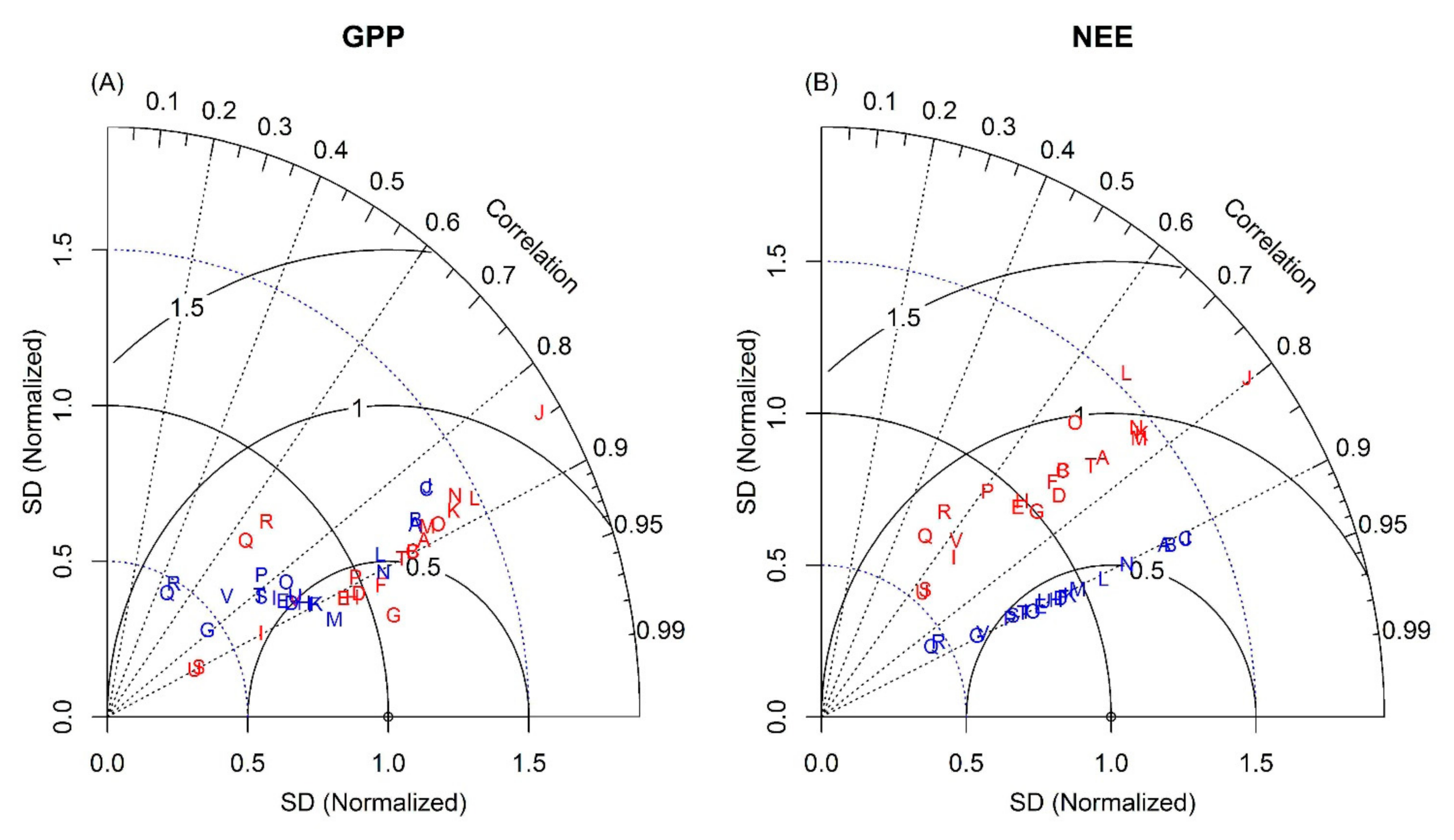
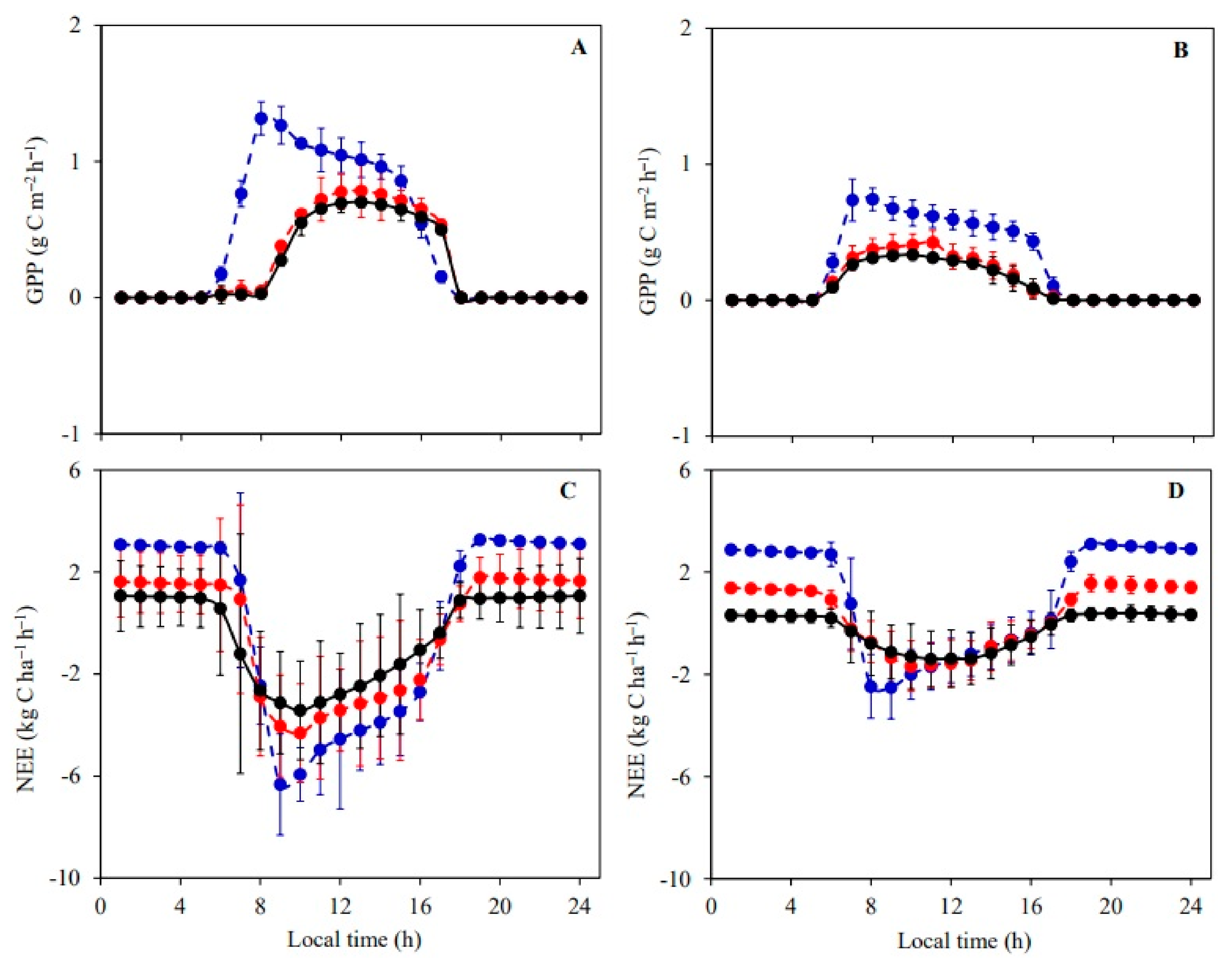
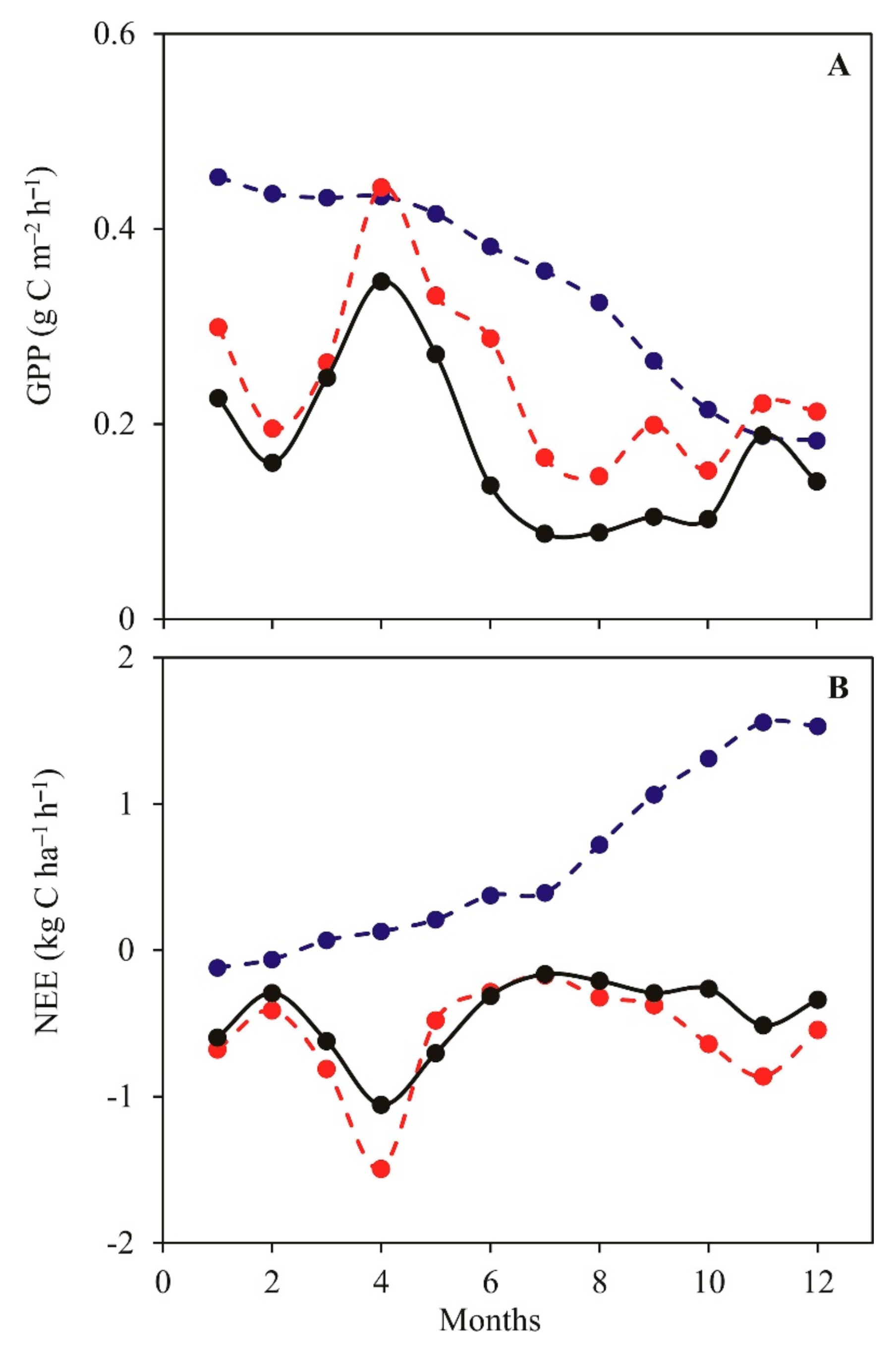
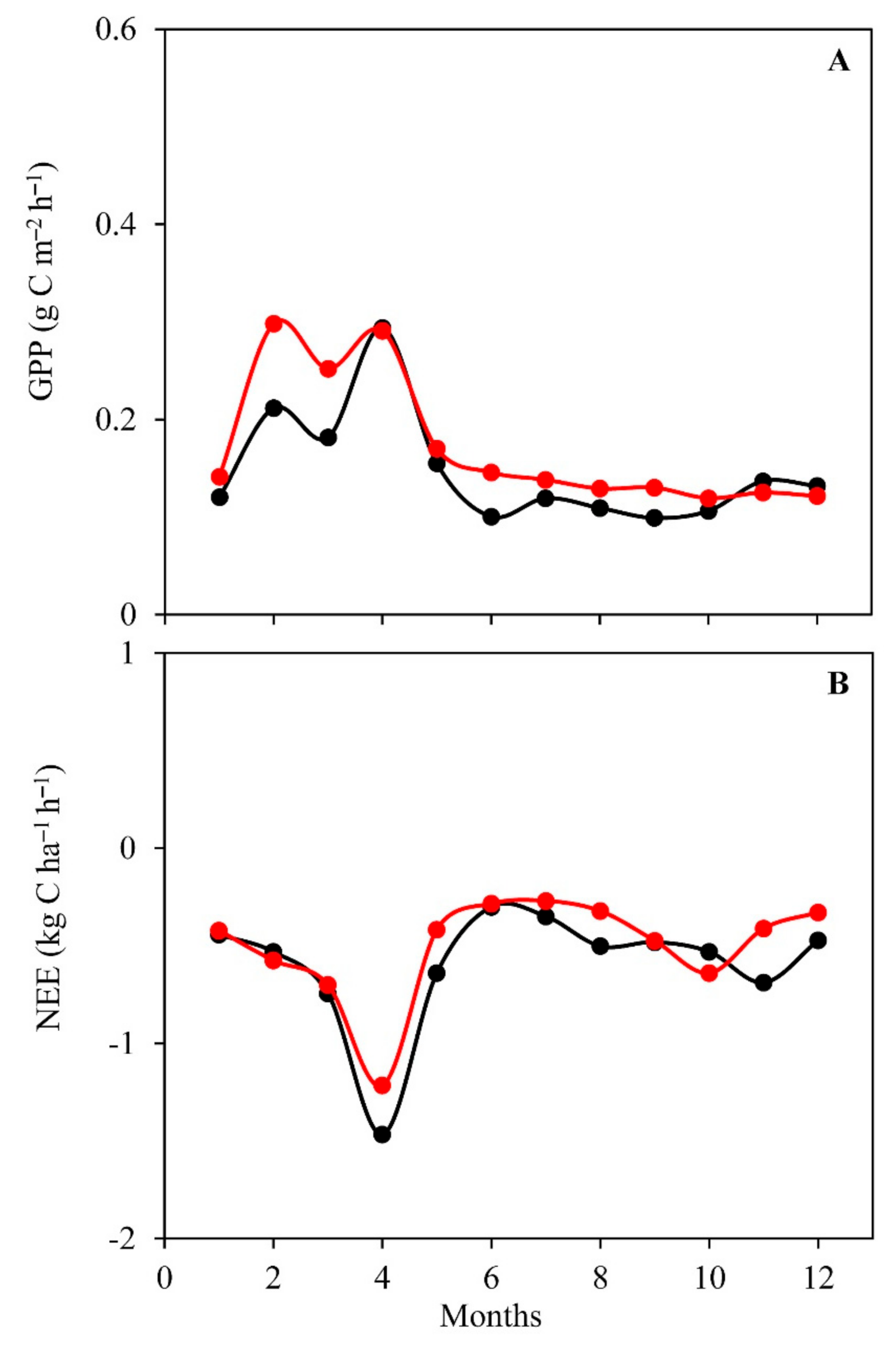
3.3. Daily Variations and Seasonal Dynamics of Simulated CO2 Fluxes
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Glotfelty, T.; Zhang, Y. Impact of future climate policy scenarios on air quality and aerosol–cloud interactions using an advanced version of CESM/CAM5: Part II. Future trend analysis and impacts of projected anthropogenic emissions. Atmos. Environ. 2017, 152, 531–552. [Google Scholar] [CrossRef] [Green Version]
- Parmesan, C.; Burrows, M.T.; Duarte, C.M.; Poloczanska, E.S.; Richardson, A.J.; Schoeman, D.S.; Singer, M.C. Beyond climate change attribution in conservation and ecological research. Ecol. Lett. 2013, 16, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Sá, J.C.D.M.; Lal, R.; Cerri, C.C.; Lorenz, K.; Hungria, M.; Carvalho, P.C.D.F. Low-carbon agriculture in South America to mitigate global climate change and advance food security. Environ. Int. 2017, 98, 102–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dombroski, J.L.D.; Praxedes, S.C.; Freitas, R.M.O.; Pontes, F.M. Water relations of Caatinga trees in the dry season. S. Afr. J. Bot. 2011, 77, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Santos, M.G.; Oliveira, M.T.; Figueiredo, K.V.; Falcão, H.M.; Arruda, E.C.P.; De Almeidacortez, J.S.; Sampaio, E.V.S.B.; Ometto, J.P.H.B.; Menezes, R.S.C.; Oliveira, A.F.M.; et al. Caatinga, the Brazilian dry tropical forest: Can it tolerate climate changes? Theor. Exp. Plant Physiol. 2014, 26, 83–99. [Google Scholar] [CrossRef]
- Koch, R.; Almeida–Cortez, J.S.; Kleinschmit, B. Revealing areas of high nature conservation importance in a seasonally dry tropical forest in Brazil: Combination of modelled plant diversity hot spots and threat patterns. J. Nat. Conserv. 2017, 35, 24–39. [Google Scholar] [CrossRef]
- Foley, J.A.; Prentice, I.C.; Ramankutty, N.; Levis, S.; Pollard, D.; Sitch, S.; Haxeltine, A. An integrated biosphere model of land surface processes, terrestrial carbon balance, and vegetation dynamics. Glob. Biogeochem. Cycles 1996, 10, 603–628. [Google Scholar] [CrossRef]
- Luyssaert, S.; Inglima, I.; Jung, M.; Richardson, A.D.; Reichstein, M.; Papale, D.; Piao, S.L.; Schulze, E.; Wingate, L.; Matteucci, G.; et al. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Glob. Chang. Biol. 2007, 13, 2509–2537. [Google Scholar] [CrossRef] [Green Version]
- Bremer, D.J.; Ham, J.M. Measurement and modeling of soil CO2 flux in a temperate grassland under mowed and burned regimes. Ecol. Appl. 2002, 12, 1318–1328. [Google Scholar]
- Hao, Y.; Wang, Y.; Huang, X.; Cui, X.; Zhou, X.; Wang, S.; Niu, H.; Jiang, G. Seasonal and interannual variation in water vapor and energy exchange over a typical steppe in Inner Mongolia, China. Agric. For. Meteorol. 2007, 146, 57–69. [Google Scholar] [CrossRef]
- Musavi, T.; Migliavacca, M.; Reichstein, M.; Kattge, J.; Wirth, C.; Black, T.A.; Janssens, I.; Knohl, A.; Loustau, D.; Roupsard, O.; et al. Stand age and species richness dampen interannual variation of ecosystem-level photosynthetic capacity. Nat. Ecol. Evol. 2017, 1, 0048. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land–use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Biudes, M.S.; Vourlitis, G.L.; Machado, N.G.; de Arruda, P.H.Z.; Neves, G.A.R.; de Almeida Lobo, F.; Neale, C.M.U.; de Souza Nogueira, J. Patterns of energy balance exchange for tropical ecosystems across a climate gradient in Mato Grosso, Brazil. Agric. For. Meterol. 2015, 202, 112–124. [Google Scholar] [CrossRef]
- Cabral, O.M.R.; Rocha, H.R.; Gash, J.H.; Freitas, H.C.; Ligo, M.A.V. Water and energy fluxes from woodland savanna (cerrado) in southeast Brazil. J. Hydrol. Reg. Stud. 2015, 4, 22–40. [Google Scholar] [CrossRef] [Green Version]
- Campos, S.; Mendes, K.R.; Da Silva, L.L.; Mutti, P.R.; Medeiros, S.S.; Amorim, L.B.; Dos Santos, C.A.; Perez-Marin, A.M.; Ramos, T.M.; Marques, T.V.; et al. Closure and partitioning of the energy balance in a preserved area of a Brazilian seasonally dry tropical forest. Agric. For. Meteorol. 2019, 271, 398–412. [Google Scholar] [CrossRef]
- Silva, P.F.; de Sousa Lima, J.R.; Antonino, A.C.D.; Souza, R.; de Souza, E.S.; Silva, J.R.I.; Alves, E.M. Seasonal patterns of carbon dioxide, water and energy fluxes over the Caatinga and grassland in the semi-arid region of Brazil. J. Arid. Environ. 2017, 147, 71–82. [Google Scholar] [CrossRef]
- Meir, P.; Metcalfe, D.B.; Costa, A.C.L.; Fisher, R.A. The fate of assimilated carbon during drought: Impacts on respiration in Amazon rainforests. Philos. Trans. R. Soc. B. 2008, 363, 1849–1855. [Google Scholar] [CrossRef]
- Sotta, E.D.; Veldkamp, E.; Schwendenmann, L.; Guimarães, B.R.; Paixão, R.K.; Ruivo, M.D.L.P.; Da Costa, A.C.L.; Meir, P. Effects of an induced drought on soil carbon dioxide (CO2) efflux and soil CO2 production in an Eastern Amazonian rainforest, Brazil. Glob. Chang. Biol. 2007, 13, 2218–2229. [Google Scholar] [CrossRef]
- Wu, J.; Guan, K.; Hayek, M.N.; Coupe, N.R.; Wiedemann, K.T.; Xu, X.; Wehr, R.; Christoffersen, B.O.; Miao, G.; Da Silva, R.; et al. Partitioning controls on Amazon forest photosynthesis between environmental and biotic factors at hourly to interannual timescales. Glob. Chang. Biol. 2016, 23, 1240–1257. [Google Scholar] [CrossRef]
- Zeri, M.; Sá, L.D.D.A.; Manzi, A.O.; Araújo, A.C.; Aguiar, R.G.; Von Randow, C.; Sampaio, G.; Cardoso, F.L.; Nobre, C.A. Variability of Carbon and Water Fluxes Following Climate Extremes over a Tropical Forest in Southwestern Amazonia. PLoS ONE 2014, 9, e88130. [Google Scholar] [CrossRef] [Green Version]
- Barbosa, H.A.; Kumar, T.V.L. Influence of rainfall variability on the vegetation dynamics over Northeastern Brazil. J. Arid Environ. 2016, 124, 377–387. [Google Scholar] [CrossRef]
- Mendes, K.R.; Campos, S.; Da Silva, L.L.; Mutti, P.R.; Ferreira, R.R.; Medeiros, S.S.; Perez-Marin, A.M.; Marques, T.V.; Ramos, T.M.; Vieira, M.M.D.L.; et al. Seasonal variation in net ecosystem CO2 exchange of a Brazilian seasonally dry tropical forest. Sci. Rep. 2020, 10, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Cunha, A.P.M.A.; Alvalá, R.C.; Kubota, P.Y.; Vieira, R.M. Impacts of land use and land cover changes on the climate over Northeast Brazil. Atmos. Sci. Lett. 2015, 16, 219–227. [Google Scholar] [CrossRef]
- De Souza, D.C.; Oyama, M.D. Climatic consequences of gradual desertification in the semi–arid area of Northeast Brazil. Theor. Appl. Climatol. 2011, 103, 345–357. [Google Scholar] [CrossRef]
- Marengo, J.A.; Ambrizzi, T.; Da Rocha, R.P.; Alves, L.M.; Cuadra, S.V.; Valverde, M.C.; Torres, R.R.; Santos, D.C.; Ferraz, S.E.T. Future change of climate in South America in the late twenty-first century: Intercomparison of scenarios from three regional climate models. Clim. Dyn. 2010, 35, 1073–1097. [Google Scholar] [CrossRef]
- Marengo, J.A.; Torres, R.R.; Alves, L.M. Drought in Northeast Brazil—Past, present, and future. Theor. Appl. Climatol. 2017, 129, 1189–1200. [Google Scholar] [CrossRef]
- Beer, C.; Reichstein, M.; Tomelleri, E.; Ciais, P.; Jung, M.; Carvalhais, N.; Rödenbeck, C.; Arain, M.A.; Baldocchi, D.; Bonan, G.B.; et al. Terrestrial gross carbon dioxide uptake: Global distribution and covariation with climate. Science 2010, 329, 834–838. [Google Scholar] [CrossRef] [Green Version]
- Cleverly, J.; Eamus, D.; Van Gorsel, E.; Chen, C.; Rumman, R.; Luo, Q.; Coupe, N.R.; Li, L.; Kljun, N.; Faux, R.; et al. Productivity and evapotranspiration of two contrasting semiarid ecosystems following the 2011 global carbon land sink anomaly. Agric. For. Meteorol. 2016, 220, 151–159. [Google Scholar] [CrossRef] [Green Version]
- Tang, X.; Carvalhais, N.; Moura, C.; Ahrens, B.; Koirala, S.; Fan, S.; Reichstein, M. Global variability of carbon use efficiency in terrestrial ecosystems. Biogeosci. Discuss. 2019. [Google Scholar] [CrossRef] [Green Version]
- Jaeger, E.B.; Stöckli, R.; Seneviratne, S.I. Analysis of planetary boundary layer fluxes and land–atmosphere coupling in the regional climate model CLM. J. Geophys. Res. 2009, 114, D17106. [Google Scholar] [CrossRef] [Green Version]
- Von Randow, C.; Zeri, M.; Coupe, N.R.; Muza, M.N.; De Goncalves, L.G.G.; Costa, M.H.; Araújo, A.C.; Manzi, A.O.; Da Rocha, H.R.; Saleska, S.R.; et al. Inter-annual variability of carbon and water fluxes in Amazonian forest, Cerrado and pasture sites, as simulated by terrestrial biosphere models. Agric. For. Meteorol. 2013, 182-183, 145–155. [Google Scholar] [CrossRef]
- Pires, W.N.; Moura, M.S.B.; Souza, L.S.B.; Silva, T.G.F.; Carvalho, H.F.S. Fluxos de radiação, energia, CO2 e vapor de água em uma área de caatinga em regeneração. Agrometoeros 2017, 25, 143–151. (In Portuguese) [Google Scholar]
- Souza, R.; Feng, X.; Antonino, A.; Montenegro, S.; Souza, E.; Porporato, A. Vegetation response to rainfall seasonality and interannual variability in tropical dry forests. Hydrol. Process. 2016, 30, 3583–3595. [Google Scholar] [CrossRef]
- Teixeira, A.H.C.; Bastiaanssen, W.G.M.; Ahmad, M.D.; Moura, M.S.B.; Bos, M.G. Analysis of energy fluxes and vegetation–atmosphere parameters in irrigated and natural ecosystems of semi–arid Brazil. J. Hydrol. 2008, 362, 110–127. [Google Scholar] [CrossRef] [Green Version]
- Santos, S.N.M.; Costa, M.H. A simple tropical ecosystem model of carbon, water and energy fluxes. Ecol. Model. 2004, 176, 291–312. [Google Scholar] [CrossRef]
- Tavares-Dasmasceno, J.P.; de Souza Silveira, J.L.G.; Câmara, T.; de Castro Stedile, P.; Macario, P.; Toledo-Lima, G.S.; Pichorim, M. Effect of drought on demography of Pileated Finch (Coryphospingus pileatus: Thraupidae) in northeastern Brazil. J. Arid Environ 2017, 147, 63–79. [Google Scholar] [CrossRef]
- Oliveira, P.T.; Santos e Silva, C.M.; Lima, K.C. Climatology and trend analysis of extreme precipitation in subregions of Northeast Brazil. Theor. Appl. Climatol. 2017, 130, 77–90. [Google Scholar] [CrossRef]
- Pagoto, M.A.; Roig, F.A.; Ribeiro, A.S.; Lisi, C.S. Influence of regional rainfall and Atlantic sea surface temperature on tree–ring growth of Poincianella pyramidalis, semiarid forest from Brazil. Dendrochronologia 2015, 35, 14–23. [Google Scholar] [CrossRef]
- Costa, C.A.G.; Lopes, J.W.B.; Pinheiro, E.A.R.; Araújo, J.C.; Gomes–Filho, R.R. Spatial behaviour of soil moisture in the root zone of the Caatinga biome. Rev. Ciênc. Agron. 2013, 44, 685–694. [Google Scholar] [CrossRef] [Green Version]
- Webb, E.K.; Pearman, G.I.; Leuning, R. Correction of flux measurements for density effects due to heat and water vapour transfer. Q. J. R. Meteorol. Soc. 1980, 106, 85–100. [Google Scholar] [CrossRef]
- Moore, C.J. Frequency response corrections for eddy correlation systems. Bound.–Layer Meteorol. 1986, 37, 17–35. [Google Scholar] [CrossRef]
- Massman, W.J. A simple method for estimating frequency response corrections for eddy covariance systems. Agric. For. Meteorol. 2000, 104, 185–198. [Google Scholar] [CrossRef]
- Papale, D.; Reichstein, M.; Aubinet, M.; Canfora, E.; Bernhofer, C.; Kutsch, W.; Longdoz, B.; Rambal, S.; Valentini, R.; Vesala, T.; et al. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: Algorithms and uncertainty estimation. Biogeosciences 2006, 3, 571–583. [Google Scholar] [CrossRef] [Green Version]
- Reichstein, M.; Falge, E.; Baldocchi, D.; Papale, D.; Aubinet, M.; Berbigier, P.; Bernhofer, C.; Buchmann, N.; Gilmanov, T.; Granier, A.; et al. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: Review and improved algorithm. Glob. Chang. Biol. 2005, 11, 1424–1439. [Google Scholar] [CrossRef]
- Jensen, R.; Herbst, M.; Fribog, T. Direct and indirect controls of the interanual variability in atmospheric CO2 exchange of three contrasting ecosystems in Denmark. Agric. For. Meteorol. 2017, 269–270, 136–144. [Google Scholar]
- Aubinet, M.; Chermanne, B.; Vendenhaute, M.; Longdoz, B.; Yernaux, M.; Laitat, E. Long term carbon dioxide exchange above a mixed forest in the Belgian Ardennes. Agric. For. Meteorol. 2001, 108, 293–315. [Google Scholar] [CrossRef]
- De Araújo, A.; Dolman, A.; Waterloo, M.; Gash, J.; Kruijt, B.; Zanchi, F.; De Lange, J.; Stoevelaar, R.; Manzi, A.; Nobre, A. The spatial variability of CO2 storage and the interpretation of eddy covariance fluxes in central Amazonia. Agric. For. Meteorol. 2010, 150, 226–237. [Google Scholar] [CrossRef]
- Lloyd, J.; Taylor, J.A. On the temperature dependence of soil respiration. Funct. Ecol. 1994, 8, 315–323. [Google Scholar] [CrossRef]
- Max Plank Institute for Biogeochemistry. Available online: http://www.bgc–jena.mpg.de/~MDIwork/eddyproc/ (accessed on 15 November 2018).
- LBA–ECO LC–31 Simple Tropical Ecosystem Model. Available online: https://daac.ornl.gov/LBA/guides/LC31_SITE.html (accessed on 10 June 2018).
- Cosby, B.J.; Hornberger, G.M.; Clapp, R.B.; Ginn, T.R. A statistical exploration of the relationships of soil moisture characteristics to the physical properties of soils. Water Resour. Res. 1984, 20, 682–690. [Google Scholar] [CrossRef] [Green Version]
- Costa, M.H. Estado-da-arte da simulação da taxa de fixação de carbono de ecossistemas tropicais. Rev. Bras. Meteorol. 2009, 24, 179–187. [Google Scholar] [CrossRef]
- Sanches, L.; Andrade, N.L.R.A.; Costa, M.H.; Alves, M.C.A.; Gaio, D. Performance evaluation of the SITE® model to estimate energy flux in a tropical semi–deciduous forest of the southern Amazon Basin. Int. J. Biometeorol. 2011, 55, 303–312. [Google Scholar] [CrossRef] [PubMed]
- Powell, T.L.; Galbraith, D.R.; Christoffersen, B.O.; Harper, A.; Imbuzeiro, H.M.; Rowland, L.; Almeida, S.; Brando, P.B.; da Costa, A.C.L.; Costa, M.H.; et al. Confronting model predictions of carbon fluxes with measurements of Amazon forests subjected to experimental drought. New Phytol. 2013, 200, 350–364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sellers, P.J.; Shuttleworth, W.J.; Dorman, J. Calibrating the Simple Biosphere Model for Amazonian Tropical Forest using field and remote sensing data. Part I: Average calibration with field data. J. Appl. Meteorol. 1989, 28, 727–759. [Google Scholar] [CrossRef] [Green Version]
- Stein, U.; Alpert, P. Factor separation in numerical simulations. J. Atmos. Sci. 1993, 50, 2107–2115. [Google Scholar] [CrossRef] [Green Version]
- Taylor, K.E. Summarizing multiple aspects of model performance in a single diagram. J. Geophys. Res. 2001, 106, 7183–7192. [Google Scholar] [CrossRef]
- Lemon, J. Plotrix: A package in the red light district of R. R–News 2006, 6, 8–12. [Google Scholar]
- Zanella De Arruda, P.H.; Vourlitis, G.L.; Santanna, F.B.; Pinto, O.B., Jr.; De Almeida Lobo, F.; De Souza Nogueira, J. Large net CO2 loss from a grass-dominated tropical savanna in south-central Brazil in response to seasonal and interannual drought. J. Geophys. Res. Biogeosci. 2016, 121, 2110–2124. [Google Scholar] [CrossRef]
- Ma, X.; Huete, A.; Cleverly, J.; Eamus, J.C.D.; Chevallier, F.; Joiner, J.; Poulter, B.; Zhang, Y.; Guanter, L.; Meyer, W.; et al. Drought rapidly diminishes the large net CO2 uptake in 2011 over semi-arid Australia. Sci. Rep. 2016, 6, 37747. [Google Scholar] [CrossRef]
- Mendes, K.R.; Granja, J.A.A.; Ometto, J.P.; Antonino, A.C.D.; Menezes, R.S.C.; Pereira, E.C.; Pompelli, M.F. Croton blanchetianus modulates its morphophysiological responses to tolerate drought in a tropical dry forest. Funct. Plant Biol. 2017, 10, 1–13. [Google Scholar] [CrossRef]
- Pinho-Pessoa, A.C.B. Interannual Variation in Temperature and Rainfall can Modulate the Physiological and Photoprotective Mechanisms of a Native Semiarid Plant Species. Indian J. Sci. Technol. 2018, 11, 1–17. [Google Scholar] [CrossRef]
- Lima–Silva, P.S.L.; Cunha, T.M.S.; Souza, A.D.; de Paula, V.F.S. Equations for leaf area estimation in some species adapted to the Brazilian Semi-arid. Rev. Caatinga 2007, 20, 18–23. [Google Scholar]
- Rezende, L.F.C.; Arenque-Musa, B.C.; Moura, M.S.B.; Aidar, S.T.; Von Randow, C.; Menezes, R.S.C.; Ometto, J.P.B.H. Calibration of the maximum carboxylation velocity (Vcmax) using data mining techniques and ecophysiological data from the Brazilian semiarid region, for use in Dynamic Global Vegetation Models. Braz. J. Biol. 2016, 76, 341–351. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, E.A.R.; Metselaar, K.; Van Lier, Q.J.; Araújo, J.C. Importance of soil–water to the Caatinga biome, Brazil. Ecohydrology 2016, 9, 1313–1327. [Google Scholar] [CrossRef]
- Pinheiro, E.A.R.; Van Lier, Q.J.; Bezerra, A.H.F. Hydrology of a Water–Limited Forest under Climate Change Scenarios: The Case of the Caatinga Biome, Brazil. Forests 2017, 8, 62. [Google Scholar] [CrossRef] [Green Version]
- Pilotto, I.L.; Rodríguez, D.A.; Tomasella, J.; Sampaio, G.; Chou, S.C. Comparisons of the Noah–MP land surface model simulations with measurements of forest and crop sites in Amazonia. Meteorol. Atmos. Phys. 2015, 127, 711–723. [Google Scholar] [CrossRef]
- Cunha, A.P.M.A.; Alvalá, R.C.S.; Sampaio, G.; Shimizu, M.H.; Costa, M.H. Calibration and Validation of the Integrated Biosphere Simulator (IBIS) for a Brazilian Semiarid Region. J. Appl. Meteorol. Clim. 2013, 52, 2753–2770. [Google Scholar] [CrossRef]
- Colello, G.D.; Grivet, C.; Sellers, P.J.; Berry, J.A. Modeling of energy, water, and CO2 flux in a temperate grassland ecosystem with SiB2: May–October 1987. Am. Meteorol. Soc. 1998, 55, 1141–1169. [Google Scholar]
- Rodrigues, T.R.; Vourlitis, G.L.; Lobo, F.A.; Oliveira, R.G.; Nogueira, J.S. Seasonal variation in energy balance and canopy conductance for a tropical savanna ecosystem of south-central Mato Grosso, Brazil. J. Geophys. Res. Biogeosci. 2014, 119, 1–13. [Google Scholar] [CrossRef]
- Falcão, H.M.; Medeiros, C.D.; Silva, B.L.; Sampaio, E.V.; Almeida-Cortez, J.; Santos, M.G. Phenotypic plasticity and ecophysiological strategies in a tropical dry forest chronosequence: A study case with Poincianella pyramidalis. For. Ecol. Manag. 2015, 340, 62–69. [Google Scholar] [CrossRef]
- Ziehn, T.; Kattge, J.; Knorr, W.; Scholze, M. Improving the predictability of global CO2 assimilation rates under climate change. Geophys. Res. Lett. 2011, 38, L10404. [Google Scholar] [CrossRef]
- Friend, A.D. Terrestrial plant production and climate change. J. Exp. Bot. 2010, 61, 1293–1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Best, M.J.; Pryor, M.; Clark, D.B.; Rooney, G.G.; Essery, R.; Menard, C.B.; Edwards, J.M.; Hendry, M.A.; Porson, A.; Gedney, N.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 1: Energy and water fluxes. Geosci. Model Dev. 2011, 4, 677–699. [Google Scholar] [CrossRef] [Green Version]
- Clark, D.B.; Mercado, L.M.; Sitch, S.; Jones, C.D.; Gedney, N.; Best, M.J.; Pryor, M.J.; Rooney, G.G.; Essery, R.L.H.; Blyth, E.M.; et al. The Joint UK Land Environment Simulator (JULES), model description—Part 2: Carbon fluxes and vegetation dynamics. Geosci. Model Dev. 2011, 4, 701–722. [Google Scholar] [CrossRef] [Green Version]
- Oleson, K.W.; Lawrence, D.M. Technical Description of Version 4.5 of the Community LandModel (CLM), NCAR Earth System Laboratory—Climate and Global Dynamics Division; Tech. Rep.TN-503+STR; National Center For Atmospheric Research: Boulder, CO, USA, 2013; Available online: http://www.cesm.ucar.edu/models/cesm1.2/clm/CLM45_Tech_Note.pdf (accessed on 1 August 2020).
- Luo, Y.; Medlyn, B.; Hui, D.; Ellsworth, D.; Reynolds, J.; Katul, G. Gross primary productivity in Duke forest: Modeling synthesis of CO2 experiment and eddy–flux data. Ecol. Appl. 2001, 11, 239–252. [Google Scholar]
- Zhan, X.; Xue, Y.; Collatz, G.J. An analytical approach for estimating CO2 and heat fluxes over the Amazonian region. Ecol. Model. 2003, 162, 97–117. [Google Scholar] [CrossRef] [Green Version]
- Antunes, W.C.; Mendes, K.R.; Chaves, A.R.D.M.; Ometto, J.P.; Jarma-Orozco, A.; Pompelli, M.F. Spondias tuberosa trees grown in tropical, wet environments are more susceptible to drought than those grown in arid environments. Rev. Colomb. Ciencia. Hortíc. 2016, 10, 9–27. [Google Scholar] [CrossRef]
- Mendes, K.R.; Marenco, R.A. Is stomatal conductance of Central Amazonian saplings influenced by circadian rhythms under natural conditions? Theor. Exp. Plant Physiol. 2014, 26, 115–125. [Google Scholar] [CrossRef]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [Green Version]
- Flexas, J.; Carriquí, M.; Coopman, R.E.; Gago, J.; Galmés, J.; Martorell, S.; Morales, F.; Diaz-Espejo, A. Stomatal and mesophyll conductances to CO2 in different plant groups: Underrated factors for predicting leaf photosynthesis responses to climate change? Plant Sci. 2014, 226, 41–48. [Google Scholar] [CrossRef]
- Delpierre, N.; Vitasse, Y.; Chuine, I.; Guillemot, J.; Bazot, S.; Rutishauser, T.; Rathgeber, C.B. Temperate and boreal forest tree phenology: From organ-scale processes to terrestrial ecosystem models. Ann. For. Sci. 2016, 73, 5–25. [Google Scholar] [CrossRef] [Green Version]
- Manoli, G.; Ivanov, V.Y.; Fatichi, S. Dry–Season Greening and Water Stress in Amazonia: The Role of Modeling Leaf Phenology. J. Geophys. Res. Biogeosci. 2018, 123, 1909–1926. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Used Value | Source |
|---|---|---|
| Height of data measurement (z) | 11 m | Measured on site |
| Height of the canopy (z1) | 8 m | Measured on site |
| Height of lower canopy (z2) | 5 m | Measured on site |
| Zero plane displacement (d) | 7.33 m | Estimated |
| Roughness above the canopy (zh) | 1.35 m | Estimated |
| Total soil porosity (Φ) | 0.41 m3 m−3 | [38,39] |
| Humidity content at field capacity (θCC) | 0.225 m3 m−3 | [38,39] |
| Moisture content of the permanent wilting point (θPM) | 0.151 m3 m−3 | [38,39] |
| Parameter | Wet Season Simulation V | Dry Season Simulation U | Wet Season Simulation M | Dry Season Simulation G | Source | |
|---|---|---|---|---|---|---|
| Initial | Energy Flux Calibrated | CO2 Flux Calibrated | ||||
| Specific leaf area (sla) | 13.0 | 14.5 | 23.5 | 14.5 | 23.5 | [61,62] |
| Typical dimension of leaves (du) | 0.072 | 0.056 | 0.032 | 0.056 | 0.032 | [63] |
| Typical dimension of stems (ds) | 0.1 | 0.05 | 0.05 | 0.05 | 0.05 | [63] |
| Leaf width (w) | 0.1 | 0.06 | 0.03 | 0.06 | 0.03 | [63] |
| Coefficient of stomatal conductance (m) | 10.0 | 8.0 | 5.0 | 8.0 | 5.0 | [4,61,62] |
| Maximum capacity of the Rubisco enzyme (Vmax) | 75 × 10−6 | 90 × 10−6 | 90 × 10−6 | 90 × 10−6 | 60 × 10−6 | [62,64] |
| Initial fraction of soil moisture (θg/θd) | 0.36 | 0.165 | 0.075 | 0.225 | 0.165 | [38,39] |
| Statistic | ||||||||
|---|---|---|---|---|---|---|---|---|
| 2014 | 2015 | |||||||
| r | MAE | RMSE | d | r | MAE | RMSE | d | |
| Energy flux | ||||||||
| Rn (W m−2) | 0.98 | 43.84 | 80.46 | 0.94 | 0.96 | 31.70 | 50.56 | 0.98 |
| H (W m−2) | 0.85 | 50.52 | 68.83 | 0.89 | 0.91 | 46.94 | 71.97 | 0.89 |
| LE (W m−2) | 0.69 | 29.97 | 65.99 | 0.72 | 0.71 | 24.25 | 53.48 | 0.74 |
| G (W m−2) | 0.90 | 9.90 | 13.66 | 0.92 | 0.90 | 11.07 | 16.23 | 0.91 |
| CO2 flux | ||||||||
| GPP (g C m−2 h−1) | 0.82 | 1.24 | 1.53 | 0.86 | 0.91 | 1.38 | 2.05 | 0.79 |
| NEE (kg C m−2 h−1) | 0.84 | 1.99 | 2.25 | 0.83 | 0.81 | 1.74 | 2.00 | 0.80 |
| Variable | 2014 | 2015 | ||
|---|---|---|---|---|
| Wet | Dry | Wet | Dry | |
| Energy flux | ||||
| Rn (W m−2) | 164.6(172.5) | 168.7(167.4) | 162.6(173.5) | 174.6(170.9) |
| H (W m−2) | 59.7(58.8) | 113.7(91.9) | 64.6(67.8) | 120.0(94.4) |
| LE (W m−2) | 71.5(81.8) | 5.2(18.6) | 48.1(56.9) | 4.4(17.5) |
| G (W m−2) | −0.8(0.4) | 5.4(4.9) | 2.3(2.8) | 6.5(4.9) |
| CO2 flux | ||||
| GPP (g C m−2 h−1) | 0.26(0.29) | 0.11(0.17) | 0.20(0.25) | 0.10(0.12) |
| NEE (kg C ha−1 h−1) | −0.66(−0.64) | −0.25(−0.34) | −0.69(−0.63) | −0.49(−0.40) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mendes, K.R.; Campos, S.; Mutti, P.R.; Ferreira, R.R.; Ramos, T.M.; Marques, T.V.; dos Reis, J.S.; de Lima Vieira, M.M.; Silva, A.C.N.; Marques, A.M.S.; et al. Assessment of SITE for CO2 and Energy Fluxes Simulations in a Seasonally Dry Tropical Forest (Caatinga Ecosystem). Forests 2021, 12, 86. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010086
Mendes KR, Campos S, Mutti PR, Ferreira RR, Ramos TM, Marques TV, dos Reis JS, de Lima Vieira MM, Silva ACN, Marques AMS, et al. Assessment of SITE for CO2 and Energy Fluxes Simulations in a Seasonally Dry Tropical Forest (Caatinga Ecosystem). Forests. 2021; 12(1):86. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010086
Chicago/Turabian StyleMendes, Keila R., Suany Campos, Pedro R. Mutti, Rosaria R. Ferreira, Tarsila M. Ramos, Thiago V. Marques, Jean S. dos Reis, Mariana M. de Lima Vieira, Any Caroline N. Silva, Ana Maria S. Marques, and et al. 2021. "Assessment of SITE for CO2 and Energy Fluxes Simulations in a Seasonally Dry Tropical Forest (Caatinga Ecosystem)" Forests 12, no. 1: 86. https://0-doi-org.brum.beds.ac.uk/10.3390/f12010086