
Thinning Effect of C Sequestration along an Elevation Gradient of Mediterranean Pinus spp. Plantations

 ,
,  , and
, and
Abstract
:1. Introduction
2. Materials and Methods
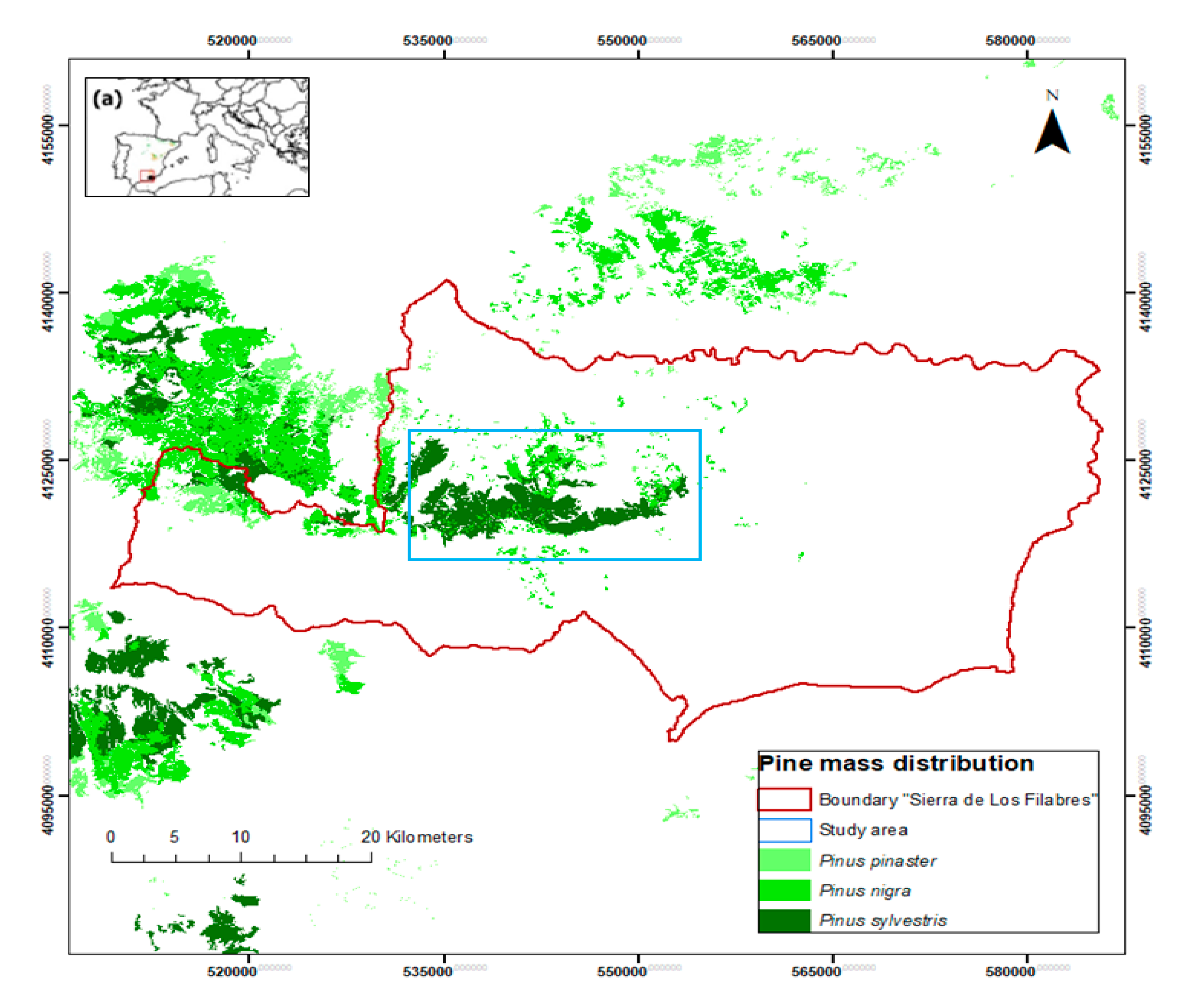
2.1. Study Area
2.2. Experimental Design and Field Data
2.3. Soil Sampling and Analysis
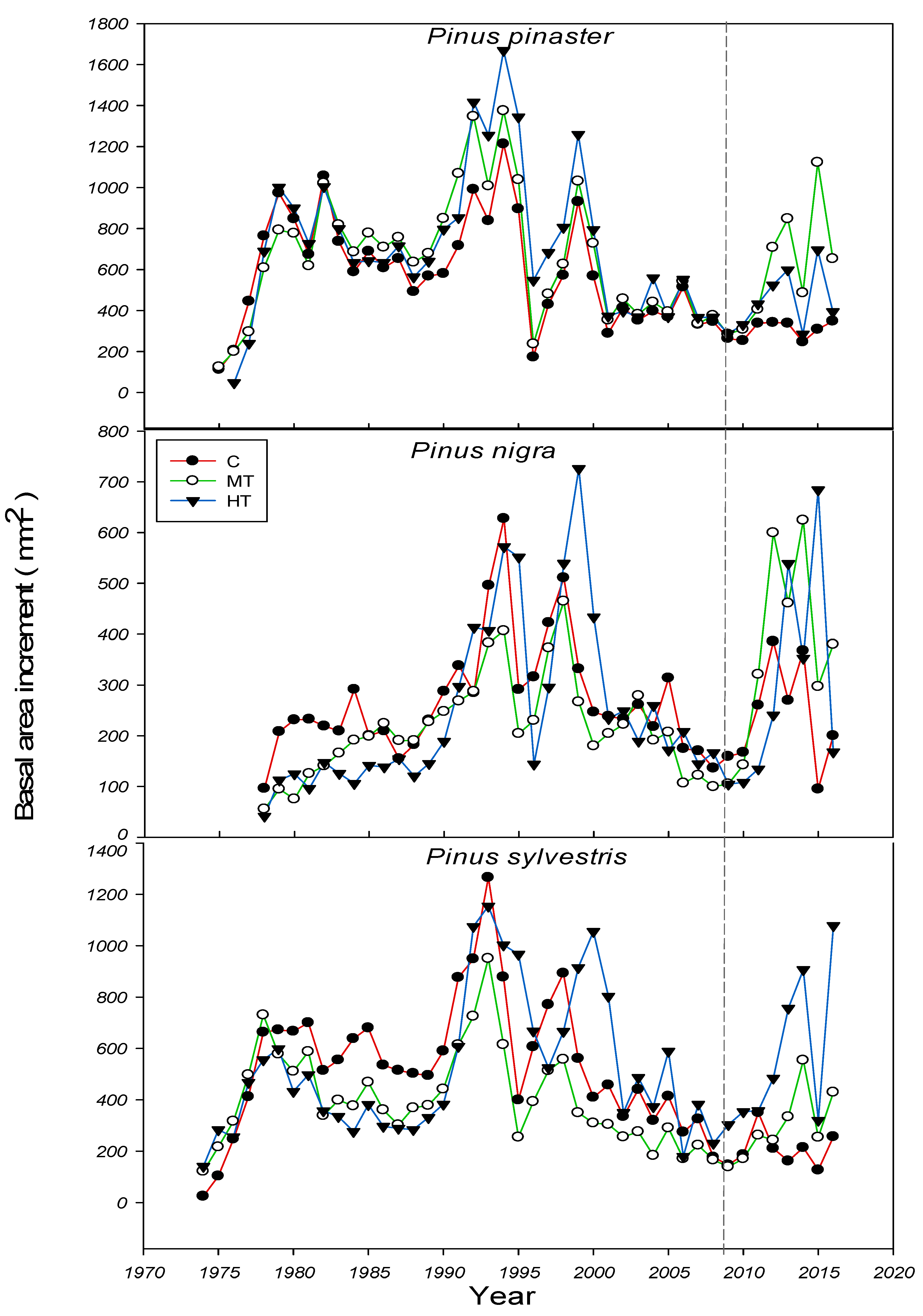
2.4. Dendrochronological Sampling and Analysis
2.5. Statistical Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Lal, R. Soil carbon sequestration to mitigate climate change. Geoderma 2004, 123, 1–22. [Google Scholar] [CrossRef]
- McCarthy, J.J.; Canziani, O.F.; Leary, N.A.; Dokken, D.J.; White, K.S. (Eds.) Climate Change 2001: Impacts, Adaptation, and Vulnerability: Contribution of Working Group II to the Third Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2001. [Google Scholar]
- Jandl, R.; Lindner, M.; Vesterdal, L.; Bauwens, B.; Baritz, R.; Hagedorn, F.; Johnson, D.W.; Minkkinen, K.; Byrne, K.A. How strongly can forest management influence soil carbon sequestration? Geoderma 2007, 137, 253–268. [Google Scholar] [CrossRef]
- Ceccherini, G.; Duveiller, G.; Grassi, G.; Lemoine, G.; Avitabile, V.; Pilli, R.; Cescatti, A. Abrupt increase in harvested forest area over Europe after 2015. Nature 2020, 583, 72–77. [Google Scholar] [CrossRef] [PubMed]
- Spake, R.; Yanou, S.; Yamaura, Y.; Kawamura, K.; Kitayama, K.; Doncaster, C.P. Meta-analysis of management effects on biodiversity in plantation and secondary forests of Japan. Conserv. Sci. Pract. 2019, 1, e14. [Google Scholar] [CrossRef]
- Bonan, G.B. Forests and climate change: Forcings, feedbacks, and the climate benefits of forests. Science 2008, 320, 1444–1449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nave, L.E.; Vance, E.D.; Swanston, C.W.; Curtis, P.S. Harvest impacts on soil carbon storage in temperate forests. For. Ecol. Manag. 2010, 259, 857–866. [Google Scholar] [CrossRef]
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; López-Senespleda, E.; Montero, G.; Río, M. Do thinnings influence biomass and soil carbon stocks in Mediterranean maritime pinewoods? Eur. J. For. Res. 2013, 132, 253–262. [Google Scholar] [CrossRef]
- Roig, S.; del Río, M.; Cañellas, I.; Montero, G. Litter fall in Mediterranean Pinus pinaster Ait. stands under different thinning regimes. For. Ecol. Manag. 2005, 206, 179–190. [Google Scholar] [CrossRef]
- Pang, X.; Bao, W.; Zhu, B.; Cheng, W. Responses of soil respiration and its temperature sensitivity to thinning in a pine plantation. Agric. For. Meteorol. 2013, 171, 57–64. [Google Scholar] [CrossRef]
- Noormets, A.; Epron, D.; Domec, J.C.; McNulty, S.G.; Fox, T.; Sun, G.; King, J.S. Effects of forest management on productivity and carbon sequestration: A review and hypothesis. For. Ecol. Manag. 2015, 355, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Horner, G.J.; Baker, P.J.; Mac Nally, R.; Cunningham, S.C.; Thomson, J.R.; Hamilton, F. Forest structure, habitat and carbon benefits from thinning floodplain forests: Managing early stand density makes a difference. For. Ecol. Manag. 2010, 259, 286–293. [Google Scholar] [CrossRef]
- Martín-Benito, D.; Del Río, M.; Heinrich, I.; Helle, G.; Cañellas, I. Response of climate-growth relationships and water use efficiency to thinning in a Pinus nigra afforestation. For. Ecol. Manag. 2010, 259, 967–975. [Google Scholar] [CrossRef]
- Sohn, J.A.; Saha, S.; Bauhus, J. Potential of forest thinning to mitigate drought stress: A meta-analysis. For. Ecol. Manag. 2016, 380, 261–273. [Google Scholar] [CrossRef]
- Smith, P.; Fang, C.; Dawson, J.J.; Moncrieff, J.B. Impact of global warming on soil organic carbon. Adv. Agron. 2008, 97, 1–43. [Google Scholar] [CrossRef]
- Janssens, I.A.; Freibauer, A.; Ciais, P.; Smith, P.; Nabuurs, G.J.; Folberth, G.; Schlamadinger, B.; Hutjes, R.W.; Ceulemans, R.; Schulze, E.D.; et al. Europe’s terrestrial biosphere absorbs 7 to 12% of European anthropogenic CO2 emissions. Science 2003, 300, 1538–1542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bottner, P.; Coûteaux, M.M.; Vallejo, V.R. Soil organic matter in Mediterranean-type ecosystems and global climatic changes: A case study—The soils of the Mediterranean Basin. In Global Change and Mediterranean-Type Ecosystems; Springer: New York, NY, USA, 1995; pp. 306–325. [Google Scholar] [CrossRef]
- Cheddadi, R.; Guiot, J.; Jolly, D. The Mediterranean vegetation: What if the atmospheric CO2 increased? Landsc. Ecol. 2001, 16, 667–675. [Google Scholar] [CrossRef]
- Schlesinger, W.H.; Amundson, R. Managing for soil carbon sequestration: Let’s get realistic. Glob. Chang. Biol. 2019, 25, 386–389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Finzi, A.C.; Giasson, M.A.; Plotkin, A.A.B.; Aber, J.D.; Boose, E.R.; Davidson, E.A.; Dietze, M.C.; Ellison, A.M.; Frey, S.D.; Goldman, E.; et al. Carbon budget of the Harvard Forest Long-Term Ecological Research site: Pattern, process, and response to global change. Ecol. Monogr. 2020, 90, e01423. [Google Scholar] [CrossRef]
- Schröter, D.; Cramer, W.; Leemans, R.; Prentice, I.C.; Araújo, M.B.; Arnell, N.W.; Bondeau, A.; Bugmann, H.; Carter, T.R.; Gracia, C.A.; et al. Ecosystem service supply and vulnerability to global change in Europe. Science 2005, 310, 1333–1337. [Google Scholar] [CrossRef] [Green Version]
- Salguero, R.S.; Cerrillo, R.M.; Camarero, J.J.; Cancio, A.F. Drought-inducted growth decline of Aleppo and maritime pine forests in south-eastern Spain. For. Syst. 2010, 19, 458–469. [Google Scholar]
- Bravo, F.; Bravo-Oviedo, A.; Diaz-Balteiro, L. Carbon sequestration in Spanish Mediterranean forests under two management alternatives: A modeling approach. Eur. J. For. Res. 2008, 127, 225–234. [Google Scholar] [CrossRef]
- Leemans, R.; Rounsevell, M.D.; Midgley, G.F.; Price, J.T.; Fischlin, A.; Dube, O.P.; Velichko, A.A.; Tarazona, J.; Gopal, B.; Turley, C. Ecosystems, their properties, goods and services. In Climate Change: Impacts, Adaptation and Vulnerability. Contribution of Working Group II to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- De Giles, J.P.; Martínez, A.M.; Sola, A.J.; Poveda, J.F.M. Análisis corológico y novedades florísticas de la Sierra de los Filabres (Andalucía Oriental, España). Lagascalia 2006, 26, 51–70. [Google Scholar]
- Nicolás, A.D.; Gandullo, J.M. Ecología de los Pinares Españoles. I. Pinus Pinaster Ait; Ministerio de Agricultura, Instituto Forestal de Investigaciones y Experiencias: Madrid, Spain, 1967. [Google Scholar]
- Queen, J.P.; Quinn, G.P.; Keough, M.J. Experimental Design and Data Analysis for Biologists; Cambridge University Press: Cambridge, UK, 2002. [Google Scholar]
- Ruiz-Peinado, R.; del Rio, M. New models for estimating the carbon sink capacity of Spanish softwood species. For. Syst. 2011, 20, 176–188. [Google Scholar] [CrossRef] [Green Version]
- Penman, J.; Gytarsky, M.; Hiraishi, T.; Krug, T.; Kruger, D.; Pipatti, R.; Buendia, L.; Miwa, K.; Ngara, T.; Tanabe, K.; et al. Good Practice Guidance for Land Use, Land- Use Change and Forestry; Intergovernmental Panel on Climate Change: IPCC National Greenhouse Gas Inventories Program, Institute for Global Environmental Strategies (IGES) for the IPCC: Kanagawa, Japan, 2003. [Google Scholar]
- Nelson, D.W.; Sommers, L.E. Total carbon, organic carbon, and organic matter. In Methods of Soil Analysis: Part 3 Chemical Methods; SSSA: Madison, WI, USA, 1996; Volume 5, pp. 961–1010. [Google Scholar] [CrossRef]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and land-use change: Processes and potential. Glob. Chang. Biol. 2000, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Mann, L.K. Changes in soil carbon storage after cultivation. Soil Sci. 1986, 142, 279–288. [Google Scholar] [CrossRef]
- Farina, R.; Marchetti, A.; Francaviglia, R.; Napoli, R.; Di Bene, C. Modeling regional soil C stocks and CO2 emissions under Mediterranean cropping systems and soil types. Agric. Ecosyst. Environ. 2017, 238, 128–141. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree-Rings and Climate; Academic Press: London, UK, 1976. [Google Scholar]
- Yamaguchi, D.K. A simple method for cross-dating increment cores from living trees. Can. J. For. Res. 1991, 21, 414–416. [Google Scholar] [CrossRef]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree Ring Bull. 1983, 43, 69–78. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018; Available online: https://www.r-project.org (accessed on 12 May 2021).
- Ruiz-Peinado, R.; Bravo-Oviedo, A.; Montero, G.; Del Río, M. Carbon stocks in a Scots pine afforestation under different thinning intensities management. Mitig. Adapt. Strateg. Glob. Chang. 2016, 21, 1059–1072. [Google Scholar] [CrossRef]
- Bravo-Oviedo, A.; Ruiz-Peinado, R.; Modrego, P.; Alonso, R.; Montero, G. Forest thinning impact on carbon stock and soil condition in Southern European populations of P. sylvestris L. For. Ecol. Manag. 2015, 357, 259–267. [Google Scholar] [CrossRef]
- Navarro-Cerrillo, R.M.; Duque-Lazo, J.; Rodríguez-Vallejo, C.; Varo-Martínez, M.; Palacios-Rodríguez, G. Airborne laser scanning cartography of on-site carbon stocks as a basis for the silviculture of Pinus halepensis plantations. Remote Sens. 2018, 10, 1660. [Google Scholar] [CrossRef] [Green Version]
- Navarro-Cerrillo, R.M.; Sánchez-Salguero, R.; Rodriguez, C.; Lazo, J.D.; Moreno-Rojas, J.M.; Palacios-Rodriguez, G.; Camarero, J.J. Is thinning an alternative when trees could die in response to drought? The case of planted Pinus nigra and P. sylvestris stands in southern Spain. For. Ecol. Manag. 2019, 433, 313–324. [Google Scholar] [CrossRef]
- Romanyà, J.; Rovira, P.; Duguy, B.; Vallejo, R.; Rubio Sánchez, A. C sequestration issues in the Mediterranean soils. Greenh. Gas Budg. Soils Chang. Clim. Land Use (BurnOut) Cost 2010, 639, 15–22. [Google Scholar]
- Conkling, B.L.; Hoover, C.M.; Smith, W.D.; Palmer, C.J. Using forest health monitoring data to integrate above and below ground carbon information. Environ. Pollut. 2002, 116, S221–S232. [Google Scholar] [CrossRef]
- Johnson, D.W.; Curtis, P.S. Effects of forest management on soil C and N storage: Meta analysis. For. Ecol. Manag. 2001, 140, 227–238. [Google Scholar] [CrossRef]
- Jones, H.S.; Garrett, L.G.; Beets, P.N.; Kimberley, M.O.; Oliver, G.R. Impacts of harvest residue management on soil carbon stocks in a plantation forest. Soil Sci. Soc. Am. J. 2008, 72, 1621–1627. [Google Scholar] [CrossRef]
- Del Río, M.; Calama, R.; Cañellas, I.; Roig, S.; Montero, G. Thinning intensity and growth response in SW-European Scots pine stands. Ann. For. Sci. 2008, 65, 308. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Species | Treat | Age | N | HO | DBHm | G |
|---|---|---|---|---|---|---|
| Pinus pinaster | T0 | 42 | 1600 | 8.0 (0.11) | 19.6 (0.01) | 48.3 (5.02) |
| T30 | 42 | 1120 | 7.8 (0.07) | 23.3 (0.01) | 47.8 (4.58) | |
| T60 | 42 | 640 | 7.9 (0.08) | 27.4 (0.01) | 37.7 (4.06) | |
| Pinus nigra | T0 | 40 | 754 | 9.9 (0.7) | 11.8 (0.9) | 8.2 (1.2) |
| T30 | 40 | 528 | 9.3 (0.8) | 12.8 (0.6) | 6.8 (0.6) | |
| T60 | 40 | 302 | 9.7 (1.1) | 13.3 (0.6) | 4.2 (0.5) | |
| Pinus sylvestris | T0 | 43 | 723 | 9.6 (0.4) | 14.7 (0.6) | 12.3 (1.0) |
| T30 | 43 | 506 | 9.5 (0.4) | 15.1 (0.9) | 9.1 (1.3) | |
| T60 | 43 | 289 | 9.6 (0.4) | 15.3 (0.8) | 5.3 (0.7) |
| Species | Control | Moderate Thinning | Heavy Thinning | |
|---|---|---|---|---|
| Biomass Fraction | C in Biomass (Mg C ha−1) | |||
| Pinus pinaster | Stems | 15.3 (0.4) | 21.0 (0.3) | 29.0 (0.3) |
| Thick–medium branches | 2.2 (0.0) | 3.8 (0.0) | 6.2 (0.1) | |
| Small branches and foliage | 6.4 (0.1) | 9.6 (0.0) | 14.0 (0.1) | |
| Roots | 9.3 (0.1) | 15.2 (0.0) | 23.9 (0.2) | |
| Total | 33.2 (0.6) a | 49.6 (0.3) a | 73.1 (0.7) a | |
| Depth (cm) | SOC (Mg C ha−1) | |||
| 0–10 | 6.1 (0.1) | 8.6 (1.2) | 3.7 (0.6) | |
| 10–20 | 5.5 (0.1) | 8.1 (1.4) | 4.6 (0.8) | |
| 20–30 | 4.0 (0.1) | 8.9 (1.9) | 4.0 (0.7) | |
| 30–40 | 2.9 (0.1) | 3.4 (0.5) | 3.8 (0.9) | |
| Total | 18.5 (1.8) a | 28.1 (3.7) b | 15.5 (2.2) a | |
| Biomass Fraction | C in biomass (Mg C ha−1) | |||
| Pinus nigra | Stems | 16.4 (0.2) | 18.0 (0.2) | 20.1 (0.2) |
| Thick–medium branches | 3.6 (0.0) | 4.3 (0.1) | 4.6 (0.1) | |
| Small branches and foliage | 5.0 (0,0) | 5.9 (0.0) | 6.4 (0.0) | |
| Roots | 3.9 (0.1) | 4.8 (0.1) | 5.3 (0.0) | |
| Total | 29.0 (0.3) a | 33.0 (0.4) a | 36.4 (0.3) a | |
| Depth (cm) | SOC (Mg C ha−1) | |||
| 0–10 | 6.4 (0.7) | 5.4 (0.5) | 5.9 (0.7) | |
| 10–20 | 5.8 (0.6) | 6.4 (0.6) | 6.1 (0.6) | |
| 20–30 | 8.5 (1.0) | 7.2 (1.0) | 6.0 (0.7) | |
| 30–40 | 7.3 (0.8) | 5.1 (0.6) | 5.2 (0.6) | |
| 30–40 | 6.60 (1.24) | 4.88 | 4.22 (0.71) | |
| Total | 27.6 (2.3) a | 23.4 (2.2) a | 22.6 (1.5) a | |
| Biomass Fraction | C in Biomass (Mg C ha−1) | |||
| Pinus sylvestris | Stems | 16.0 (0.2) | 16.7 (0.1) | 17.3 (0.1) |
| Thick–medium branches | 3.1 (0.1) | 3.3 (0.0) | 3.4 (0.0) | |
| Small branches and foliage | 5.4 (0.1) | 8.2 (0.0) | 15.6 (0.0) | |
| Roots | 14.0 (0.0) | 14.8 (0.1) | 15.2 (0.0) | |
| Total | 38.5 (0.4) a | 43.0 (0.2) a | 51.5 (0.1) a | |
| Depth (cm) | SOC (Mg C ha−1) | |||
| 0–10 | 12.2 (1.2) | 10.4 (0.9) | 9.0 (0.8) | |
| 10–20 | 10.0 (1.0) | 9.5 (0.8) | 9.8 (1.0) | |
| 20–30 | 10.1 (0.8) | 6.8 (0.5) | 8.4 (0.8) | |
| 30–40 | 6.9 (1.0) | 5.4 (0.9) | 7.3 (0.9) | |
| Total | 37.0 (2.9) a | 31.6 (2.4) a | 34.0 (2.3) a | |
| Specie | Treat | Time Span | TRW | BAI20 | BAIpreT | BAIpostT | MS | Rbar | EPS |
|---|---|---|---|---|---|---|---|---|---|
| Pinus pinaster | C | 1975–2016 | 5.46 (0.39) | 4.27 (0.37) a | - | - | 0.353 | 0.778 | 0.988 |
| MT | 1975–2016 | 6.39 (0.45) | 5.33 (0.51) a | 4.10 (0.22) a | 5.77 (0.88) a | 0.328 | 0.772 | 0.987 | |
| HT | 1975–2016 | 5.48 (0.47) | 5.23 (0.49) b | 4.19 (0.28) a | 4.43 (0.49) b | 0.347 | 0.424 | 0.804 | |
| Pinus nigra | C | 1977–2016 | 2.57 (0.18) | 2.49 (0.23) a | - | - | 0.381 | 0.500 | 0.899 |
| MT | 1977–2016 | 2.30 (0.19) | 2.79 (0.30) a | 1.66 (0.22) b | 4.03 (0.6) b | 0.320 | 0.615 | 0.938 | |
| HT | 1977–2016 | 2.59 (0.27) | 2.99 (0.39) a | 1.69 (0.17) a | 3.52 (0.69) a | 0.377 | 0.603 | 0.935 | |
| Pinus sylvestris | C | 1973–2016 | 4.74 (0.38) | 3.57 (0.42) b | - | - | 0.333 | 0.693 | 0.952 |
| MT | 1973–2016 | 3.51 (0.26) | 2.93 (0.26) a | 2.13 (0.18) a | 3.01 (0.44) a | 0.336 | 0.614 | 0.937 | |
| HT | 1973–2016 | 4.99 (0.39) | 5.50 (0.57) a | 3.62 (0.43) a | 5.87 (0.95) a | 0.398 | 0.422 | 0.874 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cachinero-Vivar, A.M.; Palacios-Rodríguez, G.; Lara-Gómez, M.A.; Navarro-Cerrillo, R.M. Thinning Effect of C Sequestration along an Elevation Gradient of Mediterranean Pinus spp. Plantations. Forests 2021, 12, 1583. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111583
Cachinero-Vivar AM, Palacios-Rodríguez G, Lara-Gómez MA, Navarro-Cerrillo RM. Thinning Effect of C Sequestration along an Elevation Gradient of Mediterranean Pinus spp. Plantations. Forests. 2021; 12(11):1583. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111583
Chicago/Turabian StyleCachinero-Vivar, Antonio M., Guillermo Palacios-Rodríguez, Miguel A. Lara-Gómez, and Rafael M. Navarro-Cerrillo. 2021. "Thinning Effect of C Sequestration along an Elevation Gradient of Mediterranean Pinus spp. Plantations" Forests 12, no. 11: 1583. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111583