
Effects of Exogenous Ethylene and Cobalt Chloride on Root Growth of Chinese Fir Seedlings under Phosphorus-Deficient Conditions

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Experimental Design
2.3. Harvest and Data Collection
2.4. Statistical Analyses
3. Results
3.1. Two-Factor Analysis of the Influence of P Supply Level and Exogenous Additive Concentration on the Determination Indices
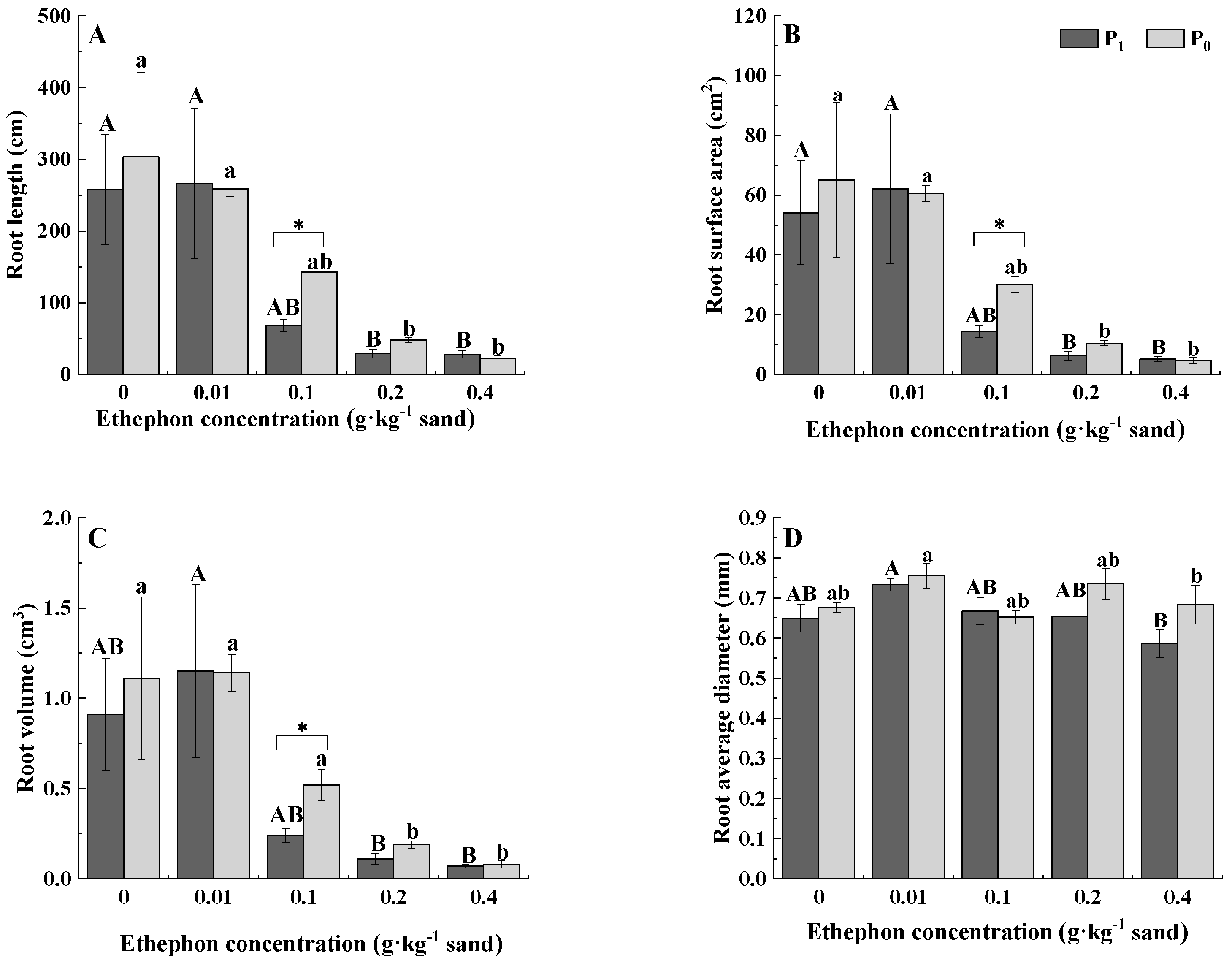
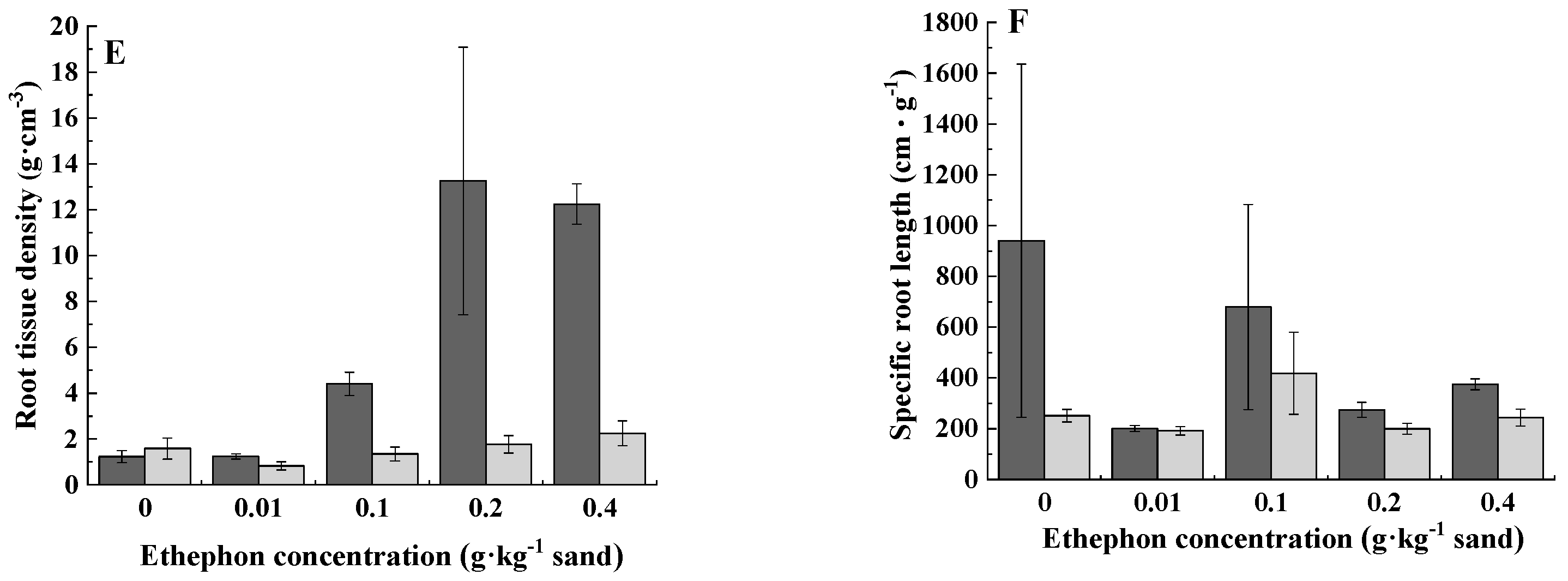
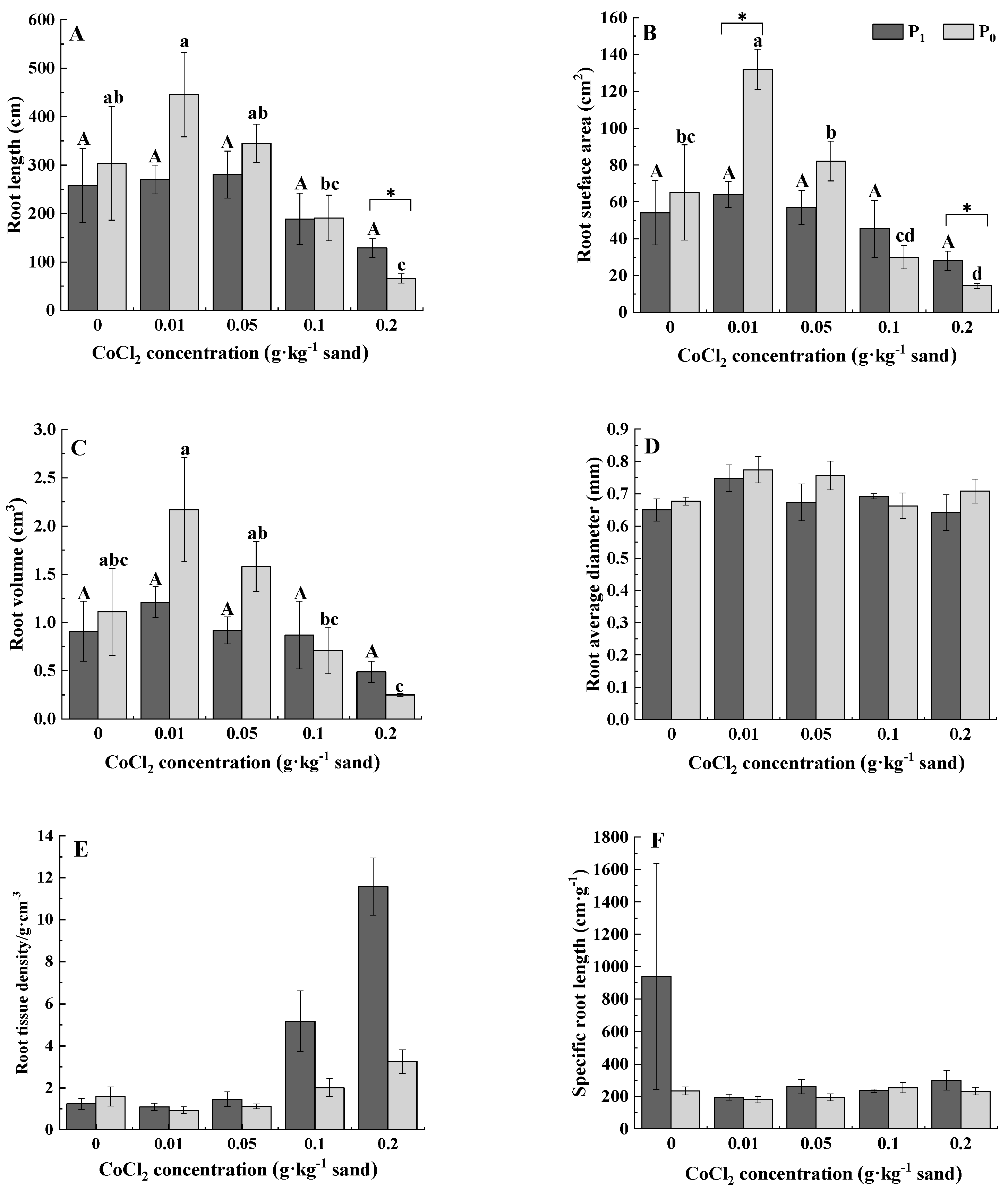
3.2. Root Morphology
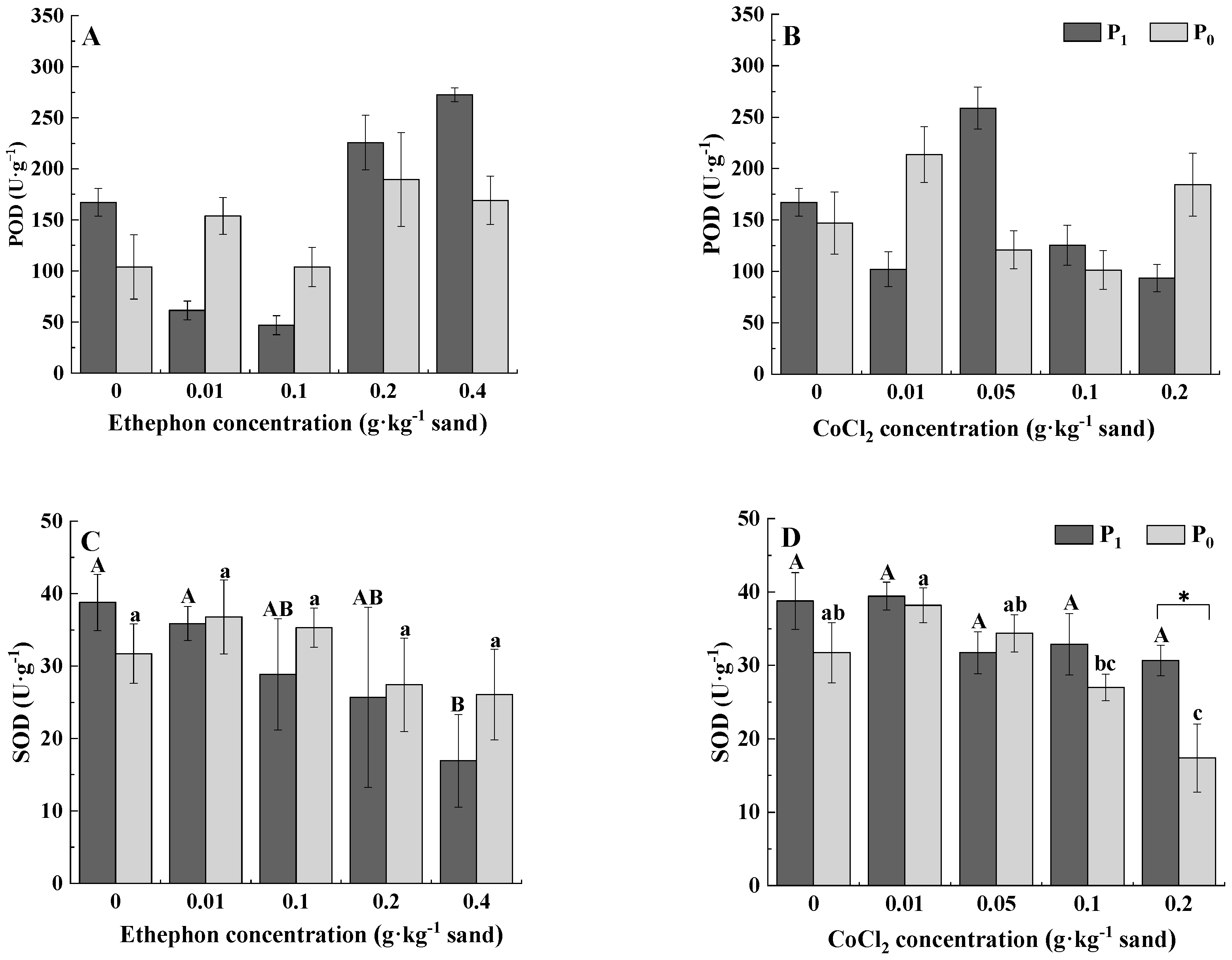
3.3. Protective Enzyme Activities
3.4. MDA Content
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Erpen, L.; Devi, H.S.; Grosser, J.W.; Dutt, M. Potential use of the DREB/ERF, MYB, NAC and WRKY transcription factors to improve abiotic and biotic stress in transgenic plants. Plant Cell Tissue Organ Cult. 2018, 132, 1–25. [Google Scholar] [CrossRef]
- Maria, J.G.; Francisco, J.R.; Carlos, L.; Esteban, A.; Rafael, P.V. Ethylene and the regulation of physiological and morphological responses to nutrient deficiencies. Plant Physiol. 2015, 169, 51–60. [Google Scholar]
- Zhang, M.; Smith, J.A.C.; Harberd, N.P.; Jiang, C.F. The regulatory roles of ethylene and reactive oxygen species (ROS) in plant salt stress responses. Plant Mol. Biol. 2016, 91, 651–659. [Google Scholar] [CrossRef]
- Yu, W.Q.; Sheng, J.P.; Zhao, R.R.; Wang, Q.; Ma, P.H.; Shen, L. Ethylene biosynthesis is involved in regulating chilling tolerance and SlCBF1 gene expression in tomato fruit. Postharvest Biol. Technol. 2019, 149, 139–147. [Google Scholar] [CrossRef]
- Valluru, R.; Davies, W.J.; Reynolds, M.P.; Dodd, I.C. Foliar abscisic acid-to-ethylene accumulation and response regulate shoot growth sensitivity to mild drought in wheat. Front. Plant Sci. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, C.L.; Cao, J.D.; Meng, F.W.; Yu, Y.Q.; Huang, J.K.; Jiang, L.; Liu, M.X.; Zhang, Z.G.; Chen, X.W.; Miyamoto, K.J.; et al. Activation of ethylene signaling pathways enhances disease resistance by regulating ROS and phytoalexin production in rice. Plant J. 2017, 89, 338–353. [Google Scholar] [CrossRef] [Green Version]
- Zou, X.H.; Hu, Y.N.; Wei, D.; Chen, S.T.; Wu, P.F.; Ma, X.Q. Correlation between endogenous hormone and the adaptability of Chinese fir with high phosphorus-use efficiency to low phosphorus stress. Chin. J. Plant Ecol. 2019, 43, 139–151. [Google Scholar]
- Xie, Y.; Li, A.; Ding, Z.J.; Zheng, S.J. Current researches on the roles of ethylene in the regulation of plant root hairs. Plant Physiol. J. 2020, 56, 2517–2525. [Google Scholar]
- Iqbal, N.; Trivellini, A.; Masood, A.; Ferrante, A.; Khan, N.A. Current understanding on ethylene signaling in plants: The influence of nutrient availability. Plant Physiol. Biochem. 2013, 73, 128–138. [Google Scholar] [CrossRef]
- Pierik, R.; Sasidharan, R.; Voesenek, L. Growth control by ethylene: Adjusting phenotypes to the environment. J. Plant Growth Regul. 2007, 26, 188–200. [Google Scholar] [CrossRef] [Green Version]
- Basu, P.; Zhang, Y.J.; Lynch, J.P.; Brown, K.M. Ethylene modulates genetic, positional, and nutritional regulation of root plagiogravitropism. Funct. Plant Biol. 2007, 34, 41–51. [Google Scholar] [CrossRef]
- Song, L.; Dong, L. Ethylene and plant responses to phosphate deficiency. Front. Plant Sci. 2015, 6, 796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lei, M.G.; Zhu, C.M.; Liu, Y.D.; Karthikeyan, A.S.; Bressan, R.A.; Raghothama, K.G.; Liu, D. Ethylene signalling is involved in regulation of phosphate starvation-induced gene expression and production of acid phosphatases and anthocyanin in Arabidopsis. New Phytol. 2011, 189, 1084–1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.Q.; Hu, Y.X.; Liu, J.; Wang, H.; Wei, J.H.; Sun, P.D.; Wu, L.F.; Zheng, H.J. Progress of ethylene action mechanism and its application on plant type formation in crops. Saudi J. Biol. Sci. 2020, 27, 1667–1673. [Google Scholar] [CrossRef] [PubMed]
- Pahwa, K.; Ghai, N.; Singh, I.; Singh, S.; Kaur, J.; Dhingra, M. Influence of ethylene and cobalt chloride on photosynthetic parameters and pedicel anatomy of pigeonpea (Cajanus cajan L.) genotypes. J. Environ. Biol. 2017, 38, 367–374. [Google Scholar] [CrossRef]
- Müller, M. Foes or friends: ABA and ethylene interaction under abiotic stress. Plants 2021, 10, 448. [Google Scholar] [CrossRef]
- Ren, H.Y.; Han, G.D.; Li, M.H.; Gao, C.P.; Jiang, L. Ethylene-regulated leaf lifespan explains divergent responses of plant productivity to warming among three hydrologically different growing seasons. Glob. Chang. Biol. 2021, 27, 4169–4180. [Google Scholar] [CrossRef]
- Chen, S.Y.; Ma, X.Q.; Wu, P.F.; Lin, W.J.; Chen, Y.L.; Zou, X.H.; Chen, N.L. Evaluation on growth and seed characters of different clones in 1.5-generation seed orchard of Chinese fir. J. Trop. Subtrop. Bot. 2014, 22, 28–291. [Google Scholar]
- Wu, P.F.; Ma, X.Q.; Tigabu, M.; Wang, C.; Liu, A.Q.; Odén, P.C. Root morphological plasticity and biomass production of two Chinese fir clones with high phosphorus efficiency under low phosphorus stress. Can. J. For. Res. 2011, 41, 228–234. [Google Scholar] [CrossRef]
- Gao, B.; Cao, C.L.; Li, T. Effect of ethylene on morphology and physiological characteristic of soybean seedlings under low-phosphorus stress. Soybean Sci. 2012, 31, 58–63. [Google Scholar]
- Zou, S.Y.; Li, D.D.; Wang, J.S.; Di, N.; Liu, J.Q.; Wang, Y.; Li, G.D.; Duan, J.; Jia, L.M.; Xi, B.Y. Response of fine roots to soil moisture of different gradients in young Populus tomentosa plantation. Sci. Silvae Sin. 2019, 55, 124–137. [Google Scholar]
- Wu, P.F.; Wang, G.Y.; Farooq, T.H.; Li, Q.; Zou, X.H.; Ma, X.Q. Low phosphorus and competition affect Chinese fir cutting growth and root organic acid content: Does neighboring root activity aggravate P nutrient deficiency? J. Soils Sediments 2017, 17, 2775–2785. [Google Scholar] [CrossRef]
- Zhang, L.L.; Ren, R.F.; Jiang, X.R.; Zhou, H.; Liu, Y. Exogenous ethylene increases the viability of cryopreserved Dendrobium protocorm-like bodies by regulating the hydrogen peroxide and its mediated oxidative stress. Plant Cell Tissue Organ Cult. 2021, 145, 19–27. [Google Scholar] [CrossRef]
- Qin, H.; Ma, C.L.; Zhou, Y.; Miao, Y.C.; Huang, R.F. Molecular modulation of root development by ethylene. Small Methods 2020, 4, 1–7. [Google Scholar] [CrossRef]
- Růzicka, K.; Ljung, K.; Vanneste, S.; Podhorska, R.; Beeckman, T.; Friml, J.; Benkova, E. Ethylene regulates root growth through effects on auxin biosynthesis and transport-dependent auxin distribution. Plant Cell 2007, 19, 2197–2212. [Google Scholar] [CrossRef] [Green Version]
- Miao, Z.Q.; Zhao, P.X.; Mao, J.L.; Yu, L.H.; Yuan, Y.; Tang, H.; Liu, Z.B.; Xiang, C.B. HOMEOBOX PROTEIN52 mediates the crosstalk between ethylene and auxin signaling during primary root elongation by modulating auxin transport-related gene expression. Plant Cell 2018, 30, 2761–2778. [Google Scholar] [CrossRef] [Green Version]
- Neumann, G. The role of ethylene in plant adaptations for phosphate acquisition in soils–A review. Front. Plant Sci. 2016, 6, 1224. [Google Scholar] [CrossRef] [Green Version]
- Lai, H.Y.; Wu, K.; Wang, N.M.; Wu, W.J.; Zou, X.H.; Ma, X.Q.; Wu, P.F. Relationship between volatile organic compounds released and growth of Cunninghamia lanceolata roots under low-phosphorus conditions. iForest 2018, 11, 713–720. [Google Scholar] [CrossRef]
- Zhang, X.L.; Wu, Q.; Tao, Y.; Zhu, X.F.; Takahashi, N.; Umeda, M.; Shen, R.F.; Ma, J.F. ANAC044 is associated with P reutilization in P deficient Arabidopsis thaliana root cell wall in an ethylene dependent manner. Environ. Exp. Bot. 2021, 185, 104386. [Google Scholar] [CrossRef]
- Fu, D.; Xiang, H.L.; Yu, C.; Zheng, X.D.; Yu, T. Colloidal chitin reduces disease incidence of wounded pear fruit inoculated by Penicillium expansum. Postharvest Biol. Technol. 2016, 111, 1–5. [Google Scholar] [CrossRef]
- Zhu, C.H.; Yang, N.; Guo, Z.F.; Qian, M.; Gan, L.J. An ethylene and ROS-dependent pathway is involved in low ammonium-induced root hair elongation in Arabidopsis seedlings. Plant Physiol. Biochem. 2016, 105, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Chiriboga, M.A.; Bordonaba, J.G.; Schotsmans, W.C.; Larrigaudière, C.; Recasens, I. Antioxidant potential of ‘Conference’ pears during cold storage and shelf life in response to 1-methylcyclopropene. LWT Food Sci. Technol. 2013, 51, 170–176. [Google Scholar] [CrossRef]
- Gamrasni, D.; Feldmesser, E.; Ben-Arie, R.; Raz, A.; Tabatznik, A.A.; Glikman, M.; Aharoni, A.; Goldway, M. Gene expression in 1-Methylcyclopropene (1-MCP) treated tomatoes during pre-climacteric ripening suggests shared regulation of methionine biosynthesis, ethylene production and respiration. Agronomy 2020, 10, 1669. [Google Scholar] [CrossRef]
- Montero-Palmero, M.B.; Ortega-Villasante, C.; Escobar, C.; Hernández, L.E. Are plant endogenous factors like ethylene modulators of the early oxidative stress induced by mercury? Front. Environ. Sci. 2014, 2, 1–8. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Seedling Traits | Ethephon | CoCl2 | ||||
|---|---|---|---|---|---|---|
| F Value | ||||||
| a | b | a × b | a | b | a × b | |
| Root length | 6.030 * | 3.702 * | 0.807 | 10.347 ** | 8.367 ** | 0.794 |
| Root surface area | 5.660 * | 3.457 * | 0.668 | 10.701 ** | 8.146 ** | 0.955 |
| Root volume | 5.196 * | 3.230 * | 0.57 | 9.982 ** | 7.329 ** | 1.054 |
| Root diameter | 4.573 * | 3.596 * | 0.986 | 1.75 | 1.805 | 0.537 |
| Root tissue density | 14.051 ** | 4.703 ** | 3.62 * | 30.672 ** | 34.790 ** | 15.723 ** |
| Specific root length | 2.003 | 1.424 | 1.406 | 1.706 | 1.643 | 1.593 |
| POD | 0.978 | 12.027 ** | 6.015 ** | 0.359 | 2.978 * | 9.380 ** |
| SOD | 0.411 | 2.461 | 0.695 | 7.190 * | 5.738 ** | 2.075 |
| MDA | 0.041 | 1.901 | 0.366 | 2.269 | 0.788 | 1.746 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, H.; Wang, N.; Zheng, S.; Chen, M.; Ma, X.; Wu, P. Effects of Exogenous Ethylene and Cobalt Chloride on Root Growth of Chinese Fir Seedlings under Phosphorus-Deficient Conditions. Forests 2021, 12, 1585. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111585
Zhang H, Wang N, Zheng S, Chen M, Ma X, Wu P. Effects of Exogenous Ethylene and Cobalt Chloride on Root Growth of Chinese Fir Seedlings under Phosphorus-Deficient Conditions. Forests. 2021; 12(11):1585. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111585
Chicago/Turabian StyleZhang, Hui, Nemin Wang, Shanshan Zheng, Min Chen, Xiangqing Ma, and Pengfei Wu. 2021. "Effects of Exogenous Ethylene and Cobalt Chloride on Root Growth of Chinese Fir Seedlings under Phosphorus-Deficient Conditions" Forests 12, no. 11: 1585. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111585