
Influence of Edaphic Salinity on Leaf Morphoanatomical Functional Traits on Juvenile and Adult Trees of Red Mangrove (Rhizophora mangle): Implications with Relation to Climate Change

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
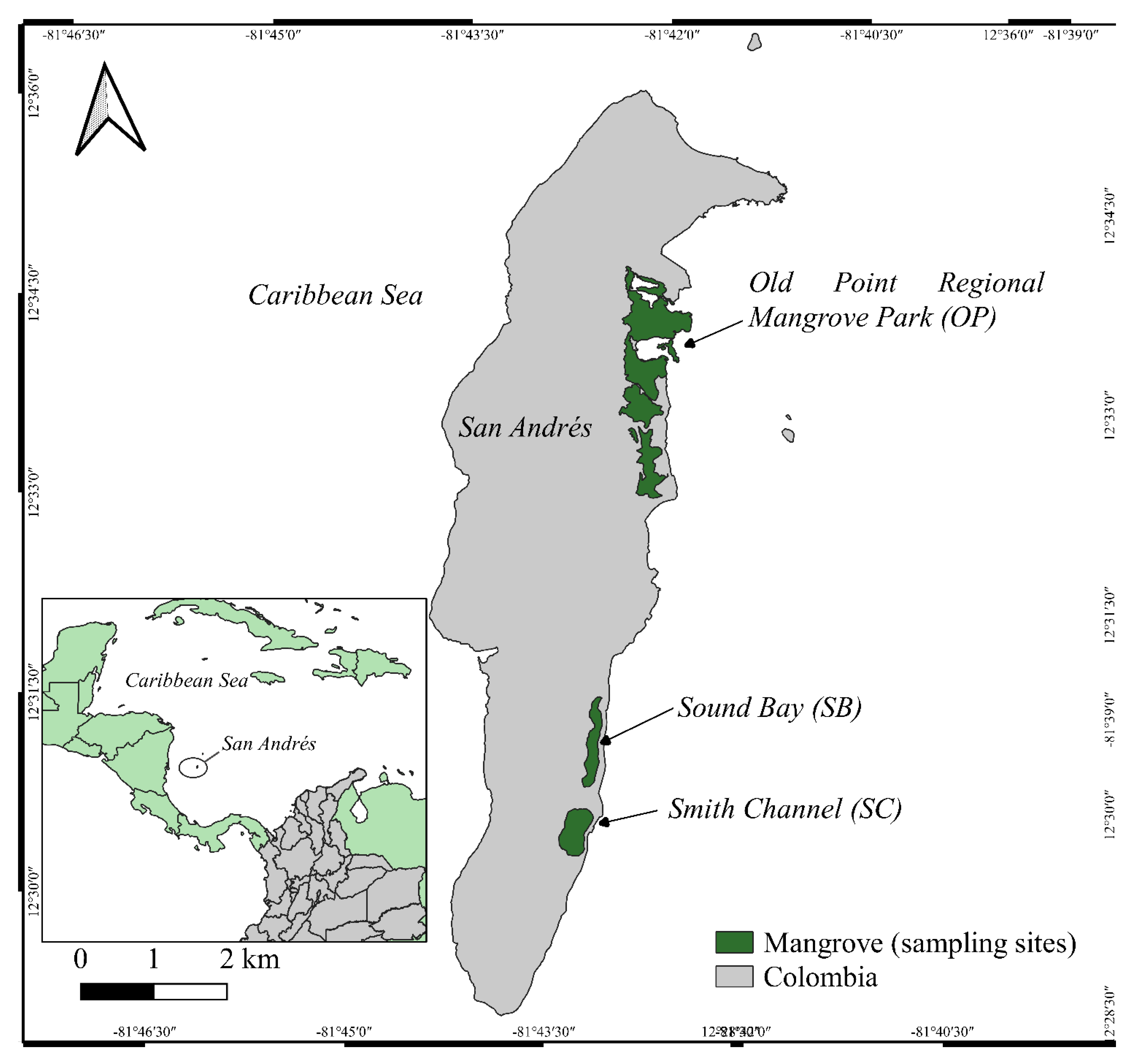
2.1. Study Area
2.2. Collection and Fixation of Plant Organs
2.3. Physicochemical Variables of Interstitial Water
2.4. Preparation of Leaves for Measuring Foliar Area
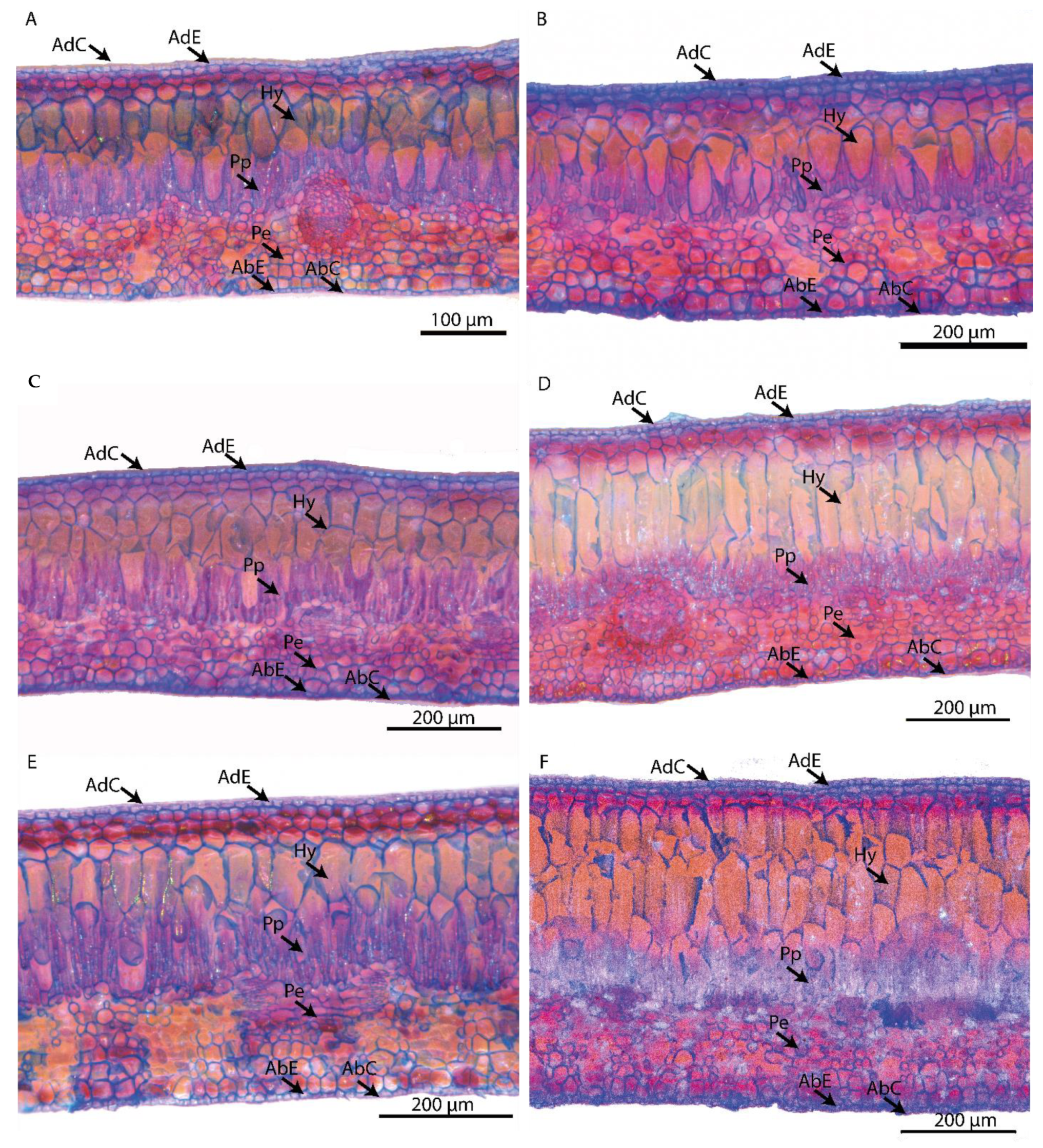
2.5. Preparation of Tissues for Fresh Histological Measurement
2.6. Measurement of Tissues
2.7. Statistical Analysis of Morphological and Anatomical Measurements
3. Results
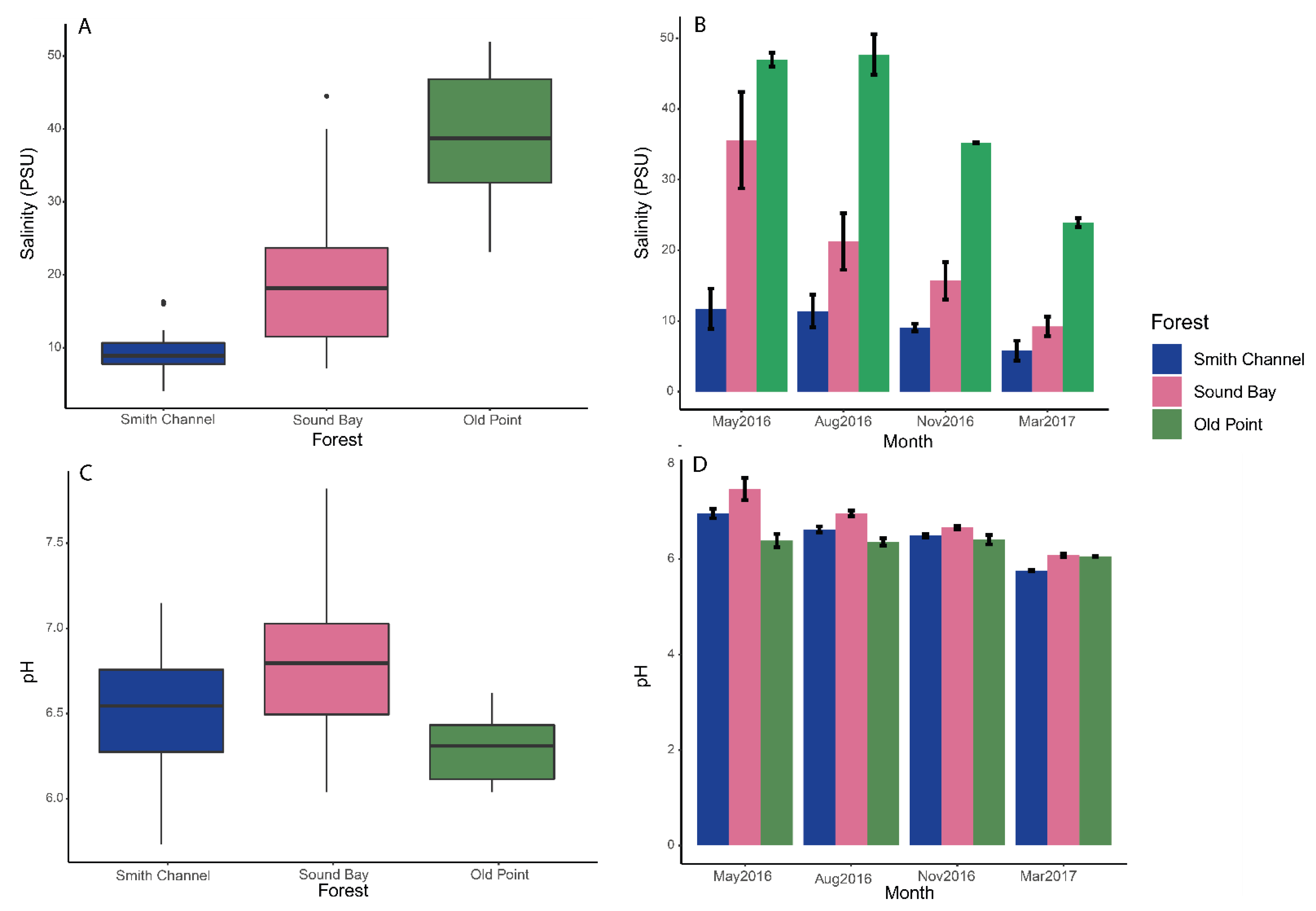
3.1. Physicochemical Variables
3.1.1. Salinity
3.1.2. pH
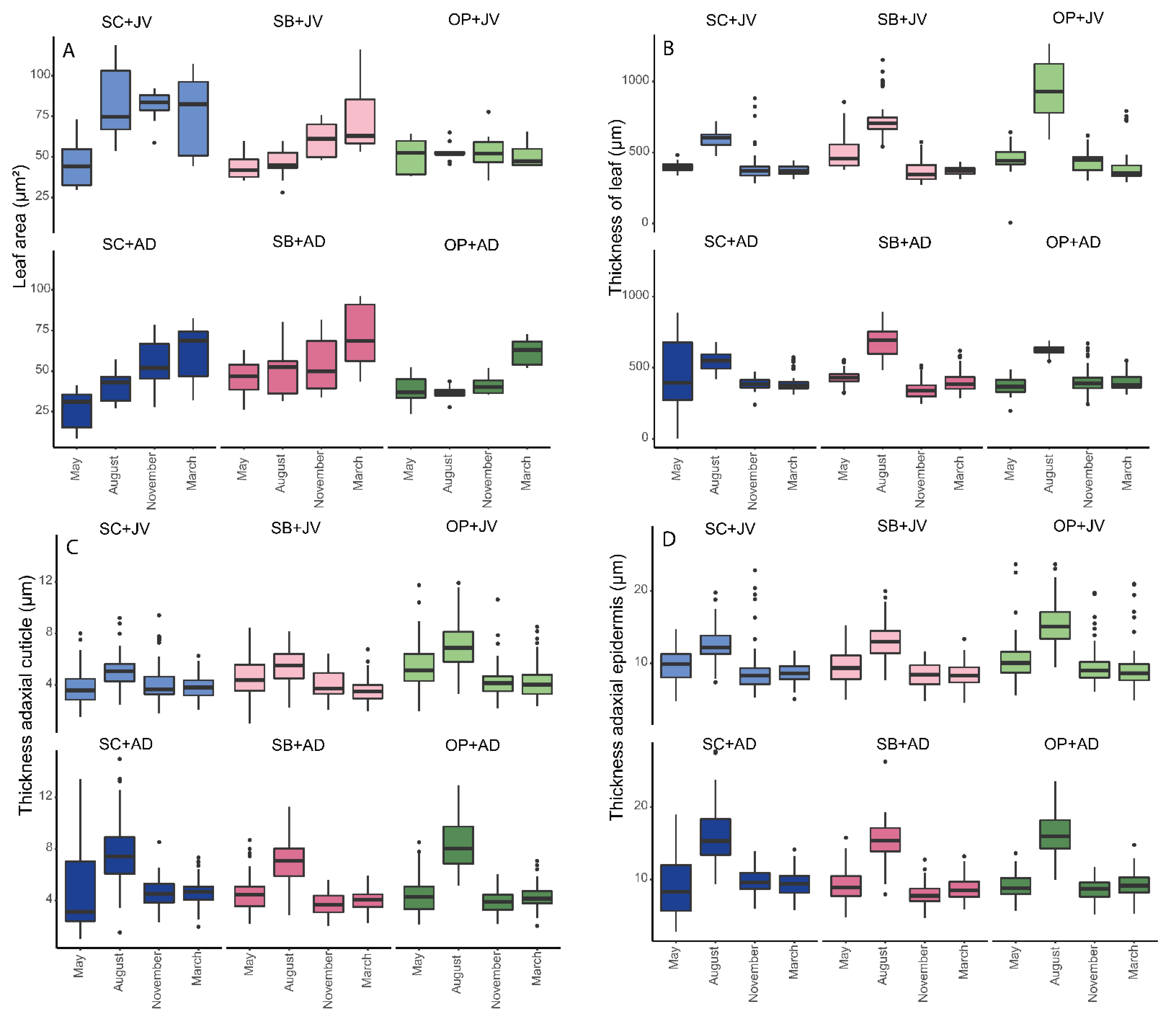
3.2. Morphoanatomical Measurements on Leaf of R. mangle
3.2.1. Comparison of Tissues between Groups
3.2.2. Spearman’s Correlation between Salinity and Morphoanatomical Traits by Age and Forest
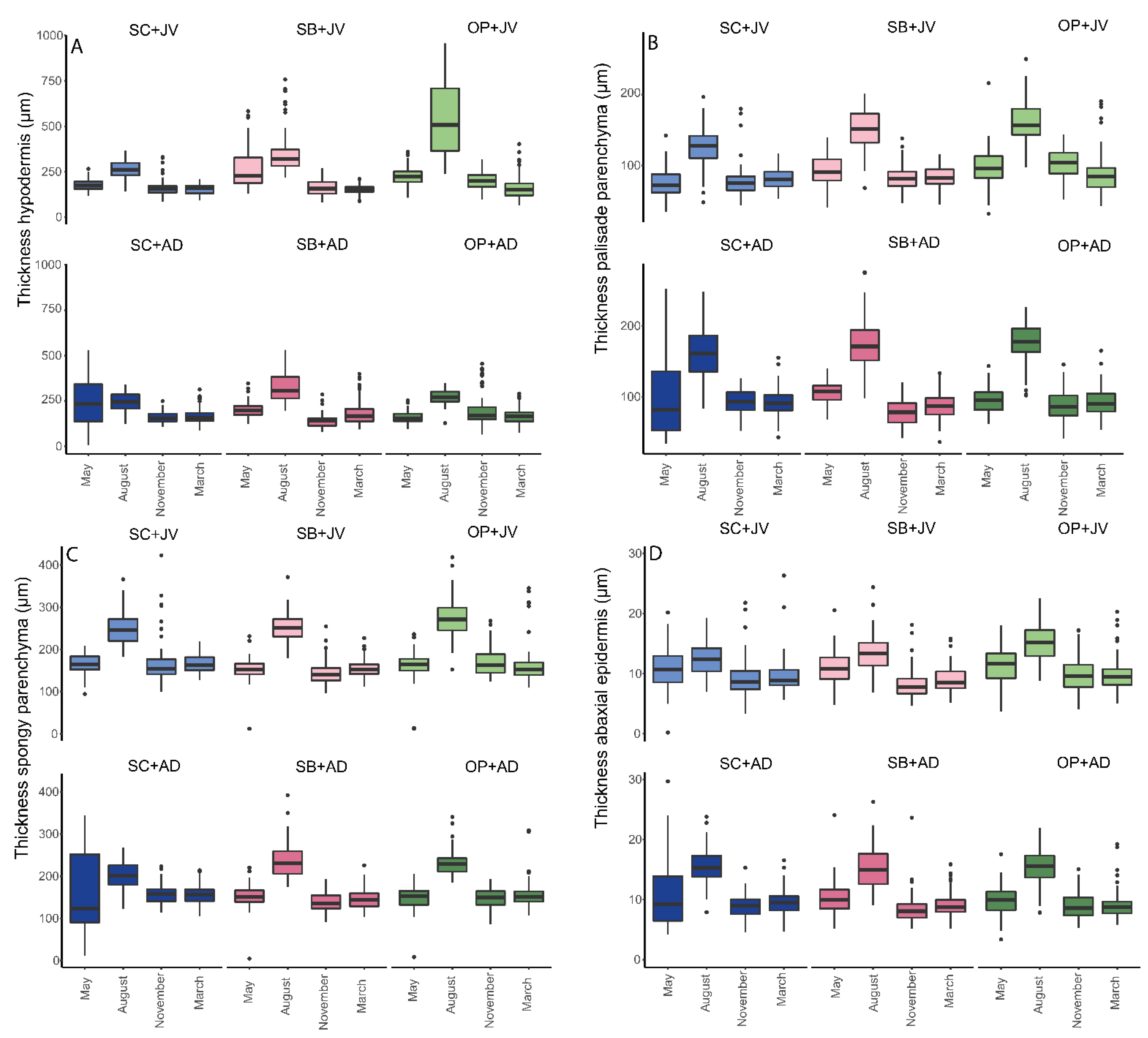
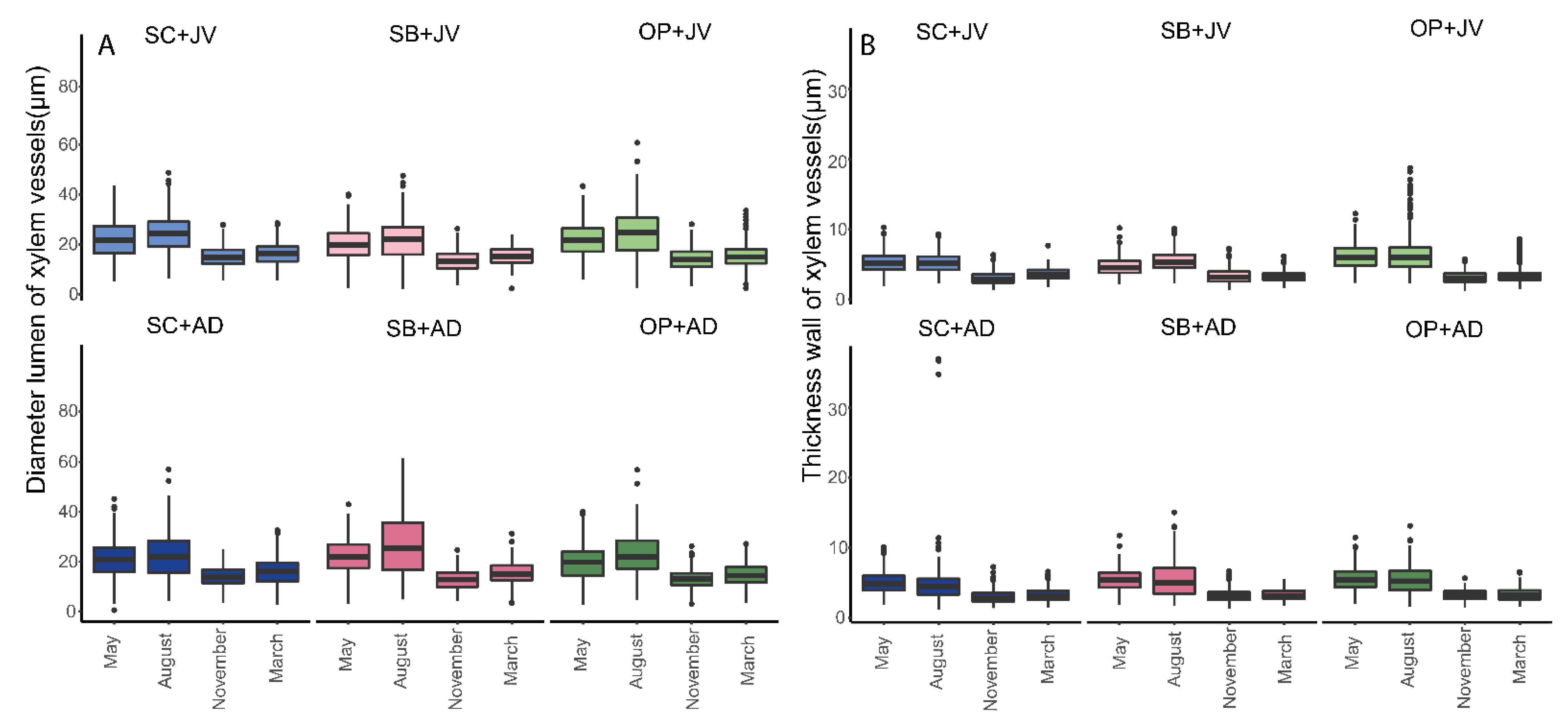
3.2.3. Measurement of Morphoanatomical Traits Evaluated (by Age and Time)
4. Discussion
4.1. Functional Traits and Response to the Salinity Ranges
4.2. Implications of the Responses of the Morphoanatomic Functional Traits of R. mangle to the Variation of Salinity in Possible Climate Change Events (Including the ENSO Phenomenon)
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Smith, S.M.; Snedaker, S.C. Salinity Responses in Two Populations of Viviparous Rhizophora mangle L. Seedlings. Biotropica 1995, 27, 435–444. [Google Scholar] [CrossRef]
- Arbeláez-Cortés, E.; Castillo-Cárdenas, M.F.; Toro-Perea, N.; Cárdenas-Henao, H. Genetic structure of the red mangrove (Rhizophora mangle L.) on the Colombian Pacific detected by microsatellite molecular markers. Hydrobiologia 2007, 583, 321–330. [Google Scholar] [CrossRef]
- Hao, G.-Y.; Jones, T.J.; Luton, C.; Zhang, Y.-J.; Manzane, E.; Scholz, F.G.; Bucci, S.J.; Cao, K.-F.; Goldstein, G. Hydraulic redistribution in dwarf Rhizophora mangle trees driven by interstitial soil water salinity gradients: Impacts on hydraulic architecture and gas exchange. Tree Physiol. 2009, 29, 697–705. [Google Scholar] [CrossRef] [Green Version]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: London, UK, 1986. [Google Scholar]
- Davis, J.H. The ecology and geologic role of mangroves in Florida. Publ. Carnegie Inst. Wash. 1940, 517, 303–412. [Google Scholar]
- Vanegas, F.H. Protocolo Para la Reproducción de Mangle Rojo (Rhizophora mangle) en Vivero; Pro natura Veracruz, A.C. e ITSZ Ingenieros: Veracruz, Mexico, 2013; p. 73. [Google Scholar]
- Scholander, P.F. How Mangroves Desalinate Seawater. Physiol. Plant. 1968, 21, 251–261. [Google Scholar] [CrossRef]
- Lugo, A.E.; Snedaker, S.C. The ecology of mangroves. Annu. Rev. Ecol. Syst. 1974, 5, 39–64. [Google Scholar] [CrossRef]
- Salas Estrada, J.B. Arboles de Nicaragua; Instituto Nicaragüense de Recursos Naturales y del Ambiente: Managua, Nicaragua, 1993. [Google Scholar]
- Ortíz Álvarez, O. El Sector Forestal de la Península Zapata, en Matanzas, Cuba: Aspectos de su Interrelación Con el Cambio Climático. Ph.D. Thesis, Universidad de Alicante, Alicante, Spain, Universidad de Pinar del Río, Pinar del Río, Cuba, 2010. [Google Scholar]
- Werner, A.; Stelzer, R. Physiological responses of the mangrove Rhizophora mangle grown in the absence and presence of NaCl. Plant Cell Environ. 1990, 13, 243–255. [Google Scholar] [CrossRef]
- Camilleri, J.C.; Ribi, G. Leaf Thickness of Mangroves (Rhizophora mangle) Growing in Different Salinities. Biotropica 1983, 15, 139. [Google Scholar] [CrossRef]
- Rodríguez, J.A.R.; Pineda, J.E.M.; Melgarejo, L.M.; Calderón, J.H.M. Functional traits of leaves and forest structure of neotropical mangroves under different salinity and nitrogen regimes. Flora 2018, 239, 52–61. [Google Scholar] [CrossRef]
- Peterson, C.A. Exodermal Casparian bands: Their significance for ion uptake by roots. Physiol. Plant. 1988, 72, 204–208. [Google Scholar] [CrossRef]
- Parida, A.K.; Jha, B. Salt tolerance mechanisms in mangroves: A review. Trees 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Lopes, D.; Tognella, M.M.P.; Falqueto, A.; Soares, M. Salinity variation effects on photosynthetic responses of the mangrove species Rhizophora mangle L. growing in natural habitats. Photosynthetica 2019, 57, 1142–1155. [Google Scholar] [CrossRef] [Green Version]
- Ramírez Correa, J.A.; Molina Grajales, E.C.; Bernal Escobar, M. Anillos Anuales y Clima en Rhizophora mangle L. de la Bahía de Cispatá; Revista Facultad Nacional de Agronomía: Medellín, Colombia, 2010; pp. 5639–5650. [Google Scholar]
- FAO. The World’s Mangroves 1980–2005; FAO: Rome, Italy, 2007. [Google Scholar]
- Spalding, M. World Atlas of Mangroves; Routledge: London, UK, 2010. [Google Scholar] [CrossRef]
- Alongi, D.M. Carbon payments for mangrove conservation: Ecosystem constraints and uncertainties of sequestration potential. Environ. Sci. Policy 2011, 14, 462–470. [Google Scholar] [CrossRef]
- Sandilyan, S.; Kathiresan, K. Mangrove conservation: A global perspective. Biodivers. Conserv. 2012, 21, 3523–3542. [Google Scholar] [CrossRef]
- Russi, D.; ten Brink, P.; Farmer, A.; Badura, T.; Coates, D.; Förster, J.; Kumar, R.; Davidson, N. The Economics of Ecosystems and Biodiversity for Water and Wetlands; IEEP: London, UK; Brussels, Belgium, 2013; Volume 78. [Google Scholar]
- Ellison, J.C. Vulnerability assessment of mangroves to climate change and sea-level rise impacts. Wetl. Ecol. Manag. 2015, 23, 115–137. [Google Scholar] [CrossRef] [Green Version]
- Ward, R.D.; Friess, D.A.; Day, R.; MacKenzie, R.A. Impacts of climate change on mangrove ecosystems: A region by region overview. Ecosyst. Health Sustain. 2016, 2, e01211. [Google Scholar] [CrossRef] [Green Version]
- Osland, M.J.; Feher, L.C.; López-Portillo, J.; Day, R.; Suman, D.O.; Menéndez, J.M.G.; Rivera-Monroy, V.H. Mangrove forests in a rapidly changing world: Global change impacts and conservation opportunities along the Gulf of Mexico coast. Estuar. Coast. Shelf Sci. 2018, 214, 120–140. [Google Scholar] [CrossRef]
- Alongi, D.M. The Impact of Climate Change on Mangrove Forests. Curr. Clim. Chang. Rep. 2015, 1, 30–39. [Google Scholar] [CrossRef]
- Díaz, M.; Manuel, J.; Díaz-Pulido, G.; Garzón-Ferreira, J.; Geister, J.; Sánchez, J.A.; Zea, S. Atlas de los Arrecifes Coralinos del Caribe Colombiano: I. Complejos Arrecifales Oceánicos; Litoflash: Santa Marta, Colombia, 1996. [Google Scholar]
- IGAC. Diccionario Geográfico de Colombia; Instituto Geográfico Agustín: Bogotá, Colombia, 1996.
- Gavio, B.P.-C. Historical analysis (2000–2005) of the coastal water quality in San Andrés Island, Sea Flower Biosphere Reserve Caribbean, Colombia. Mar. Pollut. Bull. 2010, 60, 1018–1030. [Google Scholar] [CrossRef]
- Coralina, I.; Gómez-López, D.I.; Segura-Quintero, C.; Sierra-Correa, P.C.; Garay-Tinoco, J. Atlas de la Reserva de Biósfera Seaflower. Archipiélago de San Andrés, Providencia y Santa Catalina; Serie de Publicaciones Especiales de INVEMAR # 28; Instituto de Investigaciones Marinas y Costeras “José Benito Vives De Andréis”-INVEMAR-y Corporación para el Desarrollo Sostenible del Archipiélago de San Andrés, Providencia y Santa Catalina-CORALINA: Santa Marta, Colombia, 2012; 180p. [Google Scholar]
- IDEAM. Datos de las Variables Climáticas de la Isla de San Andrés, Providenciay Santa Catalina; IDEAM: Bogotá, Colombia, 1995.
- Sánchez-Núñez, D.A.; Mancera-Pineda, J.E. Flowering patterns in three neotropical mangrove species: Evidence from a Caribbean island. Aquat. Bot. 2011, 94, 177–182. [Google Scholar] [CrossRef]
- Sánchez-Núñez, D.A.; Mancera-Pineda, J.E. Pollination and fruit set in the main neotropical mangrove species from the Southwestern Caribbean. Aquat. Bot. 2012, 103, 60–65. [Google Scholar] [CrossRef]
- Robles Sánchez, A. Respuestas Morfoanatómicas de Rhizophora mangle a Diferentes Regímenes de Salinidad en la Isla de San Andrés–Colombia-Caribe Colombiano. Master’s Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2019. [Google Scholar]
- Medina-Calderón, J.H.; Mancera-Pineda, J.E.; Castañeda-Moya, E.; Rivera-Monroy, V.H. Hydroperiod and Salinity Interactions Control Mangrove Root Dynamics in a Karstic Oceanic Island in the Caribbean Sea (San Andres, Colombia). Front. Mar. Sci. 2021, 7, 1194. [Google Scholar] [CrossRef]
- Sánchez-Núñez, D. Patrones de Floración, Polinización y Produccion de Frutos. Master’s Thesis, Universidad Nacional de Colombia, Bogotá, Colombia, 2009. [Google Scholar]
- Ito, T. The Venice system for the classification of marine waters according to salinity: Symposium on the classification of brackish waters, Venice, 8–14 April 1958. Jpn. J. Limnol. (Rikusuigaku Zasshi) 1959, 9, 119–120. [Google Scholar] [CrossRef] [Green Version]
- Medina Calderón, J.H. Estructura, Biomasa y Producción Primaria Neta de Bosques de Manglar en Ambientes Kársticos de Islas Oceánicas. Ph.D. Thesis, Universidad Nacional de Colombia sede Caribe, San Andrés, Colombia, 2016. [Google Scholar]
- McKee, K.L.; Mendelssohn, I.A.; Hester, M.W. Reexamination of pore water sulfide concentrations and redox potentials near the aerial roots of Rhizophora mangle and Avicennia germinans. Am. J. Bot. 1988, 75, 1352–1359. [Google Scholar] [CrossRef]
- Donahue, R.L.; Miller, R.W.; Shickluna, S.C. An Introduction to Soils and Plant Growth; Prentice Hall, Inc.: Englewood Cliffs, NJ, USA, 1983. [Google Scholar]
- Wakushima, S.; Kuraishi, S.; Sakurai, N. Soil salinity and pH in Japanese mangrove forests and growth of cultivated mangrove plants in different soil conditions. J. Plant Res. 1994, 107, 39–46. [Google Scholar] [CrossRef]
- Araújo, R.J.; Jaramillo, J.C.; Snedaker, S.C. LAI and leaf size differences in two red mangrove forest types in south Florida. Bull. Mar. Sci. 1997, 60, 643–647. [Google Scholar]
- Ball, M. Ecophysiology of mangroves. Trees 1988, 2, 129–142. [Google Scholar] [CrossRef]
- Reef, R.; Lovelock, C. Regulation of water balance in mangroves. Ann. Bot. 2014, 115, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Melcher, P.; Goldstein, G.; Meinzer, F.; Yount, D.; Jones, T.; Holbrook, N.; Huang, C. Water relations of coastal and estuarine Rhizophora mangle: Xylem pressure potential and dynamics of embolism formation and repair. Oecologia 2001, 126, 182–192. [Google Scholar] [CrossRef]
- Terrazas, T.; Yáñez-Espinosa, L.; López-Mata, L.; Valdez-Hernández, J.I. Wood variation in Laguncularia racemosa and its effect on fibre quality. Wood Sci. Technol. 2004, 38, 217–226. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef] [PubMed]
- Lovelock, C.E.; Ball, M.C.; Feller, I.C.; Engelbrecht, B.M.J.; Ewe, M.L. Differential effects of nitrogen and phosphorus on hydraulic conductance of mangroves along salinity gradients. Physiol. Plant. 2006, 127, 457–464. [Google Scholar] [CrossRef]
- Schmitz, N.; Verheyden, A.; Beeckman, H.; Kairo, J.G.; Koedam, N. Influence of a Salinity Gradient on the Vessel Characters of the Mangrove Species Rhizophora mucronata. Ann. Bot. 2006, 98, 1321–1330. [Google Scholar] [CrossRef] [PubMed]
- Alpert, P.; Simms, E.L. The relative advantages of plasticity and fixity in different environments: When is it good for a plant to adjust? Evol. Ecol. 2002, 16, 285–297. [Google Scholar] [CrossRef]
- Hayward, H.E.; Long, E.M. Anatomical and Physiological Responses of the Tomato to Varying Concentrations of Sodium Chloride Sodium Sulphate, and Nutrient Solutions. Bot. Gaz. 1941, 102, 437–462. [Google Scholar] [CrossRef]
- Boughalleb, F.; Denden, M.; Ben Tiba, B. Anatomical changes induced by increasing NaCl salinity in three fodder shrubs, Nitraria retusa, Atriplex halimus and Medicago arborea. Acta Physiol. Plant. 2009, 31, 947–960. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M.; Francois, L.E. Whole - plant response to salinity. In Plant-Environment Interactions; Wilkinson, R.E., Ed.; Marcel Dekker, Inc.: New York, NY, USA, 1994; pp. 199–244. [Google Scholar]
- Tomlinson, P.B. The Botany of Mangroves, 1st ed.; Cambridge University Press: New York, NY, USA, 1994. [Google Scholar]
- Sidhu, S.S. Structure of epidermis and stomatal apparatus of mangrove species. In Proceedings of the International Symposium on Biology and Management of Mangroves; Institute of Food and Agricultural Sciences, University of Florida: Gainesville, FL, USA, 1975; pp. 569–578. [Google Scholar]
- Parida, A.; Das, A.; Mittra, B. Effects of NaCl Stress on the Structure, Pigment Complex Composition, and Photosynthetic Activity of Mangrove Bruguiera parviflora Chloroplasts. Photosynthetica 2003, 41, 191–200. [Google Scholar] [CrossRef]
- Parida, A.; Das, A.B.; Mittra, B.N. Effects of salt on growth, ion accumulation, photosynthesis and leaf anatomy of the mangrove, Bruguiera parviflora. Trees 2004, 18, 167–174. [Google Scholar] [CrossRef]
- Saenger, P. Morphological, anatomical and reproductive adaptations of Australian mangroves. In Mangrove Ecosystems in Australia: Structure, Function and Management; Australia National University Press: Acton, Australia, 1982. [Google Scholar]
- Das, S. An adaptive feature of some mangroves of Sundarbans, West Bengal. J. Plant Biol. 1999, 42, 109–116. [Google Scholar] [CrossRef]
- Waisel, Y. Biology of Halophytes; Academic Press: New York, NY, USA, 1972. [Google Scholar]
- Poljakoff-Mayber, A. Morphological and anatomical changes in plants as a response to salinity stress. In Plants in Saline Environment; Springer: Heidelberg/Berlin, Germany, 1975; pp. 97–117. [Google Scholar] [CrossRef]
- Ball, M.; Farquhar, G.D. Photosynthetic and Stomatal Responses of Two Mangrove Species, Aegiceras corniculatum and Avicennia marina, to Long Term Salinity and Humidity Conditions. Plant Physiol. 1984, 74, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aziz, I.; Khan, M.A. Effect of Seawater on the Growth, Ion Content and Water Potential of Rhizophora mucronata Lam. J. Plant Res. 2001, 114, 369–373. [Google Scholar] [CrossRef]
- Hoppe-Speer, S.C.; Adams, J.; Rajkaran, A.; Bailey, D. The response of the red mangrove Rhizophora mucronata Lam. to salinity and inundation in South Africa. Aquat. Bot. 2011, 95, 71–76. [Google Scholar] [CrossRef]
- Valladares, F.; Gianoli, E.; Gómez, J.M. Ecological limits to plant phenotypic plasticity. New Phytol. 2007, 176, 749–763. [Google Scholar] [CrossRef] [PubMed]
- Poveda, G.; Mesa, Ó.J. Las fases extremas del fenómeno ENSO (El Niño y La Niña) y su influencia sobre la hidrología de Colombia. Tecnol. Cienc. Agua 1996, 11, 21–37. [Google Scholar]
- NOAA National Weather Service C for Climate Prediction. Historical El Niño/La Niña Episodes (1950-Present). 2021. Available online: https://origin.cpc.ncep.noaa.gov/products/analysis_monitoring/ensostuff/ONI_v5.php (accessed on 31 May 2021).
- Moreno, M.M.; del Valle, J.I. Influence of local climate and ENSO on the growth of Abarco (Cariniana pyriformis) in Chocó, Colombia. Trees 2015, 29, 97–107. [Google Scholar] [CrossRef]
- Blanco, J.A.; Viloria, E.A. ENSO and salinity changes in the Ciénaga Grande de Santa Marta coastal lagoon system, Colombian Caribbean. Estuarine. Coast. Shelf Sci. 2006, 66, 157–167. [Google Scholar] [CrossRef]
- Jennerjahn, T.C.; Gilman, E.; Krauss, K.W.; Lacerda, L.D.; Nordhaus, I.; Wolanski, E. Mangrove ecosystems under climate change. In Mangrove Ecosystems: A Global Biogeographic Perspective; Springer: Cham, Switzerland, 2017; pp. 211–244. [Google Scholar]
- Von Prahl, H.; Contreras, R. Manglares y Hombres del Pacífico; Fondo FEN Colombia. Editorial Presencia: Bogotá, Colombia, 1990; p. 193. [Google Scholar]
- Qin, D.; Plattner, G.K.; Tignor, M.; Allen, S.K.; Boschung, J.; Nauels, A.; Midgley, P.M. Climate Change 2013: The Physical Science Basis. Contribution of Working Group I to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (eds TF Stocker et al.); Cambridge University Press: New York, NY, USA, 2014; pp. 5–14. [Google Scholar]
- Lovelock, C.E.; Feller, I.C. Photosynthetic performance and resource utilization of two mangrove species coexisting in a hypersaline scrub forest. Oecologia 2003, 134, 455–462. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cardona, P.; Botero, L. Soil Characteristics and Vegetation Structure in a Heavily Deteriorated Mangrove Forest in the Caribbean Coast of Colombia. Biotropica 1998, 30, 24–34. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| KW | Juveniles | Adults | All Groups | |||
|---|---|---|---|---|---|---|
| Tissue | X2 | p | X2 | p | X2 | p |
| Leaf Area | 14.77 | <0.001 * | 7.49 | 0.024 * | 37.61 | <0.001 * |
| Leaf Thickness | 34.9 | <0.001 * | 0.82 | 0.0663 | 44.1 | <0.001 * |
| Adaxial Cuticle | 56.78 | <0.001 * | 9.86 | <0.001 * | 82.99 | <0.001 * |
| Adaxial Epidermis | 20.04 | <0.001 * | 9.57 | <0.001 * | 30.01 | <0.001 * |
| Hypodermis | 50.295 | <0.001 * | 1.18 | 0.554 | 58.12 | <0.001 * |
| Palisade Parenchyma | 62.43 | <0.001 * | 0.22 | 0.897 | 87.05 | <0.001 * |
| Spongy Parenchyma | 21.47 | <0.001 * | 3.91 | 0.141 | 57.88 | <0.001 * |
| Abaxial Epidermis | 21.63 | <0.001 * | 1.15 | 0.562 | 21.95 | <0.001 * |
| Abaxial Cuticle | 6.22 | 0.045 * | 0.52 | 0.771 | 17.43 | <0.001 * |
| Diameter Lumen Xylem Vessels | 42.33 | <0.001 * | 15.23 | <0.001 * | 70.54 | <0.001 * |
| Wall Xylem Vessels | 22.88 | <0.001 * | 23.66 | <0.001 * | 95.91 | <0.001 * |
| Group | Functional Traits | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Foliar Leaf | |||||||||||
| Leaf Area | Adaxial Cuticle | Adaxial Epidermis | Hypodermis | Palisade Parenchyma | Spongy Parenchyma | Abaxial Epidermis | Abaxial Cuticle | Leaf Thickness | Xylem Lumen Diameter | Xylem Vessel Thickness | |
| Juveniles | |||||||||||
| SC + JV | −0.478 | 0.065 | 0.321 | 0.369 | 0.068 | 0.207 | 0.283 | −0.041 | 0.385 | 0.417 | 0.519 |
| SB + JV | −0.696 | 0.354 | 0.334 | 0.622 | 0.308 | 0.215 | 0.397 | 0.099 | 0.591 | −0.409 | 0.734 |
| OP + JV | 0.065 | 0.573 | 0.609 | 0.756 | 0.596 | 0.594 | 0.552 | 0.339 | 0.754 | 0.644 | 0.482 |
| Adults | |||||||||||
| SC + AD | −0.674 | 0.101 | 0.121 | 0.396 | 0.180 | 0.172 | 0.223 | −0.115 | 0.276 | 0.498 | 0.331 |
| SB + AD | −0.430 | 0.277 | 0.267 | 0.371 | 0.472 | 0.309 | 0.328 | −0.073 | 0.408 | 0.538 | 0.439 |
| OP + AD | −0.701 | 0.562 | 0.550 | 0.470 | 0.597 | 0.546 | 0.600 | 0.282 | 0.507 | 0.543 | 0.462 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sánchez, A.R.; Pineda, J.E.M.; Casas, X.M.; Calderón, J.H.M. Influence of Edaphic Salinity on Leaf Morphoanatomical Functional Traits on Juvenile and Adult Trees of Red Mangrove (Rhizophora mangle): Implications with Relation to Climate Change. Forests 2021, 12, 1586. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111586
Sánchez AR, Pineda JEM, Casas XM, Calderón JHM. Influence of Edaphic Salinity on Leaf Morphoanatomical Functional Traits on Juvenile and Adult Trees of Red Mangrove (Rhizophora mangle): Implications with Relation to Climate Change. Forests. 2021; 12(11):1586. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111586
Chicago/Turabian StyleSánchez, Alejandra Robles, José Ernesto Mancera Pineda, Xavier Marquínez Casas, and Jairo Humberto Medina Calderón. 2021. "Influence of Edaphic Salinity on Leaf Morphoanatomical Functional Traits on Juvenile and Adult Trees of Red Mangrove (Rhizophora mangle): Implications with Relation to Climate Change" Forests 12, no. 11: 1586. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111586