
Land Cover and Land Use Change Decreases Net Ecosystem Production in Tropical Peatlands of West Kalimantan, Indonesia

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
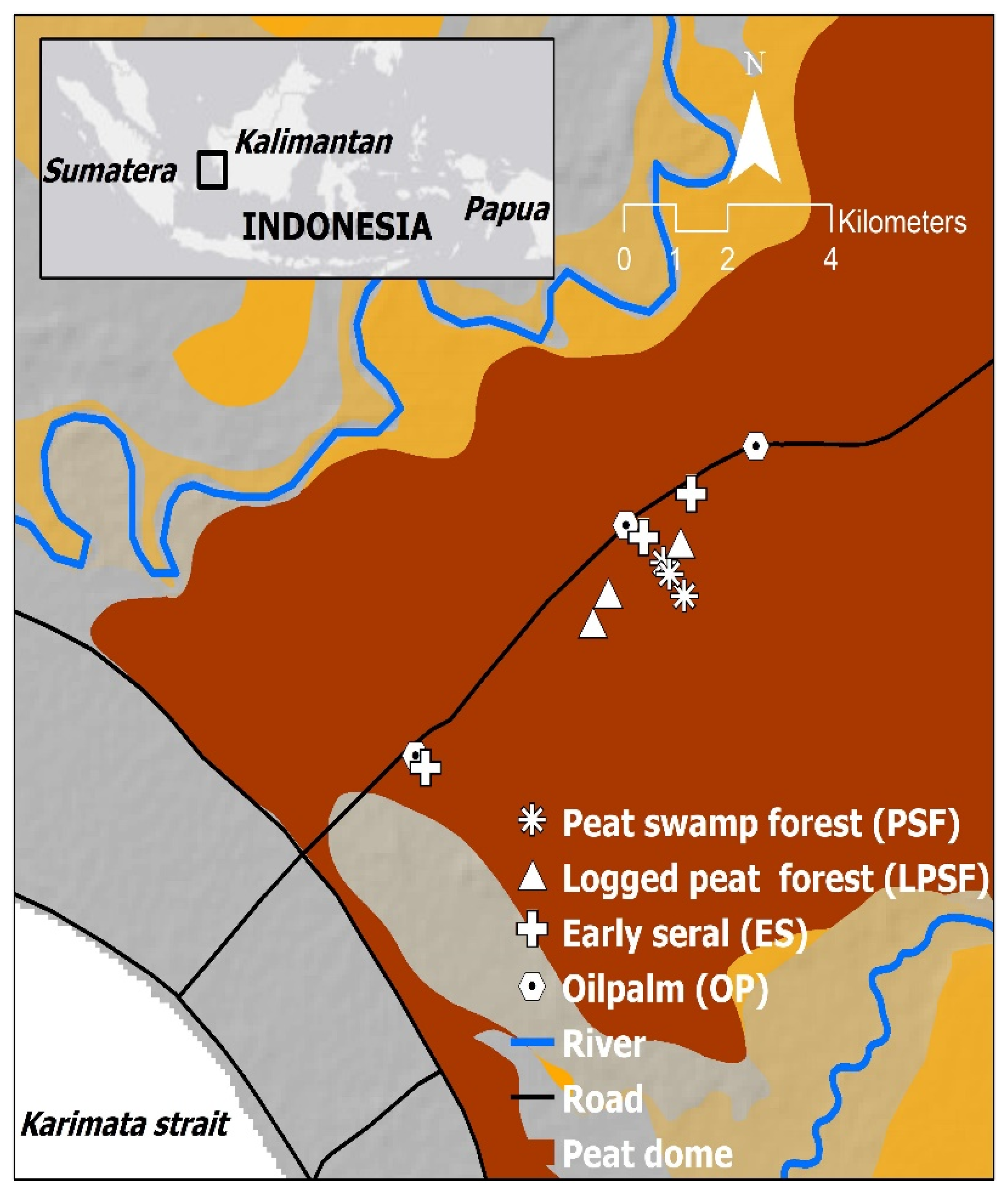
2.1. Study Site
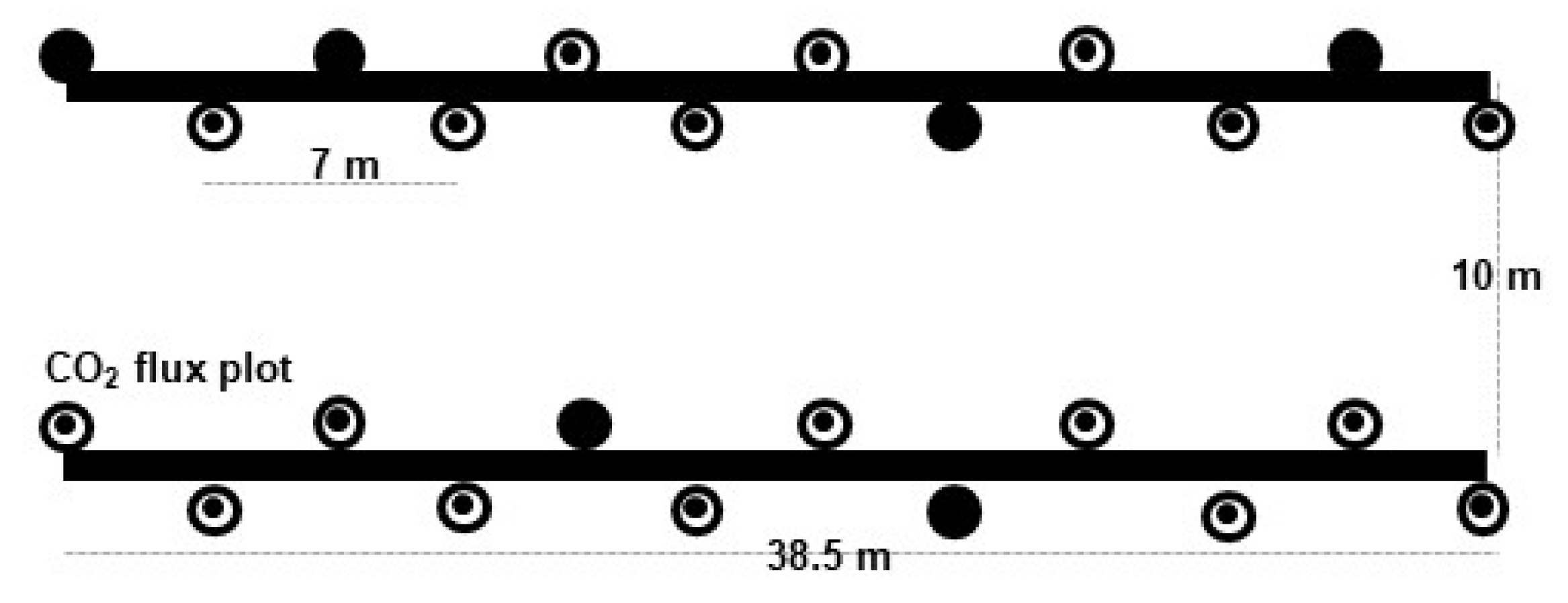
2.2. Soil Respiration: Total, Heterotrophic and Autotrophic Respiration
2.3. Net Primary Production and Net Ecosystem Production
2.4. Soil Parameters
2.5. Statistical Analyses
3. Results
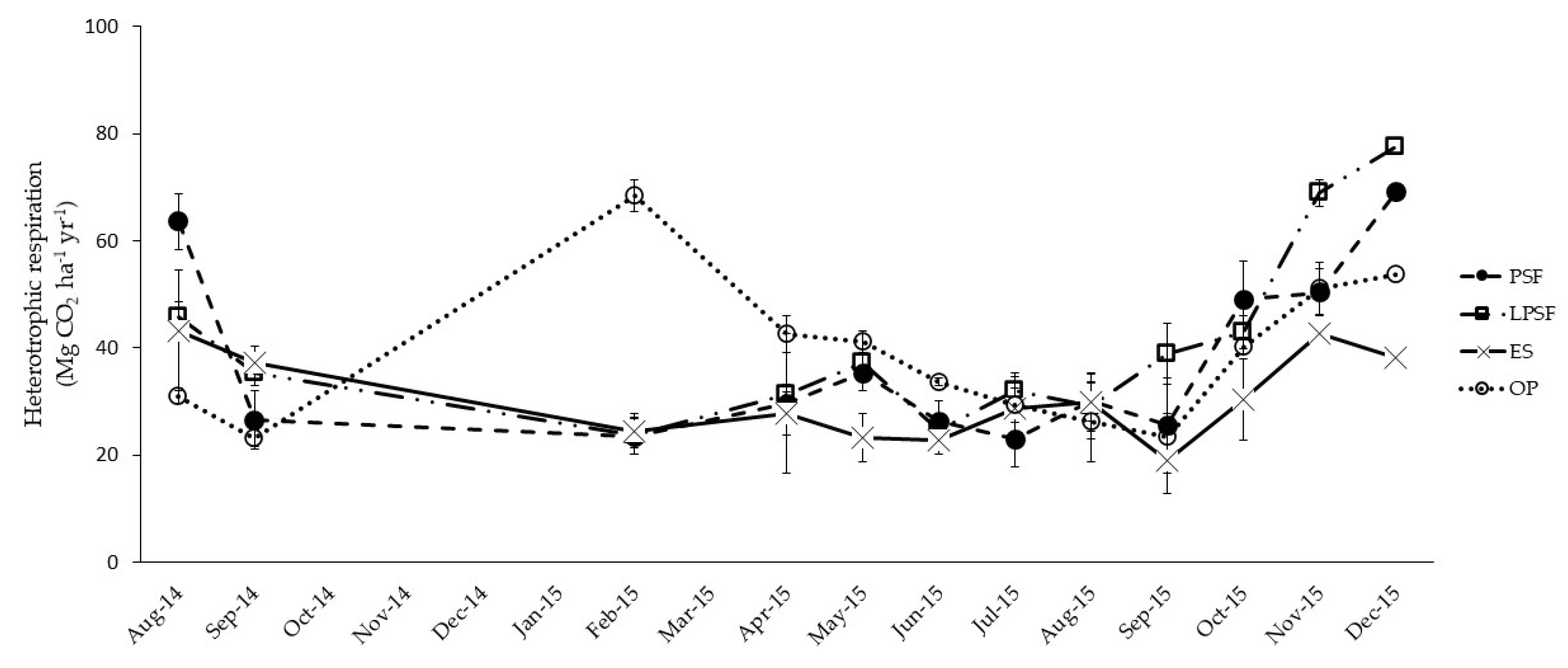
3.1. Annual Ecosystem Respiration
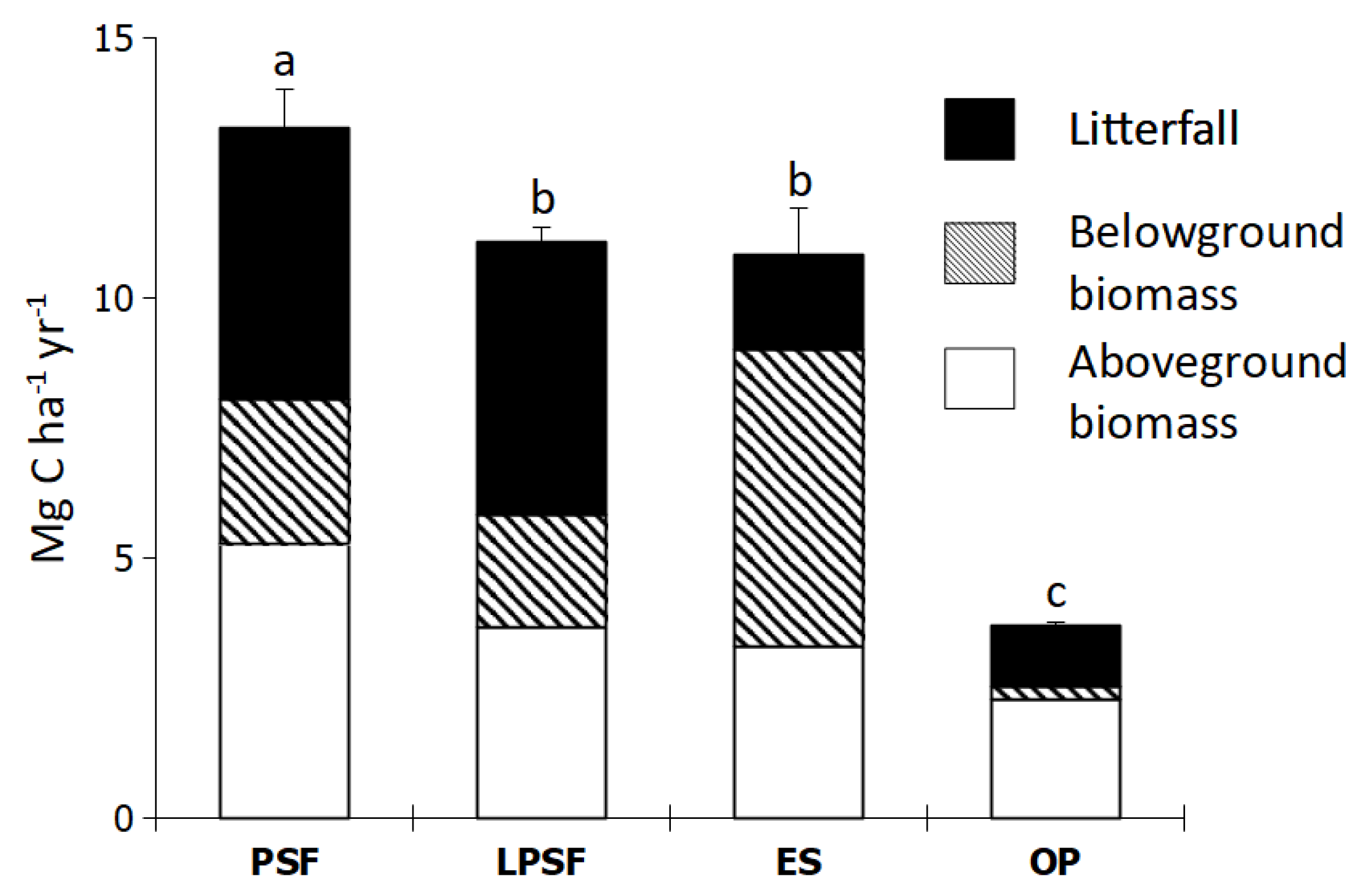
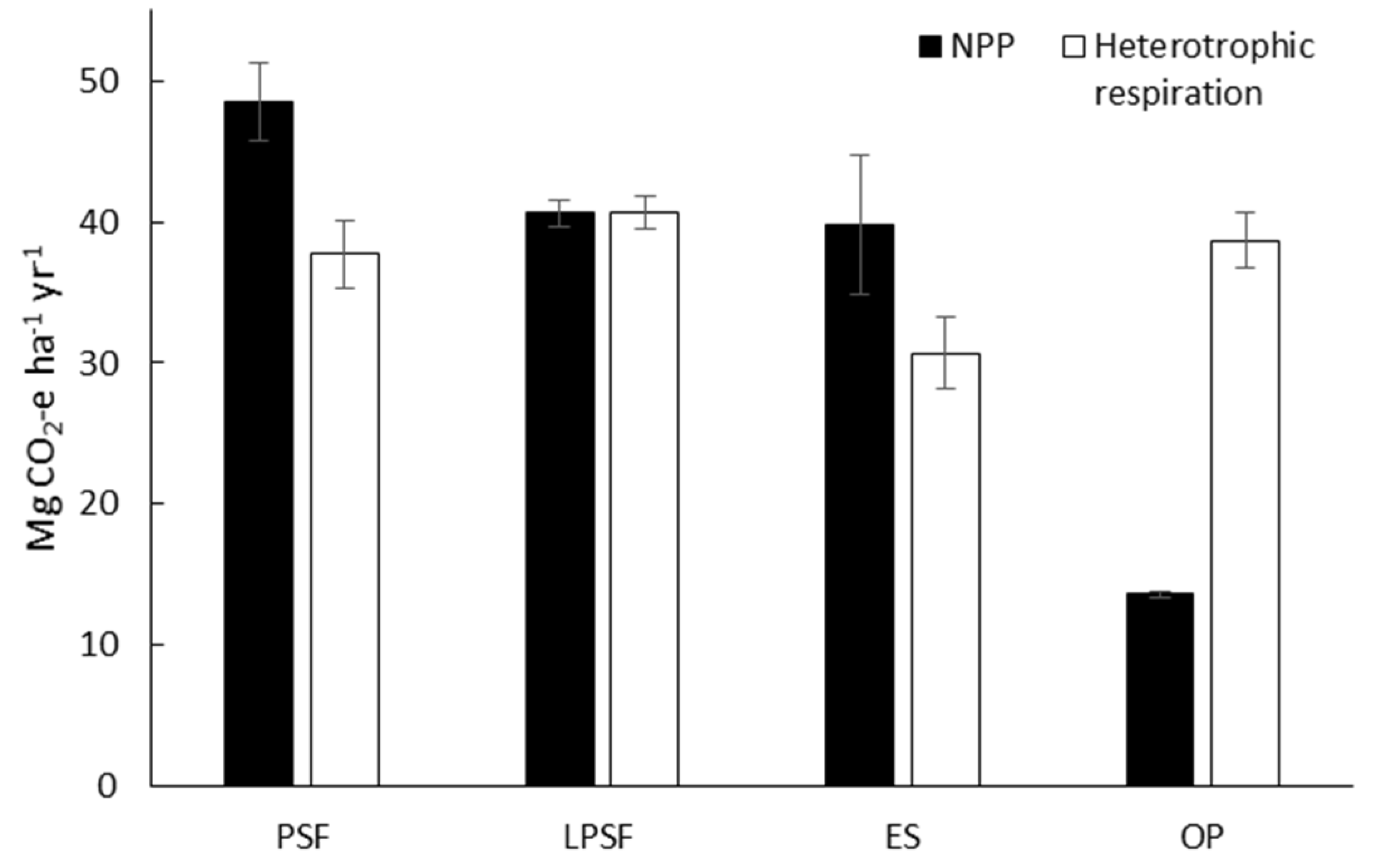
3.2. Net Primary Production in Intact and Logged Peat Swamp Forests
3.3. Net Ecosystem Production
3.4. Soil Parameters
4. Discussion
4.1. How Land Use Change Affects Soil Respiration in Tropical Peatland Ecosystems
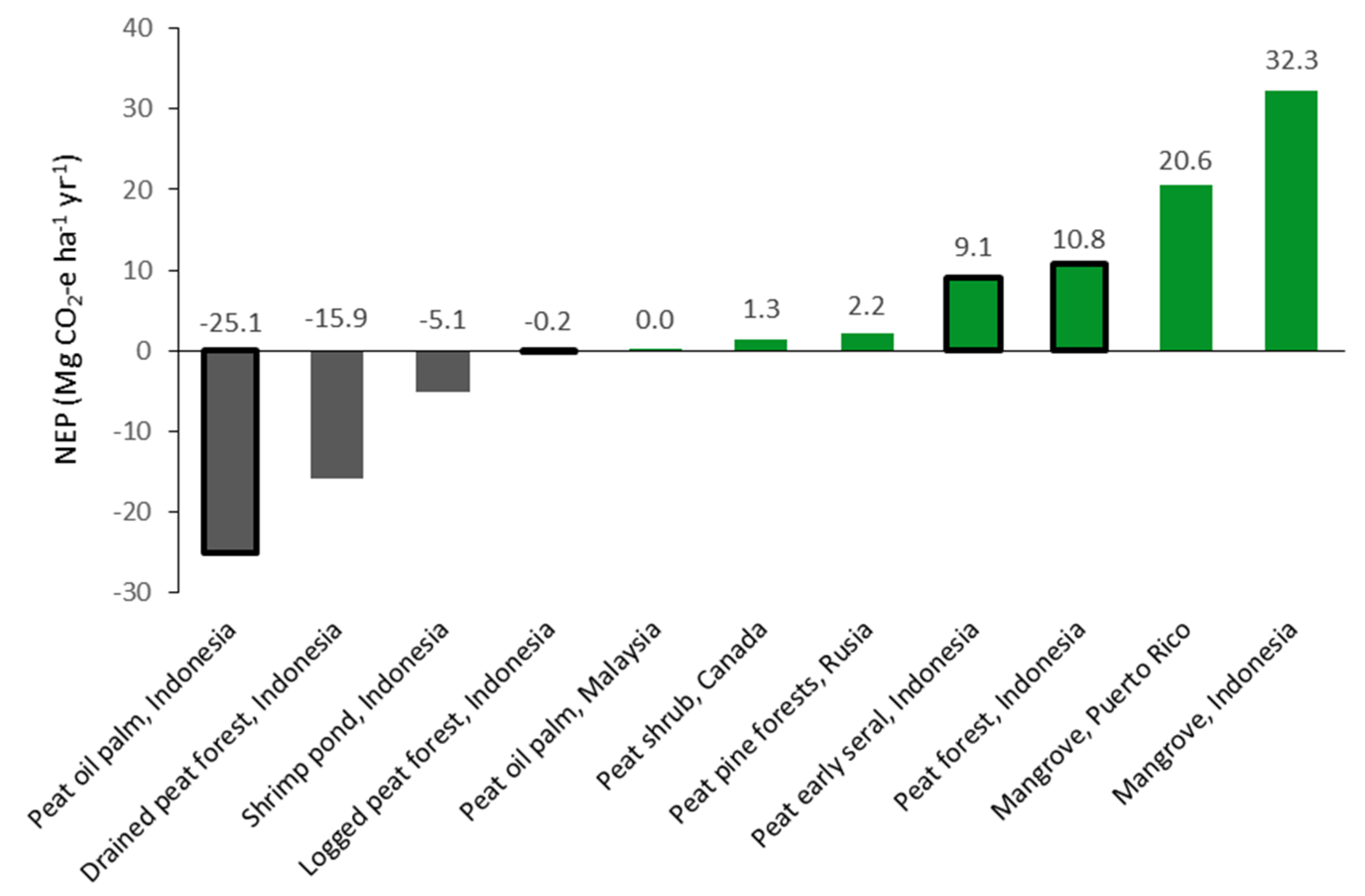
4.2. Effect of Land Use Change on Net Ecosystem Production
4.3. Implications for Tropical Peatland Management
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Murdiyarso, D.; Hergoualćh, K.; Verchot, L.V. Opportunities for reducing greenhouse gas emissions in tropical peatlands. Proc. Natl. Acad. Sci. USA 2010, 107, 19655–19660. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anda, M.; Ritung, S.; Suryani, E.; Hikmat, M.; Yatno, E.; Mulyani, A.; Subandiono, R.E. Revisiting tropical peatlands in Indonesia: Semi-detailed mapping, extent, and depth distribution assessment. Geoderma 2021, 402, 115235. [Google Scholar] [CrossRef]
- Hairiah, K.; Sitompul, S.M.; van Noordwijk, M.; Palm, C. Carbon Stocks of Tropical Land Use Systems as Part of the Global C Balance. Effects of Forest Conversion and Options for Clean Development Activities; International Centre for Research in Agroforestry, ICRAF: Bogor, Indonesia, 2001. [Google Scholar]
- Basuki, I.; Kauffman, J.B.; Murdiyarso, D.; Anshari, G. Carbon Stocks and Emissions from Degradation and Conversion of Tropical Peat Swamp Forests in West Kalimantan, Indonesia. In Proceedings of the 15th International Peat Congress, Kuching, Malaysia, 15–19 August 2016. [Google Scholar]
- Murdiyarso, D.; Donato, D.; Kauffman, J.B.; Kurnianto, S.; Stidham, M.; Kanninen, M. Carbon Storage in Mangrove and Peatland Ecosystems. A Preliminary Account from Plots in Indonesia; CIFOR: Bogor, Indonesia, 2009. [Google Scholar]
- Koh, L.P.; Miettinen, J.; Liew, S.C.; Ghazoul, J. Remotely sensed evidence of tropical peatland conversion to oil palm. Proc. Natl. Acad. Sci. USA 2011, 108, 5127–5132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miettinen, J.; Shi, C.; Liew, S.C. Land cover distribution in the peatlands of Peninsular Malaysia, Sumatera and Borneo in 2015 with changes since 1990. Glob. Ecol. Conserv. 2016, 6, 67–78. [Google Scholar] [CrossRef] [Green Version]
- Page, S.E.; Morrison, R.; Malins, C.; Hooijer, A.; Rieley, J.O.; Jauhiainen, J. Review of Peat Surface Greenhouse Gas Emissions from Oil Palm Estates in Southeast Asia; International Council on Clean Transportation: Washington, DC, USA, 2011. [Google Scholar]
- Verwer, C.; Van Der Meer, P. Carbon Pools in Tropical Peat Forest—Towards a Reference Value for Forest Biomass Carbon in Relatively Undisturbed Peat Swamp Forests in Southeast Asia; Alterra Wageningen, UR: Wageningen, The Netherlands, 2010. [Google Scholar]
- Dommain, R.; Couwenberg, J.; Glaser, P.H.; Joosten, H.; Suryadiputra, I.N.N. Carbon storage and release in Indonesian peatlands since the last deglaciation. Quat. Sci. Rev. 2014, 97, 1–32. [Google Scholar] [CrossRef]
- Anshari, G. Circularity and Singularity of Tropical Peat Swamp Forest Ecosystems. In Tropical Peatland Eco Management; Osaki, M., Tsuji, N., Foead, N., Eds.; Springer: Tokyo, Japan, 2021; pp. 463–475. [Google Scholar]
- Wösten, J.H.M.; Clymans, E.; Page, S.E.; Rieley, J.O.; Limin, S.H. Peat–water interrelationships in a tropical peatland ecosystem in Southeast Asia. Catena 2008, 73, 212–224. [Google Scholar] [CrossRef]
- Hooijer, A.; Page, S.; Jauhiainen, J.; Lee, W.A.; Lu, X.X.; Idris, A.; Anshari, G. Subsidence and carbon loss in drained tropical peatlands. Biogeosciences 2012, 9, 1053–1071. [Google Scholar] [CrossRef] [Green Version]
- Page, S.E.; Siegert, F.; Rieley, J.O.; Boehm, H.D.V.; Jaya, A.; Limin, S. The amount of carbon released from peat and forest fires in Indonesia during 1997. Nature 2002, 420, 61–65. [Google Scholar] [CrossRef]
- Basuki, I. Carbon Dynamics in Response to Land Cover Change in Tropical Peatlands, Kalimantan, Indonesia. Ph.D. Thesis, Oregon State University, Oregon, USA, 17 June 2017. [Google Scholar]
- Chapin, F.S.; Woodwell, G.M.; Randerson, J.T.; Rastetter, E.B.; Lovett, G.M.; Baldocchi, D.D.; Clark, D.A.; Harmon, M.E.; Schimel, D.S.; Valentini, R.; et al. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems 2006, 9, 1041–1050. [Google Scholar] [CrossRef] [Green Version]
- Komiyama, A.; Ong, J.E.; Poungparn, S. Allometry, biomass, and production of mangrove forests: A review. Aquat. Bot. 2008, 89, 128–137. [Google Scholar] [CrossRef]
- Clark, D.A.; Brown, S. Net primary production in tropical forests: An evaluation and synthesis of existing field data. Ecol. Appl. 2001, 11, 371–384. [Google Scholar] [CrossRef]
- Randerson, J.T.; Chapin, F.S., III; Harden, J.W.; Neff, J.C.; Harmon, M.E. Net ecosystem production: A comprehensive measure of net carbon accumulation by ecosystems. Ecol. Appl. 2002, 12, 937–947. [Google Scholar] [CrossRef]
- Woodwell, G.M.; Whittaker, R.H. Primary production in terrestrial ecosystems. Am. Zool. 1968, 8, 19–30. [Google Scholar] [CrossRef]
- Novita, N. Carbon Stocks and Soil Greenhouse Gas Emissions Associated with Forest Conversion to Oil Palm Estates in Tanjung Puting Tropical Peatlands, Indonesia. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2016. [Google Scholar]
- Chimner, R.A.; Ewel, K.C. A tropical freshwater wetland: II. Production, decomposition, and peat formation. Wetl. Ecol. Manag. 2005, 13, 671–684. [Google Scholar] [CrossRef]
- Basuki, I.; Kauffman, J.B.; Peterson, J.; Anshari, G.; Murdiyarso, D. Land cover changes reduce net primary production in tropical coastal peatlands of West Kalimantan, Indonesia. Mitig. Adapt. Strateg. Glob. Chang. 2019, 24, 557–573. [Google Scholar] [CrossRef] [Green Version]
- Melling, L.; Hatano, R.; Goh, K.J. Soil CO2 flux from three ecosystems in tropical peatland of Sarawak, Malaysia. Tellus B Chem. Phys. Meteorol. 2005, 57, 1–11. [Google Scholar] [CrossRef]
- Comeau, L.P.; Hergoualc’h, K.; Smith, J.U.; Verchot, L. Conversion of Intact Peat Swamp Forest to Oil Palm Estate: Effects on Soil CO2 Fluxes in Jambi, Sumatera; CIFOR: Bogor, Indonesia, 2013. [Google Scholar]
- Prananto, J.A.; Minasny, B.; Comeau, L.P.; Grace, P. Drainage increases CO2 and N2O emissions from tropical peat soils. Glob. Chang. Biol. 2020, 26, 4583–4600. [Google Scholar] [CrossRef] [PubMed]
- Astiani, D. Tropical peatland tree-species diversity altered by forest degradation. Biodiversitas 2016, 17, 102–109. [Google Scholar]
- Rusli, N.; Majid, M.R.; Din, A.H.M. A comparative accuracy of Google Earth height with MyGeoid, EGM96 and MSL. IOP Conf. Ser. Earth Environ. Sci. 2016, 37, 012003. [Google Scholar] [CrossRef] [Green Version]
- Carlson, K.M. Effects of Oil Palm Estate Development on Land Cover, Carbon Flux, and Streams in Indonesian Borneo. Ph.D. Thesis, Yale University, New Haven, CT, USA, 2012. [Google Scholar]
- Ritung, S.; Wahyunto; Nugroho, K.; Sukarman; Hikmatullah; Suparto; Tafakresnanto, C. Peta Lahan Gambut Indonesia; Ministry of Agriculture, Balai Besar Penelitian dan Pengembangan Sumberdaya Lahan Pertanian: Bogor, Indonesia, 2011.
- Hoscilo, A. Fire Regime, Vegetation Dynamics and Land Cover Change in Tropical Peatland, Indonesia. Ph.D. Thesis, University of Leicester, Leicester, UK, 2009. [Google Scholar]
- Bond, W.J.; Keane, R.E. Fires, Ecological Effects of. Ref. Modul. Life Sci. 2017, 1–11. [Google Scholar] [CrossRef]
- Hanson, P.J.; Edwards, N.T.; Garten, C.T.; Andrews, J.A. Separating root and soil microbial contributions to soil respiration: A review of methods and observations. Biogeochemistry 2000, 48, 115–146. [Google Scholar] [CrossRef]
- Jassal, R.S.; Black, T.A. Estimating heterotrophic and autotro-phic soil respiration using small-area trenched plot technique:theory and practice. Agric. For. Meteorol. 2006, 140, 193–202. [Google Scholar] [CrossRef]
- Jauhiainen, J.; Hidenori, T.; Juha, E.P.H.; Pertti, J.M.; Harri, V. Carbon fluxes from a tropical peat swamp forest floor. Glob. Chang. Biol. 2005, 11, 1788–1797. [Google Scholar] [CrossRef]
- Kauffman, J.B.; Virni, B.A.; Imam, B.; Sofyan, K.; Nisa, N.; Daniel, M.; Daniel, C.D.; Matthew, W.W. Protocols for the Measurement, Monitoring, and Reporting of Structure, Biomass, Carbon Stocks and Greenhouse Gas Emissions in Tropical Peat Swamp Forests; Center for International Forestry Research: Bogor, Indonesia, 2016. [Google Scholar]
- Moser, G.; Bernhard, S.; Dietrich, H.; Viviana, H.; Heinz, C.; Henry, B.; Christoph, L. Replicated throughfall exclusion experiment in an Indonesian perhumid rainforest: Wood production, litter fall and fine root growth under simulated drought. Glob. Chang. Biol. 2014, 20, 1481–1497. [Google Scholar] [CrossRef] [PubMed]
- Mokany, K.; Raison, R.; Prokushkin, A. Critical analysis of root: Shoot ratios in terrestrial biomes. Glob. Chang. Biol. 2006, 12, 84–96. [Google Scholar] [CrossRef]
- Dewi, S.; Khasanah, N.; Rahayu, S.; Ekadinata, A.; van Noordwijk, M. Carbon Footprint of Indonesian Palm Oil Production: A Pilot Study; World Agroforestry Center: Bogor, Indonesia, 2010; Available online: http://www.worldagroforestry.org/sea/Publications/files/poster/PO0236-10.PDF (accessed on 5 November 2021).
- Henson, I.E.; Dolmat, M.T. Physiological analysis of an oil palm density trial on a peat soil. J. Oil Palm. Res. 2003, 15, 1–27. [Google Scholar]
- Scurlock, J.; Olson, R. NPP Multi-Biome: Grassland, Boreal Forest, and Tropical Forest Sites, 1939–1996, [Revision] 1. Dataset, 2012. From Oak Ridge National Laboratory Distributed Active Archive Center, Oak Ridge, Tennessee. Available online: https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=653 (accessed on 5 November 2021).
- Hirano, T.; Kusin, K.; Limin, S.; Osaki, M. Carbon dioxide emissions through oxidative peat decomposition on a burnt tropical peatland. Glob. Chang. Biol. 2014, 20, 555–565. [Google Scholar] [CrossRef]
- Hergoualc’h, K.; Verchot, L.V. Stocks and fluxes of carbon associated with land use change in Southeast Asian tropical peatlands: A review. Glob. Biogeochem. Cycles 2011, 25, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Husnain, H.; Wigena, I.G.P.; Dariah, A.; Marwanto, S.; Setyanto, P.; Agus, F. CO2 emissions from tropical drained peat in Sumatera, Indonesia. Mitig. Adapt. Strateg. Glob. Chang. 2014, 19, 845–862. [Google Scholar] [CrossRef]
- Zogg, G.P.; Zak, D.R.; Ringelberg, D.B.; Macdonald, N.W.; Pregitzer, K.S.; White, D.C. Compositional and Functional Shifts in Microbial Communities Due to Soil Warming. Soil Sci. Soc. Am. J. 1997, 61, 475–481. [Google Scholar] [CrossRef] [Green Version]
- Jaatinen, K.; Laiho, R.; Vuorenmaa, A.; Del Castillo, U.; Minkkinen, K.; Pennanen, T.; Penttilä, T.; Fritze, H. Responses of aerobic microbial communities and soil respiration to water-level drawdown in a northern boreal fen. Environ. Microbiol. 2008, 10, 339–353. [Google Scholar] [CrossRef] [PubMed]
- Novita, N.; Nurul, S.L.; Mega, L.; Tatang, T.; Imam, B.; Joni, J. Geographic Setting and Groundwater Table Control Carbon Emission from Indonesian Peatland: A Meta-Analysis. Forests 2021, 12, 832. [Google Scholar] [CrossRef]
- Valentini, R.; Matteucci, G.; Dolman, A.J.; Schulze, E.D.; Rebmann, C.; Moors, E.J.; Granier, A.; Gross, P.; Jensen, N.O.; Pilegaard, K.; et al. Respiration as the main determinant of carbon balance in European forests. Nature 2000, 404, 861–865. [Google Scholar] [CrossRef] [PubMed]
- Alongi, D.; Tirendi, F.; Clough, B. Belowground decomposition of organic matter in forests of the mangroves Rhizophora stylosa and Avicennia marina along the arid coast of Western Australia. Aquat. Bot. 2000, 68, 97–122. [Google Scholar] [CrossRef]
- Golley, F.; Odum, H.T.; Wilson, R.F. The Structure and Metabolism of a Puerto Rican Red Mangrove Forest in May. Ecology 1962, 43, 9–19. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M.; Wattayakorn, G.; Pfitzner, J.; Tirendi, F.; Zagorskis, I.; Brunskill, G.J.; Davidson, A.; Clough, B.F. Organic carbon accumulation and metabolic pathways in sediments of mangrove forests in southern Thailand. Mar. Geol. 2001, 179, 85–103. [Google Scholar] [CrossRef]
- Alongi, D.M.; Mukhopadhyay, S.K. Contribution of mangroves to coastal carbon cycling in low latitude seas. Agric. For. Meteorol. 2015, 213, 266–272. [Google Scholar] [CrossRef]
- Arifanti, V.B. Influences of Land Use on Carbon Cycles in Mangrove Ecosystems of the Mahakam Delta, East Kalimantan, Indonesia. Ph.D. Thesis, Oregon State University, Corvallis, OR, USA, 2017. [Google Scholar]
- Schulze, E.D.; Prokuschkin, A.; Arneth, A.; Knorre, N.; Vaganov, E.A. Net ecosystem production and peat accumulation in a Siberian Aapa mire. Tellus B Chem. Phys. Meteorol. 2002, 54, 531–536. [Google Scholar] [CrossRef]
- Waring, R.H.; Law, B.; Bond, B. NPP Temperate Forest: OTTER Project Sites, Oregon, USA, 1989–1991. 2013. Available online: https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=472 (accessed on 5 November 2021).
- Melling, L.; Kah, J.G.; Beauvais, C.; Hatano, R. Carbon Flow and Budget in a Young Mature Oil Palm Agroecosystem on Deep Tropical Peat. Plant 2008, 84, 21–25. [Google Scholar]
- Hiraishi, T.; Krug, T.; Tanabe, K.; Srivastava, N.; Baasansuren, J.; Fukuda, M.; Troxler, T.G. 2013 Supplement to the 2006 IPCC Guidelines for National Greenhouse Gas Inventories: Wetlands; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Hirano, T.; Segah, H.; Harada, T.; Limin, S.; June, T.; Hirata, R.; Osaki, M. Carbon dioxide balance of a tropical peat swamp forest in Kalimantan, Indonesia. Glob. Chang. Biol. 2007, 13, 412–425. [Google Scholar] [CrossRef]
- Bubier, J.L.; Frolking, S.; Crill, P.M.; Linder, E. Net ecosystem production and its uncertainty in a diverse boreal peatland. J. Geophys. Res. Atmos. 1999, 104, 27683–27692. [Google Scholar] [CrossRef]
- Uhl, C.; Kauffman, J.B. Deforestation effects on fire susceptibility and the potential response of the tree species to fire in the rainforest of the eastern Amazon. Ecology 1990, 71, 437–449. [Google Scholar] [CrossRef]
- Usup, A.; Hashimoto, Y.; Takahashi, H.; Hayasaka, H. Combustion and thermal characteristics of peat fire in tropical peatland in Central Kalimantan, Indonesia. Tropics 2004, 14, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Konecny, K.; Ballhorn, U.W.E.; Navratil, P.; Jubanski, J.; Page, S.E.; Tansey, K.; Siegert, F. Variable carbon losses from recurrent fires in drained tropical peatlands. Glob. Chang. Biol. 2016, 22, 1469–1480. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Year | Pre-1988 | 1988 | 1994 | 2010 |
|---|---|---|---|---|
| Process | Subsistent use | Road and logging | Clearing/deforestation | Clearing + burning |
| Land use/land cover | Intact peat swamp forests (PSF) | Logged peat swamp forests (LPSF) | Early seral (ES) | Smallholder and industrial oil palm estate (OP) |
| Photos |  |  |  |  |
| Scheme | Heterotrophic Respiration | Autotrophic Respiration | Total Soil Respiration | Water Table Depth | Soil Temperature |
|---|---|---|---|---|---|
| -----------------Mg CO2 ha−1 yr−1--------------- | cm | °C | |||
| Peat swamp forest 1 | 40.0 ± 5.8 | 5.9 ± 2.9 | 45.9 ± 5.5 | 44.3 ± 10.4 | 27.1 ± 0.1 |
| Peat swamp forest 2 | 40.1 ± 5.3 | 13.9 ± 4.1 | 54.0 ± 5.7 | 48.4 ± 10.4 | 27.3 ± 0.2 |
| Peat swamp forest 3 | 32.9 ± 4.5 | 12.8 ± 2.3 | 45.6 ± 4.3 | 45.3 ± 10.1 | 27.2 ± 0.3 |
| Peat swamp forest mean (n = 3) | 37.7 ± 2.4 a | 10.8 ± 2.5 a | 48.5 ± 2.7 a | 46 ± 20.1 a | 27.2 ± 0.4 a |
| Logged peat swamp forest 1 | 43.0 ± 5.4 | 10.9 ± 4.6 | 53.9 ± 5.3 | 39.9 ± 10.5 | 27 ± 0.2 |
| Logged peat swamp forest 2 | 39.6 ± 5.1 | 6.1 ± 4.2 | 45.6 ± 4.9 | 34.0 ± 9.9 | 26.8 ± 0.2 |
| Logged peat swamp forest 3 | 39.4 ± 4.3 | 11.6 ± 3.4 | 51.0 ± 5.4 | 45.1 ± 10.6 | 27.3 ± 0.3 |
| Logged peat swamp forest mean (n = 3) | 40.7 ± 1.2 a | 9.5 ± 1.7 a | 50.2 ± 2.4 a | 39.7 ± 20.3 a | 27 ± 0.5 a |
| Early seral 1 | 26.0 ± 2.5 | 14.9 ± 4.1 | 39.6 ± 4.3 | 59.8 ± 13 | 30.5 ± 0.5 |
| Early seral 2 | 31.3 ± 4.0 | 6.1 ± 1.2 | 37.5 ± 4.2 | 40.4 ± 12.5 | 29.1 ± 0.3 |
| Early seral 3 | 34.6 ± 2.0 | 10.7 ± 5.8 | 45.3 ± 5.8 | 50.7 ± 9.9 | 28.9 ± 0.3 |
| Early seral mean (n = 3) | 30.7 ± 2.5 b | 10.6 ± 2.5 a | 40.8 ± 2.3 b | 50.3 ± 23.5 a | 29.5 ± 0.8 b |
| Oil palm estate 1 | 42.2 ± 7.0 | 8.6 ± 5.4 | 49.3 ± 4.3 | 88.4 ± 10.3 | 31.5 ± 0.3 |
| Oil palm estate 2 | 38.5 ± 3.3 | 5.9 ± 3.2 | 44.4 ± 3.5 | 74.4 ± 9.1 | 30 ± 0.3 |
| Oil palm estate 3 | 35.4 ± 3.6 | 13.3 ± 6.0 | 48.7 ± 4.9 | 72.2 ± 9.3 | 30 ± 0.4 |
| Oil palm estate mean (n = 3) | 38.7 ± 2.0 a | 9.3 ± 2.2 a | 47.5 ± 1.6 a | 78.3 ± 19.1 b | 30.5 ± 0.8 b |
| Site | Soil Respiration | |||||
|---|---|---|---|---|---|---|
| Dry Months (August to October) | Wet Months (November to July) | |||||
| Heterotrophic | Autotrophic | Total | Heterotrophic | Autotrophic | Total | |
| ----------------------------------Mg CO2 ha−1 yr−1----------------------------------- | ||||||
| Peat swamp forest 1 | 41.2 ± 6.3 | 9.9 ± 4.2 | 51.1 ± 6.5 | 39.2 ± 5.9 | 3.0 ± 1.1 | 42.2 ± 5.0 |
| Peat swamp forest 2 | 40.2 ± 5.3 | 18.9 ± 5.1 | 59.2 ± 7.4 | 40.1 ± 5.6 | 10.2 ± 3.2 | 50.3 ± 4.6 |
| Peat swamp forest 3 | 35.5 ± 4.2 | 14.5 ± 3.5 | 50.0 ± 3.5 | 31.0 ± 4.9 | 11.5 ± 1.2 | 42.5 ± 4.9 |
| Peat swamp forest mean | 39.0 ± 10.1 | 14.4 ± 8.3 | 53.4 ± 11.5 | 36.7 ± 10.8 | 8.3 ± 4.5 | 45.0 ± 9.4 |
| Logged peat swamp forest 1 | 42.4 ± 2.9 | 17.9 ± 6.3 | 60.2 ± 5.3 | 43.5 ± 7.0 | 5.9 ± 2.5 | 49.4 ± 5.3 |
| Logged peat swamp forest 2 | 36.7 ± 2.4 | 14.3 ± 4.5 | 51.0 ± 5.7 | 41.6 ± 6.6 | 0.2 ± 3.2 | 41.8 ± 4.4 |
| Logged peat swamp forest 3 | 36.2 ± 2.1 | 16.8 ± 5.0 | 53.0 ± 6.8 | 41.7 ± 5.4 | 7.9 ± 1.1 | 49.5 ± 4.7 |
| Logged peat swamp forest mean | 38.4 ± 4.9 | 16.3 ± 9.9 | 54.7 ± 11.3 | 42.2 ± 12.1 | 4.7 ± 5.0 | 46.9 ± 9.4 |
| Early seral 1 | 27.5 ± 3.3 | 13.6 ± 6.2 | 37.9 ± 5.8 | 25.0 ± 2.1 | 15.9 ± 2.1 | 40.9 ± 3.4 |
| Early seral 2 | 30.8 ± 4.8 | 8.4 ± 1.2 | 39.2 ± 4.9 | 31.8 ± 3.6 | 4.5 ± 1.1 | 36.2 ± 4.1 |
| Early seral 3 | 37.6 ± 2.1 | 15.8 ± 9.1 | 53.4 ± 8.3 | 32.4 ± 1.9 | 7.0 ± 2.0 | 39.4 ± 2.7 |
| Early seral mean | 31.9 ± 7.1 | 12.6 ± 12 | 43.5 ± 12.7 | 29.7 ± 5.4 | 9.1 ± 4.5 | 38.8 ± 6.6 |
| Oil palm estate 1 | 30.1 ± 2.1 | 18.7 ± 4.3 | 45.3 ± 3.6 | 50.8 ± 8.3 | 1.4 ± 5.4 | 52.2 ± 4.8 |
| Oil palm estate 2 | 31.1 ± 3.0 | 10.4 ± 4.3 | 41.5 ± 4.7 | 43.8 ± 2.6 | 2.7 ± 2.0 | 46.5 ± 2.5 |
| Oil palm estate 3 | 25.3 ± 2.9 | 24.4 ± 7.0 | 49.7 ± 7.8 | 42.6 ± 2.5 | 5.4 ± 4.2 | 48.0 ± 2.1 |
| Oil palm estate mean | 28.8 ± 5.2 | 17.8 ± 10.5 | 45.5 ± 10.6 | 45.7 ± 10.1 | 3.2 ± 7.9 | 48.9 ± 6.5 |
| Site | Total NPP | Heterotrophic Respiration | NEP |
|---|---|---|---|
| -----------------Mg CO2 ha−1 yr−1------------------- | |||
| Peat swamp forest 1 | 50.4 | 40.0 ± 5.8 | 10.4 |
| Peat swamp forest 2 | 42.4 | 40.1 ± 5.3 | 2.3 |
| Peat swamp forest 3 | 52.8 | 32.9 ± 4.5 | 19.9 |
| Peat swamp forest mean | 48.5 ± 2.8 | 37.7 ± 2.4 | 10.8 ± 5.1 |
| Logged peat swamp forest 1 | 38.8 | 43.0 ± 5.4 | −4.2 |
| Logged peat swamp forest 2 | 40.7 | 39.6 ± 5.1 | 1.1 |
| Logged peat swamp forest 3 | 42.2 | 39.4 ± 4.3 | 2.8 |
| Logged peat swamp forest mean | 40.6 ± 1.0 | 40.7 ± 1.2 | −0.1 ± 2.1 |
| Early seral 1 | 31.3 ± 12.8 | 26 ± 2.5 | 5.3 |
| Early seral 2 | 48.1 ± 19.7 | 31.3 ± 4.0 | 16.8 |
| Early seral 3 | 39.9 ± 16.3 | 34.6 ± 2.0 | 5.3 |
| Early seral mean | 39.8 ± 4.9 | 30.7 ± 2.5 | 9.1 ± 3.8 |
| Oil palm estate 1 | 14.1 ± 1.4 | 42.2 ± 7.0 | −28.1 |
| Oil palm estate 2 | 13.4 ± 3.5 | 38.5 ± 3.3 | −25.1 |
| Oil palm estate 3 | 13.4 ± 1.0 | 35.4 ± 3.6 | −22.0 |
| Oil palm estate mean | 13.6 ± 0.2 | 38.7 ± 2.0 | −25.1 ± 1.8 |
| Site | Water Table Depth Level (cm) | Soil Temperature (°C) | ||||
|---|---|---|---|---|---|---|
| Dry Months | Wet Months | Annual | Dry Months | Wet Months | Annual | |
| Peat swamp forest | 81 ± 13 | 21 ± 8 | 46 ± 6 a | 27.3 ± 0.5 | 27.1 ± 0.3 | 27.2 ± 0.1 a |
| Logged peat swamp forest | 74 ± 13 | 15 ± 9 | 40 ± 6 a | 27.0 ± 0.5 | 27.1 ± 0.6 | 27.0 ± 0.1 a |
| Early seral | 84 ± 21 | 26 ± 13 | 50 ± 7 a | 30.0 ± 0.8 | 28.8 ± 0.8 | 29.5 ± 0.2 b |
| Oil palm estate | 105 ± 15 | 60 ± 13 | 78 ± 6 b | 30.6 ± 0.8 | 30.3 ± 0.8 | 30.5 ± 1.6 c |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Basuki, I.; Kauffman, J.B.; Peterson, J.T.; Anshari, G.Z.; Murdiyarso, D. Land Cover and Land Use Change Decreases Net Ecosystem Production in Tropical Peatlands of West Kalimantan, Indonesia. Forests 2021, 12, 1587. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111587
Basuki I, Kauffman JB, Peterson JT, Anshari GZ, Murdiyarso D. Land Cover and Land Use Change Decreases Net Ecosystem Production in Tropical Peatlands of West Kalimantan, Indonesia. Forests. 2021; 12(11):1587. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111587
Chicago/Turabian StyleBasuki, Imam, J. Boone Kauffman, James T. Peterson, Gusti Z. Anshari, and Daniel Murdiyarso. 2021. "Land Cover and Land Use Change Decreases Net Ecosystem Production in Tropical Peatlands of West Kalimantan, Indonesia" Forests 12, no. 11: 1587. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111587