1. Introduction
Indonesia is a tropical country that offers a suitable environment for a huge number of termite species. Of the 3106 species of termite that have been recorded worldwide, 300 species (11.5% of them) were discovered in this country [
1,
2]. In this circumstance, the Indo-Malayan termite
Macrotermes gilvus Hagen (Isoptera: Termitidae) has the most extensive geographical distribution out of the termite species in the country [
3]. The existence of this termite species has been reported in almost all parts of the country, including all districts of Jakarta, the capital city of Indonesia [
2,
4]. Even within the South Jakarta landscape, the existence of this species was more dominant than other termite species. A colony of this termite species lives in nest systems in the form of a mound, with a complex network of tunnels inside. This serves to protect the colony from the influence of extreme external environmental conditions [
5]. Subterranean termites of the subfamily
Macrotermitinae, particularly
Macrotermes gilvus Hagen (Isoptera: Termitidae), have the most extensive geographical distribution in Indonesia [
3]. These termites create fungus combs or fungus gardens inside their nest as a food source for the colony members [
6]. Therefore, this termite, as well as the other members of the subfamily
Macrotermitinae, is known as a fungus-cultivating termite.
The fungus comb is a special structure created by termite colonies from the sub-family Macrotermitinae (Isoptera: Termitidae) in their nests as a substrate for the growth of the fungus
Termitomyces sp. [
7]. This special structure is in the form of a mammalian brain with a volume of 44.17 ± 7.36 cm and has interconnected burrows from the surface to the bottom [
5]. Architecturally, the fungus comb consists of two structural parts: the fresh comb at the top and the old comb at the bottom. The density of old combs is higher compared to the fresh comb, which functions as a strong foundation for fungus-comb structure [
5]. Singh et al. [
8] reported that the structure of a fungus comb, which is ventilated and connected to the nest wall by particular tunnels, allows the exchange of CO
2 with the surrounding atmosphere, therefore maintaining a stable temperature inside the fungus comb throughout the year. Fungus comb has abundant clusters of nodules, small (about 1 mm in diameter) white spherules of
Termitomyces species [
7]. Fungal mycelium grows in these fungus combs and produces
Termytomyces nodules as a food source for the Macrotermitinae colonies. The nodules are the asexual phase composed of a hyphae collection with short cells, conidium, and sphaerocyst [
9]. When the fungus comb fails to be created by the termite colony, they eventually lack food and are unable to survive. However,
Termitomyces fungus does not always produce fruiting bodies, as termites consume the fungi in their nodule form [
10].
The termite colony creates conditions in which there is only one genus of fungus, i.e.,
Termitomyces, that can grow in the fungus comb [
11,
12], including in the fungus comb of
Macrotermes sp.,
Odontotermes sp., and
Ancistrotermes sp. [
13] In addition, Batra and Batra [
14] reveal that termite saliva has antifungal activity that inhibits the growth of other fungus genera, except
Termytomyces. Moreover, Thomas [
15] reported that the micro climate inside the termite mound was steadily maintained, including a temperature around 30 °C, which is very suitable for the growth of
Termytomyces. However, until today, there has been no comprehensive scientific information available regarding the mechanism of how
Termitomyces sp. became the only species that grows in fungus combs. Thus, the authors predict that chemical properties of the fungus comb are also responsible for stimulating the growth of
Termitomyces fungus species and suppressing others. Based on this proposition,
M. gilvus mounds can be potentially considered as a source of particular active compounds for an environmentally friendly organic fungicide. With such curiosity, the chemical properties of fungus combs and their potential as an active source of antifungal ingredients are or great research interest. Recently, the authors studied the architectural and physical properties of fungus combs from
Macrotermes gilvus mounds to understand their shape, volume, structure, the dimension of their burrows, texture, color, and density, as well as the existence of the
Termitomyces nodules [
5]. In addition, Anwar et al. [
16] successfully demonstrated fungus comb establishment under in vitro conditions using
Termitomyces cylindricus (Basidiomycota: Agaricales) basidiospores. However, the bioactivity of fungus-comb extract from Indo-Malayan termite
M. gilvus mounds as an antifungal has not been reported. Therefore, this research will be a pioneering study to determine the chemical composition of fungus comb from Indo-Malayan termite
M. gilvus mounds and will hopefully offer considerable contributions to the development of environmentally friendly organic fungicides, which are needed to prevent wood-staining fungi attacks on various wood species used in Indonesian wood industries.
Stain fungi are filamentous fungi that cause blue, greyish, green, and black discoloration on wood surfaces. Colonization of the fungi initiates a mineralization process occurs on the wood surface, leading to discoloration. The is discoloration influenced by melanin crystallization around hyphae or from secretion of extracellular material [
17]. Salman et al. [
18] reported that
Aspergillus foetidus caused the worst discoloration on air-dried rubberwood, which is processed for furniture products in West Java Province, Indonesia. The growth rate of fungus on the wood surface reached 90.5%. Meanwhile, Oldertrøen et al. [
19] found three stain-fungi species on rubberwood:
Aspergillus niger, Aspergillus flavus, and
Penicillium citrinum.According to George [
20] wood-staining fungi can seriously attack rubberwood within one day of felling. As a result of fungi attacks, rubberwood experiences discoloration, causing a significant decrease in its aesthetic value. An example of stain fungi is
Botryodiplodia theobromae, which occurs together with the surface molds
Aspergillus sp. and
Penicillium spp., which also cause considerable loss in aesthetic value of the attacked wood. The susceptibility of rubberwood to stain fungi and mold colonization on its surface is considered a major concern for furniture and wooden-toy manufacturers. The tropical climate in Indonesia, characterized by temperatures above 25 °C and humidity of 80–100%, imposes challenging conditions for the protection of rubberwood from stain fungi attack. Such environmental conditions are perfect for the growth of stain fungi and therefore facilitate the colonization of stain fungi [
21,
22]. Particularly in the rainy season, rainwater containing nutrients wets the wood, thus promoting biodeterioration activities of fungi [
23]. In 2018, economic loss due to stain fungi attacks on rubberwood raw material in Indonesian furniture industry alone reached almost USD 15,246 million [
18]. In fact, this economic loss is higher than the value since others light-color wood species used in the furniture industry were also susceptible to stain fungi. Even still, it is expected that this value will increase in the future if there are no sufficient control techniques developed.
The prevention wood-staining fungi attacks is critical, considering that 85% of the log production for Indonesian wood industries (47.9 million m
3/year) comes from plantation forests [
24]. Unfortunately, most of the wood from the plantation forests is highly susceptible to biodeterioration, including susceptibility to wood-staining fungi attacks [
25]. Such attacks contribute to the discoloration of the wood surface, i.e., changing it to blue, grayish, green, or black, considerably lessening its aesthetic value [
18]. Valiante et al. [
26] argued that discoloration is caused by pigmentation on the hyphae or melanin synthesis on the conidia during the sporulation process.
One of the most widely found wood-staining fungi that attacks wooden raw materials used in the Indonesian wood industry, including rubberwood (
Hevea brasiliensis Muell. Arg.), is
Aspergillus spp. [
18]. This situation requires multiple approaches, including the exploration of organic fungicide that is safe for humans and the environment, the active ingredients of which are originally produced from indigenous natural resources.
4. Conclusions
In addition to having a high enough moisture content (25.94%), the fungus combs from Indo-Malayan termite M. gilvus mounds also had a high crude-ash (30.57%) and crude-fiber (25.46%) content and was enriched with adequate carbohydrates (7.76%) and proteins (5,80%). In addition, the fungus combs contained 17 amino acids, with a total content of 5.53%, of which arginine, leucine, glutamate, and aspartic acid were the majority. Compared to other studies, protein the content of fungus comb from Indo-Malayan termite M. gilvus was much higher.
The fungus combs were rich in secondary metabolites, particularly from the phenol hydroquinone, steroid, terpenoids, and saponin groups. In addition, extractive substances from the fungus combs were dominated by nonpolar compounds in which the n-hexane extract consisted of 18 compounds, with 1,2,3-trimethyl-benzene, methyl palmitate, benzenepropanoic acid, 3,5-bis (1,1 dimethylethyl)-4-hydroxy-,methyl ester, methyl linolelaidate, methyl oleate, and bis(2-ethylhexyl) phthalate were identified as the dominant compounds. In the ethyl-acetate extract, 14 compounds were found, generally in the phenol group. Among the extractive substances, four dominant compounds, i.e., 2-methoxy-phenol, 2,6-dimethoxy-phenol, and benzenepropanoic acid, 3,5-bis (1,1 dimethylethyl)-4-hydroxy-,methyl ester were reported to display antifungal activities.
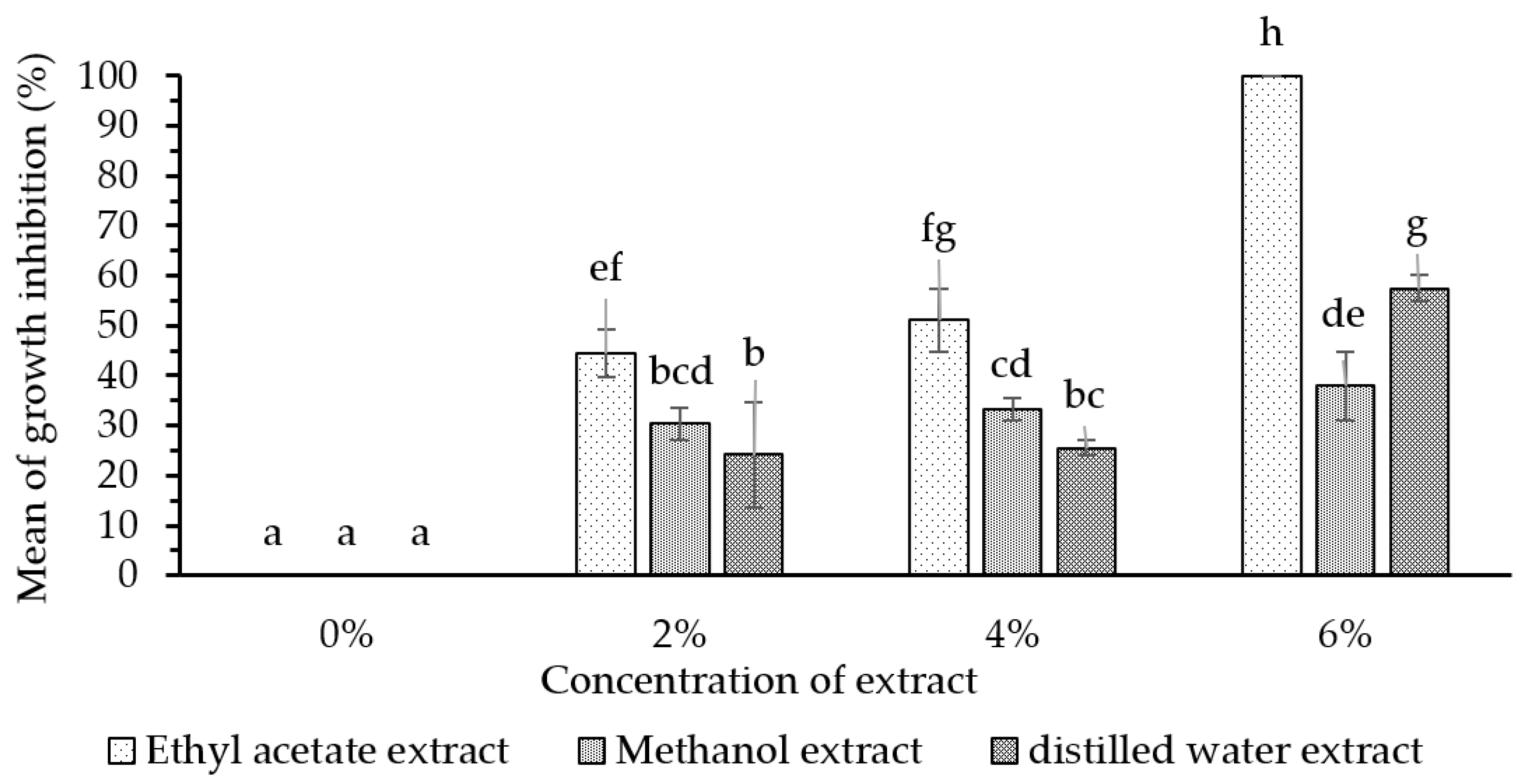
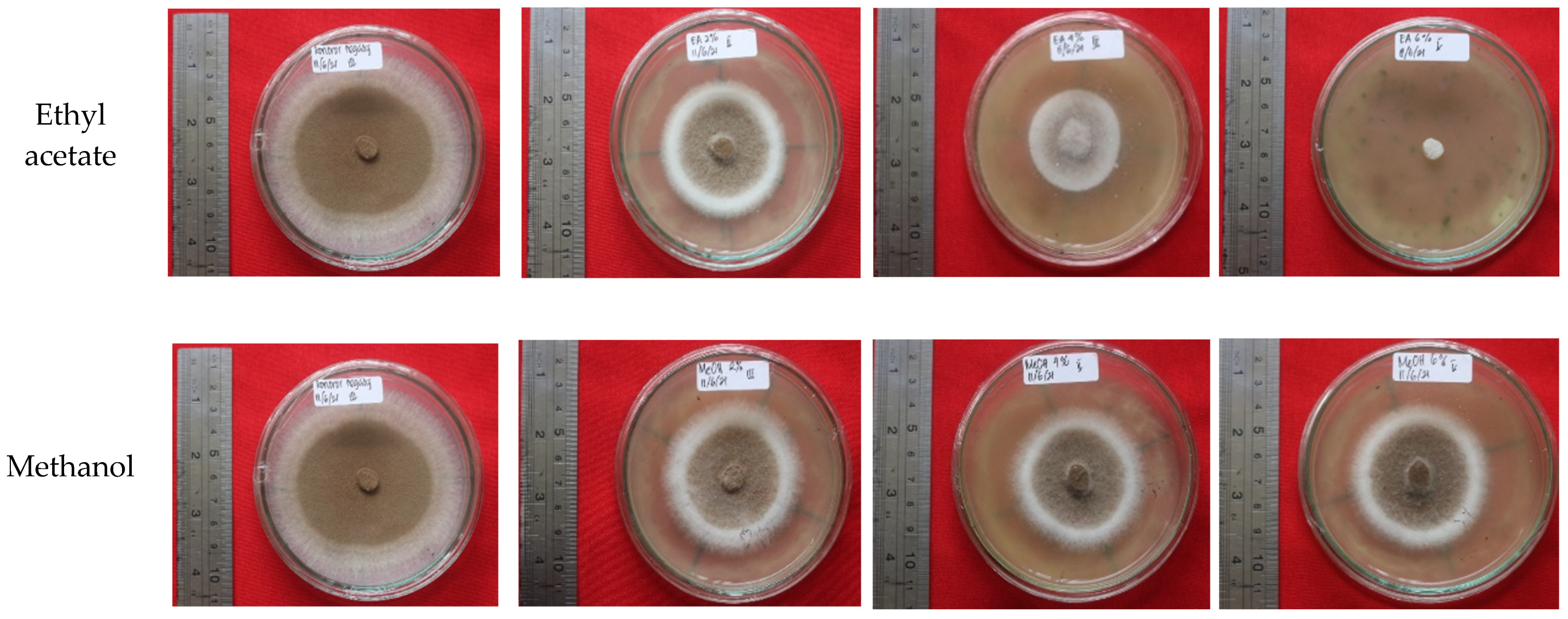
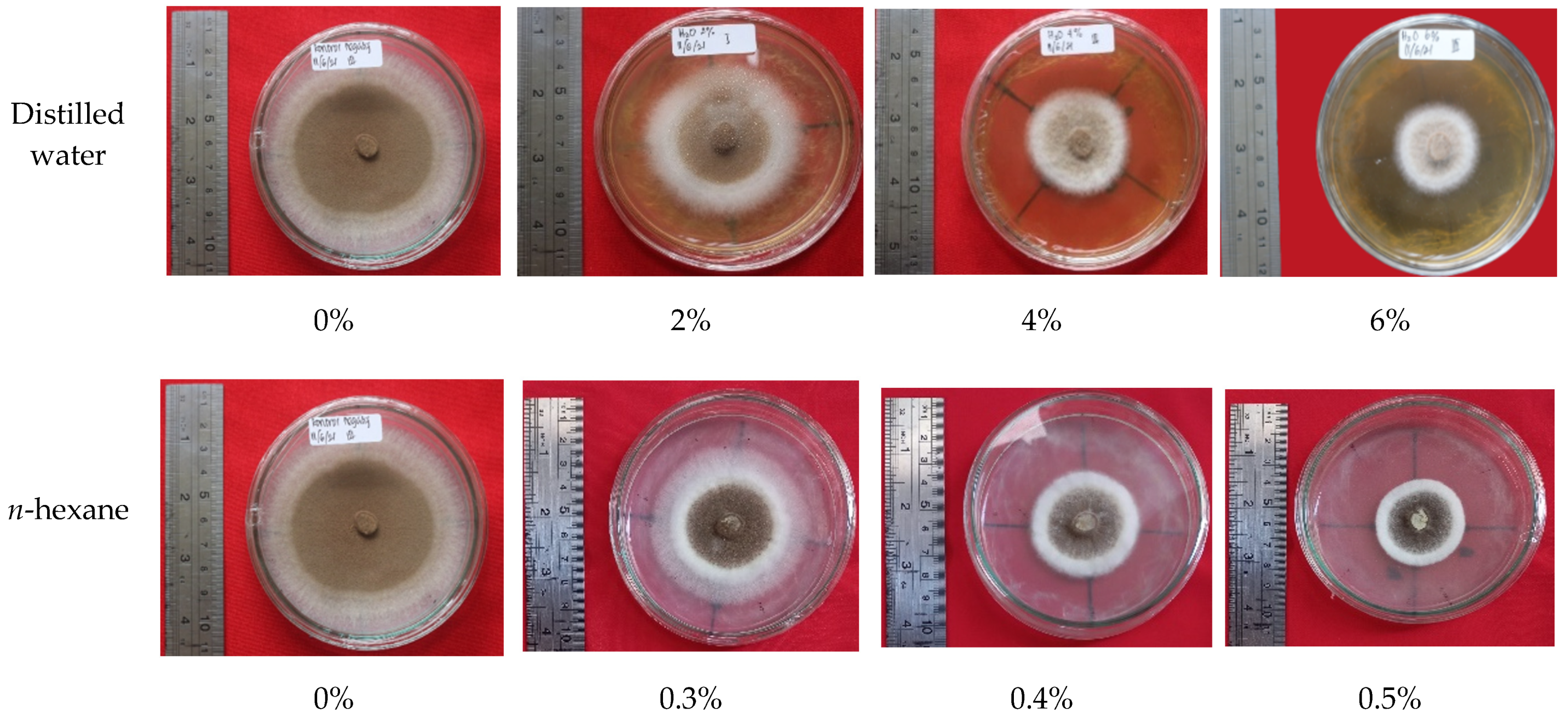
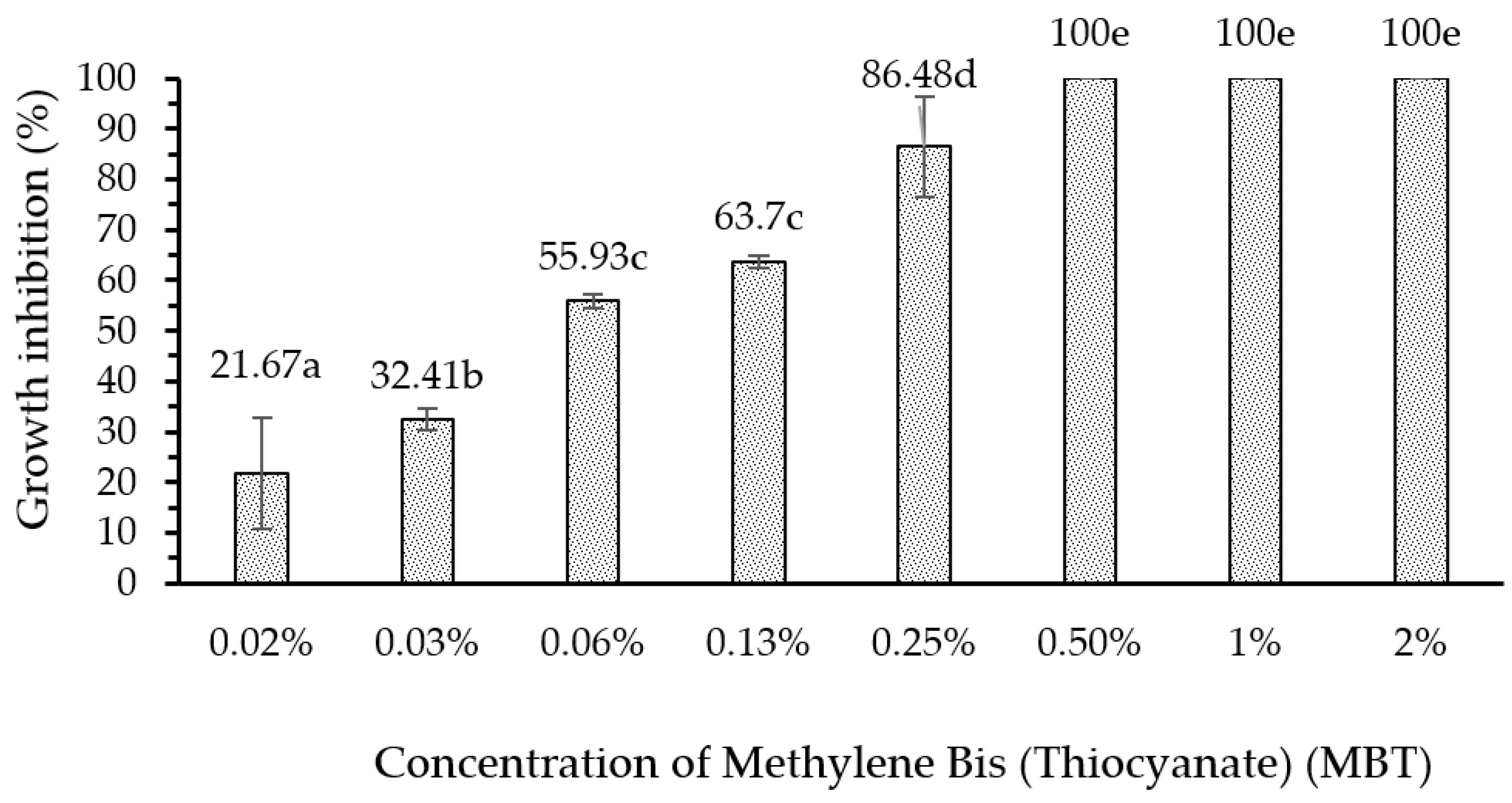
Ethyl-acetate extract and n-hexane extract from the fungus combs, with concentrations of 6% and 0.5%, respectively, showed remarkable bioactivity in terms of inhibiting wood-staining fungus A. foetidus. In addition, the bioactivity of the 6% ethyl-acetate extract showed a maximum inhibiting rate (100%) on wood-staining fungus growth equal to the efficacy of methylene bis-thiocyanate (MBT), which is widely used as an active ingredient in commercial synthetic wood preservatives. Further investigations are needed to identify the chemical structure of the active compounds in the fungus-comb extracts responsible for wood-staining-fungi-inhibition.



 ,
, 






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}