Tree Regeneration Patterns on Contrasting Slopes at Treeline Ecotones in Eastern Tibet


1
Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, Chinese Academy of Sciences, Beijing 100101, China
2
School of Life Science, Taizhou University, Taizhou 318000, China
3
College of Resources and Environment, University of Chinese Academy of Sciences, Beijing 100190, China
4
Institute of Wetland Ecology & Clone Ecology/Zhejiang Provincial Key Laboratory of Plant Evolutionary Ecology and Conservation, Taizhou University, Taizhou 318000, China
*
Author to whom correspondence should be addressed.
Forests 2021, 12(11), 1605; https://0-doi-org.brum.beds.ac.uk/10.3390/f12111605
Submission received: 8 October 2021
/
Revised: 10 November 2021
/
Accepted: 15 November 2021
/
Published: 20 November 2021
(This article belongs to the Section Forest Ecology and Management)
Abstract
:Alpine treelines are projected to shift upslope in response to climate warming, but empirical studies have yielded inconsistent results, with both upshifted and stable alpine treelines. Additionally, treelines on different slope aspects of the same mountain can differ. Thus, for a better understanding of the mechanisms of treeline formation and treeline responses to climate change, we need to elucidate the population dynamics at treelines on different slope aspects. Here, we quantified the population dynamics of Balfour spruce (Picea likiangensis var. rubescens) at treeline ecotones on contrasting north- and east-facing slopes on the eastern Tibetan Plateau based on field surveys. The alpine treeline positions of Balfour spruce have not advanced toward higher altitudes on the contrasting slopes in recent decades. Compared with the east-facing slope, more recruits occurred on the north-facing slope above the present treeline, indicating a more favorable regeneration condition. However, on the north-facing slope, the individual growth rate of Balfour spruce was lower, and the number of adult trees above the present treeline was higher than that on the east-facing slope. Thus, slope aspects mediate a trade-off between the growth and survival of treeline species, explaining the absence of an impact of slope aspects on treeline responses to climate change. Our results highlight the importance of considering the effect of topography on population dynamics in predicting alpine treeline dynamics under the scenario of climate change.
1. Introduction
Alpine treelines represent one of the most obvious vegetation boundaries in the world [1]. Due to low temperature, treeline trees acclimate to harsh living conditions close to their physiological limits [1]. As a consequence, alpine treelines are highly sensitive to climate warming and are frequently used as an indicator of climatic changes [2]. Previous studies have reported that some alpine treelines advanced to higher altitudes in response to climate warming, but others did not respond or even decline to lower elevations [3,4,5]. Such inconsistent results highlight the importance of better understanding the population dynamics of treeline trees in order to disentangle the sensitivity of treeline regeneration in response to climatic changes.
Population dynamics at treelines are closely associated with climate change-induced treeline dynamics [6,7], which usually include treeline elevation shift, tree regeneration, and annual tree growth [1]. Among these processes, treeline elevation shift largely depends on the latter two, mainly because only successful seedling establishment and seedling-to-tree transition above the present treeline can contribute to treeline shifts [1,7,8]. Therefore, although treeline shift is the simplest descriptor of treeline dynamics [9], population dynamics related to recruitment and annual growth at treelines have received considerable attention in the context of climate warming [10,11,12]. Previous studies have revealed that climate parameters such as temperature, precipitation, and relocation of snow can exert a direct influence on the population dynamics at treelines, including seed germination [13,14], seedling establishment [15,16,17], and survival and growth of seedlings, juveniles, and adults [18,19,20,21]. To better understand treeline responses to climate warming, it is essential to focus on the ecological processes of population dynamics under a changing climate.
The elevations of alpine treelines differ along latitudes [1,8]; generally, they decrease with increasing latitudes due to thermal deficiency [1,8]. Additionally, slope aspects can influence the elevations of alpine treelines, usually with higher elevations on sunny slopes than on shady ones [8]. Differences in species composition, seedling survival, tree growth, and forest structure among different slope aspects near treelines have been documented [8,11,22,23]. However, it is still unclear how these processes relate to treeline shift. Against the background of a changing climate, the responses of alpine treelines to climate warming could be leveled off by the differences in treeline elevations caused by slope aspects. Therefore, determining the effects of fine-scale variation in slope aspects on the population dynamics of treeline species is crucial to understand the sensitivity of alpine treelines in response to climate warming from landscape to regional scale.
On the Tibetan Plateau, the climate has been experiencing a dramatic warming trend [24,25,26]. In this region, the most diverse and highest alpine treelines in the Northern Hemisphere are distributed [27]. Some studies have observed an upward shift of the treeline in recent years [4,5], whereas others have found relatively stable treelines [7,9,28]. Most studies observed that regeneration increased drastically in response to climate warming [29,30], whereas in some studies, regeneration decreased due to anthropogenic disturbance [31,32]. However, all these studies did not concern the effects of slope aspects on responses of treeline dynamics to climate warming. To better understand this issue in this region, we quantified the population dynamics of Balfour spruce (Picea likiangensis var. rubescens) at treeline ecotones on north- and east-facing mountain slopes on the eastern Tibetan Plateau based on field surveys. Specifically, we addressed two questions: (1) Does the alpine treeline elevation shift under climate change? (2) Do tree regeneration and growth at alpine treeline ecotones differ among different slope aspects?
2. Materials and Methods
2.1. Study Area and Climate
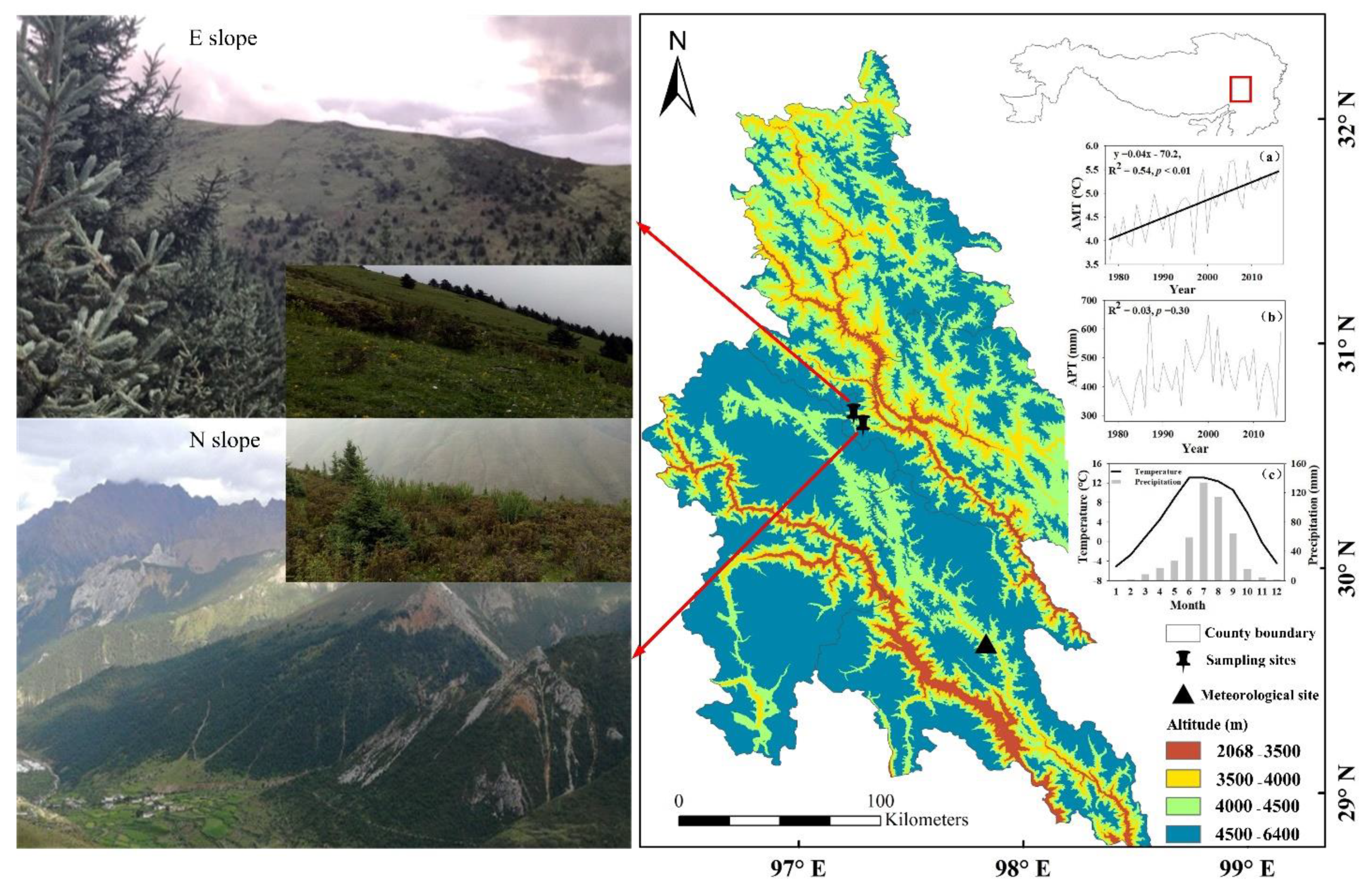
The study area was located at Langlashan mountain, Chaya County, Changdu Prefecture of the Tibet Autonomous Region, eastern Tibetan Plateau, under control by East Asian monsoon climate (30°38′ N, 97°17′ E, 4597 m asl, Figure 1). Based on the records of the nearest meteorological station in Zuogong County (29°40′ N, 97°50′ E, 3780 m asl.), annual mean temperature is 4.7 °C, with a mean temperature range from −5.1 to 13.1 °C. January and July are the coldest and warmest months, respectively. Annual precipitation is 449 mm, of which 83% fall as rain during June to September. Annual mean temperature shows a warming trend (R2 = 0.54, p < 0.01), and annual precipitation shows no apparent variation trend since 1978 (R2 = 0.03, p > 0.05, Figure 1).
2.2. Tree Species
Picea likiangensis var· rubescens (Rehd·et Wils) (Balfour spruce, Pinaceae) is one of the dominant tree species forming alpine treelines on the eastern Tibetan Plateau. Balfour spruce frequently forms pure forest stands or mixed stands with other junipers (e.g., Juniperus tibetica) and extends toward elevations above 4000 m asl. One of our sampling sites was located on the north-facing slope where pure stands of Balfour spruce are distributed, with dense shrubs of Rhododendron nyingchiense. Average treeline and timberline elevations on this slope are 4438 and 4414 m, respectively. The other one was on the east-facing slope, where stands of Balfour spruce are mixed with sporadically occurring junipers, with relatively sparse shrubs of R. nyingchiense and high abundance of short sedge (e.g., Kobresia humilis). Average treeline and timberline elevations on this slope are 4483 and 4464 m, respectively. Treeline elevations in our study sites are equivalent to neighboring treeline sites [33]. Due to high elevations, there is no human disturbance on these two slopes, and thus, the treeline in our study should be considered a “climatic treeline” (in contrast to the anthropogenic treeline due to human activities such as timber harvesting and livestock production).
2.3. Field Sampling
Tree species line was determined by the uppermost distribution of the treeline species. Treeline position was defined by the maximum altitude at which upright trees higher than 2 m clumped, and the upper limit of continuous forest with a forest cover higher than 20% was regarded as timberline. The treeline ecotone was defined as the transition zone from timberline to tree species line [1,8].
We surveyed two sites of Balfour spruce treeline ecotones (north- and east-facing slopes) in the summer of 2018, with three spatially independent transects along the treeline ecotone as three replicates for each slope (N1, N2, and N3 for the north-facing slope; E1, E2, and E3 for the east-facing slope). Each transect was a rectangle plot with a width of 30 m (perpendicular to the slope) and a length of 150–390 m (along the slope), from the species line to at least 50 m below the timberline, spanning the entire treeline ecotone. We selected a topographically uniform area and avoided rugged terrain, landslides, and debris flow while establishing each transect. In each transect, we used a diameter tape to determine the base diameter and diameter at breast height (DBH) of each individual tree. We used a measuring stick to determine the height of trees shorter than 2 m and a clinometer for those higher than 2 m. The location of each Balfour spruce individual was mapped on the xy-plane [9]. Simultaneously, for the adult trees higher than 2 m, we took one or two cores at stem base perpendicular to the slope, as close as possible to the ground level, with an increment borer, collecting a total of 463 cores. For the juveniles (0.5 m < height < 2 m) and seedlings (<0.5 m), we counted the number of branch whorls and the bud scars along the main stem to determine their ages in a non-destructive way [34,35,36]. Previous studies have proved that this method can estimate seedling and juvenile age within an acceptable error limit [4,9,37].
The cores were taken to the laboratory for tree ring recording. They were air-dried, tied to a narrow slot with white latex, then polished with finer grits of sanding paper to make the annual rings visible, followed by determining tree ring width and ages by standard dendrochronological procedures [38,39,40], with a LINTAB™ 5 tree-ring measurement equipment under a stereomicroscope at a resolution of 0.01 mm (Rinntech, Heidelberg, Germany) [41]. Tree-ring series were cross-dated visually and statistically to the calendar year, and the COFECHA program was used to check the accuracy of cross-dating [42,43,44].
Adult tree age was determined by counting the number of rings in the collected sampling cores. If a core did not reach the pith, but was close to it, we used a geometrical method to estimate the missing years [45] and then added the missing years to the years obtained from tree cores. For the individual with rotten pith, we used the tree age–DBH or tree age–height relationship to estimate its age. Based on tree age–height relationship, we also estimated the time required for a seedling to reach the breast height and 2 m for the two slope aspects, respectively. The time required for a tree to reach breast height (1.37 m) and 2 m was 23 and 28 years on the north-facing slope, respectively, and 14 and 18 years on the east-facing slope, respectively.
2.4. Identifying the Dynamics of the Balfour Spruce Treeline
Treeline dynamics are mainly reflected by treeline elevation shifts and the intensity of regeneration in the treeline ecotones. First, we figured out the establishing year of each individual at a 50-year time interval. Second, we calculated the number of establishing individuals on a decadal interval and defined the significant increase in recruitment as the number of individuals that increased by 50% compared with that in the previous age class [9]. The two-sample Kolmogorov–Smirnov test was employed to examine the differences between temporal recruitment patterns on both slopes. Third, as successful tree establishment is essential to treeline dynamics [1], we focused on two young age classes (2001–2010 and 2011–2020) to depict regeneration dynamics by comparing tree height, base diameter, and growth rate on the two slopes. For these analyses, we pooled data from the three transects on the same slope.
Two-sample Kolmogorov–Smirnov test, and the independent-samples t test were conducted with the SPSS software 20.0 (IBM SPSS Statistics for Windows, IBM Corp., Armonk, NY, USA).
3. Results
3.1. Treeline Dynamics
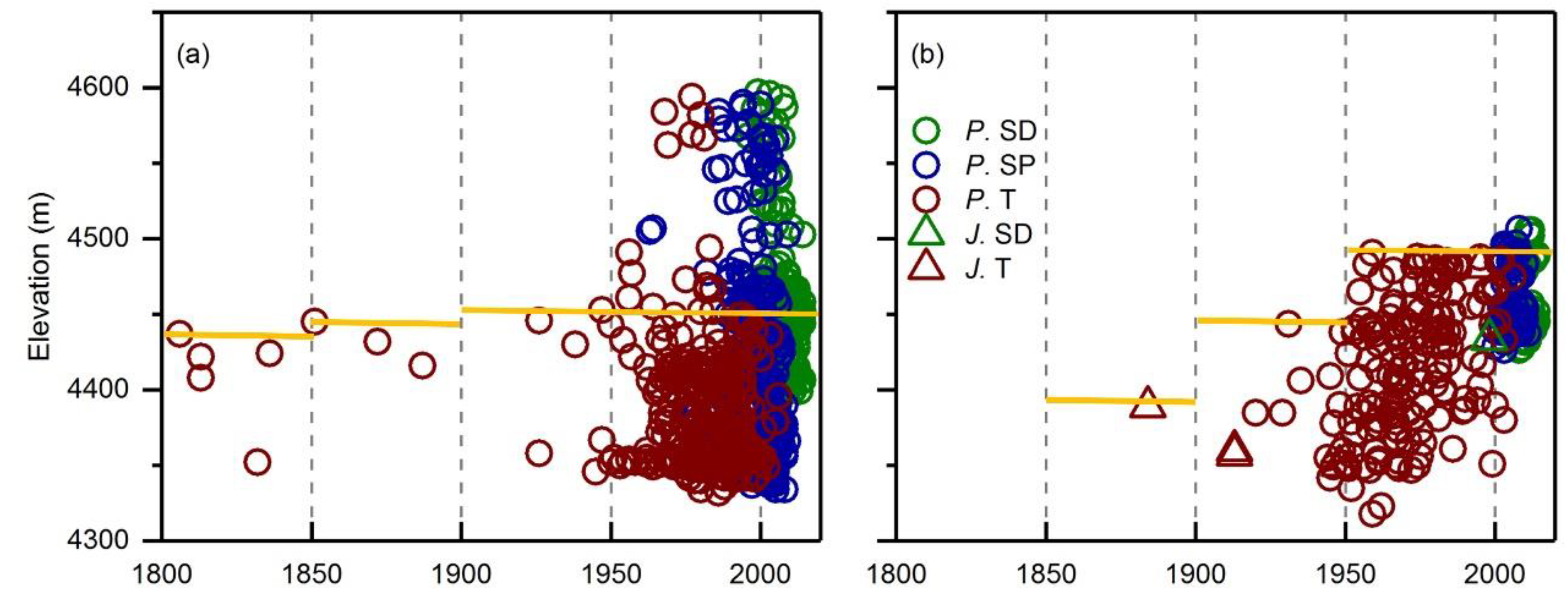
The treeline elevation positions on the north-facing slope were significantly lower than those on the east-facing slope (Table 1). The treeline was quite stable on the north-facing slope (Figure 2a), but showed a slight advancement on the east-facing slope from 1900 onwards (Figure 2b). Notably, on the east-facing slope, few regenerated trees could be observed above the current treeline, indicating poor regeneration potential (Figure 2b). However, on the north-facing slope, a growing number of juveniles and seedlings, and even sporadic adult trees, established towards the higher altitudes, indicating a larger potential of regeneration and treeline advancement (Figure 2a).
3.2. Age Structure and Growth Characteristics of Regenerated Balfour Spruce
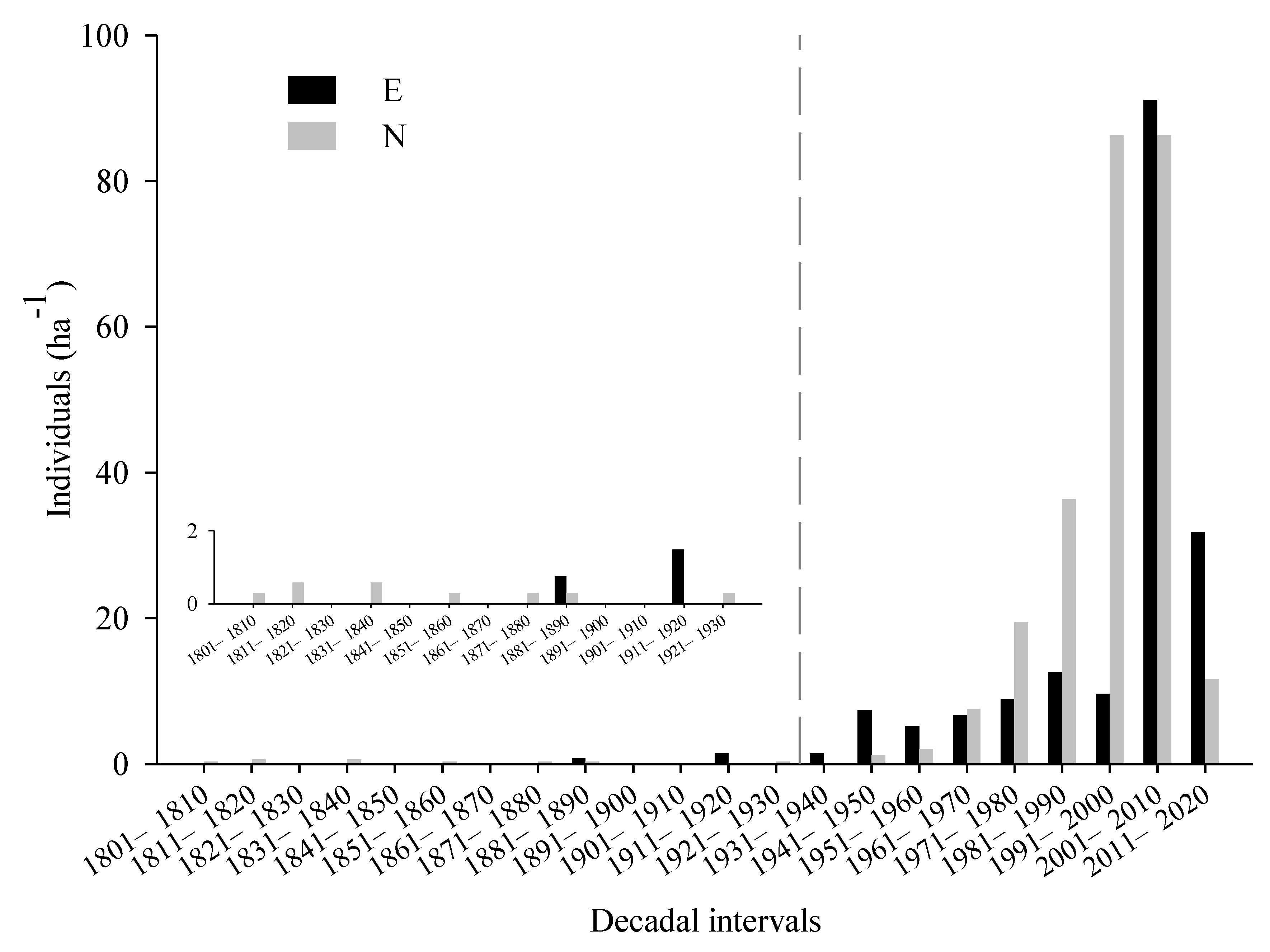
The age structure of Balfour spruce showed a reverse J-shape on both slopes, i.e., the number of adult trees decreased with increasing age (Figure 3). Tree density on the north-facing slope was significantly higher than that on the east-facing slope (N: 249 ± 5 individuals/hm2; E: 162 ± 16 individuals/hm2). The number of adult trees was higher on the north-facing slope than that on the east-facing slope (Table 1 and Figure 3). The oldest tree on the north-facing slope was 212 years, considerably older than the oldest tree of 135 years on the east-facing slope (Figure 2 and Figure 3).
The Kolmogorov–Smirnov test showed that the patterns of tree recruitment on the two slopes were insignificantly different (p > 0.05, Figure 3). Tree recruitment showed a continuous increase from 1961 to 2010 on the north-facing slope, especially pronounced from the 1980s on, accounting for 80% of the total recruits (Figure 3). However, tree recruitment showed an abrupt increase from 1941 to 1950 on the east-facing slope, then maintained a constant rate until 2000, followed by a second increase pulse during the period 2001–2010 (Figure 3).
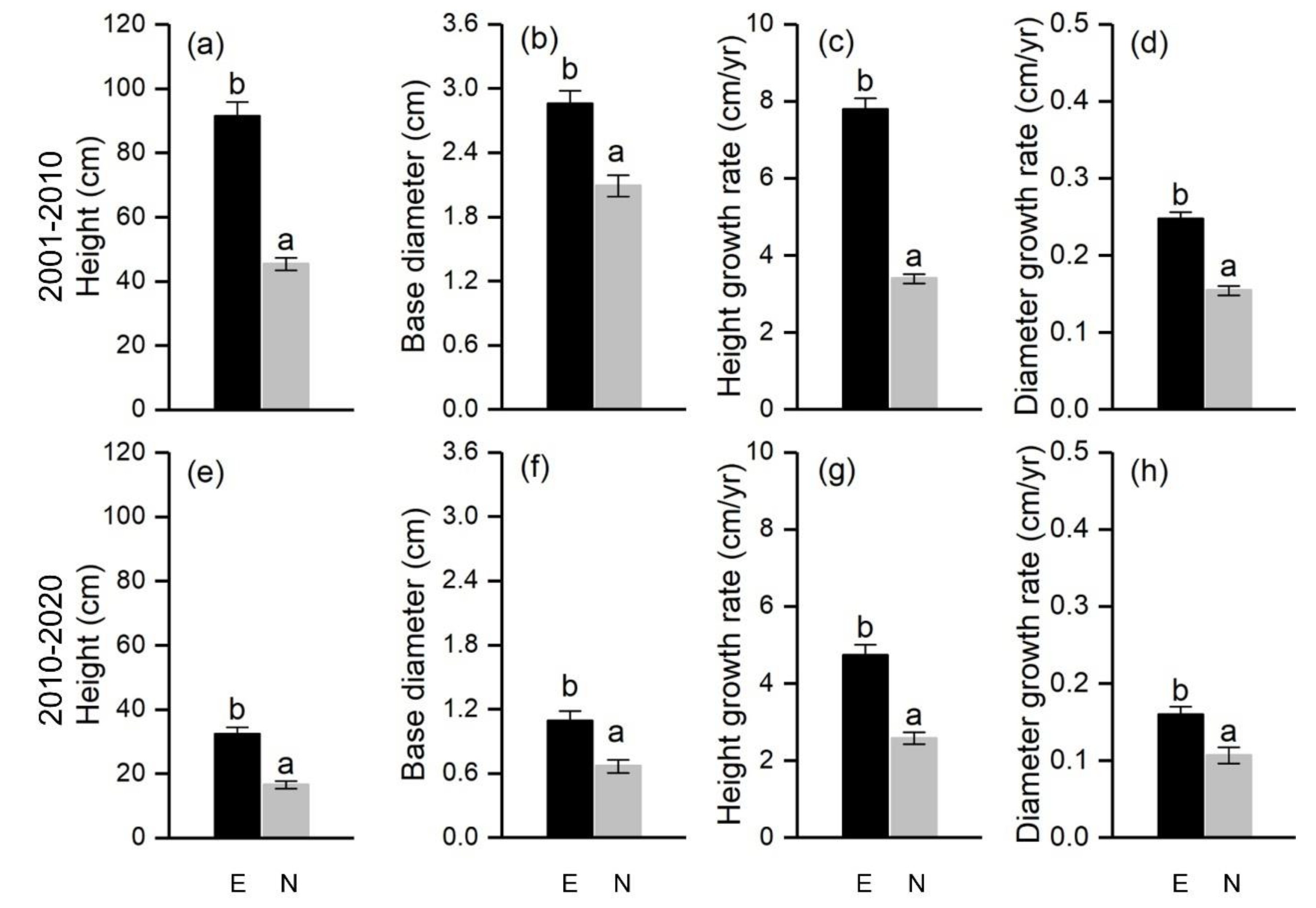
For the two age classes 2001–2010 and 2011–2020, tree height, base diameter, height growth rate, and diameter growth rate were significantly higher on the east-facing slope than that on the north-facing slope, indicating that individuals on the east-facing slope regenerated more rapidly than those on the north-facing slope (Figure 4).
4. Discussion
In our study, we observed no apparent upward shift of the Balfour spruce alpine treeline on both slopes on the eastern Tibetan Plateau, although the area has experienced dramatic warming in recent decades [24,25,26]. Interestingly, we found a favorable regeneration pattern on the shady (north-facing) slope above the current treeline, while an unfavorable one on the sunny (east-facing) slope. Unexpectedly, young individuals grew much more slowly on the shady slope than on the sunny slope, suggesting the effect of slope aspect induced microhabitat conditions on population dynamics at treeline ecotones in a warming climate.
4.1. Slight Advancement in Treeline Position
Generally, treeline elevation is higher on sunny than on shady slopes due to thermal advantage [8]. Our results support this notion and showed that treeline position was tens of meters higher on the east-facing slope than on the north-facing slope. The main explanation for this phenomenon is that sunny slopes are usually warmer compared to shady slopes because of the higher solar radiation in the same elevation [8]. Treelines are commonly limited by low temperatures, and thus, a warming climate can promote treeline shifts to higher altitudes or latitudes [1]. Against the background of a warming climate, treeline elevation shifts have occurred in mountain ranges of Europe [34,46,47], North America [48,49], and Asia [4,5,37]. However, our result indicates that no advancement of alpine treelines has been explored on both slopes, which seems contradictive to common sense. Other surveys in the eastern Tibetan Plateau are also in line with our findings [30,33], and the results have been explained by plant–plant interactions [30] and differences in treeline forms [7]. On the shady slope, the establishment of seedlings and saplings was enhanced under dense shrubs but are difficult to recruit into adult due to death under insufficient light, self-thinning, and competition with shrubs [50,51,52]. In contrast, regeneration was subjected to drought and exposure to radiation and thus recruitment is very rare on the sunny slope [8]. Additionally, trade-off between the growth and survival of tree species, mediated by slope aspects, is another possible reason we want to address later.
4.2. Population Regeneration
We found a successive regeneration pattern above the current treeline on the shady slope, indicating a somewhat successful seedling establishment. Generally, seedling establishment is controlled by abiotic and biotic factors at the alpine treeline ecotones. The growth and development of alpine plants are limited by low temperatures [1]. As the sunny slope is characterized by higher temperatures, the vegetation on such slopes has heat advantages over that on shady slopes. Sufficient heat at high altitudes favors seedling growth, resulting in higher growth rates. Moreover, strong solar radiation facilitates an earlier snowmelt, thereby prolonging the growing season [7] and, consequently, increasing the annual tree growth. In addition, plant interactions, such as competition, can influence tree regeneration, as smaller seedlings are more sensitive to competition than larger ones [53]. In our case, the surrounding shrubs were denser and higher on the shady slope than on the sunny slope and may therefore more fiercely compete with Balfour spruce seedlings for sunlight, water, and nutrients. Such denser and higher surrounding shrubs can also lead to less sky exposure, which can increase needle temperatures at nighttime, decrease insolation during early morning hours, and substantially reduce photosynthetic carbon gain and cause a lower growth rate [54,55,56]. Furthermore, the slow growth rate on the shady slope is likely to be an adaptation mechanism of seedlings to maximize survival in harsh habitats [57]. In harsh environments such as alpine treeline ecotones, rapidly growing seedlings may not be successful in long-term seedling establishment [58].
Coincidently, we observed a lower seedling abundance and distribution on the sunny slop, indicating lower seedling survival, which is in line with previous studies [12,59]. There are some explanations for this phenomenon. First, the sunny slopes contained more open areas because of the sparse distribution of shrubs. In such open areas, solar radiation is stronger, with a higher surface temperature and more intense evapotranspiration [8]; excessive solar radiation can cause higher percentages of red leaves (indicator of high concentrations of photoprotective anthocyanins) and increase seedling mortality [10,12]. Contrarily, shelters provided by ambient shrubs and herbs can largely increase survival rate due to less sky exposure [12,60]. Second, open areas generally have lower nighttime temperatures due to long-wave irradiance, with more frequent freezing events in the growing seasons [1,59]. Following the statistical approach detailed in Shen et al. [59], our data logger records somehow confirmed this. The frequency (28 days for the N slope, 33 days for the E slope) and duration (168 h for N, 186 h for E) of growing season freezing events were higher on the sunny than on the shady slope (unpublished data). Third, also the occurrence of extreme cold events has decreased with climate warming [61], winter conditions can be extremely challenging for seedling survival at high-altitude treeline ecotones, with frequent freezing injuries and winter desiccation [8,62], which is fatal in the initial life stage of young seedlings with less protection from surroundings on sunny slopes.
Overall, on both slopes, high growth rates were accompanied by low survival and vice versa. A trade-off between growth and survival of Balfour spruce mediated by slope aspects appeared, which might control the responses of alpine treeline to climate change at the local scale.
4.3. Implication of Population Dynamics to Treeline Shift
Alpine treelines are expected to shift upslope in response to a warming climate [1]. Previous studies on treeline dynamics, however, have not drawn a consistent conclusion [3]. In the past century, a substantial tree densification has occurred at alpine treeline ecotones, which seems to be more common than elevational shifts upward under climate warming [7,9,50,63]. A growing number of studies are reporting this mismatch between population dynamics and treeline elevation shift [9,63,64]. Generally, only if individuals above the present treeline can survive and successfully finish the seedling-to-tree transition, upward treeline shift occurs [1,8]. Hence, seedling establishment is prerequisite for treeline elevation shift. As previously mentioned, climatic variables, as abiotic factors, control the upper tree distribution boundaries at landscape to regional scale [2]. Biotic factors also play an important role in seedling establishment at local scales [10,65], which in turn determines whether treeline dynamics can keep pace with climate warming at local or larger scales. Therefore, a comprehensive analysis including both regeneration pattern and elevation shift should be considered to objectively reflect alpine treeline dynamics under the scenario of climate change.
5. Conclusions
Based on our findings, the alpine treeline of Balfour spruce is relatively stable, in spite of the rapidly warming climate in eastern Tibet. However, more recruits, including sporadic adult trees, appeared above the present treeline on the north-facing slope than that on the east-facing slope. Additionally, contrary to the north-facing slope, a poor tree regeneration condition occurred on the east-facing slope, while with a high growth rate. A trade-off between the growth and survival in population regeneration mediated by slope aspects may constrain the treeline range shifts on both slopes. These results have answered the questions we raised above, suggesting more biophysiological mechanisms are need to be disclosed at alpine treeline ecotones to accurately predict treeline expansion in response to climate change.
Author Contributions
L.Z. and P.S. conceived of the project and planed the analytical approach. L.Z., T.Z. and G.H. conducted the field survey and analyzed the data. L.Z. and P.S. visualized the tables and figures. L.Z. wrote the original paper. P.S., M.S. and F.Y. contributed to data interpretation and edited the paper. All authors have read and agreed to the published version of the manuscript.
Funding
This study was supported by the National Natural Science Foundation of China and ICIMOD Joint Research Project (No. 41661144045).
Acknowledgments
Many thanks go to Xu Min, Zhiwei Lai, Zhichang Cheng and Zhenkun Wang, as well as to two Tibetan drivers Dawaduoji and Pingcuodengzeng for the difficult field survey. We also thank Zhipeng Wang for the climate data acquisition during the data analysis.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Körner, C. Alpine Treelines; Springer: Basel, Switzerland, 2012. [Google Scholar]
- Körner, C.; Paulsen, J. A world-wide study of high altitude treeline temperatures. J. Biogeogr. 2004, 31, 713–732. [Google Scholar] [CrossRef]
- Harsch, M.A.; Hulme, P.E.; McGlone, M.S.; Duncan, R. Are treelines advancing? A global meta-analysis of treeline response to climate warming. Ecol. Lett. 2009, 12, 1040–1049. [Google Scholar] [CrossRef]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Borgaonkar, H.P. Treeline dynamics with climate change at the central Nepal Himalaya. Clim. Past 2014, 10, 1277–1290. [Google Scholar] [CrossRef] [Green Version]
- Gaire, N.P.; Koirala, M.; Bhuju, D.R.; Carrer, M. Site- and species-specific treeline responses to climatic variability in eastern Nepal Himalaya. Dendrochronologia 2017, 41, 44–56. [Google Scholar] [CrossRef]
- Conlisk, E.; Castanha, C.; Germino, M.J.; Veblen, T.T.; Smith, J.M.; Kueppers, L. Declines in low-elevation subalpine tree populations outpace growth in high-elevation populations with warming. J. Ecol. 2017, 105, 1347–1357. [Google Scholar] [CrossRef] [Green Version]
- Schickhoff, U.; Bobrowski, M.; Böhner, J.; Bürzle, B.; Chaudhary, R.P.; Gerlitz, L.; Heyken, H.; Lange, J.; Müller, M.; Scholten, T.; et al. Do Himalayan treelines respond to recent climate change? An evaluation of sensitivity indicators. Earth Syst. Dyn. 2015, 6, 245–265. [Google Scholar] [CrossRef]
- Holtmeier, F.-K. Mountain Timberlines—Ecology, Patchiness, and Dynamics; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Liang, E.; Wang, Y.; Eckstein, D.; Luo, T. Little change in the fir tree-line position on the southeastern Tibetan Plateau after 200 years of warming. New Phytol. 2011, 190, 760–769. [Google Scholar] [CrossRef] [PubMed]
- Akhalkatsi, M.; Abdaladze, O.; Nakhutsrishvili, G.; Smith, W.K. Facilitation of Seedling Microsites by Rhododendron caucasicum Extends the Betula litwinowii Alpine Treeline, Caucasus Mountains, Republic of Georgia. Arct. Antarct. Alp. Res. 2006, 38, 481–488. [Google Scholar] [CrossRef] [Green Version]
- Elliott, G.P.; Cowell, C.M. Slope Aspect Mediates Fine-Scale Tree Establishment Patterns at Upper Treeline during Wet and Dry Periods of the 20th Century. Arct. Antarct. Alp. Res. 2015, 47, 681–692. [Google Scholar] [CrossRef] [Green Version]
- Germino, M.J.; Smith, W.K.; Resor, A.C. Conifer seedling distribution and survival in an alpine-treeline ecotone. Plant Ecol. 2002, 162, 157–168. [Google Scholar] [CrossRef]
- Olson, A.R.; Richards, J.H.; Richards, A.R.O.H. Temperature Responses of Germination in Arctic Poppy (Papaver radicatum Rottb.) Seeds. Arct. Alp. Res. 1979, 11, 343. [Google Scholar] [CrossRef]
- Hoyle, G.L.; Venn, S.; Steadman, K.; Good, R.B.; McAuliffe, E.J.; Williams, E.R.; Nicotra, A. Soil warming increases plant species richness but decreases germination from the alpine soil seed bank. Glob. Chang. Biol. 2013, 19, 1549–1561. [Google Scholar] [CrossRef]
- Germino, M.J.; Graumlich, L.J.; Maher, E.J. Recent Relationships of Tree Establishment and Climate in Alpine Treelines of the Rocky Mountains. In Proceedings of the AGU Fall Meeting Abstracts, San Francisco, CA, USA, 10 December 2007; p. GC33B-05. [Google Scholar]
- Loranger, H.; Zotz, G.; Bader, M.Y. Early establishment of trees at the alpine treeline: Idiosyncratic species responses to temperature-moisture interactions. AoB Plants 2016, 8, plw053. [Google Scholar] [CrossRef] [Green Version]
- Lett, S.; Dorrepaal, E. Global drivers of tree seedling establishment at alpine treelines in a changing climate. Funct. Ecol. 2018, 32, 1666–1680. [Google Scholar] [CrossRef] [Green Version]
- Shi, C.; Shen, M.; Wu, X.; Cheng, X.; Li, X.; Fan, T.; Li, Z.; Zhang, Y.; Fan, Z.; Shi, F.; et al. Growth response of alpine treeline forests to a warmer and drier climate on the southeastern Tibetan Plateau. Agric. For. Meteorol. 2019, 264, 73–79. [Google Scholar] [CrossRef]
- Reich, P.B.; Oleksyn, J. Climate warming will reduce growth and survival of Scots pine except in the far north. Ecol. Lett. 2008, 11, 588–597. [Google Scholar] [CrossRef] [PubMed]
- Batllori, E.; Camarero, J.J.; Ninot, J.M.; Gutiérrez, E. Seedling recruitment, survival and facilitation in alpine Pinus uncinate tree line ecotones. Implications and potential responses to climate warming. Glob. Ecol. Biogeogr. 2009, 18, 460–472. [Google Scholar] [CrossRef]
- Zheng, L.; Gaire, N.P.; Shi, P. High-altitude tree growth responses to climate change across the Hindu Kush Himalaya. Journal of Plant Ecology 2021, 14, 829–842. [Google Scholar] [CrossRef]
- Salzer, M.; Larson, E.R.; Bunn, A.G.; Hughes, M.K. Changing climate response in near-treeline bristlecone pine with elevation and aspect. Environ. Res. Lett. 2014, 9, 114007. [Google Scholar] [CrossRef]
- Yirdaw, E.; Starr, M.; Negash, M.; Yimer, F. Influence of topographic aspect on floristic diversity, structure and treeline of afromontane cloud forests in the Bale Mountains, Ethiopia. J. For. Res. 2015, 26, 919–931. [Google Scholar] [CrossRef]
- IPCC. Climate change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergov-ernmental Panel on Climate Change; 9291691437; IPCC: Geneva, Switzerland, 2014. [Google Scholar]
- Liu, X.D.; Chen, B.D. Climatic warming in the Tibetan Plateau during recent decades. Int. J. Climatol. 2000, 20, 1729–1742. [Google Scholar] [CrossRef]
- You, Q.; Min, J.; Kang, S. Rapid warming in the Tibetan Plateau from observations and CMIP5 models in recent decades. Int. J. Clim. 2015, 36, 2660–2670. [Google Scholar] [CrossRef]
- Miehe, G.; Miehe, S.; Vogel, J.; Co, S.; La, D. Highest Treeline in the Northern Hemisphere Found in Southern Tibet. Mt. Res. Dev. 2007, 27, 169–173. [Google Scholar] [CrossRef] [Green Version]
- Lu, X.; Liang, E.; Wang, Y.; Babst, F.; Camarero, J.J. Mountain treelines climb slowly despite rapid climate warming. Glob. Ecol. Biogeogr. 2021, 30, 305–315. [Google Scholar] [CrossRef]
- Gaire, N.P.; Dhakal, Y.R.; Lekhak, H.C.; Bhuju, D.R.; Shah, S.K. Dynamics of Abies spectabilis in Relation to Climate Change at the Treeline Ecotone in Langtang National Park. Nepal J. Sci. Technol. 2012, 12, 220–229. [Google Scholar] [CrossRef] [Green Version]
- Liang, E.; Wang, Y.; Piao, S.; Lu, X.; Camarero, J.J.; Zhu, H.; Zhu, L.; Ellison, A.M.; Ciais, P.; Penuelas, J. Species interactions slow warming-induced upward shifts of treelines on the Tibetan Plateau. Proc. Natl. Acad. Sci. USA 2016, 113, 4380–4385. [Google Scholar] [CrossRef] [Green Version]
- Rawal, R.S.; Pangtey, Y.P.S. High Altitude Forests with Special Reference to The Timber Line in Kumaun, Central Himalaya; Pangtey, Y.P.S., Rawal, R.S., Eds.; Gyanodaya Prakashan: Nainital, India, 1994. [Google Scholar]
- Wang, Y.; Sylvester, S.P.; Lu, X.; Dawadi, B.; Sigdel, S.R.; Liang, E.; Camarero, J.J. The stability of spruce treelines on the eastern Tibetan Plateau over the last century is explained by pastoral disturbance. For. Ecol. Manag. 2019, 442, 34–45. [Google Scholar] [CrossRef]
- Lyu, L.; Zhang, Q.-B.; Deng, X.; Mäkinen, H. Fine-scale distribution of treeline trees and the nurse plant facilitation on the eastern Tibetan Plateau. Ecol. Indic. 2016, 66, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Camarero, J.J.; Gutiérrez, E. Pace and pattern of recent treeline dynamics: Response to ecotones to climate variability in the Spanish Pyrenees. Clim. Chang. 2004, 63, 181–200. [Google Scholar] [CrossRef]
- Batllori, E.; Gutierrez, E. Regional tree line dynamics in response to global change in the Pyrenees. J. Ecol. 2008, 96, 1275–1288. [Google Scholar] [CrossRef]
- Chhetri, P.K.; Cairns, D.M. Low recruitment above treeline indicates treeline stability under changing climate in Dhorpatan Hunting Reserve, Western Nepal. Phys. Geogr. 2018, 39, 329–342. [Google Scholar] [CrossRef]
- Chhetri, P.K.; Cairns, D.M. Contemporary and historic population structure of Abies spectabilis at treeline in Barun valley, eastern Nepal Himalaya. J. Mt. Sci. 2015, 12, 558–570. [Google Scholar] [CrossRef]
- Cook, E.R.; Kairiukstis, L.A. Methods of Dendrochronology: Applications in the Environmental Sciences; Springer: Dordrecht, The Netherlands, 1990; p. 394. [Google Scholar]
- Speer, J.H. Fundamentals of Tree Ring Research; Indiana State University: Terre Haute, IN, USA, 2010. [Google Scholar]
- Fritts, H.C. Tree Rings and Climate; University Press Cambridge: Cambridge, UK, 1976. [Google Scholar]
- Rinn, F. TSAP-Win User Reference Manual; Rinntech: Heidelberg, Germany, 2003. [Google Scholar]
- Holmes, R.L. Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull. 1983, 43, 69–78. [Google Scholar]
- Grissino-Mayer, H.D. Evaluating crossdating accuracy: A manual and tutorial for the computer program COFECHA. Tree-Ring Res. 2001, 57, 205–221. [Google Scholar]
- Stokes, M.A.; Smiley, T.L. An Introduction to Tree-Ring Dating; University of Chicago Press: Chicago, IL, USA, 1968. [Google Scholar]
- Norton, D.; Palmer, J.; Ogden, J. Dendroecological studies in New Zealand 1. An evaluation of tree age estimates based on increment cores. New Zealand J. Bot. 1987, 25, 373–383. [Google Scholar] [CrossRef]
- Kirdyanov, A.V.; Hagedorn, F.; Knorre, A.A.; Fedotova, E.V.; Vaganov, E.A.; Naurzbaev, M.M.; Moiseev, P.A.; Rigling, A. 20th century tree-line advance and vegetation changes along an altitudinal transect in the Putorana Mountains, northern Siberia. Boreas 2012, 41, 56–67. [Google Scholar] [CrossRef]
- Hagedorn, F.; Shiyatov, S.G.; Mazepa, V.S.; Devi, N.M.; Grigor’Ev, A.A.; Bartysh, A.A.; Fomin, V.V.; Kapralov, D.S.; Terent’Ev, M.; Bugman, H.; et al. Treeline advances along the Urals mountain range—Driven by improved winter conditions? Glob. Chang. Biol. 2014, 20, 3530–3543. [Google Scholar] [CrossRef]
- Brown, R. Response of Alpine Treeline Ecotones to 20th Century Climate Change: A Comparative Analysis from Kananaskis Country, Alberta; The University of Guelph: Guelph, ON, Canada, 2013. [Google Scholar]
- Millar, C.I.; Westfall, R.D.; Delany, D.L.; King, J.C.; Graumlich, L.J. Response of Subalpine Conifers in the Sierra Nevada, California, U.S.A., to 20th-Century Warming and Decadal Climate Variability. Arct. Antarct. Alp. Res. 2004, 36, 181–200. [Google Scholar] [CrossRef] [Green Version]
- Singh, R.B.; Schickhoff, U.; Mal, S. Climate Change, Glacier Response, and Vegetation Dynamics in the Himalaya; Springer: Geneva, Switzerland, 2016. [Google Scholar]
- Caccia, F.D.; Ballaré, C.L. Effects of tree cover, understory vegetation, and litter on regeneration of Douglas-fir (Pseudotsuga menziesii) in southwestern Argentina. Can. J. For. Res. 1998, 28, 683–692. [Google Scholar] [CrossRef]
- Wang, Y.; Camarero, J.J.; Luo, T.; Liang, E. Spatial patterns of Smith fir alpine treelines on the south-eastern Tibetan Plateau support that contingent local conditions drive recent treeline patterns. Plant Ecol. Divers. 2012, 5, 311–321. [Google Scholar] [CrossRef]
- Greenwood, S.; Chen, J.; Chen, C.; Jump, A.S. Temperature and sheltering determine patterns of seedling establishment in an advancing subtropical treeline. J. Veg. Sci. 2015, 26, 711–721. [Google Scholar] [CrossRef] [Green Version]
- Germino, M.J.; Smith, W.K. Sky exposure, crown architecture, and low-temperature photoinhibition in conifer seedlings at alpine treeline. Plant Cell Environ. 1999, 22, 407–415. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K. High resistance to low-temperature photoinhibition in two alpine, snowbank species. Physiol. Plant. 2000, 110, 89–95. [Google Scholar] [CrossRef]
- Germino, M.J.; Smith, W.K. Differences in microsite, plant form, and low-temperature photoinhibition in alpine plants. Arct. Antarct. Alp. Res. 2000, 32, 388–396. [Google Scholar] [CrossRef]
- Petit, G.; Anfodillo, T.; Carraro, V.; Grani, F.; Carrer, M. Hydraulic constraints limit height growth in trees at high altitude. New Phytol. 2011, 189, 241–252. [Google Scholar] [CrossRef] [PubMed]
- Smithers, B.V.; Alongi, F.; North, M.P. Live fast, die young: Climate shifts may favor Great Basin bristlecone pine or limber pine in sub-alpine forest establishment. For. Ecol. Manag. 2021, 494, 119339. [Google Scholar] [CrossRef]
- Shen, W.; Zhang, L.; Liu, X.; Luo, T. Seed-based treeline seedlings are vulnerable to freezing events in the early growing season under a warmer climate: Evidence from a reciprocal transplant experiment in the Sergyemla Mountains, southeast Tibet. Agric. For. Meteorol. 2014, 187, 83–92. [Google Scholar] [CrossRef]
- Smith, W.K.; Germino, M.J.; Hancock, T.E.; Johnson, D.M. Another perspective on altitudinal limits of alpine timberlines. Tree Physiol. 2003, 23, 1101–1112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.-B.; Ren, G.-Y.; Shrestha, A.B.; Ren, Y.-Y.; You, Q.-L.; Zhan, Y.-J.; Xu, Y.; Rajbhandari, R. Changes in extreme temperature events over the Hindu Kush Himalaya during 1961–2015. Adv. Clim. Chang. Res. 2017, 8, 157–165. [Google Scholar] [CrossRef]
- Renard, S.M.; McIntire, E.J.; Fajardo, A. Winter conditions—Not summer temperature—Influence establishment of seedlings at white spruce alpine treeline in Eastern Quebec. J. Veg. Sci. 2015, 27, 29–39. [Google Scholar] [CrossRef]
- Sigdel, S.R.; Liang, E.; Wang, Y.; Dawadi, B.; Camarero, J.J. Tree-to-tree interactions slow down Himalayan treeline shifts as inferred from tree spatial patterns. J. Biogeogr. 2020, 47, 1816–1826. [Google Scholar] [CrossRef]
- Feuillet, T.; Birre, D.; Milian, J.; Godard, V.; Clauzel, C.; Serrano-Notivoli, R. Spatial dynamics of alpine tree lines under global warming: What explains the mismatch between tree densification and elevational upward shifts at the tree line ecotone? J. Biogeogr. 2019, 47, 1056–1068. [Google Scholar] [CrossRef]
- Cavieres, L.A.; Badano, E.I.; Sierra-Almeida, A.; Molina-Montenegro, M.A. Microclimatic Modifications of Cushion Plants and Their Consequences for Seedling Survival of Native and Non-native Herbaceous Species in the High Andes of Central Chile. Arct. Antarct. Alp. Res. 2007, 39, 229–236. [Google Scholar] [CrossRef] [Green Version]
Figure 1.
Study area and the annual mean temperature (a), annual precipitation (b), and monthly mean temperature and total precipitation (c) at the nearest meteorological station in Zuogong County. The R2 and p values of fitted curves are given.
Figure 1.
Study area and the annual mean temperature (a), annual precipitation (b), and monthly mean temperature and total precipitation (c) at the nearest meteorological station in Zuogong County. The R2 and p values of fitted curves are given.

Figure 2.
Elevation positions of Balfour spruce adult trees, juveniles, and seedlings on the north- (a) and east-facing slopes (b). Red color represents adult trees, blue color represents juveniles, and green color represents seedlings. Circles stand for Balfour spruce; triangles for junipers. P.SD stands for Balfour spruce seedlings, P.SP for Balfour spruce saplings, P.T for Balfour spruce adult trees, J.SD for juniper seedlings, J.T for juniper adult trees. Yellow lines indicate averaged treeline position.
Figure 2.
Elevation positions of Balfour spruce adult trees, juveniles, and seedlings on the north- (a) and east-facing slopes (b). Red color represents adult trees, blue color represents juveniles, and green color represents seedlings. Circles stand for Balfour spruce; triangles for junipers. P.SD stands for Balfour spruce seedlings, P.SP for Balfour spruce saplings, P.T for Balfour spruce adult trees, J.SD for juniper seedlings, J.T for juniper adult trees. Yellow lines indicate averaged treeline position.

Figure 3.
Age structures of Balfour spruce on the north- (gray bars) and east-facing slopes (dark bars). The inset shows the currently existing trees during 1801–1930.
Figure 3.
Age structures of Balfour spruce on the north- (gray bars) and east-facing slopes (dark bars). The inset shows the currently existing trees during 1801–1930.

Figure 4.
Height (a,e), base diameter (b,f), height growth rate (c,g), and diameter growth rate (d,h) of Balfour spruce during 2001–2010 (upper panels) and 2010–2020 (bottom panels) on the north- and east-facing slopes. Different letters indicate significant differences at p < 0.05.
Figure 4.
Height (a,e), base diameter (b,f), height growth rate (c,g), and diameter growth rate (d,h) of Balfour spruce during 2001–2010 (upper panels) and 2010–2020 (bottom panels) on the north- and east-facing slopes. Different letters indicate significant differences at p < 0.05.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Characteristics of the study sites.
| Sites | N1 | N2 | N3 | E1 | E2 | E3 |
|---|---|---|---|---|---|---|
| Latitude | 30.63043° N | 30.6303° N | 30.63016° N | 30.68192° N | 30.68168° N | 30.68144° N |
| Longitude | 97.2853° E | 97.28554° E | 97.28584° E | 97.2447° E | 97.24485° E | 97.24498° E |
| Slope | 32° | 31° | 30° | 32° | 33° | 31° |
| Aspect | North | North | North | East | East | East |
| Length (m) | 378 | 380 | 390 | 150 | 150 | 150 |
| Width (m) | 30 | 30 | 30 | 30 | 30 | 30 |
| Forest line (m) | 4416 | 4419 | 4408 | 4470 | 4464 | 4457 |
| Treeline (m) | 4437 | 4453 | 4424 | 4487 | 4491 | 4470 |
| Species limit (m) | 4597 | 4594 | 4590 | 4508 | 4506 | 4504 |
| Seedlings | 75 | 48 | 49 | 18 | 25 | 20 |
| Juveniles | 126 | 157 | 126 | 39 | 34 | 25 |
| Adult trees | 84 | 89 | 105 | 22 | 22 | 13 |
| Treeline shift over the past 100 y (m) | 0 | 29 | 46 | —— | —— | 26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zheng, L.; Shi, P.; Zhou, T.; Hou, G.; Song, M.; Yu, F. Tree Regeneration Patterns on Contrasting Slopes at Treeline Ecotones in Eastern Tibet. Forests 2021, 12, 1605. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111605
AMA Style
Zheng L, Shi P, Zhou T, Hou G, Song M, Yu F. Tree Regeneration Patterns on Contrasting Slopes at Treeline Ecotones in Eastern Tibet. Forests. 2021; 12(11):1605. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111605
Chicago/Turabian StyleZheng, Lili, Peili Shi, Tiancai Zhou, Ge Hou, Minghua Song, and Feihai Yu. 2021. "Tree Regeneration Patterns on Contrasting Slopes at Treeline Ecotones in Eastern Tibet" Forests 12, no. 11: 1605. https://0-doi-org.brum.beds.ac.uk/10.3390/f12111605
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.