
Data Mining Methods to Detect Airborne Pollen of Spring Flowering Arboreal Taxa

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
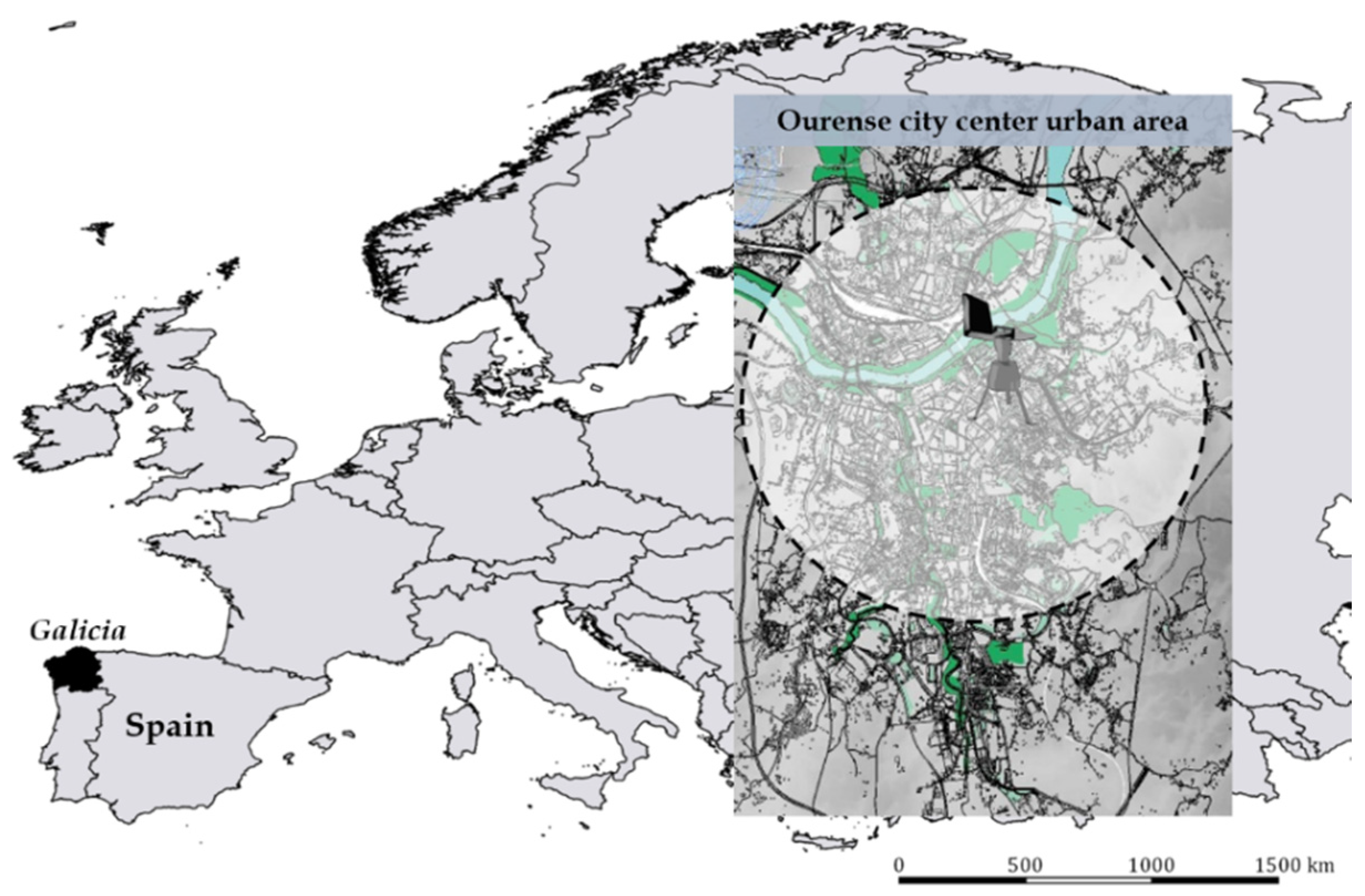
2.1. Study Area and Meteorological Data
2.2. Aerobiological Data
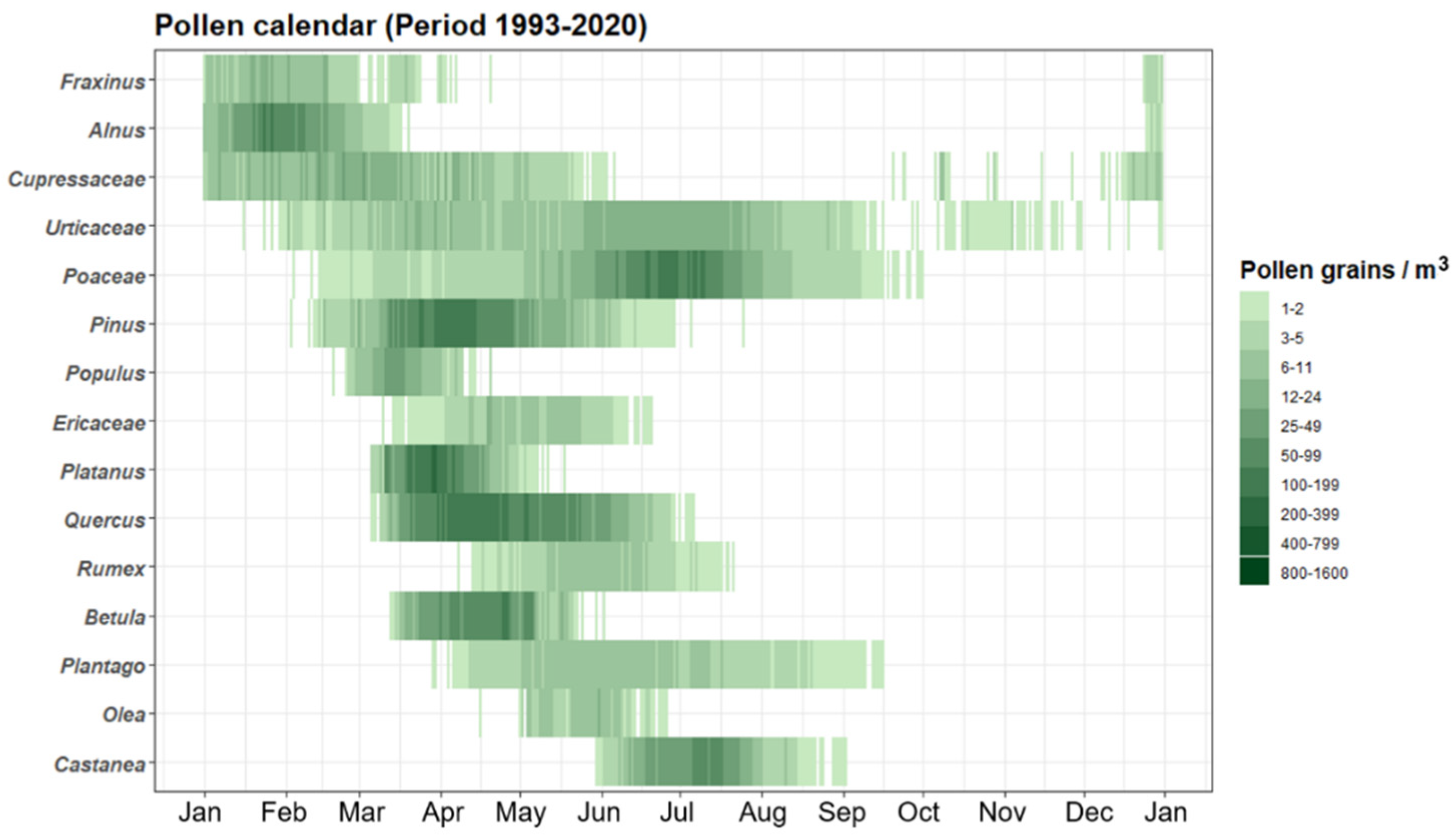
2.3. Pollen Calendar and Pollen Trends
2.4. Statistical Analysis
2.4.1. Correlation Analysis
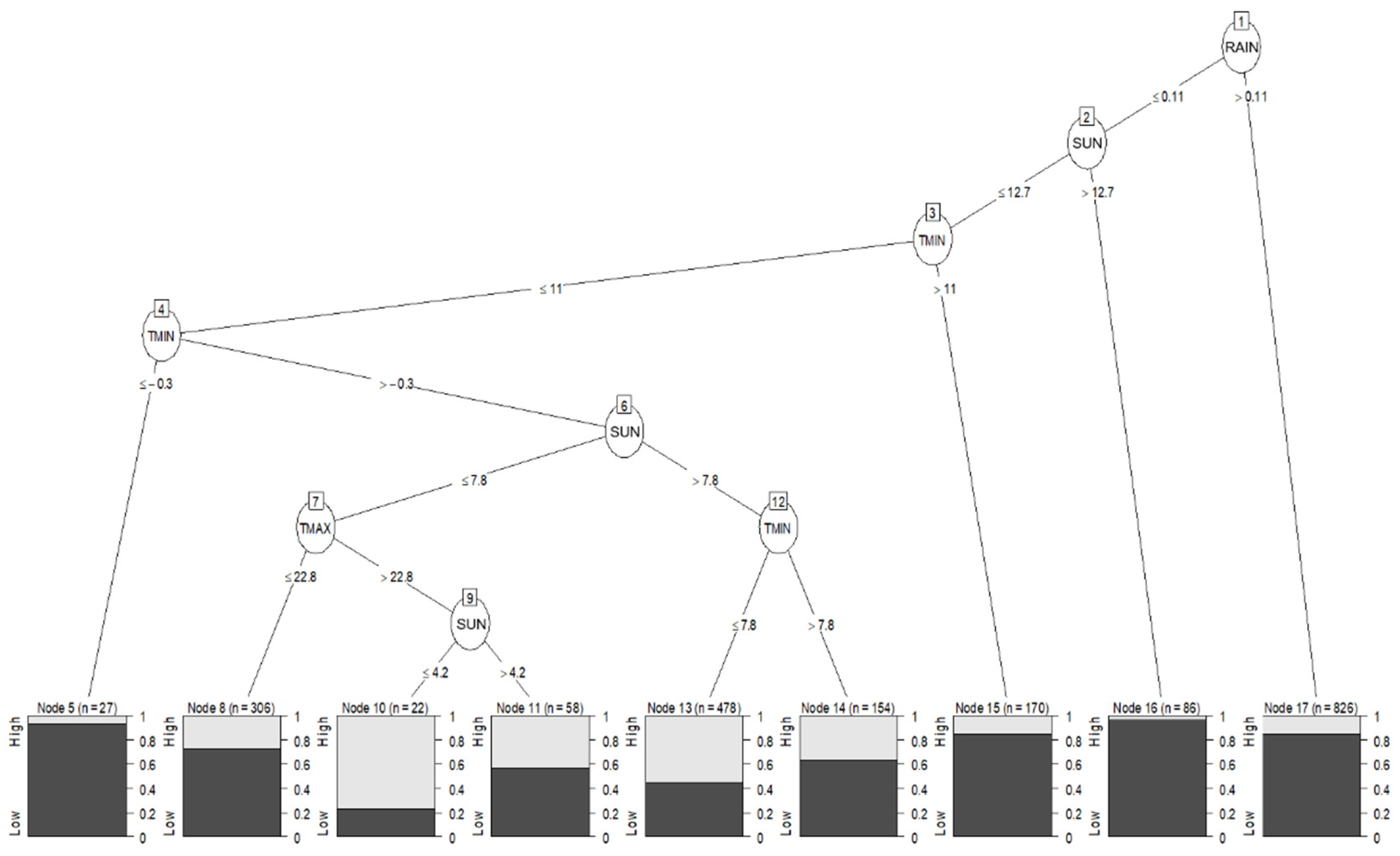
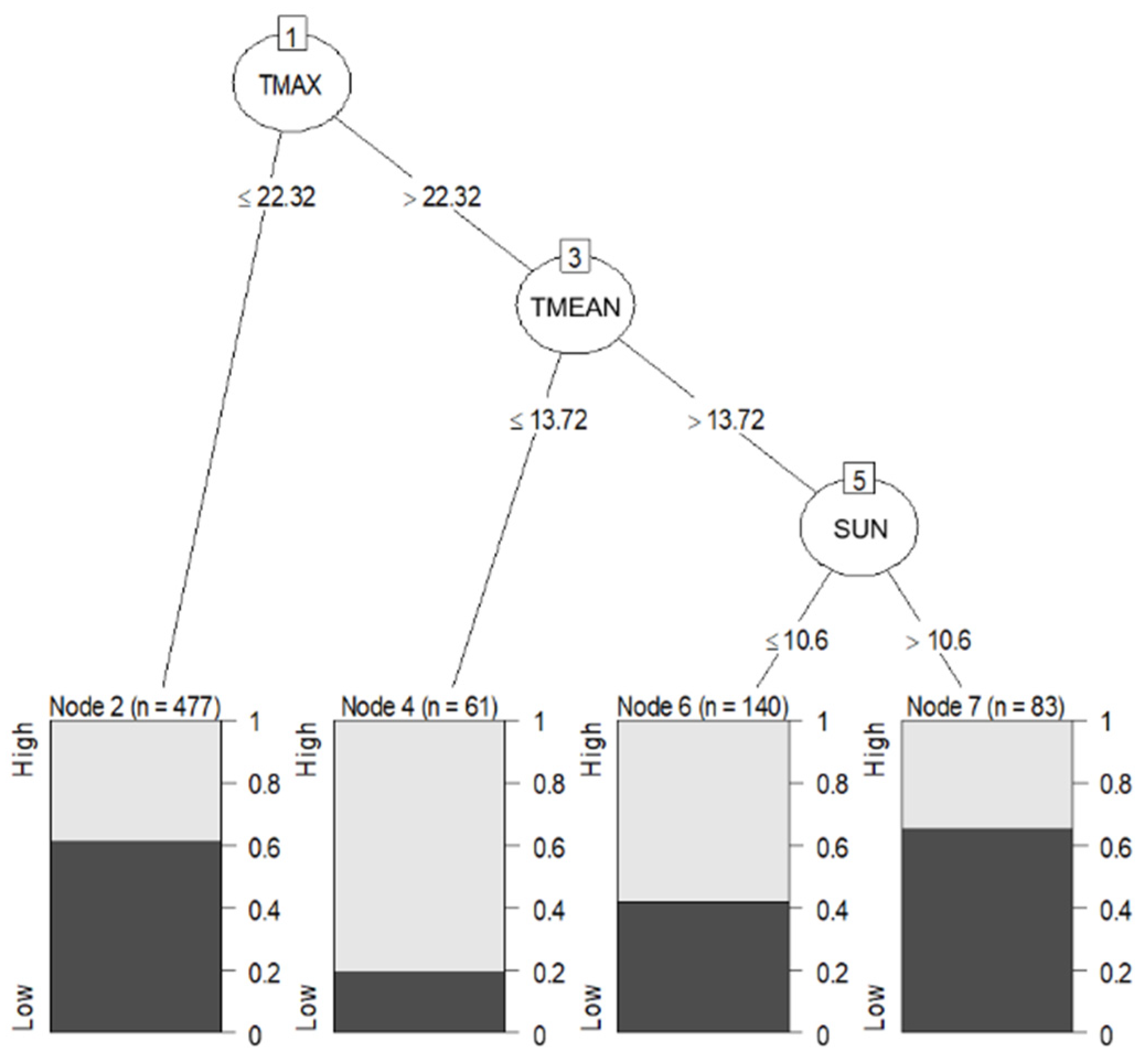
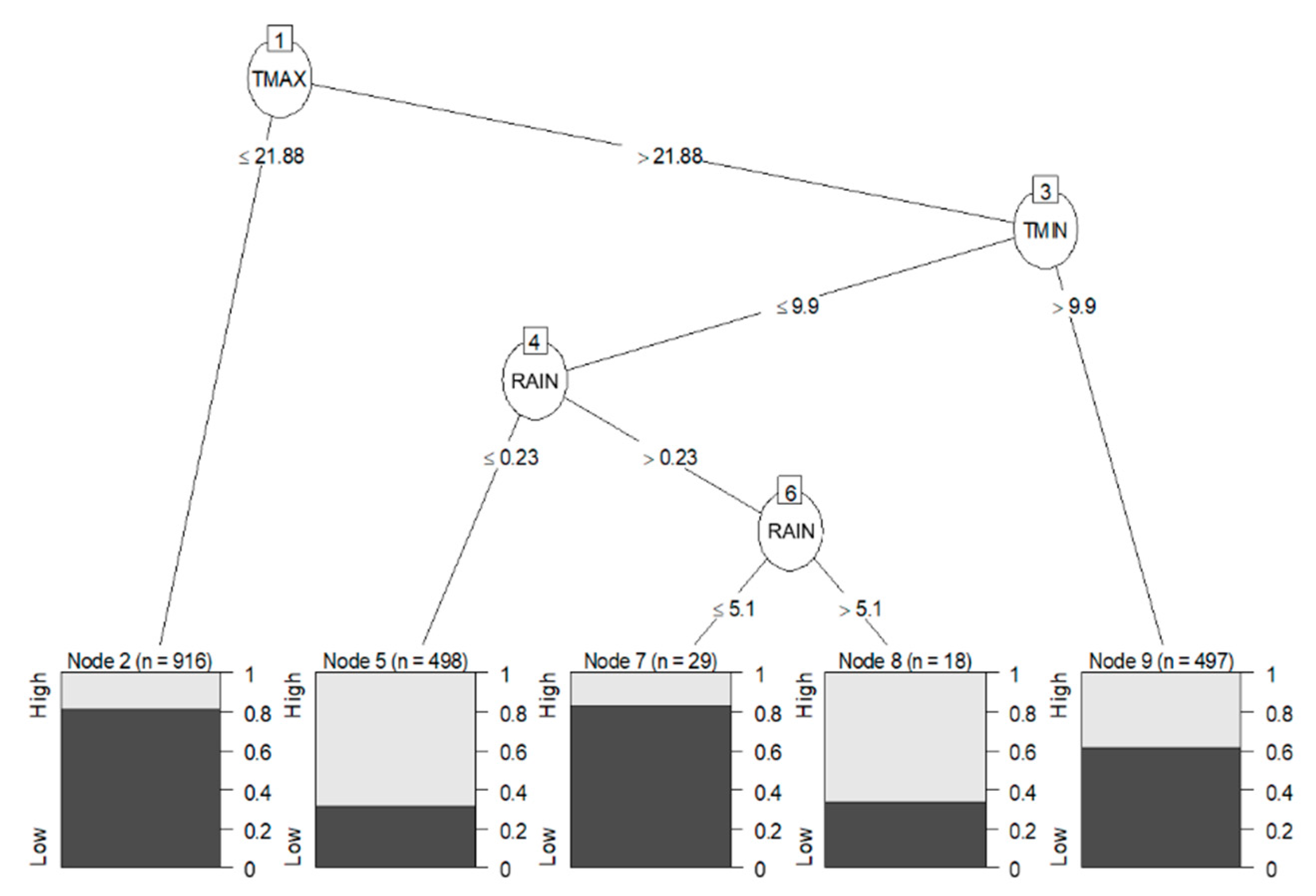
2.4.2. Data Mining Algorithm: C5.0 Decision Trees and Rule-Based Models
3. Results
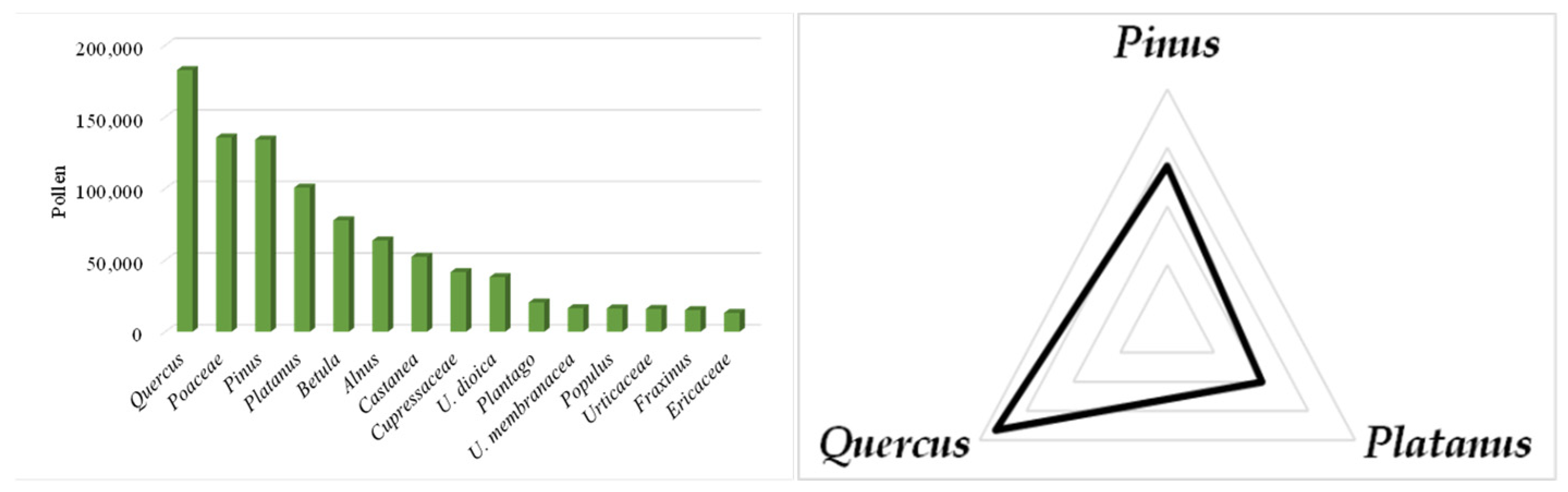
3.1. Pollen Seasonal Distribution
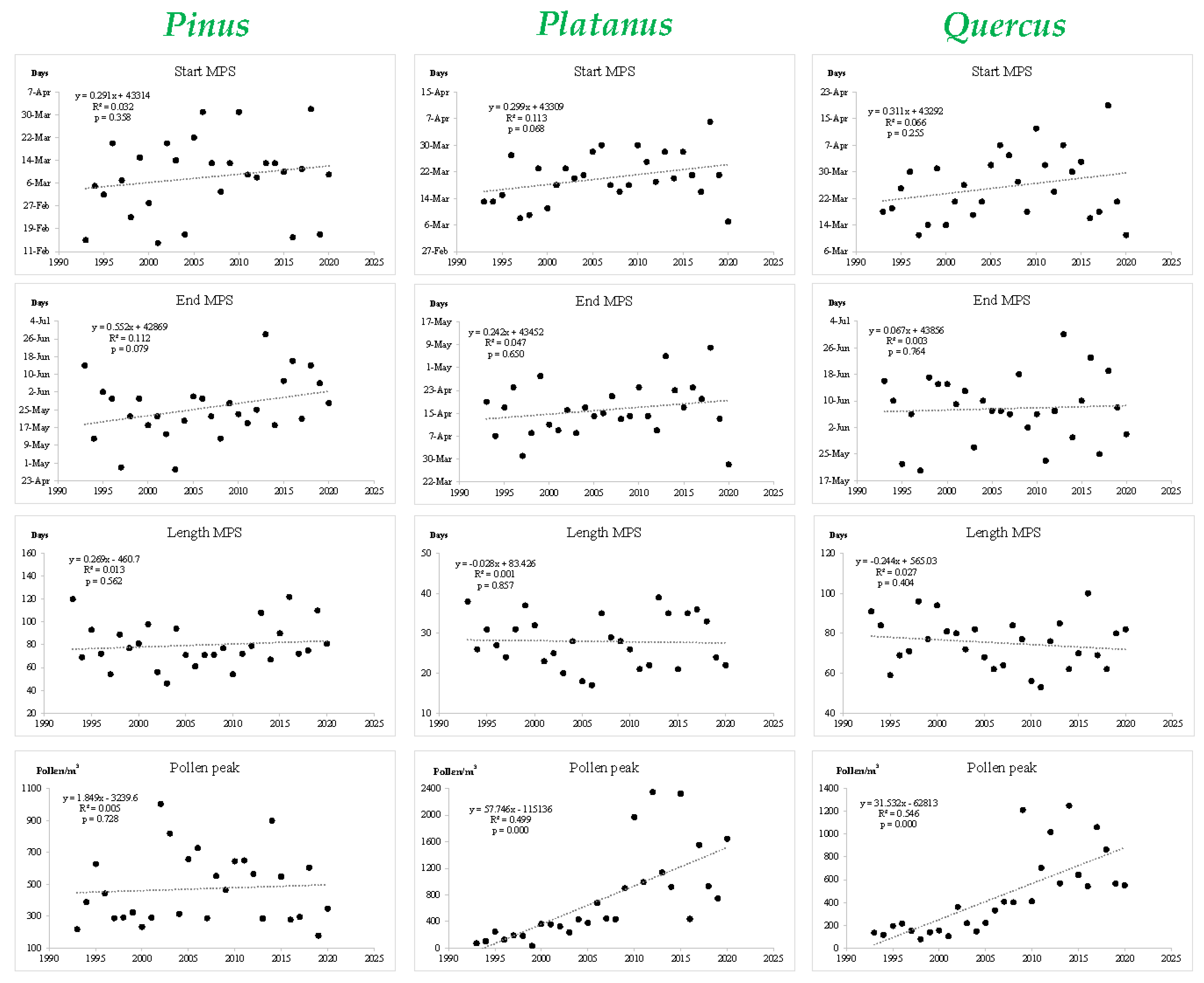
3.2. Main Seasonal Indexes of the Considered Pollen Types
3.3. Pollen Trends
3.4. Statistical Analysis: Influence of Meteorological Conditions
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Fernández-González, F.; Loidi, J.; Moreno, J.C. Impacts on Plant Biodiversity. Impacts of Climatic Change in Spain; Cambio climático: Impactos, vulnerabilidad y adaptación. Ministerio para la Transición Ecológica y el Reto Demográfico. Gobierno de España. pp. 179–242. Available online: https://www.miteco.gob.es/es/cambio-climatico/temas/impactos-vulnerabilidad-y-adaptacion/05_Impacts%20on%20plant%20biodiversity_ing_tcm30-178519.pdf (accessed on 28 June 2021).
- Lo, F.; Bitz, C.M.; Battisti, D.S.; Hess, J.J. Pollen calendars and maps of allergenic pollen in North America. Aerobiologia 2019, 35, 613–633. [Google Scholar] [CrossRef] [Green Version]
- Bertin, R.I. Plant Phenology and Distribution in Relation to Recent Climate Change. J. Torrey Bot. 2008, 135, 126–146. [Google Scholar] [CrossRef] [Green Version]
- Gordo, O.; Sanz, J.J. Impact of climate change on plant phenology in Mediterranean ecosystems. Glob. Chang. Biol. 2010, 16, 1082–1106. [Google Scholar] [CrossRef]
- Menzel, A. Trends in phenological phases in Europe between 1951 and 1996. Int. J. Biometeorol. 2000, 44, 76–81. [Google Scholar] [CrossRef]
- Walther, G.R.; Post, E.; Convey, P.; Menzel, A.; Parmesan, C.; Beebee, T.J.C.; Fromentin, J.M.; Hoegh-Guldberg, O.; Bairlein, F. Ecological Responses to Recent Climate Change. Nature 2002, 416, 389–395. [Google Scholar] [CrossRef]
- Menzel, A.; Sparks, T.H.; Estrella, N.; Koch, E.; Aasa, A.; Ahas, R.; Alm-Kübler, K.; Bissolli, P.; Braslavská, O.G.; Briede, A.; et al. European phenological response to climate change matches the warming pattern. Glob. Chang. Biol. 2006, 12, 1969–1976. [Google Scholar] [CrossRef]
- Schwartz, M.D.; Ahas, R.; Aasa, A. Onset of spring starting earlier across the Northern Hemisphere. Glob. Chang. Biol. 2006, 12, 343–351. [Google Scholar] [CrossRef]
- Anderegg, W.R.L.; Abatzoglou, J.T.; Anderegg, L.D.L.; Bielory, L.; Kinney, P.L.; Ziska, L. Anthropogenic climate change is worsening North American pollen seasons. PNAS 2021, 118, e2013284118. [Google Scholar] [CrossRef]
- Ziska, L.; Knowlton, K.; Rogers, C.A.; Dalan, D.; Tierney, N.; Elder, M.A.; Filley, W.; Shropshire, J.; Ford, L.B.; Hedberg, C.; et al. Recent warming by latitude associated with increased length of ragweed pollen season in central North America. Proc. Natl. Acad. Sci. USA 2011, 108, 4248–4251. [Google Scholar] [CrossRef] [Green Version]
- Ziska, L.; Makra, L.; Harry, S.K.; Bruffaerts, N.; Hendrickx, M.; Coates, F.; Saarto, A.; Thibaudon, M.; Oliver, G.; Damialis, A.; et al. Temperature-related changes in airborne allergenic pollen abundance and seasonality across the northern hemisphere: A retrospective data analysis. Lancet Planet. Health 2019, 3, e124–e131. [Google Scholar] [CrossRef] [Green Version]
- Frei, T.; Gassner, E. Climate change and its impact on birch pollen quantities and the start of the pollen season an example from Switzerland for the period 1969–2006. Int. J. Biometeorol. 2008, 52, 667–674. [Google Scholar] [CrossRef]
- Ziello, C.; Sparks, T.H.; Estrella, N.; Belmonte, J.; Bergmann, K.C.; Bucher, E.; Brighetti, M.A.; Damialis, A.; Detandt, M.; Galán, C.; et al. Changes to Airborne Pollen Counts across Europe. PLoS ONE 2012, 7, e34076. [Google Scholar] [CrossRef]
- Mimet, A.; Pellsier, V.; Quénol, H.; Aguejdad, R.; Dubreuil, V.; Rozé, H. Urbanisation induces early flowering: Evidence from Platanus acerifolia and Prunus cerasus. Int. J. Biometeorol. 2009, 53, 287–298. [Google Scholar] [CrossRef]
- Rodríguez-Rajo, F.J.; Fdez-Sevilla, D.; Stach, A.; Jato, V. Assessment between pollen seasons in areas with different urbanization level related to local vegetation sources and differences in allergen exposure. Aerobiologia 2010, 26, 1–14. [Google Scholar] [CrossRef]
- Pawankar, R. Allergic diseases and asthma: A global public health concern and call to action. World Allergy Organ. J. 2014, 7, 12. [Google Scholar] [CrossRef] [Green Version]
- Cariñanos, P.; Casares-Porcel, M. Urban Green Zones and related pollen allergy: A Review. Some guidelines for designing spaces with low allergy impact. Landsc. Urban Plan. 2011, 101, 205–214. [Google Scholar] [CrossRef]
- Von Döhren, P.; Haase, D. Ecosystem disservices research: A review of the state of the art with a focus on cities. Ecol. Indic. 2015, 52, 490–497. [Google Scholar] [CrossRef]
- Cariñanos, P.; Sánchez-Mesa, J.A.; Prieto-Baena, J.C.; López, A.; Guerra, F.; Moreno, C.; Domínguez, E.; Galán, C. Pollen allergy related to the area of residence in the city of Córdoba, south-west Spain. J. Environ. Monit. 2002, 4, 734–738. [Google Scholar] [CrossRef] [Green Version]
- Kasprzyk, I. Comparative study of seasonal and intradiurnal variation of airborne herbaceous pollen in urban and rural areas. Aerobiologia 2006, 22, 185–195. [Google Scholar] [CrossRef]
- Ziska, L.; Bunce, J.A.; Goins, E.W. Characterization of an urban-rural CO2/temperature gradient and associated changes in initial plant productivity during secondary succession. Oecologia 2004, 139, 454–458. [Google Scholar] [CrossRef]
- Oliveira, M.; Ribeiro, H.; Delgado, J.L.; Abreu, I. Seasonal and intradiurnal variation of allergenic fungal spores in urban and rural areas of the North of Portugal. Aerobiologia 2009, 25, 85–98. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Clark, J.S. Pollen production by Pinus taeda growing in elevated atmospheric CO2. Funct. Ecol. 2006, 20, 541–547. [Google Scholar] [CrossRef]
- Rogers, C.A.; Wayne, P.M.; Macklin, E.A.; Muilenberg, M.L.; Wagner, C.J.; Epstein, P.R.; Bazzaz, F.A. Interaction of the onset of spring and elevated atmospheric CO2 on ragweed (Ambrosia artemisiifolia L.) pollen production. Environ. Health Perspect. 2006, 114, 865–869. [Google Scholar] [CrossRef] [Green Version]
- Wan, S.; Yuan, T.; Bowdish, S.; Wallace, L.; Russell, S.D.; Luo, Y. Response of an allergenic species, Ambrosia psilostachya (Asteraceae), to experimental warming and clipping: Implications for public health. Am. J. Bot. 2002, 89, 1843–1846. [Google Scholar] [CrossRef] [Green Version]
- Damialis, A.; Fotiou, C.; Halley, J.M.; Vokou, D. Effects of environmental factors on pollen production in anemophilous woody species. Trees 2011, 25, 253–264. [Google Scholar] [CrossRef]
- Faegri, K.; Iversen, J. Textbook of Pollen Analysis, 4th ed.; Faegri, K., Kalland, P.E., Krzywinski, K., Eds.; Wiley: Chichester, UK, 1989. [Google Scholar]
- Moore, P.D.; Webb, J.A.; Collinson, M.E. Pollen Analysis, 2nd ed.; Blackwell Scientific Publications: London, UK, 1991; pp. 42–46, 181–182. [Google Scholar]
- Rodríguez-Guitián, M.A.; Ramil-Rego, P. Clasificaciones climáticas aplicadas a Galicia: Revisión desde una perspectiva biogeográfica. Recur. Rurais 2007, 1, 31–53. [Google Scholar] [CrossRef]
- MeteoGalicia 2021. Galician Institute for Meteorology and Oceanography, Santiago de Compostela, Spain. Environment, Territory and Infrastructure Department of Galician Regional Government. Available online: http://www.meteogalicia.gal/web/index.action (accessed on 12 July 2021).
- PXOM. Plan Xeral De Ordenación Municipal De Ourense; Concello de Ourense: Galicia, Spain, 2013. [Google Scholar]
- Galán, C.; Cariñanos, P.; Alcázar, P.; Domínguez, E. Spanish Aerobiology Network (REA): Management and Quality Manual; University of Córdoba Publication Service: Córdoba, Spain, 2007; p. 61. [Google Scholar]
- Hirst, J.M. An Automatic Volumetric Spore Trap. Ann. Appl. Biol. 1952, 39, 257–265. [Google Scholar] [CrossRef]
- Andersen, T.B. A model to predict the beginning of the pollen season. Grana 1991, 30, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Galán, C.; Ariatti, A.; Bonini, M.; Clot, B.; Crouzy, B.; Dahl, A.; Fernandez-González, D.; Frenguelli, G.; Gehrig, R.; Isard, S.; et al. Recommended terminology for aerobiological studies. Aerobiologia 2017, 33, 293–295. [Google Scholar] [CrossRef]
- Rojo, J.; Picornell, A.; Oteros, J. AeRobiology: The computational tool for biological data in the air. Methods in Ecology and Evolution. Methods Ecol. Evol. 2019, 10, 1371–1376. [Google Scholar] [CrossRef] [Green Version]
- R Core Team 2020. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 15 July 2021).
- Rojo, J.; Picornell, A.; Oteros, J. Package ‘Aerobiology’. A Computational Tool for Aerobiological Data; Version 2.0.1; CRAN Repository: A Coruña, Spain, 2019; CRAN Mirror Spain (A Coruña). [Google Scholar]
- Maimon, O.; Rokach, L. Data Mining and Knowledge Discovery Handbook, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2010. [Google Scholar]
- Maimon, O.; Rokach, L. Introduction to Supervised Methods. In Data Mining and Knowledge Discovery Handbook; Maimon, O., Rokach, L., Eds.; Springer: Boston, MA, USA, 2005. [Google Scholar]
- Kuhn, M.; Wetson, S.; Culp, M.; Coulter, N.; Quinlan, R. Package ‘C50’. C5.0 Decision Trees and Rule-Based Models; Version 0.1.4; CRAN Repository: A Coruña, Spain, 2021; CRAN mirror Spain (A Coruña). [Google Scholar]
- Pang, S.-L.; Gong, J.-Z. C5.0 Classification Algorithm and Application on Individual Credit Evaluation of Banks. Syst. Eng. Theory Pract. 2009, 29, 94–104. [Google Scholar] [CrossRef]
- Fernández-González, M.; Lara, B.; González-Fernández, E.; Rojo, J.; Pérez-Badia, R.; Rodríguez-Rajo, F.J. Pinus Pollen Emission Patterns in Different Bioclimatic Areas of the Iberian Peninsula. Forests 2021, 12, 688. [Google Scholar] [CrossRef]
- Richardson, D.M.; Rundel, P.W. Ecology and biogeography of Pinus: An introduction. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 3–49. [Google Scholar]
- De Linares, C.; Delgado, R.; Aira, M.J.; Alcázar, P.; Alonso-Pérez, S.; Boi, M.; Cariñanos, P.; Cuevas, E.; de la Guardia, C.D.; Elvira-Rendueles, B.; et al. Changes in the Mediterranean pine forest: Pollination patterns and annual trends of airborne pollen. Aerobiologia 2017, 33, 375–391. [Google Scholar] [CrossRef]
- San-Miguel-Ayanz, J.; De Rigo, D.; Caudullo, G.; Houston Durrant, T.; Mauri, A.; Tinner, W.; Ballian, D.; Beck, P.; Birks, H.J.B.; Eaton, E.; et al. European Atlas of Forest Tree Species; Publications Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Barberó, M.; Loisel, R.; Quézel, P.; Richardson, D.M.; Romane, F. Pines of the Mediterranean basin. In Ecology and Biogeography of Pinus; Richardson, D.M., Ed.; Cambridge University Press: Cambridge, UK, 1998; pp. 153–170. [Google Scholar]
- Sheffer, E. A review of the development of Mediterranean pine–oak ecosystems after land abandonment and afforestation: Are they novel ecosystems? Ann. For. Sci. 2012, 69, 429–443. [Google Scholar] [CrossRef] [Green Version]
- Jato, V.; Rodríguez-Rajo, F.J.; Fernández-González, M.; Aira, M.J. Assessment of Quercus flowering trends in NW Spain. Int. J. Biometeorol. 2015, 59, 517–531. [Google Scholar] [CrossRef]
- Vrinceanu, D.; Berghi, O.N.; Cergan, R.; Dumitru, M.; Ciuluvica, R.C.; Giurcaneanu, C.; Neagos, A. Urban allergy review: Allergic rhinitis and asthma with plane tree sensitization (Review). Exp. Ther. Med. 2021, 21, 275. [Google Scholar] [CrossRef] [PubMed]
- Christenhusz, M.; Byng, J. The number of known plants species in the world and its annual increase. Phytotaxa 2016, 261, 201–217. [Google Scholar] [CrossRef] [Green Version]
- Cariñanos, P.; Ruiz Peñuela, S.; Valle, A.M.; de la Guardia, C.D. Assessing pollination disservices of urban street trees: The case of London plane tree (Platanus x hispanica Mill. ex Münchh). Sci. Total Environ. 2020, 737, 139722. [Google Scholar] [CrossRef]
- Willis, K.J.; Petrokofsky, G. The natural capital of city trees. Science 2017, 356, 374–376. [Google Scholar] [CrossRef]
- Baldachini, C.; Castanheiro, A.; Maghakyan, N.; Sgrigna, G.; Verhelst, J.; Alonso, R.; Amorim, J.H.; Bellan, P.; Bojović, D.Đ.; Breuste, J.; et al. How does the amount and composition of PM deposited on Platanus acerifolia leaves change across different cities in Europe? Environ. Sci. Technol. 2017, 51, 1147–1156. [Google Scholar] [CrossRef]
- Iglesias, I.; Rodríguez-Rajo, F.J.; Méndez, J. Behavior of Platanus hispanica Pollen, an Important Spring Aeroallergen in Northwestern Spain. J. Investig. Allergol. Clin. Immunol. 2007, 17, 145–156. [Google Scholar]
- Fernández-González, M.; Guedes, A.; Abreu, I.; Rodríguez-Rajo, F.J. Pla a_1 aeroallergen immunodetection related to the airborne Platanus pollen content. Sci. Total Environ. 2013, 463–464, 855–860. [Google Scholar] [CrossRef]
- Maya-Manzano, J.M.; Fernández-Rodríguez, S.; Monroy-Colín, A.; Silva-Palacios, I.; Tormo-Molina, R.; Gonzalo-Garijo, A. Allergenic pollen of ornamental plane trees in a Mediterranean environment and urban planning as a prevention tool. Urban For. Urban Gree. 2017, 27, 352–362. [Google Scholar] [CrossRef]
- Fernández-González, M.; Ribeiro, H.; Pereira, J.R.S.; Rodríguez-Rajo, F.J.; Abreu, I. Assessment between Platanus pollen and Pla a 1 allergen in two cities of North-western Iberian Peninsula. Aerobiologia 2019, 35, 463–475. [Google Scholar] [CrossRef]
- Bousquet, J.; Heinzerling, L.; Bachert, C.; Papadopoulos, N.G.; Bousquet, P.J.; Burney, P.G.; Canonica, G.W.; Carlsen, K.H.; Cox, L.; Haahtela, T.; et al. Practical guide to skin prick tests in allergy to aeroallergens. Allergy 2012, 67, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Cipriani, F.; Tripodi, S.; Panetta, V.; Perna, S.; Potapova, E.; Dondi, A.; Bernardini, R.; Caffarelli, C.; Casani, A.; Cervone, R.; et al. Early molecular biomarkers predicting the evolution of allergic rhinitis and its comorbidities: A longitudinal multicenter study of a patient cohort. Pediatr. Allergy Immunol. 2019, 30, 325–334. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.R.; Oh, J.-W.; Woo, S.-Y.; Seo, Y.A.; Choi, Y.-J.; Kim, H.S.; Lee, W.Y.; Kim, B.-J. Does the increase in ambient CO2 concentration elevate allergy risks posed by oak pollen? Int. J. Biometeorol. 2018, 62, 1587–1594. [Google Scholar] [CrossRef]
- Ziska, L.; Caulfield, F. Rising CO2 and pollen production of common ragweed (Ambrosia artemisiifolia L.), a known allergy-inducing species: Implications for public health. Funct. Plant Biol. 2000, 27, 893–898. [Google Scholar] [CrossRef]
- Sharma, C.M.; Khanduri, V.P.; Chamoli, N. The influence of air temperature and humidity on the anthesis and anther dehiscence in P. roxburghhi at three different altitudes. Curr. Sci. 1998, 75, 761–763. [Google Scholar]
- Khanduri, V.P.; Sharma, C.M. Development of groups of male strobili, anthesis and microsporangium dehiscence in Pinus roxburghii. Grana 2000, 39, 169–174. [Google Scholar] [CrossRef]
- Kluska, K.; Piotrowicz, K.; Kasprzyk, I. The impact of rainfall on the diurnal patterns of atmospheric pollen concentrations. Agric. For. Meteorol. 2020, 291, 108042. [Google Scholar] [CrossRef]
- Ribeiro, H.; Cunha, M.; Abreu, I. Airborne pollen concentration in the region of Braga, Portugal, and its relationship with meteorological parameters. Aerobiologia 2003, 19, 21–27. [Google Scholar] [CrossRef]
- Alan, Ş.; Şahin, A.A.; Sarışahin, T.; Şahin, S.; Kaplan, A.; Pinar, N.M. The effect of geographical and climatic properties on grass pollen and Phl p 5 allergen release. Int. J. Biometeorol. 2018, 62, 1325–1337. [Google Scholar] [CrossRef] [PubMed]
- Bruffaerts, N.; De Smedt, T.; Delcloo, A.; Simons, K.; Hoebeke, L.; Verstraeten, C.; Van Nieuwenhuyse, A.; Packeu, A.; Hendrickx, M. Comparative long-term trend analysis of daily weather conditions with daily pollen concentrations in Brussels, Belgium. Int. J. Biometeorol. 2018, 62 (Suppl. 1), 483–491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rasmussen, A. The effects of climate change on the birch pollen season in Denmark. Aerobiologia 2002, 18, 253–265. [Google Scholar] [CrossRef]
- Green, B.J.; Dettman, M.; Yli-Panula, E.; Rutherford, S.; Simpson, R. Atmospheric Poaceae pollen frequencies and associations with meteorological parameters in Brisbane, Australia: A 5-year record, 1994–1999. Int. J. Biometeorol. 2004, 48, 172–178. [Google Scholar] [CrossRef] [PubMed]
- Janati, A.; Bouziane, H.; Trigo, M.M.; Kadiri, M.; Kazzaz, M. Poaceae pollen in the atmosphere of Tetouan (NW Morocco): Effect of meteorological parameters and forecast of daily pollen concentration. Aerobiologia 2017, 33, 517–528. [Google Scholar] [CrossRef]
- Peternel, R.; Srnec, L.; Culig, J.; Zaninovic, K.; Mitic, B.; Vukusic, I. Atmospheric pollen season in Zagreb (Croatia) and its relationship with temperature and precipitation. Int. J. Biometeorol. 2004, 48, 186–191. [Google Scholar] [CrossRef]
- Majeed, H.T.; Periago, C.; Alarcón, M.; Belmonte, J. Airborne pollen parameters and their relationship with meteorological variables in NE Iberian Peninsula. Aerobiologia 2018, 34, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Timerman, D.; Greene, D.F.; Urzay, J.; Ackerman, J.D. Turbulence-induced resonance vibrations cause pollen release in wind-pollinated Plantago lanceolata L. (Plantaginaceae). J. R. Soc. Interface 2014, 11, 20140866. [Google Scholar] [CrossRef] [Green Version]
- Green, B.J.; Yli-Panula, E.; Dettmann, M.; Rutherford, S.; Simpson, R. Airborne Pinus pollen in the atmosphere of Brisbane, Australia and relationships with meteorological parameters. Aerobiologia 2003, 19, 47–55. [Google Scholar] [CrossRef]
- García-Mozo, H.; Galán, C.; Gómez-Casero, M.T.; Domínguez, E. A comparative study of different temperature accumulation methods for predicting the start of the Quercus pollen season in Cordoba (South West Spain). Grana 2000, 39, 194–199. [Google Scholar] [CrossRef]
- Faust, M. Physiology of Temperate Zone Fruit Trees; John Wiley & Sons: New York, NY, USA, 1989; p. 338. [Google Scholar]
- Richardson, E.A.; Seeley, S.D.; Walker, D.R. A model for estimating the completion of rest for ‘Redhaven’ and ‘Elberta’ peach trees. HortScience 1974, 9, 331–332. [Google Scholar]
- Oke, T.R. Boundary Layer Climates, 2nd ed.; Routledge: London, UK, 1987; p. 464. [Google Scholar]
- Snider, J.L.; Oosterhuis, D.M. How does timing, duration and severity of heat stress influence pollen-pistil interactions in angiosperms? Plant Signal. Behav. 2011, 6, 930–933. [Google Scholar] [CrossRef] [Green Version]
- Galán, C.; García-Mozo, H.; Vázquez, L.; Ruiz-Valenzuela, L.; Díaz de la Guardia, C.; Trigo Pérez, M. Heat requirement for the onset of the Olea europaea L. pollen season in several places of Andalusia region and the effect of the expected future climate change. Int. J. Biometeorol. 2005, 49, 184–188. [Google Scholar] [CrossRef] [PubMed]
- Emberlin, J.; Jäeger, S.; Domínguez, E.; Galán, C.; Hodal, L.; Mandrioli, P.; Rantio-Lehtimäki, A.; Savage, M.; Spieksma, F.T.; Barlett, C. Temporal and geographical variations in grass pollen seasons in areas of western Europe: An analysis of season dates at sites of the European pollen information system. Aerobiologia 2000, 16, 373–379. [Google Scholar] [CrossRef]
- Jato, V.; Rodríguez-Rajo, F.J.; Alcázar, P.; De Nuntiis, P.; Galán, C.; Mandrioli, P. May the definition of pollen season influence aerobiological results? Aerobiologia 2006, 22, 13–25. [Google Scholar] [CrossRef] [Green Version]
- Picornell, A.; Oteros, J.; Ruiz-Mata, R.; Recio, M.; Trigo, M.M.; Martínez-Bracero, M.; Lara, B.; Serrano-García, A.; Galán, C.; García-Mozo, H.; et al. Methods for interpolating missing data in aerobiological databases. Environ. Res. 2021, 200, 111391. [Google Scholar] [CrossRef]
- Damialis, A.; Häring, F.; Gökkaya, M.; Rauer, D.; Reiger, M.; Bezold, S.; Bounas-Pyrros, N.; Eyerich, K.; Todorova, A.; Hammel, G.; et al. Human exposure to airborne pollen and relationships with symptoms and immune responses: Indoors versus outdoors, circadian patterns and meteorological effects in alpine and urban environments. STOTEN 2019, 653, 190–199. [Google Scholar] [CrossRef]
- Demain, J.G.; Choi, Y.-J.; Oh, J.-W. The Impact of Climate Change on the Pollen Allergy and Sporulation of Allergic Fungi. Curr. Treat. Options Allergy 2021, 8, 60–73. [Google Scholar] [CrossRef]
- Gilles, S.; Akdis, C.; Lauener, R.; Schmid-Grendelmeier, P.; Bieber, T.; Schäppi, G.; Traidl-Hoffmann, C. The role of environmental factors in allergy: A critical reappraisal. Exp. Dermatol. 2018, 27, 1193–1200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gökkaya, M.; Damialis, A.; Nussbaumer, T.; Beck, I.; Bounas-Pyrros, N.; Bezold, S.; Amisi, M.M.; Kolek, F.; Todorova, A.; Chaker, A.; et al. Defining biomarkers to predict symptoms in subjects with and without allergy under natural pollen exposure. J. Allergy Clin. Immunol. 2020, 146, 583–594.e6. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pollen Type | GROUP | Low | Moderate | High |
|---|---|---|---|---|
| Pinus | 4 | 1–50 | 51–200 | >200 |
| Platanus | 4 | 1–50 | 51–200 | >200 |
| Quercus | 4 | 1–50 | 51–200 | >200 |
| Pollen Type | Pinus | Platanus | Quercus |
|---|---|---|---|
| Start Date | 0.3 | 0.3 | 0.3 |
| End Date | 0.6 | 0.3 | 0.1 |
| Peak Date | 0.3 | 0.1 | 0.3 |
| Total Pollen | 106.9 * | 269.4 ** | 442 ** |
| Variables | Pinus | Platanus | Quercus |
|---|---|---|---|
| RAIN | −0.338 ** | −0.114 ** | −0.425 ** |
| TMAX | 0.224 ** | 0.146 ** | 0.408 ** |
| TMIN | −0.246 ** | −0.037 | −0.133 ** |
| TMEAN | 0.028 | 0.056 | 0.208 ** |
| SUN | 0.294 ** | 0.066 | 0.414 ** |
| DATASET | Pinus | Platanus | Quercus |
|---|---|---|---|
| Training | 74.802 | 64.160 | 72.803 |
| Validation | 71.267 | 51.282 | 73.684 |
| Entire | 74.277 | 62.228 | 73.266 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
González-Fernández, E.; Álvarez-López, S.; Piña-Rey, A.; Fernández-González, M.; Rodríguez-Rajo, F.J. Data Mining Methods to Detect Airborne Pollen of Spring Flowering Arboreal Taxa. Forests 2021, 12, 1801. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121801
González-Fernández E, Álvarez-López S, Piña-Rey A, Fernández-González M, Rodríguez-Rajo FJ. Data Mining Methods to Detect Airborne Pollen of Spring Flowering Arboreal Taxa. Forests. 2021; 12(12):1801. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121801
Chicago/Turabian StyleGonzález-Fernández, Estefanía, Sabela Álvarez-López, Alba Piña-Rey, María Fernández-González, and Francisco Javier Rodríguez-Rajo. 2021. "Data Mining Methods to Detect Airborne Pollen of Spring Flowering Arboreal Taxa" Forests 12, no. 12: 1801. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121801