
Variation of Soil Organic Carbon Density with Plantation Age and Initial Vegetation Types in the Liupan Mountains Areas of Northwest China


Abstract
:1. Introduction
2. Materials and Methods
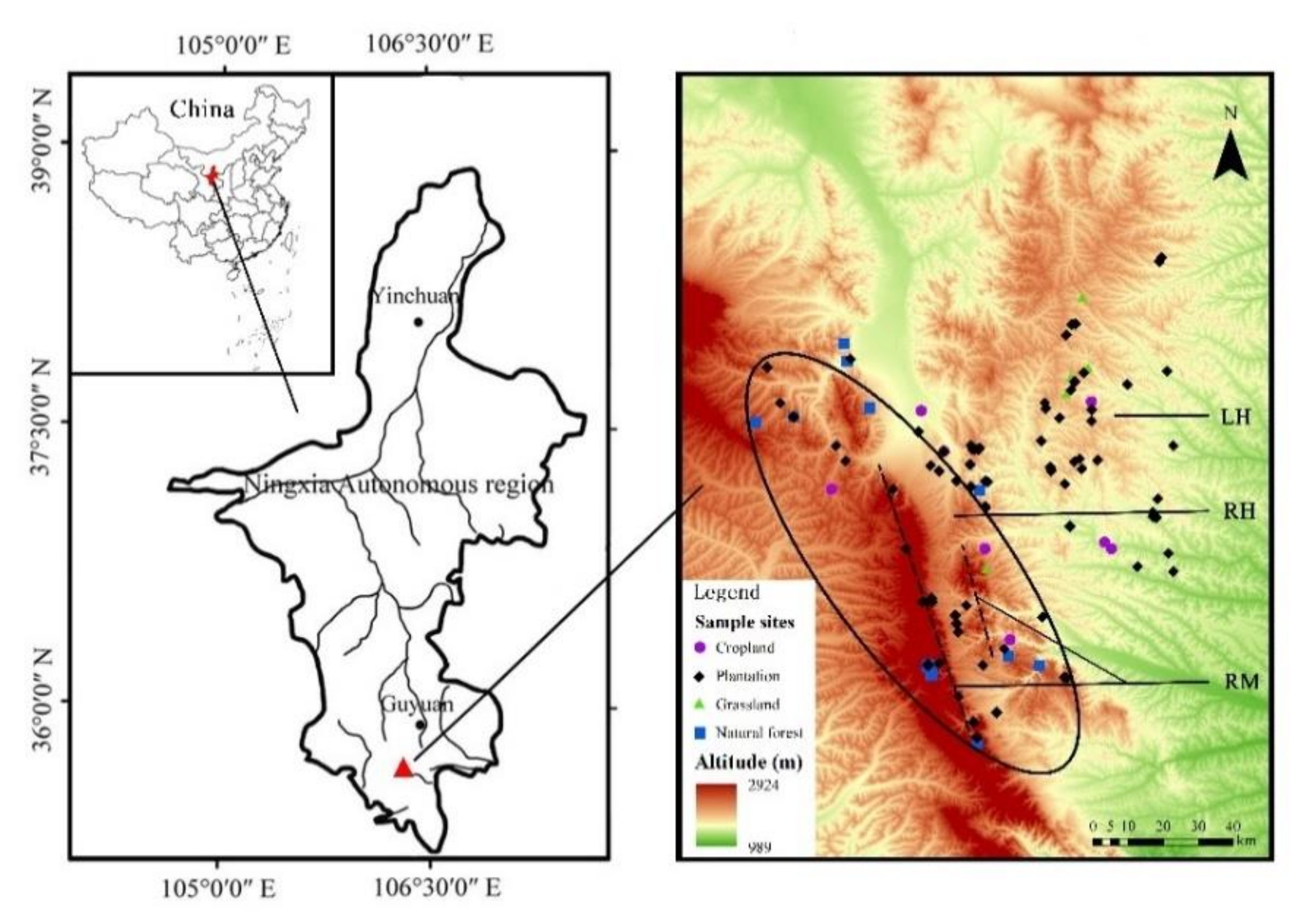
2.1. Study Area
2.2. Sample Plots Selection and Investigation
2.3. Soil Sampling and Analysis
2.4. Calculation and Correction of SOCD
2.5. The Functions of Temporal Variation of SOCD
2.6. TheΔSOCD Models Coupling the Effects of Multiple Factors
3. Results
3.1. Difference in SOCD among Sub-Areas and Vegetation Types
3.2. The Temporal Variation of SOCD
3.3. The Main Factors Influencing SOCD Changes
4. Discussion
4.1. The Difference in SOCD Due to Topographic Conditions
4.2. The Effects of Individual Factors on SOCD Change after Forestation
4.2.1. Initial SOCD
4.2.2. Organic Matter/Carbon Input
4.2.3. Climatic Factors
4.2.4. Altitude
4.3. Temporal Variation Patterns of SOCD after Forestation
4.4. Implications for the Assessment and Management of SOCD of Plantations
5. Conclusions
- (1)
- Wide variation of SOCD. The mean SOCD of main vegetation types follows the order of natural forests > grassland > plantation > cropland, and decreases from the sub-areas of RM, to RH, to LH with lowering altitudes and annual precipitation.
- (2)
- The important role of initial SOCD. The variation of SOCD is strongly affected by the counteracting processes of SOCD loss, mainly due to SOC mineralization and the SOCD input mainly from the litter fall of vegetation, besides the effects of pre-forestation soil processing. Higher initial SOCD can lead to a more obvious SOCD loss in the early periods after forestation and a delayed and lowered net SOCD increase after forestation. In average, the forestation mode from cropland to plantation presents a sustain increase of SOCD within 0–40 years after forestation because of the negligible SOCD loss due to low initial SOCD, while the forestation modes from grassland or natural forests to plantation presents firstly a slight or obvious decrease and then an increase of SOCD within 0−40 years because of the less significant or significant SOCD loss due to the relatively high or high initial SOCD.
- (3)
- Many factors to be considered. The site-quality-dependent tree growth, the SOCD response pattern, and the net SOCD increase after forestation are also greatly affected by the initial vegetation and site-specific initial SOCD, SOCD capacity, and their differences. All these points should be considered for a precise assessment and management of the SOCD of plantations.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| SOC | Soil organic carbon |
| SOCD | Soil organic carbon density |
| ΔSOCD | Soil organic carbon density change |
| MAT | Mean annual air temperature |
| MAP | Mean annual precipitation |
| RM | Rocky mountainous sub-area |
| RH | Rocky hilly sub-area |
| LH | Loess hilly sub-area |
| NP | Natural-forests-to-plantation |
| GP | Grassland-to-plantation |
| CP | Cropland-to-plantation |
| Bio | Biomass of trees |
| SOCD0 | Initial soil organic carbon density |
References
- Tang, X.; Zhao, X.; Bai, Y.; Tang, Z.; Wang, W.; Zhao, Y.; Wan, H.; Xie, Z.; Shi, X.; Wu, B.; et al. Carbon Pools in China’s Terrestrial Ecosystems: New Estimates Based on an Intensive Field Survey. Proc. Natl. Acad. Sci. USA 2018, 115, 4021–4026. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.; Park, T.; Wang, X.; Piao, S.; Xu, B.; Chaturvedi, R.K.; Fuchs, R.; Brovkin, V.; Ciais, P.; Fensholt, R.; et al. China and India Lead in Greening of the World through Land-Use Management. Nat. Sustain. 2019, 2, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Shao, M.; Wang, Y. Effect of Environmental Factors on Regional Soil Organic Carbon Stocks across the Loess Plateau Region, China. Agric. Ecosyst. Environ. 2011, 142, 184–194. [Google Scholar] [CrossRef]
- Wang, J.; Fu, B.; Qiu, Y.; Chen, L. Analysis on Soil Nutrient Characteristics for Sustainable Land Use in Danangou Catchment of the Loess Plateau, China. Catena 2003, 54, 17–29. [Google Scholar] [CrossRef]
- Zhou, G.; Liu, S.; Li, Z.; Zhang, D.; Tang, X.; Zhou, C.; Yan, J.; Mo, J. Old-Growth Forests Can Accumulate Carbon in Soils. Science 2006, 314, 1417. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Huang, Y. Determinants of Soil Organic Carbon Sequestration and Its Contribution to Ecosystem Carbon Sinks of Planted Forests. Glob. Change Biol. 2020, 26, 3163–3173. [Google Scholar] [CrossRef]
- Detwiler, R.P. Land Use Change and the Global Carbon Cycle: The Role of Tropical Soils. Biogeochemistry 1986, 2, 67–93. [Google Scholar] [CrossRef]
- Don, A.; Schumacher, J.; Freibauer, A. Impact of Tropical Land-Use Change on Soil Organic Carbon Stocks—A Meta-Analysis. Glob. Change Biol. 2011, 17, 1658–1670. [Google Scholar] [CrossRef] [Green Version]
- Poeplau, C.; Don, A.; Vesterdal, L.; Leifeld, J.; Van Wesemael, B.; Schumacher, J.; Gensior, A. Temporal Dynamics of Soil Organic Carbon after Land-Use Change in the Temperate Zone—Carbon Response Functions as a Model Approach. Glob. Change Biol. 2011, 17, 2415–2427. [Google Scholar] [CrossRef]
- Friggens, N.L.; Hester, A.J.; Mitchell, R.J.; Parker, T.C.; Subke, J.; Wookey, P.A. Tree Planting in Organic Soils Does Not Result in Net Carbon Sequestration on Decadal Timescales. Glob. Change Biol. 2020, 26, 5178–5188. [Google Scholar] [CrossRef]
- Wang, K.; Deng, L.; Ren, Z.; Shi, W.; Chen, Y.; Shang-Guan, Z. Dynamics of Ecosystem Carbon Stocks during Vegetation Restoration on the Loess Plateau of China. J. Arid Land 2016, 8, 207–220. [Google Scholar] [CrossRef] [Green Version]
- Cao, Z.; Fang, X.; Xiang, W.; Lei, P.; Peng, C. The Vertical Differences in the Change Rates and Controlling Factors of Soil Organic Carbon and Total Nitrogen along Vegetation Restoration in a Subtropical Area of China. Sustainability 2020, 12, 6443. [Google Scholar] [CrossRef]
- Kirschbaum, M.U.F.; Guo, L.; Gifford, R.M. Observed and Modelled Soil Carbon and Nitrogen Changes after Planting a Pinus Radiata Stand onto Former Pasture. Soil Biol. Biochem. 2008, 40, 247–257. [Google Scholar] [CrossRef]
- Wang, H.; Yue, C.; Mao, Q.; Zhao, J.; Ciais, P.; Li, W.; Yu, Q.; Mu, X. Vegetation and Species Impacts on Soil Organic Carbon Sequestration Following Ecological Restoration over the Loess Plateau, China. Geoderma 2020, 371, 114389. [Google Scholar] [CrossRef]
- Li, W.; Ciais, P.; Guenet, B.; Peng, S.; Chang, J.; Chaplot, V.; Khudyaev, S.; Peregon, A.; Piao, S.; Wang, Y.; et al. Temporal Response of Soil Organic Carbon after Grassland-Related Land-Use Change. Glob. Change Biol. 2018, 24, 4731–4746. [Google Scholar] [CrossRef] [PubMed]
- Tian, A.; Wang, Y.; Webb, A.A.; Liu, Z.; Ma, J.; Yu, P.; Wang, X. Water Yield Variation with Elevation, Tree Age and Density of Larch Plantation in the Liupan Mountains of the Loess Plateau and Its Forest Management Implications. Sci. Total. Environ. 2021, 752, 141752. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.B. Status and Countermeasures of Forest Resources Construction in Yuanzhou District of Guyuan City. Mod. Agric. Sci. Technol. 2018, 8, 165–166, (In Chinese with English abstract). Available online: http://en.cnki.com.cn/Article_en/CJFDTotal-ANHE201808106.htm (accessed on 18 October 2021).
- Pi, H.Y.; Zhang, M.X.; Xia, J.X. Changes in Land Use and Soil Erosion in the Loess Plateau Driven by the Grain for Green Program: A Case Study of Jingyuan Ningxia. J. Basic Sci. Eng. 2020, 28, 522–534, (In Chinese with English abstract). [Google Scholar] [CrossRef]
- Huaizhu, L. On the Current Situation and Development Plan of Mixed Needle-Broad Water Containment Forest Project Construction in Liupan Mountain Area of Ningxia. Ningxia J. Agric. For. Sci. Technol. 1999, 3, 23–25. (In Chinese). Available online: http://www.cnki.com.cn/Article/CJFDTOTAL-NXNL903.006.htm (accessed on 18 October 2021).
- Wang, F.; Pan, X.; Gerlein-Safdi, C.; Cao, X.; Wang, S.; Gu, L.; Wang, D.; Lu, Q. Vegetation Restoration in Northern China: A Contrasted Picture. Land Degrad. Dev. 2020, 31, 669–676. [Google Scholar] [CrossRef]
- Scheu, S.; Wolters, V. Influence of fragmentation and bioturbation on the decomposition of C-14-labeled beech leaf litter. Soil Biol. Biochem. 1991, 23, 1029–1034. [Google Scholar] [CrossRef]
- Rethemeyer, J.; Kramer, C.; Gleixner, G.; John, B.; Yamashita, T.; Flessa, H.; Andersen, N.; Nadeau, M.-J.; Grootes, P.M. Transformation of Organic Matter in Agricultural Soils: Radiocarbon Concentration versus Soil Depth. Geoderma 2005, 128, 94–105. [Google Scholar] [CrossRef]
- Guo, L.B.; Gifford, R.M. Soil Carbon Stocks and Land Use Change: A Meta Analysis. Glob. Change Biol. 2002, 8, 345–360. [Google Scholar] [CrossRef]
- Chang, R.; Fu, B.; Liu, G.; Wang, S.; Yao, X. The Effects of Afforestation on Soil Organic and Inorganic Carbon: A Case Study of the Loess Plateau of China. Catena 2012, 95, 145–152. [Google Scholar] [CrossRef]
- Lal, R.; Kimble, J.M. Importance of soil bulk density and methods of its importance. In Assessment Methods for Soil Carbon; Lal, R., Kimble, J.M., Follell, R.F., Stewart, B.A., Eds.; Taylor & Francis: London, UK, 2000; pp. 31–44. [Google Scholar]
- Henderson, D.; Ellert, B.; Naeth, M.A. Grazing and soil carbon along a gradient of Alberta rangeland. J. Range Manage. 2004, 57, 402–410. [Google Scholar] [CrossRef]
- Piñeiro, G.; Paruelo, J.M.; Jobbágy, E.G.; Jackson, R.B.; Oesterheld, M. Grazing Effects on Belowground C and N Stocks along a Network of Cattle Exclosures in Temperate and Subtropical Grasslands of South America. Glob. Biogeochem. Cycles 2009, 23, GB2003. [Google Scholar] [CrossRef] [Green Version]
- Sumfleth, K.; Duttmann, D. Prediction of soil property distribution in paddy soil landscapes using terrain data and satellite information as indicators. Ecol. Indic. 2008, 8, 485–501. [Google Scholar] [CrossRef]
- Zhang, W.; Wang, K.; Chen, H.; He, X.; Zhang, J. Ancillary Information Improves Kriging on Soil Organic Carbon Data for a Typical Karst Peak Cluster Depression Landscape. J. Sci. Food Agric. 2012, 92, 1094–1102. [Google Scholar] [CrossRef]
- Hartley, I.P.; Ineson, P. Substrate quality and the temperature sensitivity of soil organic matter decomposition. Soil Biol. Biochem. 2008, 40, 1567–1574. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.; Liski, J.; Chang, R.Y.; Akujärvi, A.; Wu, X.; Jin, T.T.; Wang, Y.F.; Fu, B.J. Soil Organic Carbon Dynamics of Black Locust Plantations in the Middle Loess Plateau Area of China. Biogeosciences 2013, 10, 7053–7063. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Liu, G.-B.; Xu, M.-X.; Zhang, J.; Wang, Y.; Tang, L. Temporal and Spatial Variations in Soil Organic Carbon Sequestration Following Revegetation in the Hilly Loess Plateau, China. Catena 2012, 99, 26–33. [Google Scholar] [CrossRef]
- Dou, X.; Deng, Q.; Li, M.; Wang, W.; Zhang, Q.; Cheng, X. Reforestation of Pinus Massoniana Alters Soil Organic Carbon and Nitrogen Dynamics in Eroded Soil in South China. Ecol. Eng. 2013, 52, 154–160. [Google Scholar] [CrossRef]
- Deng, L.; Liu, G.; Shangguan, Z. Land-Use Conversion and Changing Soil Carbon Stocks in China’s ‘Grain-for-Green’ Program: A Synthesis. Glob. Change Biol. 2014, 20, 3544–3556. [Google Scholar] [CrossRef] [PubMed]
- Hong, S.; Yin, G.; Piao, S.; Dybzinski, R.; Cong, N.; Li, X.; Wang, K.; Peñuelas, J.; Zeng, H.; Chen, A. Divergent Responses of Soil Organic Carbon to Afforestation. Nat. Sustain. 2020, 3, 694–700. [Google Scholar] [CrossRef]
- Don, A.; Rebmann, C.; Kolle, O.; Scherer-Lorenzen, M.; Schulze, E.-D. Impact of Afforestation-Associated Management Changes on the Carbon Balance of Grassland. Glob. Change Biol. 2009, 15, 1990–2002. [Google Scholar] [CrossRef]
- Laganière, J.; Angers, D.A.; Paré, D. Carbon Accumulation in Agricultural Soils after Afforestation: A Meta-Analysis. Glob. Change Biol. 2010, 16, 439–453. [Google Scholar] [CrossRef]
- Song, Y.; Zhai, J.; Zhang, J.; Qiao, L.; Wang, G.; Ma, L.; Xue, S. Forest Management Practices of Pinus Tabulaeformis Plantations Alter Soil Organic Carbon Stability by Adjusting Microbial Characteristics on the Loess Plateau of China. Sci. Total. Environ. 2021, 766, 144209. [Google Scholar] [CrossRef]
- Deng, L.; Shangguan, Z.; Sweeney, S. “Grain for Green” Driven Land Use Change and Carbon Sequestration on the Loess Plateau, China. Sci. Rep. 2015, 4, 7039. [Google Scholar] [CrossRef]
- Xiao, H.B.; Li, Z.W.; Chang, X.F.; Huang, B.; Nie, X.D.; Liu, C.; Liu, L.; Wang, D.Y.; Jiang, J.Y. The mineralization and sequestration of organic carbon in relation to agricultural soil erosion. Geoderma 2018, 329, 73–81. [Google Scholar] [CrossRef]
- Davidson, E.A.; Janssens, I.A. Temperature sensitivity of soil carbon decomposition and feedbacks to climate change. Nature 2006, 440, 165–173. [Google Scholar] [CrossRef]
- Oren, R.; Ellsworth, D.S.; Johnsen, K.H.; Phillips, N.; Ewers, B.E.; Maier, C.; Schäfer, K.V.R.; McCarthy, H.; Hendrey, G.; McNulty, S.G.; et al. Soil Fertility Limits Carbon Sequestration by Forest Ecosystems in a CO2-Enriched Atmosphere. Nature 2001, 411, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Huang, M.; Hou, J.; Tang, X.L.; Hao, M. Response of vegetation and soil carbon accumulation rate for China’s mature forest on climate change. Chin. J. Plant. Ecol. 2016, 40, 416–424, (In Chinese with English abstract). [Google Scholar] [CrossRef] [Green Version]
- Post, W.M.; Kwon, K.C. Soil carbon sequestration and landuse change: Processes and potential. Glob. Change Biol. 2010, 6, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Thornley, J. Temperate Grassland Responses to Climate Change: An Analysis Using the Hurley Pasture Model. Ann. Bot. 1997, 80, 205–221. [Google Scholar] [CrossRef] [Green Version]
- Trumbore, S.E.; Chadwick, O.A.; Amundson, R. Rapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change. Science 1996, 272, 393–396. [Google Scholar] [CrossRef] [Green Version]
- Garten, C.T.; Post, W.M.; Hanson, P.J.; Cooper, L.W. Forest soil carbon inventories and dynamics along an elevation gradient in the southern Appalachian Mountains. Biogeochemistry 1999, 45, 115–162. [Google Scholar] [CrossRef]
- Lal, R.; Follett, R.F.; Stewart, B.A.; Kimble, J.M. Soil Carbon Sequestration to Mitigate Climate Change and Advance Food Security. Soil Sci. 2007, 172, 943–956. [Google Scholar] [CrossRef]
- Tan, Z.X.; Lal, R.; Semeck, N.E.; Calhoun, F.G. Relationships between surface soil organic carbon pool and site variables. Geoderma 2004, 121, 187–195. [Google Scholar] [CrossRef]
- Dai, W.; Huang, Y. Relation of soil organic matter concentration to climate and altitude in zonal soils of China. Catena 2006, 65, 87–94. [Google Scholar] [CrossRef]
- Martínez-Mena, M.; Rogel, J.A.; Castillo, V.; Albaladejo, J. Organic Carbon and Nitrogen Losses Influenced by Vegetation Removal in a Semiarid Mediterranean Soil. Biogeochemistry 2002, 61, 309–321. [Google Scholar] [CrossRef]
- Deng, L.; Wang, K.-B.; Chen, M.-L.; Shangguan, Z.-P.; Sweeney, S. Soil Organic Carbon Storage Capacity Positively Related to Forest Succession on the Loess Plateau, China. Catena 2013, 110, 1–7. [Google Scholar] [CrossRef]
- Kuzyakov, Y.; Friedel, J.K.; Stahr, K. Review of Mechanisms and Quantification of Priming Effects. Soil Biol. Biochem. 2000, 32, 1485–1498. [Google Scholar] [CrossRef]
- Kuzyakov, Y. Priming Effects: Interactions between Living and Dead Organic Matter. Soil Biol. Biochem. 2010, 42, 1363–1371. [Google Scholar] [CrossRef]
- Ritter, E. Carbon, Nitrogen and Phosphorus in Volcanic Soils Following Afforestation with Native Birch (Betula Pubescens) and Introduced Larch (Larix Sibirica) in Iceland. Plant. Soil 2007, 295, 239–251. [Google Scholar] [CrossRef]
- Thuille, A.; Schulze, E.-D. Carbon Dynamics in Successional and Afforested Spruce Stands in Thuringia and the Alps. Glob. Change Biol. 2006, 12, 325–342. [Google Scholar] [CrossRef]
- Zhu, J.; Hu, H.; Tao, S.; Chi, X.; Li, P.; Jiang, L.; Ji, C.; Zhu, J.; Tang, Z.; Pan, Y.; et al. Carbon Stocks and Changes of Dead Organic Matter in China’s Forests. Nat. Commun. 2017, 8, 151. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Land Use, Land-Use Change, and Forestry; Cambridge University Press: Cambridge, UK, 2000. [Google Scholar]
- Bastin, J.-F.; Finegold, Y.; Garcia, C.; Mollicone, D.; Rezende, M.; Routh, D.; Zohner, C.M.; Crowther, T.W. The Global Tree Restoration Potential. Science 2019, 365, 76–79. [Google Scholar] [CrossRef] [PubMed]
- Bedison, J.E.; Scatena, F.N.; Mead, J.V. Influences on the Spatial Pattern of Soil Carbon and Nitrogen in Forested and Non-Forested Riparian Zones in the Atlantic Coastal Plain of the Delaware River Basin. For. Ecol. Manage. 2013, 302, 200–209. [Google Scholar] [CrossRef]
- Song, B.-L.; Yan, M.-J.; Hou, H.; Guan, J.-H.; Shi, W.-Y.; Li, G.-Q.; Du, S. Distribution of Soil Carbon and Nitrogen in Two Typical Forests in the Semiarid Region of the Loess Plateau, China. Catena 2016, 143, 159–166. [Google Scholar] [CrossRef]
- Jin, Z.; Dong, Y.; Wang, Y.; Wei, X.; Wang, Y.; Cui, B.; Zhou, W. Natural Vegetation Restoration Is More Beneficial to Soil Surface Organic and Inorganic Carbon Sequestration than Tree Plantation on the Loess Plateau of China. Sci. Total. Environ. 2014, 485–486, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Liu, W.; Wei, J.; Cheng, J.; Li, W. Profile Distribution of Soil Inorganic Carbon along a Chronosequence of Grassland Restoration on a 22-Year Scale in the Chinese Loess Plateau. Catena 2014, 121, 321–329. [Google Scholar] [CrossRef]
- Wang, Y.; Xiong, W.; Gampe, S.; Coles, N.A.; Yu, P.; Xu, L.; Zuo, H.; Wang, Y. A Water Yield-Oriented Practical Approach for Multifunctional Forest Management and Its Application in Dryland Regions of China. J. Am. Water Resour Assoc. 2015, 51, 689–703. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sub-Areas | Forestation Modes | Group Quantity | Plot Quantity | Tree Density (Trees·ha−1) | Tree Height (m) | Tree Breast Dimension (cm) | Forest Age (years) | Main Tree Species |
|---|---|---|---|---|---|---|---|---|
| RM | GP | 1 | 9 | 1940 ± 449 | 8.3 ± 0.9 | 9.8 ± 0.4 | 27 ± 6 | Larix principis-rupprechtii |
| NP | 2 | 31 | 1116 ± 511 | 15.5 ± 2.8 | 19.7 ± 2.1 | 32 ± 3 | ||
| RH | CP | 1 | 9 | 3244 ± 1995 | 6.2 ± 3.5 | 6.9 ± 2.6 | 17 ± 8 | Robinia pseudoacacia |
| GP | 2 | 13 | 1271 ± 602 | 8.4 ± 5.2 | 15.0 ± 4.5 | 16 ± 8 | Pinus tabulaeformis, Armeniaca sibirica | |
| NP | 2 | 5 | 4140 ± 1927 | 7.3 ± 1.8 | 8.7 ± 1.6 | 21 ± 8 | Pinus tabulaeformis, Populus davidiana | |
| LH | CP | 4 | 29 | 1790 ± 965 | 4.8 ± 3.3 | 7.3 ± 3.9 | 14 ± 8 | Prunus davidiana, Pinus tabulaeformis, Pyrus sorotina |
| GP | 2 | 10 | 1789 ± 472 | 11.2 ± 2.9 | 14.6 ± 8.3 | 26 ± 9 | L. principis-rupprechtii, Hippophae rhamnoides | |
| NP | 1 | 9 | 3000 ± 1868 | 6.3 ± 5.7 | 7.6 ± 4.0 | 14 ± 7 | Robinia pseudoacacia |
| Sub-Areas | Forestation Modes | MAT (°C) | MAP (mm) | Alt (m) | Soil Bulk Density (g·cm−3) | Initial SOCD (t·ha−1) | Total N (t·ha−1) | Total P (t·ha−1) | Total K (t·ha−1) |
|---|---|---|---|---|---|---|---|---|---|
| RM | GP | 5.1 ± 0.5 | 564.0 ± 46.2 | 2069 ± 29 | 1.11 ± 0.06 | 257.33 | 28.9. ± 9.6 | 5.7 ± 1.2 | 220.2 ± 12.2 |
| NP | 5.1 ± 1.1 | 601.4 ± 20.0 | 2294 ± 158 | 1.17 ± 0.11 | 247.99 | 24.6 ± 6.2 | 7.7 ± 1.6 | 233.2 ± 28.5 | |
| RH | CP | 6.0 ± 0.7 | 562.5 ± 38.8 | 1806 ± 99 | 1.04 ± 0.07 | 73.70 | 11.8 ± 5.0 | 7.2 ± 1.4 | 213.4 ± 31.8 |
| GP | 6.9 ± 1.5 | 506.9 ± 52.3 | 1930 ± 116 | 1.12 ± 0.10 | 173.26 | 19.8 ± 7.0 | 7.1 ± 0.9 | 200.5 ± 35.0 | |
| NP | 7.2 ± 0.9 | 426.2 ± 19.1 | 1914 ± 247 | 1.09 ± 0.04 | 187.09 | 20.2 ± 11.0 | 8.1 ± 2.6 | 226.5 ± 18.8 | |
| LH | CP | 7.7 ± 0.8 | 484.8 ± 26.6 | 1836 ± 208 | 1.13 ± 0.08 | 45.66 | 7.9 ± 4.0 | 7.2 ± 1.2 | 200.1 ± 25.0 |
| GP | 7.5 ± 0.4 | 477.0 ± 10.1 | 1914 ± 247 | 1.02 ± 0.11 | 100.85 | 15.6 ± 7.1 | 7.4 ± 1.7 | 201.4 ± 27.5 | |
| NP | 7.8 ± 0.4 | 476.1 ± 14.4 | 2113 ± 155 | 1.09 ± 0.07 | 245.16 | 8.2 ± 4.2 | 7.1 ± 1.4 | 189.0 ± 14.3 |
| Foresta-tion Modes | Curve | Equation Number | Response Function Types | Fitted Response Equations | Inflection Points | Values at 40 Years (t·ha−1) | |
|---|---|---|---|---|---|---|---|
| Value (t·ha−1) | Time (Years) | ||||||
| NP | SOCDupper | (11) | Logistic | 389.67–181.41/[1 + (Time/24.40)4.40] | 389.7 | 40 | -- |
| SOCDbottom | (12) | Polynomial | 147.03–10.61 × Time + 0.31 × Time2 | 55.8 | 17 | -- | |
| SOCD average | (13) | Composite | ((11) + (12))/2 | 140.0 | 12 | 293.2 | |
| ΔSOCDupper | (14) | Logistic | 207.87–207.87/[1 + (Time/21.25)4.00] | 207.9 | 44 | -- | |
| ΔSOCDbottom | (15) | Polynomial | −21.91 × Time + 0.55 × Time2 | −216.6 | 20 | -- | |
| ΔSOCD average | (16) | Composite | ((14) + (15))/2 | −82.8 | 13 | 101.3 | |
| GP | SOCDupper | (17) | Logistic | 307.49–131.63/[1 + (Time/16.89)2.75] | 307.5 | 35 | -- |
| SOCDbottom | (18) | Polynomial | 121.90–2.63 × Time + 0.14 × Time2 | 109.3 | 10 | -- | |
| SOCD average | (19) | Composite | ((17) + (18))/2 | 145.9 | 4 | 266.5 | |
| ΔSOCDupper | (20) | Logistic | 182.36–182.36/[1 + (Time/15.51)3.17] | 182.4 | 39 | -- | |
| ΔSOCDbottom | (21) | Polynomial | −8.90 × Time + 0.30 × Time2 | −65.8 | 15 | -- | |
| ΔSOCD average | (22) | Composite | ((20) + (21))/2 | −17.1 | 6 | 149.6 | |
| CP | SOCDupper | (23) | Logistic | 201.17–127.47/[1 + (Time/0.17)2.78] | 201.2 | 25 | -- |
| SOCDbottom | (24) | Polynomial | 51.18–1.86 × Time + 0.08 × Time2 | 40.9 | 10 | -- | |
| SOCD average | (25) | Composite | ((23) + (24))/2 | -- | -- | 154.5 | |
| ΔSOCDupper | (26) | Logistic | 99.40–99.40/[1 + (Time/6.46)3.16] | 99.4 | 16 | -- | |
| ΔSOCDbottom | (27) | Polynomial | −1.34 × Time + 0.08 × Time2 | −5.4 | 8 | -- | |
| ΔSOCD average | (28) | Composite | ((26) + (27))/2 | -- | -- | 90.0 | |
| Forestation Modes | Possible Factors | Averages (Standard Error) | Max | Min | Correlation Coefficient | Regression | Explained Importance | |
|---|---|---|---|---|---|---|---|---|
| PRC | Pr | |||||||
| NP | Time | 27±9 | 40 | 3 | 0.34 * | 0.34 | 0.006 ** | 28% |
| Bio | 129.7±83.4 | 358.1 | 12.0 | 0.39 ** | 0.15 | 0.187 | 13% | |
| SOCD0 | 254.7±111.8 | 378.4 | 117.6 | −0.66 ** | −0.52 | 0.001 ** | 43% | |
| MAT | 5.9±1.5 | 8.7 | 4.2 | 0.19 | ||||
| MAP | 556.9±70.9 | 617.2 | 398.0 | 0.03 | ||||
| Alt | 2214±195 | 2525 | 1737 | −0.46 ** | −0.19 | 0.199 | 16% | |
| GP | Time | 23±9 | 36 | 4 | 0.47 ** | 0.42 | 0.002 ** | 42% |
| Bio | 85.3±64.4 | 218.1 | 1.9 | 0.33 | ||||
| SOCD0 | 167.4±68.9 | 257.3 | 69.0 | −0.63 ** | −0.59 | 0.000 ** | 58% | |
| MAT | 6.6±1.4 | 8.5 | 4.6 | 0.15 | ||||
| MAP | 513.6±53.2 | 603.2 | 450.4 | −0.16 | ||||
| Alt | 1876±209 | 2289 | 1506 | −0.18 | ||||
| CP | Time | 15±8 | 35 | 5 | 0.51 ** | 0.375 | 0.009 ** | 40% |
| Bio | 41.8±30.9 | 124.7 | 1.2 | 0.04 | ||||
| SOCD0 | 52.4±16.3 | 73.7 | 23.4 | 0.56 ** | 0.263 | 0.121 | 28% | |
| MAT | 7.3±1.1 | 9.5 | 4.6 | −0.35 * | −0.022 | 0.899 | 2% | |
| MAP | 503.2±44.5 | 603.2 | 434.6 | 0.51 ** | 0.288 | 0.114 | 30% | |
| Alt | 1829±187 | 2185 | 1446 | 0.11 | ||||
| Forestation Modes | Plot Quantity | Influencing Factors | Response Functions (Models) of △SOCD | R2 | Sig. (P) | Equation Number |
|---|---|---|---|---|---|---|
| NP | 45 | Time | 103.94–103.94/[1 + (Time/21.25)4.00]−10.96 × Time + 0.28 × Time2 | 0.14 | 0.01 | (16) |
| Time, SOCD0, Alt, Bio | {13.37–13.37 / [1 + (Time/58.52)11.49] −0.022 × Time + 0.0006 × Time2} × (4.58 × SOCD0 −0.53 × Alt−2.82 × Bio + 1029.41) | 0.56 | 0.000 | (29) | ||
| GP | 32 | Time | 91.18–91.18/[1 + (Time/15.51)3.17]−4.45 × Time + 0.15 × Time2 | 0.23 | 0.006 | (22) |
| Time, SOCD0 | {91.18- 91.18/ [1 + (Time/15.51)3.17] −4.45 × Time + 0.15 × Time2} × (−0.01 × SOCD0 + 1.92) | 0.54 | 0.000 | (30) | ||
| CP | 38 | Time | 49.70–49.70/[1 + (Time/6.46)3.16]−0.67 × Time + 0.04 × Time2 | 0.26 | 0.001 | (28) |
| Time, SOCD0, MAP, MAT | {3.40–3.40/[1 + (Time/18.26)0.02] + 0.71 × Time−0.01 × Time2} × (0.07 × SOCD0 + 0.02 × MAP + 0.29 × MAT −13.55) | 0.51 | 0.000 | (31) |
| SOC/SOCD Response Patterns | Range of Forest Age (Year) | Inflection Time (Year) | Initial Vegetation Type (SOCD0, t·ha−1) | Soil Layer (cm) | Country | Longitude(°) | Latitude(°) | Plantation Type after Afforestation | Reference |
|---|---|---|---|---|---|---|---|---|---|
| Decrease | 0–18 | Grassland | 0–100 | Australia | Pinus radiata | [13] | |||
| From decrease to increase | 0–30 | 15–30 | Grassland | 0–100 | China | 109.25 E | 36.77N | Robinia pseudoacacia | [11] |
| 0–30 | 10–15 | Grassland | 0–100 | China | 107.77–109.95 E | 35–38.80 N | Robinia pseudoacacia | [31] | |
| From slow increase to rapid increase | 0–35 | 10 | Cropland (7.7) | 0–20 | China | 109.24 E | 36.77 N | Robinia pseudoacacia | [32] |
| 0–30 | 10 | Forest | 0–10 | China | 116.30–116.48 E | 25.55–25.80 N | Pinus massoniana | [33] | |
| From increase to levelling off | 0–60 | 50 | Cropland | 0–100 | China | 108.58–109.25 E | 35.28–36.78 N | Coniferous | [14] |
| 0–40 | ≥16 years | Cropland (52.4) | 0–100 | China | 105.78–107.11 E | 35.32–36.58 N | Coniferous, deciduous broad-leaved | This study, Figure 3f | |
| From decrease to increase and finally levelling off | 0–250 | 5, 40 | Cropland (78.0) | 0–100 | China | Nationwide | Coniferous | [34] | |
| 10, 30 | Broad-leaved | ||||||||
| 5, 40 | Evergreen | ||||||||
| 10, 30 | Deciduous | ||||||||
| 0–40 | ≥39 years | Grassland (167.4) | 0–100 | China | 105.78–107.11 E | 35.32–36.58 N | Coniferous, deciduous broad-leaved | This study, Figure 3e | |
| 0–40 | ≥44 years | Forest (254.7) | 0–100 | China | 105.78–107.11 E | 35.32–36.58 N | Coniferous, deciduous broad-leaved | This study, Figure 3d | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Guo, J.; Wang, Y.; Yu, P.; Wang, X. Variation of Soil Organic Carbon Density with Plantation Age and Initial Vegetation Types in the Liupan Mountains Areas of Northwest China. Forests 2021, 12, 1811. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121811
Zhang Z, Guo J, Wang Y, Yu P, Wang X. Variation of Soil Organic Carbon Density with Plantation Age and Initial Vegetation Types in the Liupan Mountains Areas of Northwest China. Forests. 2021; 12(12):1811. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121811
Chicago/Turabian StyleZhang, Ziyou, Jianbin Guo, Yanhui Wang, Pengtao Yu, and Xiao Wang. 2021. "Variation of Soil Organic Carbon Density with Plantation Age and Initial Vegetation Types in the Liupan Mountains Areas of Northwest China" Forests 12, no. 12: 1811. https://0-doi-org.brum.beds.ac.uk/10.3390/f12121811