
Diversity of Phytophthora Species Detected in Disturbed and Undisturbed British Soils Using High-Throughput Sequencing Targeting ITS rRNA and COI mtDNA Regions

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
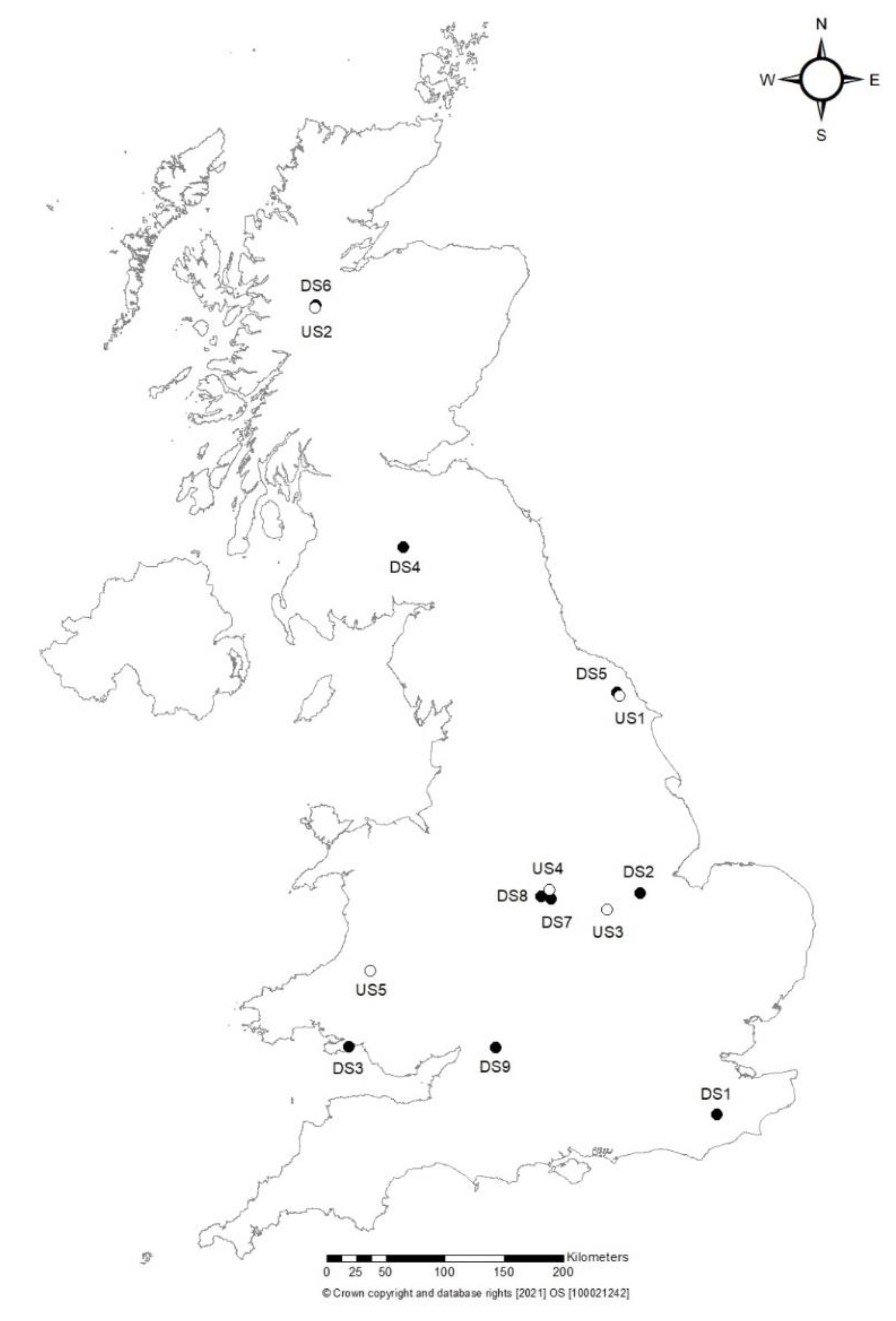
2.1. Soil Sampling and Phytophthora Isolation
2.2. Soil DNA Extraction and PCR Amplification
2.3. Positive Controls for Illumina Sequencing
2.4. Illumina Sequencing Library Preparation and Sequencing
2.5. Bioinformatic and Statistical Analyses
3. Results
3.1. Sequencing Output and Performance of Control Reactions
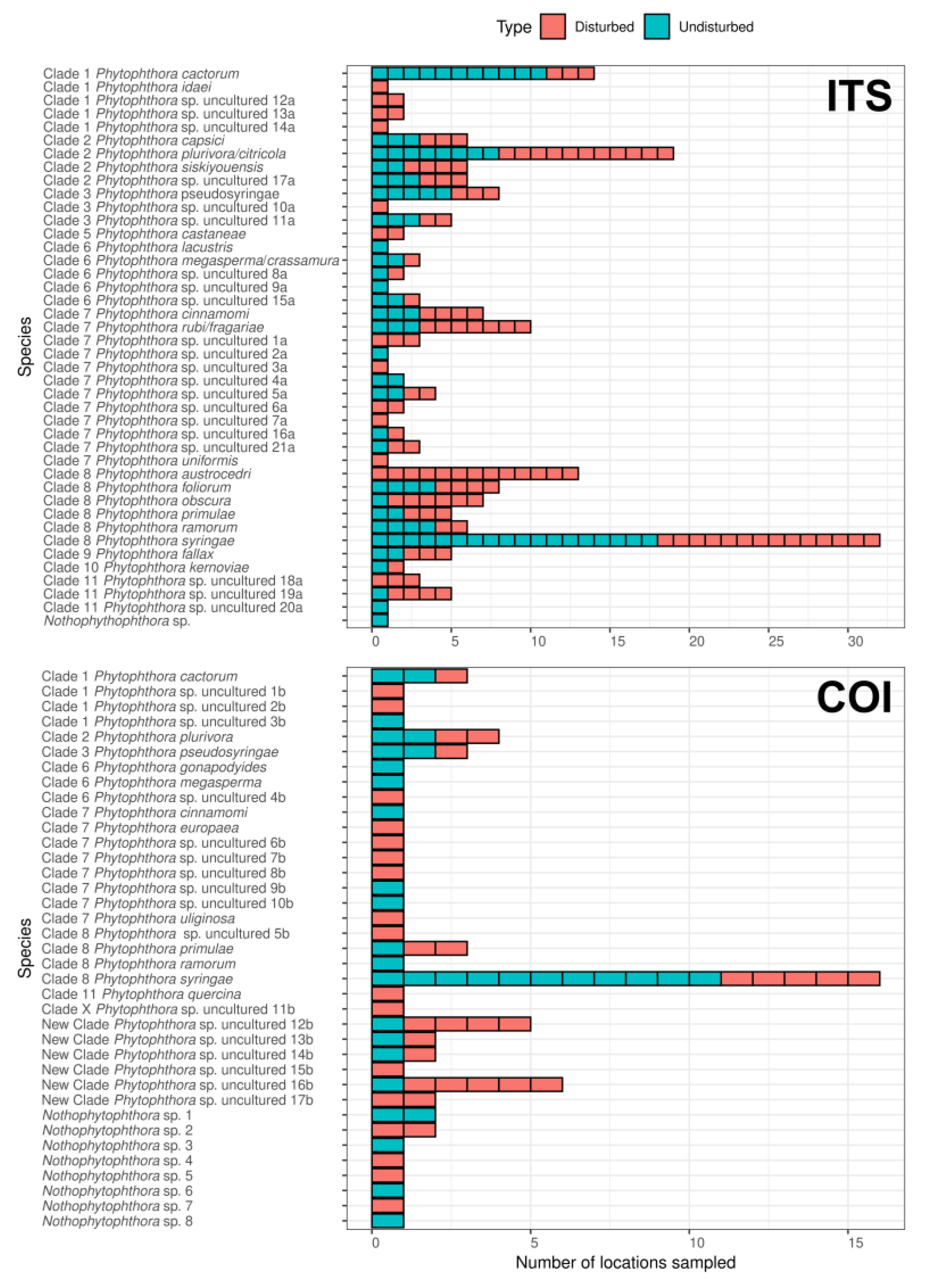
3.2. Analysis of Sequences from British Soil Samples
3.3. Identification of Phytophthora Phylotypes in Britain by NGS
3.4. Identification of Phytophthora by Isolation
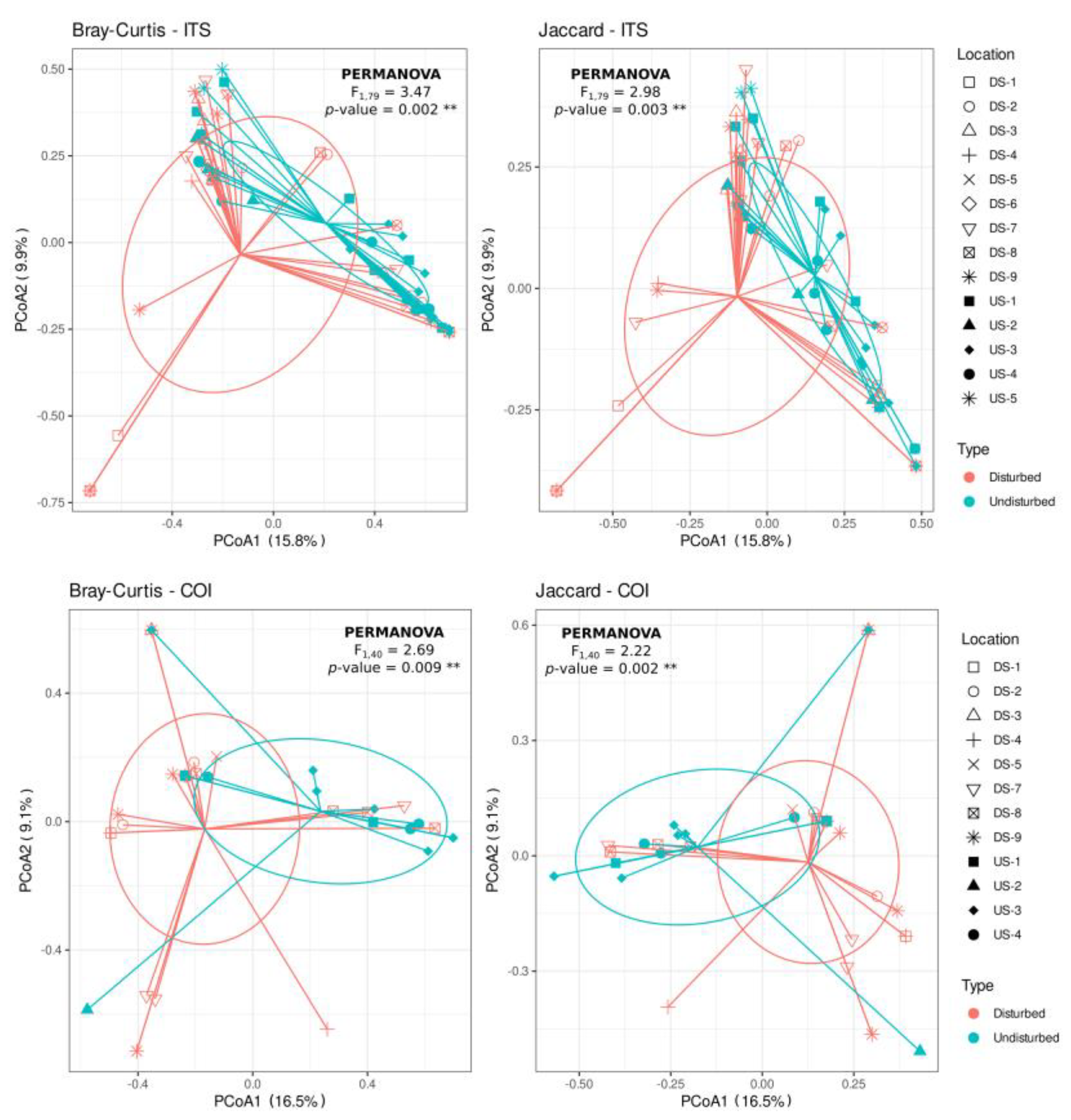
3.5. Phytophthora spp. Community Composition Differs between Disturbed and Undisturbed Sites
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brasier, C.M.; Beales, P.A.; Kirk, S.A.; Denman, S.; Rose, J. Phytophthora kernoviae sp. nov., an invasive pathogen causing bleeding stem lesions on forest trees and foliar necrosis of ornamentals in the UK. Mycol. Res. 2005, 109, 853–859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasier, C.; Webber, J. Sudden larch death. Nat. Cell Biol. 2010, 466, 824–825. [Google Scholar] [CrossRef] [PubMed]
- Robin, C.; Piou, D.; Feau, N.; Douzon, G.; Schenck, N.; Hansen, E.M. Root and aerial infections of Chamaecyparis lawsoniana by Phytophthora lateralis: A new threat for European countries. For. Pathol. 2010, 41, 417–424. [Google Scholar] [CrossRef]
- Green, S.; Brasier, C.M.; Schlenzig, A.; McCracken, A.; Macaskill, G.A.; Wilson, M.; Webber, J.F. The destructive invasive pathogen Phytophthora lateralis found on Chamaecyparis lawsoniana across the UK. For. Pathol. 2012, 43, 19–28. [Google Scholar] [CrossRef]
- Hansen, E.M. Phytophthora Species Emerging as Pathogens of Forest Trees. Curr. For. Rep. 2015, 1, 16–24. [Google Scholar] [CrossRef] [Green Version]
- Scanu, B.; Webber, J.F. Dieback and mortality of Nothofagus in Britain: Ecology, pathogenicity and sporulation potential of the causal agent Phytophthora pseudosyringae. Plant Pathol. 2016, 65, 26–36. [Google Scholar] [CrossRef]
- Greslebin, A.G.; Vélez, M.L.; Green, S. Phytophthora Diseases: Mal del Ciprés; American Phytopathological Society: St. Paul, MN, USA, 2018. [Google Scholar]
- Jung, T.; Orlikowski, L.; Henricot, B.; Abad-Campos, P.; Aday, A.G.; Casal, O.A.; Bakonyi, J.; Cacciola, S.O.; Cech, T.; Chavarriaga, D.; et al. Widespread Phytophthora infestations in European nurseries put forest, semi-natural and horticultural ecosystems at high risk of Phytophthora diseases. For. Pathol. 2015, 46, 134–163. [Google Scholar] [CrossRef] [Green Version]
- Redekar, N.R.; Eberhart, J.L.; Parke, J.L. Diversity of Phytophthora, Pythium, and Phytopythium Species in Recycled Irrigation Water in a Container Nursery. Phytobiomes J. 2019, 3, 31–45. [Google Scholar] [CrossRef] [Green Version]
- Rooney-Latham, S.; Blomquist, C.L.; Kosta, K.L.; Gou, Y.Y.; Woods, P.W. Phytophthora Species are Common on Nursery Stock Grown for Restoration and Revegetation Purposes in California. Plant Dis. 2019, 103, 448–455. [Google Scholar] [CrossRef] [Green Version]
- Molnar, C.; Nikolaeva, E.; Kim, S.; Olson, T.; Bily, D.; Kim, J.-E.; Kang, S. Phytophthora Diversity in Pennsylvania Nurseries and Greenhouses Inferred from Clinical Samples Collected over Four Decades. Microorganism 2020, 8, 1056. [Google Scholar] [CrossRef]
- Scott, P.; Bader, M.K.-F.; Burgess, T.; Hardy, G.; Williams, N. Global biogeography and invasion risk of the plant pathogen genus Phytophthora. Environ. Sci. Policy 2019, 101, 175–182. [Google Scholar] [CrossRef]
- Hulbert, J.M.; Agne, M.C.; Burgess, T.I.; Roets, F.; Wingfield, M.J. Urban environments provide opportunities for early detections of Phytophthora invasions. Biol. Invasions 2017, 19, 3629–3644. [Google Scholar] [CrossRef] [Green Version]
- Paap, T.; Burgess, T.I.; Wingfield, M.J. Urban trees: Bridge-heads for forest pest invasions and sentinels for early detection. Biol. Invasions 2017, 19, 3515–3526. [Google Scholar] [CrossRef] [Green Version]
- Hüberli, D.; Hardy, G.E.S.J.; White, D.; Williams, N.; Burgess, T.I. Fishing for Phytophthora from Western Australia’s waterways: A distribution and diversity survey. Australas. Plant Pathol. 2013, 42, 251–260. [Google Scholar] [CrossRef]
- Jung, T.; Jung, M.H.; Cacciola, S.O.; Cech, T.; Bakonyi, J.; Seress, D.; Mosca, S.; Schena, L.; Seddaiu, S.; Pane, A.; et al. Multiple new cryptic pathogenic Phytophthora species from Fagaceae forests in Austria, Italy and Portugal. IMA Fungus 2017, 8, 219–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; Jung, M.H.; Scanu, B.; Seress, D.; Kovács, M.G.; Maia, C.; Pérez-Sierra, A.; Chang, T.-T.; Chandelier, A.; Heungens, K.; et al. Six new Phytophthora species from ITS Clade 7a including two sexually functional heterothallic hybrid species detected in natural ecosystems in Taiwan. Pers. Mol. Phylogeny Evol. Fungi 2017, 38, 100–135. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Pérez-Sierra, A.; Durán, A.; Jung, M.H.; Balci, Y.; Scanu, B. Canker and decline diseases caused by soil- and airborne Phytophthora species in forests and woodlands. Pers. Mol. Phylogeny Evol. Fungi 2018, 40, 182–220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jung, T.; La Spada, F.; Pane, A.; Aloi, F.; Evoli, M.; Jung, M.H.; Scanu, B.; Faedda, R.; Rizza, C.; Puglisi, I.; et al. Diversity and Distribution of Phytophthora Species in Protected Natural Areas in Sicily. Forests 2019, 10, 259. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Scanu, B.; Brasier, C.M.; Webber, J.; Milenković, I.; Corcobado, T.; Tomšovský, M.; Pánek, M.; Bakonyi, J.; Maia, C.; et al. A Survey in Natural Forest Ecosystems of Vietnam Reveals High Diversity of both New and Described Phytophthora Taxa including P. ramorum. Forests 2020, 11, 93. [Google Scholar] [CrossRef] [Green Version]
- Burgess, T.I.; White, D.; McDougall, K.M.; Garnas, J.; Dunstan, W.A.; Català, S.; Carnegie, A.J.; Worboys, S.; Cahill, D.; Vettraino, A.-M.; et al. Distribution and diversity of Phytophthora across Australia. Pac. Conserv. Biol. 2017, 23, 150–162. [Google Scholar] [CrossRef] [Green Version]
- Bourret, B.T.; Aram, K.; Mehl, K.H.; Rizzo, M.D.; Rooney-Latham, S.; Swiecki, J.T.; Frankel, J.S. Ten new provisional species of Phytophthora and Nothophytophthora from California. In Proceedings of the Seventh Sudden Oak Death Science and Management Symposium: Healthy Plants in a World with Phytophthora, San Francisco, CA, USA, 25–27 June 2019; General Technical Report PSW-GTR-268. US Forest Service: Albany, CA, USA, 2020; Volume 268, pp. 46–47. [Google Scholar]
- Brasier, C.M. The biosecurity threat to the UK and global environment from international trade in plants. Plant Pathol. 2008, 57, 792–808. [Google Scholar] [CrossRef]
- Moralejo, E.; Pérez-Sierra, A.M.; Álvarez, L.A.; Belbahri, L.; Lefort, F.; Descals, E. Multiple alien Phytophthora taxa discovered on diseased ornamental plants in Spain. Plant Pathol. 2009, 58, 100–110. [Google Scholar] [CrossRef]
- O’Hanlon, R.; Choiseul, J.; Corrigan, M.; Catarame, T.; Destefanis, M. Diversity and detections of Phytophthora species from trade and non-trade environments in Ireland. EPPO Bull. 2016, 46, 594–602. [Google Scholar] [CrossRef]
- Frankel, S.J.; Conforti, C.; Hillman, J.; Ingolia, M.; Shor, A.; Benner, D.; Alexander, J.M.; Bernhardt, E.; Swiecki, T.J. Phytophthora Introductions in Restoration Areas: Responding to Protect California Native Flora from Human-Assisted Pathogen Spread. Forests 2020, 11, 1291. [Google Scholar] [CrossRef]
- Giordana, G.; Kitzberger, T.; La Manna, L. Anthropogenic Factors Control the Distribution of a Southern Conifer Phytophthora Disease in a Peri-Urban Area of Northern Patagonia, Argentina. Forests 2020, 11, 1183. [Google Scholar] [CrossRef]
- Cooke, D.; Drenth, A.; Duncan, J.; Wagels, G.; Brasier, C. A Molecular Phylogeny of Phytophthora and Related Oomycetes. Fungal Genet. Biol. 2000, 30, 17–32. [Google Scholar] [CrossRef]
- Seifert, K.A. Progress towards DNA barcoding of fungi. Mol. Ecol. Resour. 2009, 9, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Grünwald, N.J.; Martin, F.N.; Larsen, M.M.; Sullivan, C.M.; Press, C.M.; Coffey, M.D.; Hansen, E.M.; Parke, J.L. Phytophthora-ID.org: A Sequence-Based Phytophthora Identification Tool. Plant Dis. 2011, 95, 337–342. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Hong, C. Differential Usefulness of Nine Commonly Used Genetic Markers for Identifying Phytophthora Species. Front. Microbiol. 2018, 9, 2334. [Google Scholar] [CrossRef] [Green Version]
- Català, S.; Pérez-Sierra, A.; Abad-Campos, P. The Use of Genus-Specific Amplicon Pyrosequencing to Assess Phytophthora Species Diversity Using eDNA from Soil and Water in Northern Spain. PLoS ONE 2015, 10, e0119311. [Google Scholar] [CrossRef] [Green Version]
- Vannini, A.; Bruni, N.; Tomassini, A.; Franceschini, S.; Vettraino, A.M. Pyrosequencing of environmental soil samples reveals biodiversity of the Phytophthora resident community in chestnut forests. FEMS Microbiol. Ecol. 2013, 85, 433–442. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Prigigallo, M.I.; Abdelfattah, A.; Cacciola, S.O.; Faedda, R.; Sanzani, S.M.; Cooke, D.E.L.; Schena, L. Metabarcoding Analysis of Phytophthora Diversity Using Genus-Specific Primers and 454 Pyrosequencing. Phytopathology 2016, 106, 305–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riddell, C.E.; Frederickson-Matika, D.; Armstrong, A.C.; Elliot, M.; Forster, J.; Hedley, P.E.; Morris, J.; Thorpe, P.; El Cooke, D.; Pritchard, L.; et al. Metabarcoding reveals a high diversity of woody host-associated Phytophthora spp. in soils at public gardens and amenity woodlands in Britain. PeerJ 2019, 7, e6931. [Google Scholar] [CrossRef] [Green Version]
- Kroon, L.; Bakker, F.; Bosch, G.V.D.; Bonants, P.; Flier, W. Phylogenetic analysis of Phytophthora species based on mitochondrial and nuclear DNA sequences. Fungal Genet. Biol. 2004, 41, 766–782. [Google Scholar] [CrossRef] [PubMed]
- Blair, J.E.; Coffey, M.D.; Park, S.-Y.; Geiser, D.M.; Kang, S. A multi-locus phylogeny for Phytophthora utilizing markers derived from complete genome sequences. Fungal Genet. Biol. 2008, 45, 266–277. [Google Scholar] [CrossRef]
- Martin, F.N.; Blair, J.E.; Coffey, M.D. A combined mitochondrial and nuclear multilocus phylogeny of the genus Phytophthora. Fungal Genet. Biol. 2014, 66, 19–32. [Google Scholar] [CrossRef] [PubMed]
- Robideau, G.P.; De Cock, A.W.A.M.; Coffey, M.D.; Voglmayr, H.; Brouwer, H.; Bala, K.; Chitty, D.W.; Désaulniers, N.; Eggertson, Q.A.; Gachon, C.M.M.; et al. DNA barcoding of oomycetes with cytochrome c oxidase subunit I and internal transcribed spacer. Mol. Ecol. Resour. 2011, 11, 1002–1011. [Google Scholar] [CrossRef] [PubMed]
- Choi, Y.-J.; Beakes, G.; Glockling, S.; Kruse, J.; Nam, B.; Nigrelli, L.; Ploch, S.; Shin, H.-D.; Shivas, R.G.; Telle, S.; et al. Towards a universal barcode of oomycetes—A comparison of the cox1 and cox2 loci. Mol. Ecol. Resour. 2015, 15, 1275–1288. [Google Scholar] [CrossRef] [Green Version]
- Brasier, C.M.; Kirk, S.A. Comparative aggressiveness of standard and variant hybrid alder phytophthoras, Phytophthora cambivora and other Phytophthora species on bark of Alnus, Quercus and other woody hosts. Plant Pathol. 2001, 50, 218–229. [Google Scholar] [CrossRef]
- Man, W.A.; de Cock, A.W.; Summerbell, R.C. Natural hybrids of resident and introduced Phytophthora species proliferating on multiple new hosts. Eur. J. Plant Pathol. 2006, 117, 25–33. [Google Scholar] [CrossRef]
- Nagel, J.H.; Gryzenhout, M.; Slippers, B.; Wingfield, M.J.; Hardy, G.E.S.J.; Stukely, M.J.; Burgess, T.I. Characterization of Phytophthora hybrids from ITS clade 6 associated with riparian ecosystems in South Africa and Australia. Fungal Biol. 2013, 117, 329–347. [Google Scholar] [CrossRef] [Green Version]
- Jung, T.; Blaschke, H.; Neumann, P. Isolation, identification and pathogenicity of Phytophthora species from declining oak stands. For. Pathol. 1996, 26, 253–272. [Google Scholar] [CrossRef]
- Jung, T.; Blaschke, H.; Osswald, W. Involvement of soilborne Phytophthora species in Central European oak decline and the effect of site factors on the disease. Plant Pathol. 2000, 49, 706–718. [Google Scholar] [CrossRef]
- Scibetta, S.; Schena, L.; Chimento, A.; Cacciola, S.O.; Cooke, D.E. A molecular method to assess Phytophthora diversity in environmental samples. J. Microbiol. Methods 2012, 88, 356–368. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M., Gelfand, D., Shinsky, J., White, T., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Illumina. 16S Metagenomic Sequencing Library Preparation; Illumina: San Diego, CA, USA, 2013; p. 28. [Google Scholar]
- Krueger, F. Trim Galore!: A Wrapper Tool Around Cutadapt and FastQC to Consistently Apply Quality and Adapter Trimming to FastQ Files; Babraham Institute: Babraham, UK, 2015. [Google Scholar]
- Babraham Bioinformatics. FastQC v. 0.11.2. Available online: http://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 10 January 2020).
- Ewels, P.; Magnusson, M.; Lundin, S.; Käller, M. MultiQC: Summarize analysis results for multiple tools and samples in a single report. Bioinformatics 2016, 32, 3047–3048. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; Mcmurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High-resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed]
- Ratnasingham, S.; Hebert, P.D.N. BARCODING: Bold: The Barcode of Life Data System. Mol. Ecol. Notes 2007, 7, 355–364. Available online: http://www.barcodinglife.org (accessed on 14 February 2021). [CrossRef] [PubMed] [Green Version]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in Performance and Usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Capella-Gutiérrez, S.; Silla-Martínez, J.M.; Gabaldón, T. trimAl: A tool for automated alignment trimming in large-scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef]
- Darriba, D.; Posada, D.; Kozlov, A.M.; Stamatakis, A.; Morel, B.; Flouri, T. ModelTest-NG: A New and Scalable Tool for the Selection of DNA and Protein Evolutionary Models. Mol. Biol. Evol. 2020, 37, 291–294. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, L.-T.; Schmidt, H.A.; Von Haeseler, A.; Minh, B.Q. IQ-TREE: A Fast and Effective Stochastic Algorithm for Estimating Maximum-Likelihood Phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef] [PubMed]
- Hoang, D.T.; Chernomor, O.; Von Haeseler, A.; Minh, B.Q.; Vinh, L.S. UFBoot2: Improving the Ultrafast Bootstrap Approximation. Mol. Biol. Evol. 2018, 35, 518–522. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.J. A new method for non-parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46. [Google Scholar] [CrossRef]
- Legendre, P.; Gallagher, E.D. Ecologically meaningful transformations for ordination of species data. Oecologia 2001, 129, 271–280. [Google Scholar] [CrossRef]
- Anderson, M.J. Distance-Based Tests for Homogeneity of Multivariate Dispersions. Biometrics 2005, 62, 245–253. [Google Scholar] [CrossRef]
- Oksanen, J.; Blanchet, F.G.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, R.B.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-2. 2018. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 14 February 2021).
- Cooke, D.E.L. Threats Posed by Phytophthora to Scottish Plant Health; A Review of Previous Findings, Pathways of Entry and Further Spread and the Status of Diagnostic Techniques. RESAS Phytophthora Risk Review. Scottish Government Report. Available online: https://cran.r-project.org/web/packages/vegan/index.html (accessed on 28 November 2020).
- Farr, D.F.; Rossman, A.Y. Fungal Databases, US National Fungus Collections, ARS, USDA. Available online: https://nt.ars-grin.gov/fungaldatabases/ (accessed on 14 February 2021).
- Roesch, L.F.W.; Fulthorpe, R.R.; Riva, A.; Casella, G.; Hadwin, A.K.M.; Kent, A.D.; Daroub, S.H.; Camargo, F.A.O.; Farmerie, W.G.; Triplett, E.W. Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J. 2007, 1, 283–290. [Google Scholar] [CrossRef] [PubMed]
- Jumpponen, A.; Jones, K.L. Massively parallel 454 sequencing indicates hyperdiverse fungal communities in temperate Quercus macrocarpa phyllosphere. New Phytol. 2009, 184, 438–448. [Google Scholar] [CrossRef] [PubMed]
- Vettraino, A.; Bonants, P.; Tomassini, A.; Bruni, N.; Vannini, A. Pyrosequencing as a tool for the detection of Phytophthora species: Error rate and risk of false Molecular Operational Taxonomic Units. Lett. Appl. Microbiol. 2012, 55, 390–396. [Google Scholar] [CrossRef]
- Jung, T.; Scanu, B.; Bakonyi, J.; Seress, D.; Kovács, G.; Durán, A.; Von Stowasser, E.S.; Schena, L.; Mosca, S.; Thu, P.; et al. Nothophytophthora gen. nov., a new sister genus of Phytophthora from natural and semi-natural ecosystems. Pers. Mol. Phylogeny Evol. Fungi 2017, 39, 143–174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlenzig, A.; Purser, E.; Perez-Sierra, A. First finding of Phytophthora foliorum in the United Kingdom. New Dis. Rep. 2016, 34, 2. [Google Scholar] [CrossRef] [Green Version]
- Donahoo, R.; Blomquist, C.L.; Thomas, S.L.; Moulton, J.K.; Cooke, D.E.; Lamour, K.H. Phytophthora foliorum sp. nov., a new species causing leaf blight of azalea. Mycol. Res. 2006, 110, 1309–1322. [Google Scholar] [CrossRef]
- Green, S.; Hendry, S.; Macaskill, G.; Laue, B.; Steele, H. Dieback and mortality of Juniperus communis in Britain associated with Phytophthora austrocedrae. New Dis. Rep. 2012, 26, 2. [Google Scholar] [CrossRef] [Green Version]
- Green, S.; Elliot, M.; Armstrong, A.; Hendry, S.J. Phytophthora austrocedrae emerges as a serious threat to juniper (Juniperus communis) in Britain. Plant Pathol. 2014, 64, 456–466. [Google Scholar] [CrossRef]
- Henricot, B.; Pérez-Sierra, A.; Armstrong, A.C.; Sharp, P.M.; Green, S. Morphological and Genetic Analyses of the Invasive Forest Pathogen Phytophthora austrocedri Reveal that Two Clonal Lineages Colonized Britain and Argentina from a Common Ancestral Population. Phytopathology 2017, 107, 1532–1540. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greslebin, A.G.; Hansen, E.M.; Sutton, W. Phytophthora austrocedrae sp. nov., a new species associated with Austrocedrus chilensis mortality in Patagonia (Argentina). Mycol. Res. 2007, 111, 308–316. [Google Scholar] [CrossRef] [PubMed]
- Werres, S.; Marwitz, R.; In’T Veld, W.A.M.; De Cock, A.W.A.M.; Bonants, P.J.M.; De Weerdt, M.; Themann, K.; Ilieva, E.; Baayen, R.P. Phytophthora ramorum sp. nov., a new pathogen on Rhododendron and Viburnum. Mycol. Res. 2001, 105, 1155–1165. [Google Scholar] [CrossRef]
- Lane, C.R.; Beales, P.A.; Hughes, K.J.D.; Griffin, R.L.; Munro, D.; Brasier, C.M.; Webber, J.F. First outbreak of Phytophthora ramorum in England, on Viburnum tinus. Plant Pathol. 2003, 52, 414. [Google Scholar] [CrossRef]
- Brasier, C.M.; Denman, S.; Rose, J.; Kirk, S.A.; Hughes, K.J.D.; Griffin, R.L.; Lane, C.R.; Inman, A.J.; Webber, J.F. First report of ramorum bleeding canker on Quercus falcata, caused by Phytophthora ramorum. Plant Pathol. 2004, 53, 804. [Google Scholar] [CrossRef]
- Denman, S.; Kirk, S.A.; Brasier, C.M.; Hughes, K.J.D.; Griffin, R.; Hobdon, E.; Webber, J.F. Foliar infection of sweet chestnut (Castanea sativa) by Phytophthora ramorum in the UK. Plant Pathol. 2005, 54, 581. [Google Scholar] [CrossRef]
- Giltrap, P.M.; Inman, A.J.; Barton, V.C.; Barnes, A.V.; Lane, C.R.; Hughes, K.J.D.; Tomlinson, J.; Dean, M.L.; Izzard, K. First report of ramorum dieback (Phytophthora ramorum) on Hamamelis virginiana in the UK. Plant Pathol. 2004, 53, 526. [Google Scholar] [CrossRef]
- Hughes, K.J.D.; Giltrap, P.M.; Barton, V.C.; Hobden, E.; Tomlinson, J.A.; Barber, P. On-site real-time PCR detection of Phytophthora ramorum causing dieback of Parrotia persica in the UK. Plant Pathol. 2006, 55, 813. [Google Scholar] [CrossRef]
- Inman, A.J.; Townend, V.C.; Barnes, A.V.; Lane, C.R.; Hughes, K.J.D.; Griffin, R.L.; Eales, S.J. First report of ramorum dieback (Phytophthora ramorum) on Pieris in England. Plant Pathol. 2003, 52, 785. [Google Scholar] [CrossRef]
- Webber, J.; Mullett, M.; Brasier, C. Dieback and mortality of plantation Japanese larch (Larix kaempferi) associated with infection by Phytophthora ramorum. New Dis. Rep. 2010, 22, 19. [Google Scholar] [CrossRef] [Green Version]
- Grünwald, N.; Werres, S.; Goss, E.M.; Taylor, C.R.; Fieland, V.J. Phytophthora obscura sp. nov., a new species of the novel Phytophthora subclade 8d. Plant Pathol. 2011, 61, 610–622. [Google Scholar] [CrossRef]
- Català, S.; Berbegal, M.; Pérez-Sierra, A.; Abad-Campos, P. Metabarcoding and development of new real-time specific assays reveal Phytophthora species diversity in holm oak forests in eastern Spain. Plant Pathol. 2016, 66, 115–123. [Google Scholar] [CrossRef]
- Reeser, P.W.; Sutton, W.; Hansen, E.M.; Remigi, P.; Adams, G.C. Phytophthora species in forest streams in Oregon and Alaska. Mycologia 2011, 103, 22–35. [Google Scholar] [CrossRef]
- Brasier, C.M.; Rose, J.; Gibbs, J.N. An unusual Phytophthora associated with widespread alder mortality in Britain. Plant Pathol. 1995, 44, 999–1007. [Google Scholar] [CrossRef]
- Fraser, S.; Gomez-Gallego, M.; Gardner, J.; Bulman, L.S.; Denman, S.; Williams, N.M. Impact of weather variables and season on sporulation of Phytophthora pluvialis and Phytophthora kernoviae. For. Pathol. 2020, 50, e12588. [Google Scholar] [CrossRef]
- Dick, M.A.; Dobbie, K.; Cooke, D.E.; Brasier, C.M. Phytophthora captiosa sp. nov. and P. fallax sp. nov. causing crown dieback of Eucalyptus in New Zealand. Mycol. Res. 2006, 110, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Wheller, T.; Erwin, D.C.; Ribeiro, O.K. Phytophthora Diseases Worldwide. Mycologia 1998, 90, 1092. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Site Code (Site Type) | Location | Surveyed Location and Nearest Hosts | Signs of Ill-Health Tree Status | ITS | COI | Isolation |
|---|---|---|---|---|---|---|
| DS1 (Arboreta) | DS1-1 | Mixed Pinus species and Abies | Healthy | P. austrocedri | Clade 8 Phytophthora sp. uncultured 5b | P. cryptogea |
| DS1-2 | Mixed new planting Asian species (broadleaved trees and conifers) | Healthy | P. austrocedri | - | - | |
| DS1-3 | Heathland | Healthy | P. austrocedri | - | - | |
| DS1-4 | Pinus sp. | Declining | P. austrocedri | - | P. cinnamomi | |
| DS1-5 | Acid grassland, mainly conifers | Healthy | P. austrocedri | - | P. cinnamomi | |
| DS1-6 | Cedrus sp. | Healthy | P. austrocedri | - | P. cinnamomi | |
| DS1-7 | New planting Betula, Nothofagus, Quercus | Healthy | P. austrocedri, P. pseudosyringae | - | - | |
| DS1-8 | Mixed broadleaved trees and conifers new plantings | Healthy | P. austrocedri | New Clade Phytophthora sp. uncultured 12b | P. megasperma | |
| DS1-9 | Mixed broadleaved trees and conifers new plantings | Healthy | - | - | - | |
| DS1-10 | Acid grassland, conifer and broadleaved trees, new plantings | Healthy | - | - | P. cinnamomi | |
| DS2 (Woodland/Recreational Park) | DS2-1 | Fraxinus | Declining | - | - | - |
| DS2-2 | Quercus | Dead trees | Clade 7 Phytophthora sp. uncultured 3a and 21a | P. europaea | - | |
| DS2-3 | Fraxinus | Dead trees | P. cactorum, P. syringae, P. cinnamomi | P. cactorum | - | |
| DS2-4 | Quercus | Declining | P. cactorum, P. syringae, Clade 7 Phytophthora sp. uncultured 21a | P. uliginosa, Clade 1 Phytophthora sp. uncultured 2b, New Clade Phytophthora sp. uncultured 12b | - | |
| DS2-5 | Larix | Dead | - | - | - | |
| DS2-6 | Betula | Dead | - | - | - | |
| DS2-7 | Quercus | Declining | - | - | - | |
| DS2-8 | Acer campestre | Declining | P. cactorum, P. megasperma/crassamura, P. ramorum, P. rubi/fragariae, Clade 7 Phytophthora sp. uncultured 5a and 6a | Clade X Phytophthora sp. uncultured 11b | - | |
| DS2-9 | Acer and Fraxinus | Declining | P. plurivora/citricola, P. syringae | - | - | |
| DS2-10 | Larix in area with Corylus, Fraxinus and Quercus | Healthy | P. capsici, P. obscura, P. plurivora/citricola, P. rubi, P. siskiyouensis, P. syringae, Clade 2 Phytophthora sp. uncultured 17a | - | P. plurivora | |
| DS3 (Garden) | DS3-1 | Border with herbaceous plants and shrubs, mixed planting | Healthy | Clade 3 Phytophthora sp. uncultured 10a | - | P. plurivora |
| DS3-2 | Kalmia, Quercus and Rhododendron | Healthy | - | - | ||
| DS3-3 | Quercus, Rhododendron and Sorbus | Healthy | Clade 11 Phytophthora sp. uncultured 19a | - | ||
| DS3-4 | Area with silt from a stream with no trees in the last 8 years surrounded by Acer, Larix and Quercus | Healthy | P. obscura, Clade 7 Phytophthora sp. uncultured 1a | - | P. cinnamomi | |
| DS3-5 | Border, the old kitchen garden, herbaceous plants and shrubs | Healthy | P. primulae, P. syringae | P. primulae | P. chlamydospora | |
| DS3-6 | Burning site for the garden | NA | P. obscura, P. rubi/fragariae | - | P. plurivora | |
| DS3-7 | Quercus and Rhododendron | Healthy | P. rubi/fragariae | - | - | |
| DS3-8 | Composting area of the garden | NA | Clade 1 Phytophthora sp. uncultured 14a, Clade 11 Phytophthora sp. uncultured 18a and 19a | - | P. plurivora | |
| DS3-9 | Mature Fagus, Fraxinus and Quercus | Healthy | - | - | P. cinnamomi | |
| DS3-10 | Mature Quercus and Rhododendron | Healthy | - | - | - | |
| DS4 (New plantings and a link to nurseries) | DS4-1 | Young Sorbus | Dead | - | - | - |
| DS4-2 | Young Quercus | Declining | P. obscura | - | - | |
| DS4-3 | Young Salix | Declining | P. austrocedri, P. plurivora/citricola, P. siskiyouensis | - | - | |
| DS4-4 | Young Juniperus | Declining | - | - | - | |
| DS4-5 | Young Salix | Declining | - | P. syringae, New Clade Phytophthora sp. uncultured 16b | - | |
| DS4-6 | Young Sorbus | Declining | P. ramorum | - | - | |
| DS4-7 | Young Betula | Declining | - | - | - | |
| DS4-8 | Young Prunus | Declining | - | - | - | |
| DS4-9 | Young Crataegus | Declining | - | New Clade Phytophthora sp. uncultured 12b | - | |
| DS4-10 | Young Corylus | Healthy | - | - | - | |
| DS5 (Woodland/Recreational Park) | DS5-1 | Betula | Declining | - | - | - |
| DS5-2 | Betula | Declining | - | - | - | |
| DS5-3 | Quercus | Declining | P. siskiyouensis | - | - | |
| DS5-4 | Sorbus | Declining | P. plurivora/citricola, P. syringae | P. plurivora | P. plurivora | |
| DS5-5 | Abies | Dead | - | - | - | |
| DS5-6 | Quercus robur | Declining | P. plurivora/citricola | - | - | |
| DS5-7 | Pinus sylvestris | Dead | - | - | - | |
| DS5-8 | Pinus sylvestris | Dead | - | - | - | |
| DS5-9 | Fagus | Declining | - | - | - | |
| DS5-10 | Fagus | Healthy | - | - | - | |
| DS6 (New plantings and a link to nurseries) | DS6-1 | Sorbus aucuparia | Healthy | - | - | - |
| DS6-2 | Betula | Healthy | - | - | - | |
| DS6-3 | Juniperus communis | Healthy | - | - | - | |
| DS6-4 | Sorbus aucuparia | Rust | - | - | - | |
| DS6-5 | Alnus | Healthy | - | - | - | |
| DS6-6 | Pinus sylvestris | Healthy | P. kernoviae | - | - | |
| DS6-7 | Betula | Declining | - | - | - | |
| DS6-8 | Alnus | Healthy | - | - | - | |
| DS6-9 | Alnus | Declining | - | - | - | |
| DS6-10 | Prunus | Declining | - | - | - | |
| DS7 (Woodland/Recreational Park) | DS7-1 | Alnus | Dead | P. austrocedri, Clade 11a Phytophthora sp. uncultured 18a and 19a | P. quercina, New Clade Phytophthora sp. uncultured 14b, New Clade Phytophthora sp. uncultured 16b | - |
| DS7-2 | Alnus | Declining | P. plurivora/citricola, P. pseudosyringae, P. syringae, Clade 7 Phytophthora sp. uncultured 1a, Clade 3 Phytophthora sp. uncultured 11a | P. plurivora, P. syringae | - | |
| DS7-3 | Young Abies grandis in Alnus and Betula forest | Healthy | P. cinnamomi, P. uniformis, Clade 7 Phytophthora sp. uncultured 1a and 16a | Clade 7 Phytophthora sp. uncultured 7b and 8b, New Clade Phytophthora sp. uncultured 16b, Nothophytophthora sp. 4 | - | |
| DS7-4 | Quercus in mature woodland | Healthy | P. plurivora/citricola | - | - | |
| DS7-5 | Crataegus in mature woodland | Dead | P. capsici, P. castaneae, P. cinnamomi, P. fallax, P. foliorum, P. obscura, P. plurivora/citricola, P. rubi/fragariae, P. siskiyouensis, Clade 2 Phytophthora sp. uncultured 17a | - | - | |
| DS8 (Woodland/Recreational Park) | DS8-1 | Larix | Dead | - | Nothophytophthora sp. 2 | - |
| DS8-2 | Fraxinus | Dead | P. syringae | P. syringae, New Clade Phytophthora sp. uncultured 13b, Nothophytophthora sp. 2 | - | |
| DS8-3 | Alnus | Healthy | P. pseudosyringae, P. syringae, Clade 7 Phytophthora sp. uncultured 6a, Clade 3 Phytophthora sp. uncultured 11a | P. pseudosyringae, P. syringae | - | |
| DS8-4 | Pinus nigra | Healthy | Clade 7 Phytophthora sp. uncultured 7a | Nothophytophthora sp. 5 | - | |
| DS8-5 | Fraxinus stump, decayed tree in mature woodland | NA | P. capsici, P. castaneae, P. fallax, P. foliorum, P. obscura, P. plurivora/citricola, P. rubi/fragariae, P. syringae, Clade 7 Phytophthora sp. uncultured 5a, Clade 6 Phytophthora sp. uncultured 8a, Clade 1 Phytophthora sp. uncultured 13a | P. syringae, Clade 1 Phytophthora sp. uncultured 1b, Clade 6 Phytophthora sp. uncultured 4b, Clade 7 Phytophthora sp. uncultured 6b | - | |
| DS9 (Arboreta) | DS9-1 | Chamaecyparis thyoides, Picea brachytyla | Healthy | P. austrocedri, P. cinnamomi, P. plurivora/citricola | New Clade Phytophthora sp. uncultured 16b | P. chlamydospora |
| DS9-2 | Grassland, Parrotia persica, Pseudotsuga menziesii | Healthy | P. syringae | Nothophytophthora sp. 7 | P. chlamydospora, P. ramorum | |
| DS9-3 | Quercus cerris | Healthy | - | - | - | |
| DS9-4 | Pinus sylvestris | Healthy | P. primulae | P. primulae | - | |
| DS9-5 | Quercus robur | Healthy | P. fallax, P. foliorum, P. idaei, P. plurivora/citricola, Clade 1 Phytophthora sp. uncultured 12a and 13a, Clade 2 Phytophthora sp. uncultured 17a | New Clade Phytophthora sp. uncultured 15b, New Clade Phytophthora sp. uncultured 17b | - | |
| DS9-6 | Taxus baccata | Decline | P. syringae, Clade 6 Phytophthora sp. uncultured 15a | - | P. gonapodyides | |
| DS9-7 | Mixed species, new plantings: Acer, Cornus, Calocedrus, Fraxinus | Healthy | P. austrocedri | New Clade Phytophthora sp. uncultured 12b, New Clade Phytophthora sp. uncultured 17b | - | |
| DS9-8 | Cedrus atlantica | Healthy | - | - | - | |
| DS9-9 | Quercus petraea, Quercus ilex | Healthy | P. foliorum, P. rubi/fragariae, Clade 11 Phytophthora sp. uncultured 18a and 19a | - | P. plurivora | |
| DS9-10 | Prumnopitys andina, Pinus contorta, boundary with Larix | Healthy | P. primulae | New Clade Phytophthora sp. uncultured 16b | - | |
| DS9-11 | Pseudotsuga menziesii | Healthy | P. austrocedri | - | - | |
| US1 (Undisturbed site 1) | US1-1 | Larix | Dead | - | - | - |
| US1-2 | Sorbus aucuparia | Declining | P. obscura, P. ramorum | P. ramorum | - | |
| US1-3 | Crataegus | Declining | P. syringae | - | - | |
| US1-4 | Betula | Dead | P. ramorum | - | - | |
| US1-5 | Betula | Declining | - | - | - | |
| US1-6 | Sorbus aucuparia | Dead | P. plurivora/citricola, Clade 2 Phytophthora sp. uncultured 17a | - | - | |
| US1-7 | Quercus | Healthy | - | - | - | |
| US1-8 | Malus | Declining | P. syringae, Clade 6 Phytophthora sp. uncultured 15a | P. syringae, P. gonapodyides | - | |
| US1-9 | Fraxinus | Declining | P. fallax, P. plurivora/citricola, P. rubi/fragariae, P. siskiyouensis, P. syringae | - | - | |
| US1-10 | Alnus | Healthy | P. plurivora/citricola, P. pseudosyringae, P. syringae | - | - | |
| US1-11 | Alnus | Healthy | P. cactorum, P. syringae | - | - | |
| US2 (Undisturbed site 2) | US2-1 | Betula pubescens | Declining | Clade 7 Phytophthora sp. uncultured 21a | - | - |
| US2-2 | Juniperus communis | Dead | P. kernoviae | - | - | |
| US2-3 | Mature Pinus sylvestris | Healthy | P. cactorum | - | - | |
| US2-4 | Sorbus aucuparia | Healthy | P. syringae, Clade 7 Phytophthora sp. uncultured 2a | - | - | |
| US2-5 | Betula pubescens | Declining | Clade 11 Phytophthora sp. uncultured 19a | - | - | |
| US2-6 | Pinus sylvestris | Healthy | - | - | - | |
| US2-7 | Pinus sylvestris | Healthy | - | - | - | |
| US2-8 | Quercus | Healthy | - | New Clade Phytophthora sp. uncultured 12b, New Clade Phytophthora sp. uncultured 16b | - | |
| US2-9 | Quercus | Declining | - | - | - | |
| US2-10 | Juniperus communis | Declining | - | - | - | |
| US3 (Undisturbed site 3) | US3-1 | Fraxinus | Declining | P. cactorum, P. syringae | P. syringae | - |
| US3-2 | Corylus | Declining | P. cactorum, P. capsici, P. fallax, P. foliorum, P. plurivora/citricola, P. siskiyouensis, P. syringae | P. plurivora, P. syringae, Clade 7 Phytophthora sp. uncultured 9b, Nothophytophthora sp. 6b | P. plurivora | |
| US3-3 | Fraxinus windblown trees | NA | P. cactorum, P. plurivora/citricola, P. primulae, P. syringae | P. cactorum, P. plurivora/citricola, P. syringae, Nothophytophthora sp. 1 | - | |
| US3-4 | Decayed Acer campestre | NA | P. cactorum, P. megasperma/crassamura, P. syringae, Clade 7 Phytophthora sp. uncultured 5a | P. cactorum, P. syringae, Clade 7 Phytophthora sp. uncultured 10b, Nothophytophthora sp. 1 | - | |
| US3-5 | Fraxinus | Dead | - | - | - | |
| US3-6 | Decayed Fagus | Dead | P. syringae | P. syringae | - | |
| US3-7 | Acer campestre | Dead | P. ramorum, P. syringae | P. syringae | - | |
| US3-8 | Betula | Declining | P. cactorum, P. foliorum, P. plurivora/citricola, P. rubi/fragariae, P. syringae, Clade 6 Phytophthora sp. uncultured 15a | P. syringae | P. chlamydospora | |
| US3-9 | Alnus | Declining | P. cactorum, P. pseudosyringae, P. syringae, | P. syringae, New Clade Phytophthora sp. uncultured 14b | P. cactorum, P. plurivora | |
| US3-10 | Betula | Dead | P. cactorum, P. primulae, P. syringae, Clade 3 Phytophthora sp. uncultured 11a | P. primulae | P. plurivora | |
| US4 (Undisturbed site 4) | US4-1 | Decayed Betula | Dead | P. cinnamomi | - | - |
| US4-2 | Decayed Quercus | Dead | P. cinnamomi, P. pseudosyringae | - | - | |
| US4-3 | Fraxinus | Healthy | P. plurivora/citricola, P. pseudosyringae, P. syringae, Clade 3 Phytophthora sp. uncultured 11a, Clade 11 Phytophthora sp. uncultured 20a | P. pseudosyringae, P. syringae, New Clade Phytophthora sp. uncultured 13b | - | |
| US4-4 | Castanea sativa | Healthy | - | - | - | |
| US4-5 | Fagus sylvatica | Healthy | - | - | - | |
| US4-6 | Betula | Dead | P. cactorum, P. cinnamomi, P. lacustris, P. megasperma/crassamura, P. syringae, Clade 7 Phytophthora sp. uncultured 4a and 5a, Clade 6 Phytophthora sp. uncultured 8a | P. cinnamomi, P. megasperma, P. syringae | P. chlamydospora, P. gonapodyides | |
| US4-7 | Fagus sylvatica | Healthy | P. pseudosyringae, Clade 3 Phytophthora sp. uncultured 11a | P. pseudosyringae | - | |
| US4-8 | Quercus | Healthy | - | - | - | |
| US4-9 | Ulmus glabra | Healthy | P. cactorum, P. syringae | Clade 1 Phytophthora sp. uncultured 3b | - | |
| US4-10 | Acer campestre | Declining | P. syringae, Clade 7 Phytophthora sp. uncultured 4a, Clade 6 Phytophthora sp. uncultured 9a, Nothophytophthora sp. | Nothophytophthora sp. 3, Nothophytophthora sp. 8 | P. plurivora | |
| US5 (Undisturbed site 5) | US5-1 | Decayed Quercus robur | Dead | P. ramorum | - | - |
| US5-2 | Tsuga heterophylla | Healthy | - | - | - | |
| US5-3 | Picea sitchensis | Healthy | - | - | - | |
| US5-4 | Picea sitchensis | Declining | - | - | - | |
| US5-5 | Bog | NA | - | - | - | |
| US5-6 | Salix | Healthy | - | - | - | |
| US5-7 | Pinus contorta | Declining | P. capsici, P. foliorum, P. rubi/fragariae, Clade 2 Phytophthora sp. uncultured 17a | - | - | |
| US5-8 | Decayed Betula | Dead | P. capsici, P. foliorum, P. plurivora/citricola, Clade 2 Phytophthora sp. uncultured 17a | - | - | |
| US5-9 | Fagus sylvatica | Healthy | - | - | - | |
| US5-10 | Abies procera | Healthy | - | - | - |
| ITS | COI | ||||
|---|---|---|---|---|---|
| Clade | Phytophthora spp. | Reads (%) | Incidence (%) | Reads (%) | Incidence (%) |
| 10 | P. boehmeriae | 0.0 | 0.0 | 5.5 | 100.0 |
| 2 | P. capsici | 16.1 | 100.0 | 30.8 | 100.0 |
| 9 | P. fallax | 3.4 | 100.0 | 13.7 | 100.0 |
| 8 | P. foliorum | 3.8 | 66.7 | 0.0 | 0.0 |
| 1 | P. idaei | 1.6 | 66.7 | 9.1 | 66.7 |
| 5 | P. castaneae | 12.6 | 100.0 | 7.9 | 100.0 |
| 8 | P. obscura | 12.0 | 100.0 | 13.1 | 100.0 |
| 2 | P. plurivora | 27.3 | 100.0 | 6.1 | 66.7 |
| 7 | P. rubi/fragariae | 17.9 | 100.0 | 1.5 | 33.3 |
| 2 | P. siskiyouensis | 5.4 | 66.7 | 12.4 | 100.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Landa, B.B.; Arias-Giraldo, L.F.; Henricot, B.; Montes-Borrego, M.; Shuttleworth, L.A.; Pérez-Sierra, A. Diversity of Phytophthora Species Detected in Disturbed and Undisturbed British Soils Using High-Throughput Sequencing Targeting ITS rRNA and COI mtDNA Regions. Forests 2021, 12, 229. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020229
Landa BB, Arias-Giraldo LF, Henricot B, Montes-Borrego M, Shuttleworth LA, Pérez-Sierra A. Diversity of Phytophthora Species Detected in Disturbed and Undisturbed British Soils Using High-Throughput Sequencing Targeting ITS rRNA and COI mtDNA Regions. Forests. 2021; 12(2):229. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020229
Chicago/Turabian StyleLanda, Blanca B., Luis F. Arias-Giraldo, Béatrice Henricot, Miguel Montes-Borrego, Lucas A. Shuttleworth, and Ana Pérez-Sierra. 2021. "Diversity of Phytophthora Species Detected in Disturbed and Undisturbed British Soils Using High-Throughput Sequencing Targeting ITS rRNA and COI mtDNA Regions" Forests 12, no. 2: 229. https://0-doi-org.brum.beds.ac.uk/10.3390/f12020229