
Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy



Abstract
:1. Introduction
2. Materials and Methods
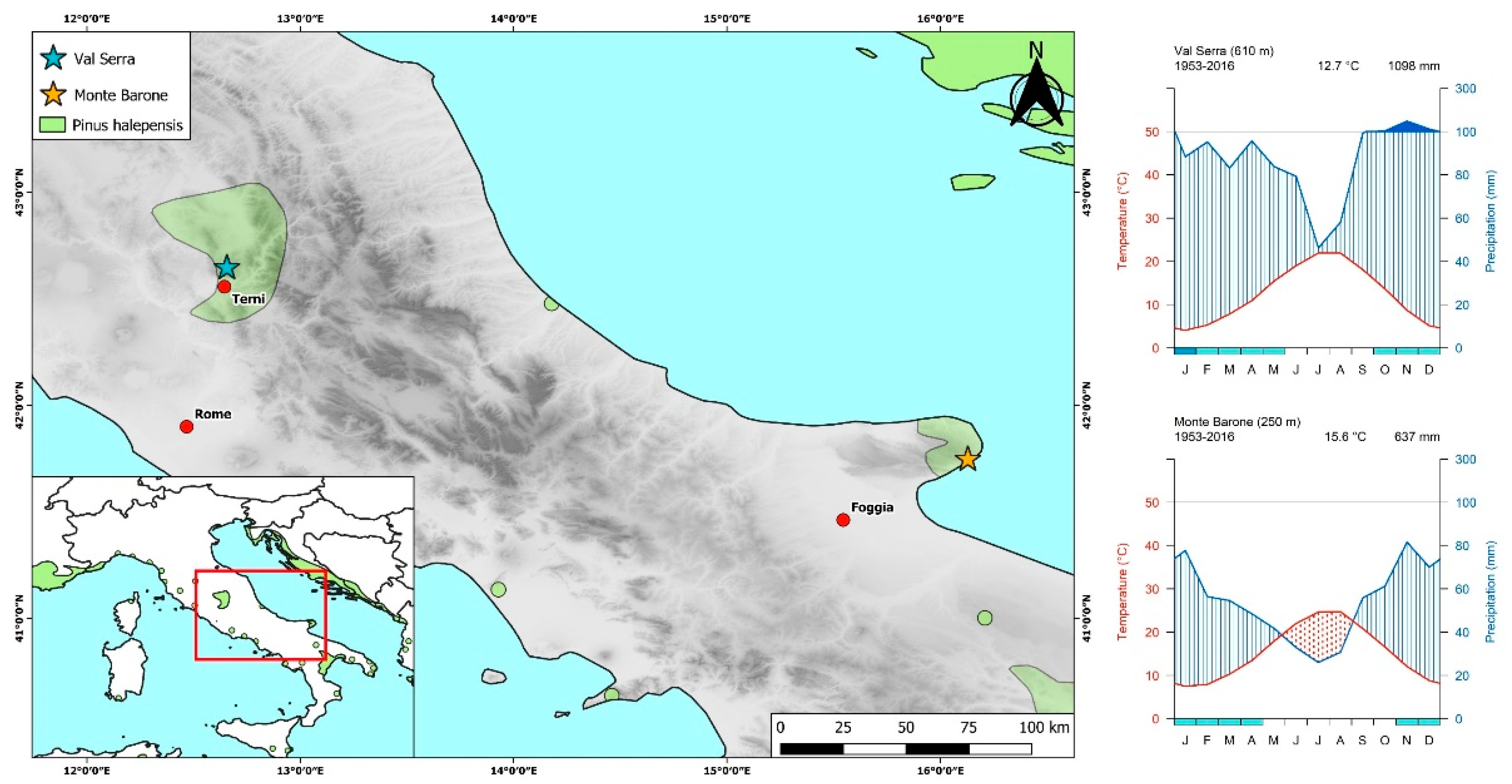
2.1. Study Sites
2.2. Tree-Ring Data Collection and Analysis
2.3. The Climate-Growth Relationships
2.4. Satellite Data Analysis
3. Results
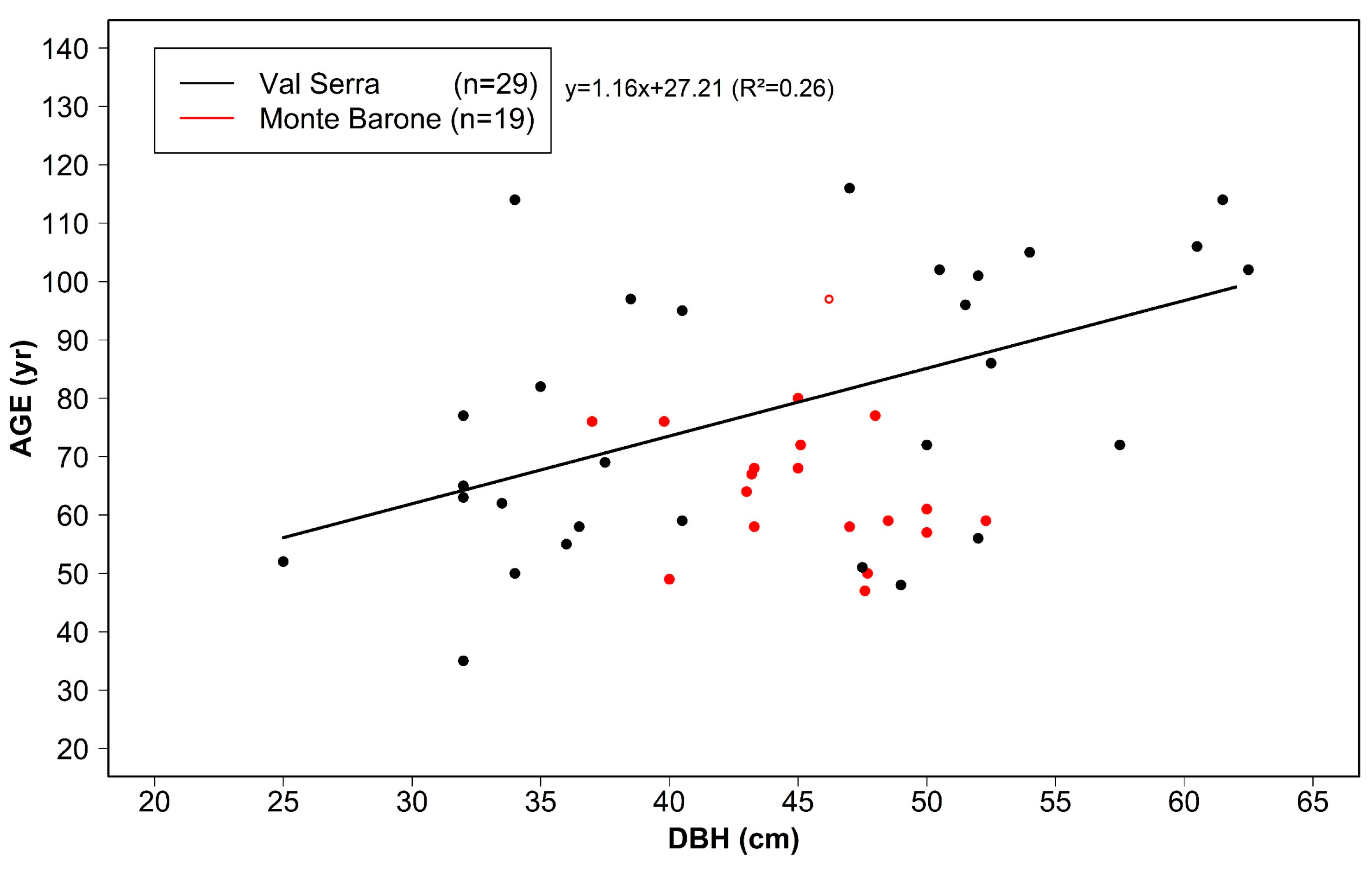
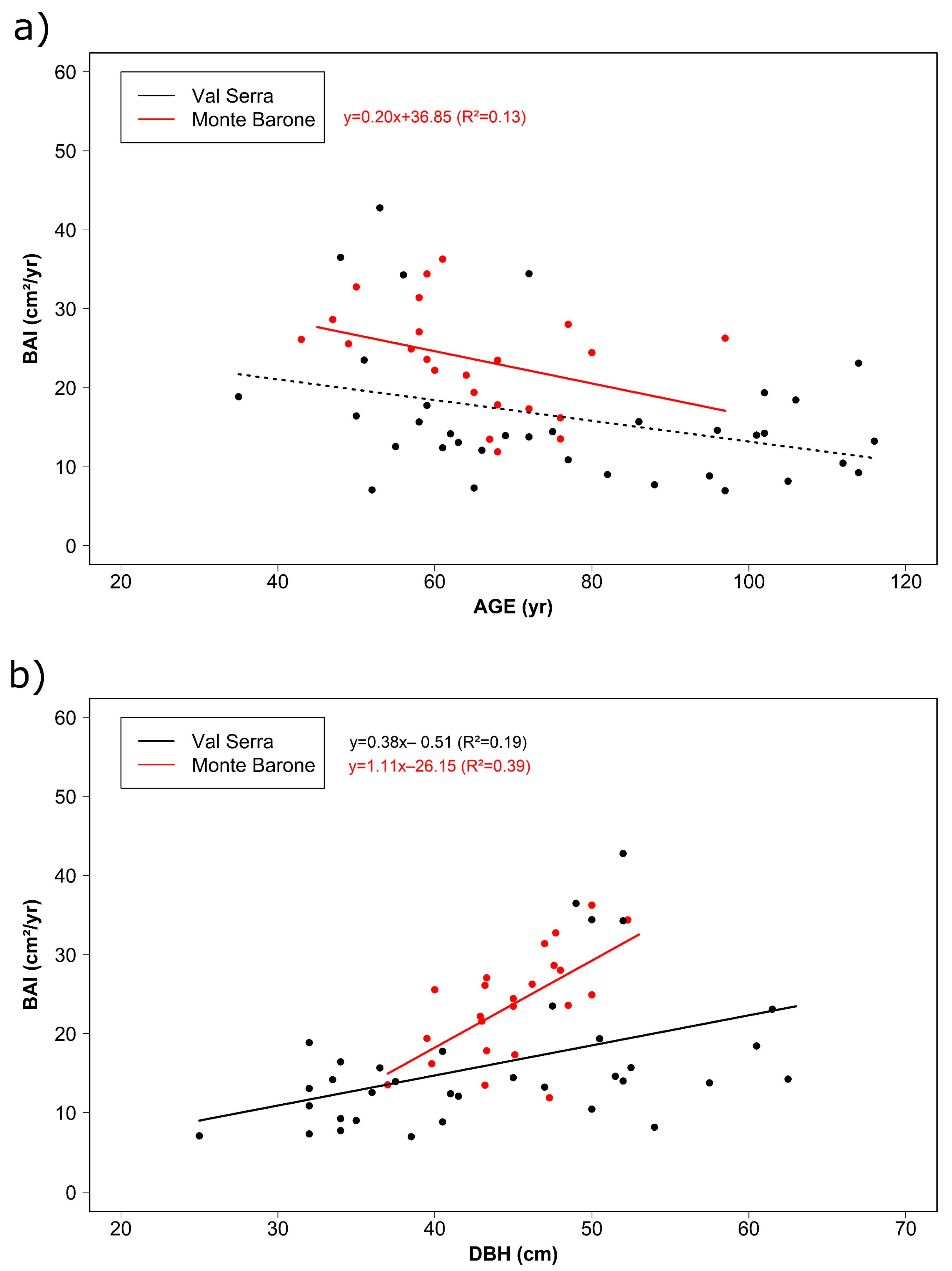
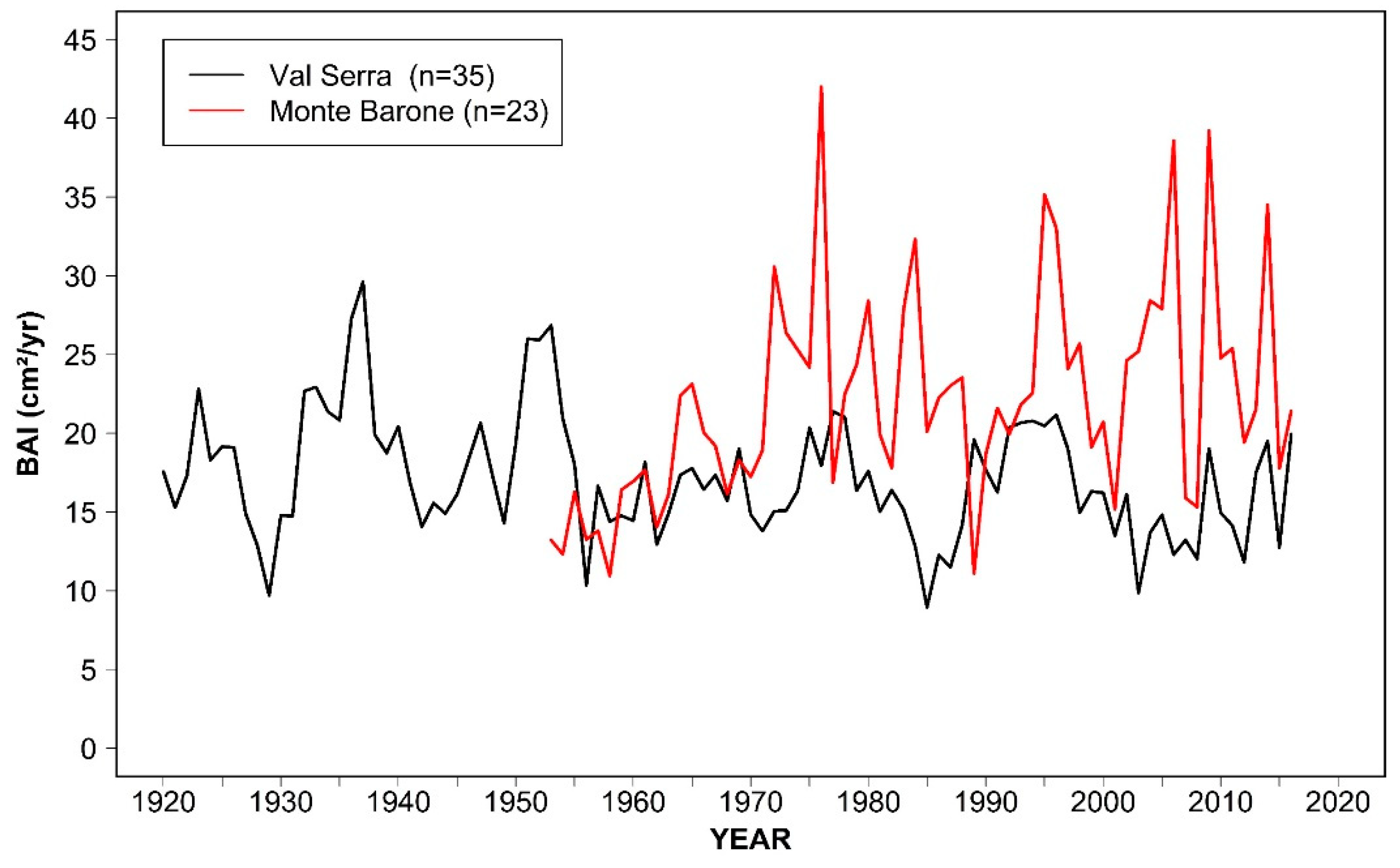
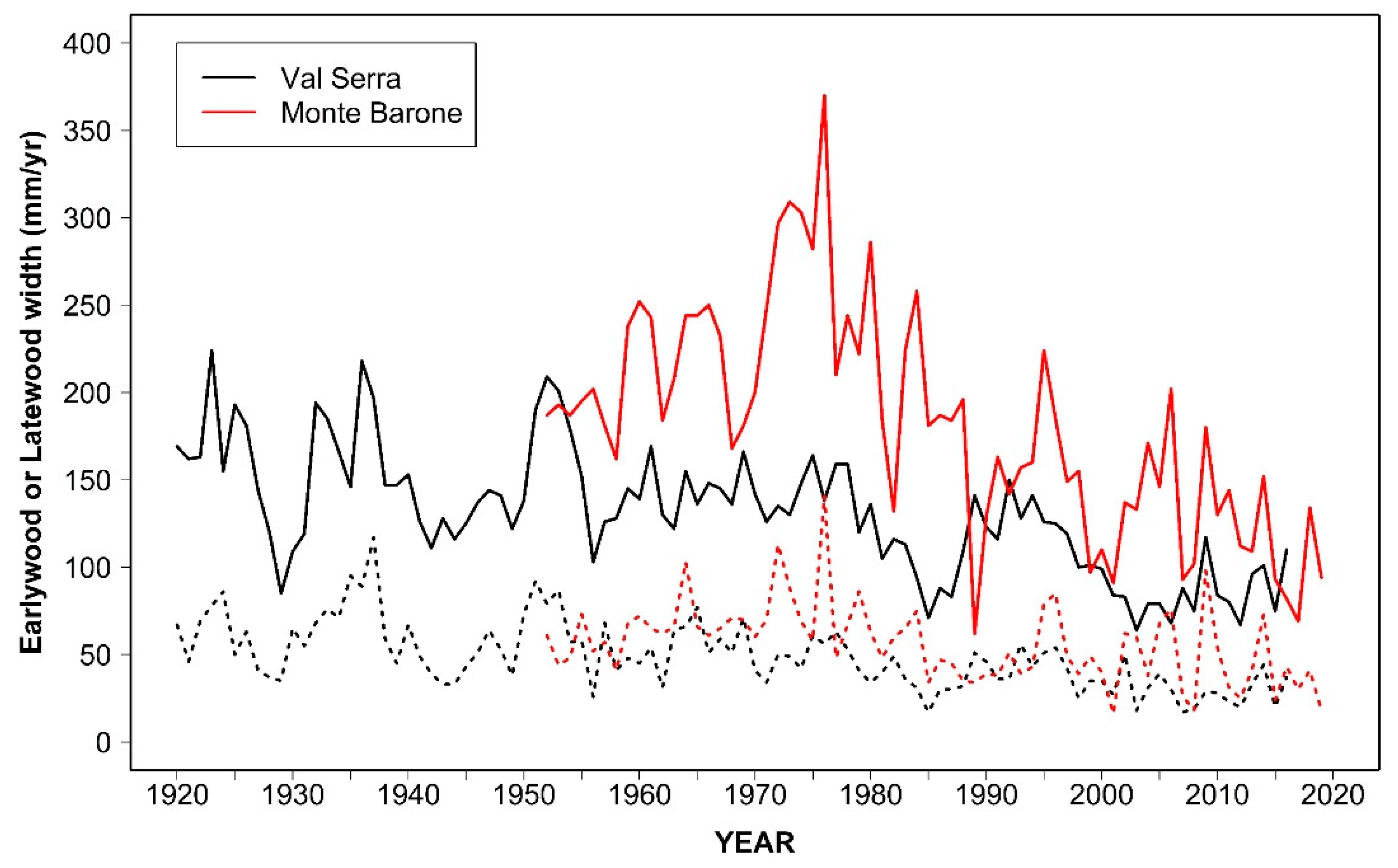
3.1. Tree Age, Size, and Growth Rates
3.2. Climate-Growth Relationships
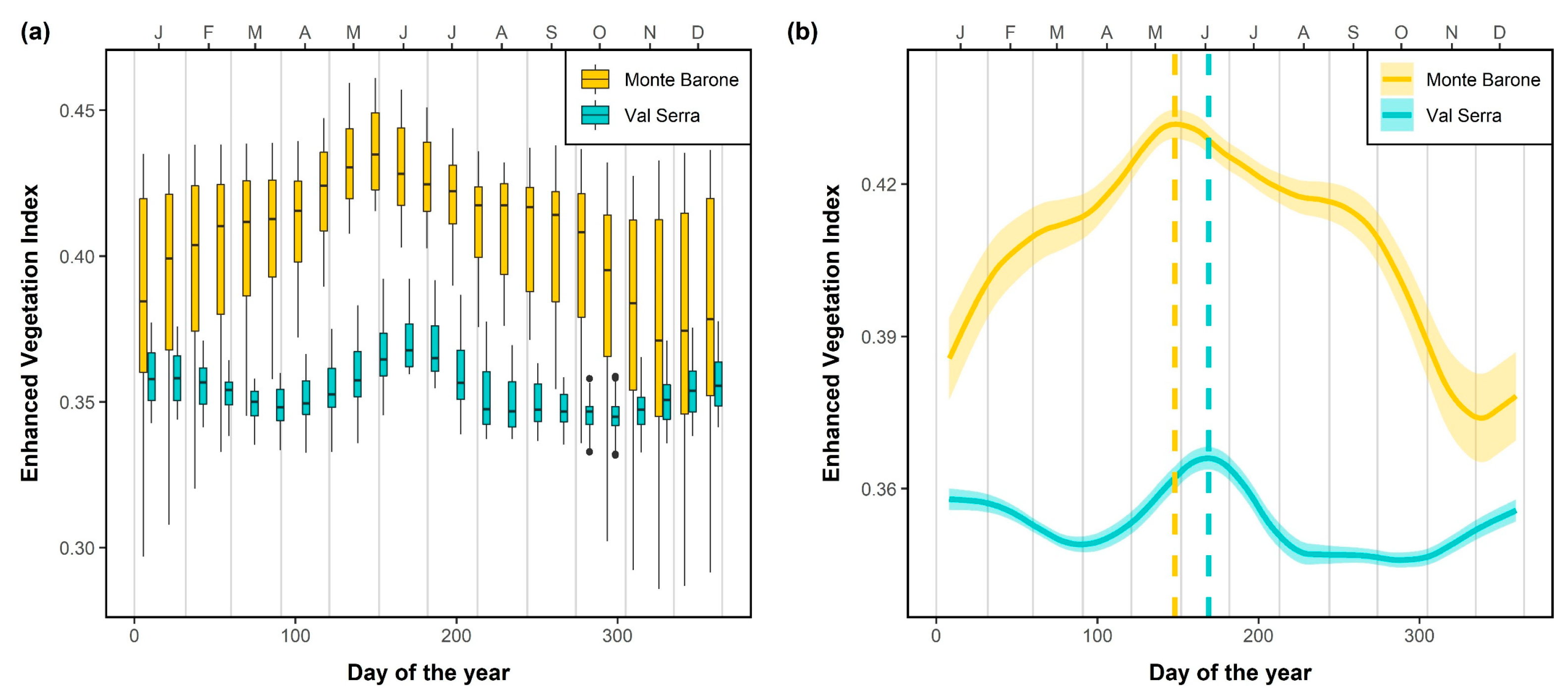
3.3. Satellite Data Analysis
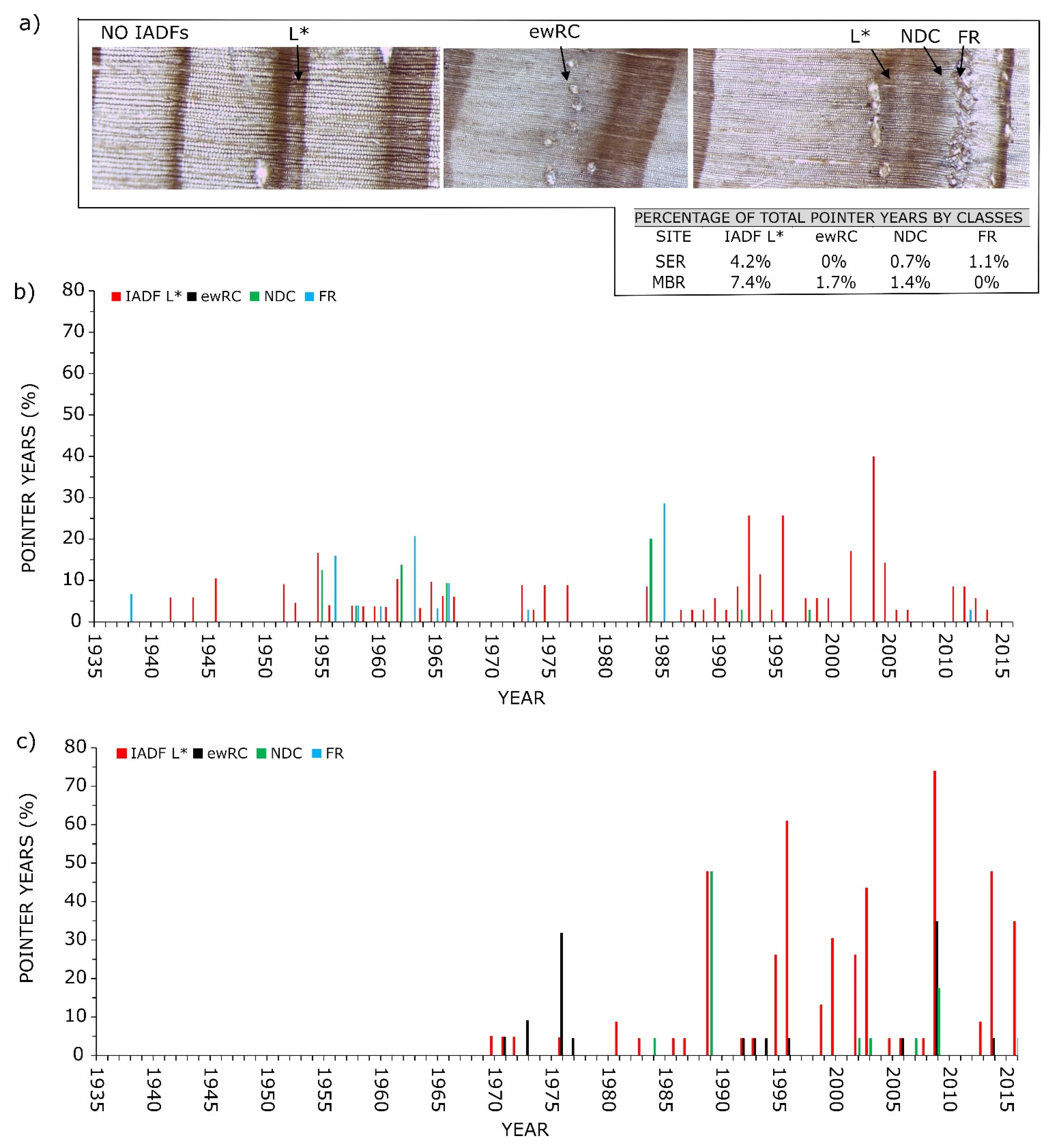
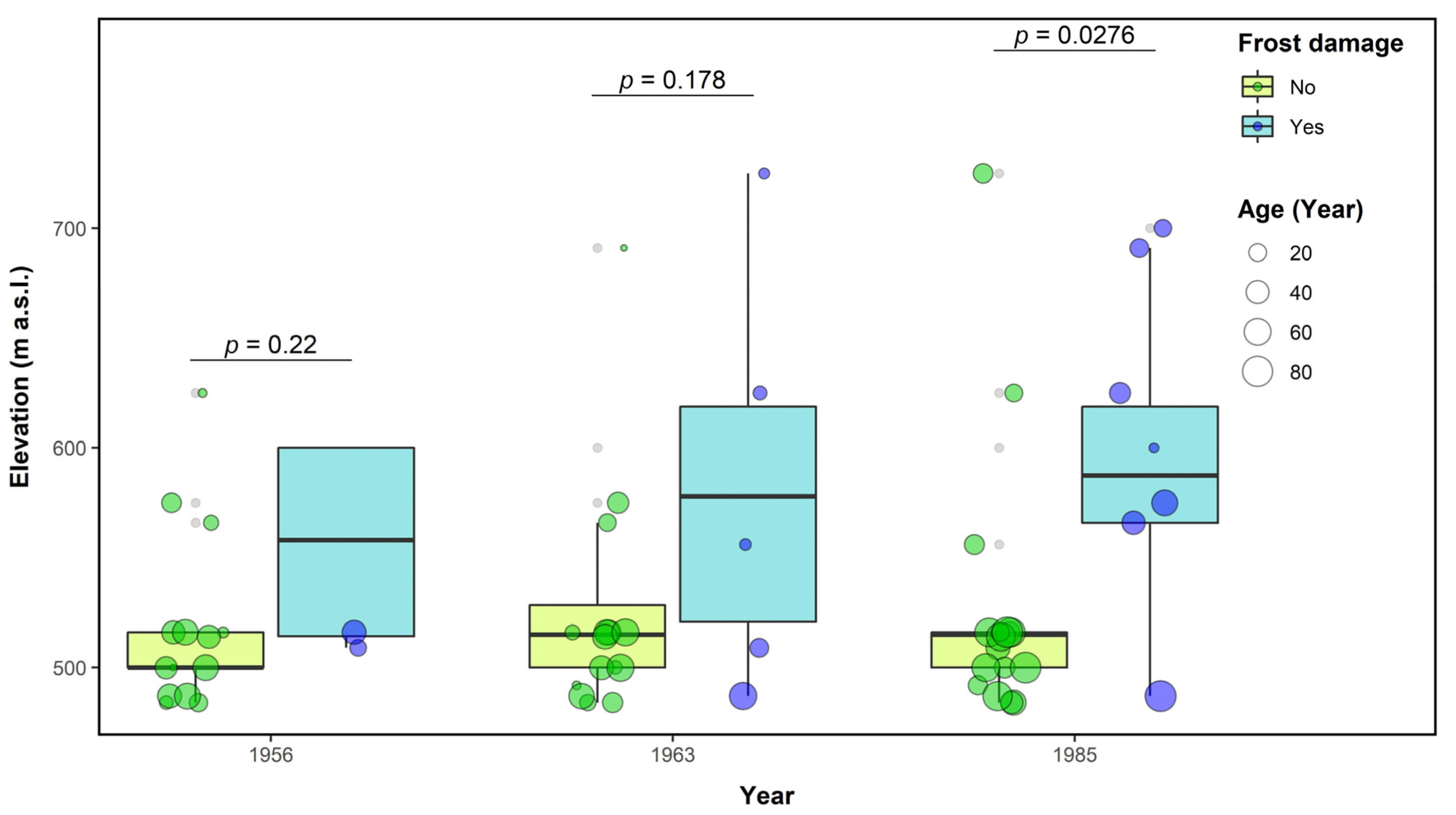
3.4. Pointer Years and Frost Events
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Cuttelod, A.; García, N.; Malak, D.A.; Temple, H.; Katariya, V. The Mediterranean: A biodiversity hotspot under threat. In Wildlife in a Changing World—An Analysis of the 2008 IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2008. [Google Scholar]
- Hoerling, M.; Eischeid, J.; Perlwitz, J.; Quan, X.; Zhang, T.; Pegion, P. On the increased frequency of mediterranean drought. J. Clim. 2012, 25, 2146–2161. [Google Scholar] [CrossRef] [Green Version]
- Lionello, P.; Scarascia, L. The relation between climate change in the Mediterranean region and global warming. Reg. Environ. Chang. 2018, 18, 1481–1493. [Google Scholar] [CrossRef]
- Sánchez, E.; Gallardo, C.; Gaertner, M.A.; Arribas, A.; Castro, M. Future climate extreme events in the Mediterranean simulated by a regional climate model: A first approach. Glob. Planet. Chang. 2004, 44, 163–180. [Google Scholar] [CrossRef]
- Chambel, M.R.; Climent, J.; Pichot, C.; Ducci, F. Mediterranean Pines (Pinus halepensis Mill. and brutia Ten.). In Forest Tree Breeding in Europe; Springer: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-6145-2. [Google Scholar]
- De Luis, M.; Čufar, K.; Di Filippo, A.; Novak, K.; Papadopoulos, A.; Piovesan, G.; Rathgeber, C.B.K.; Raventós, J.; Saz, M.A.; Smith, K.T. Plasticity in dendroclimatic response across the distribution range of Aleppo pine (Pinus halepensis). PLoS ONE 2013, 8, e83550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borghetti, M.; Cinnirella, S.; Magnani, F.; Saracino, A. Impact of long-term drought on xylem embolism and growth in Pinus halepensis Mill. Trees Struct. Funct. 1998, 12, 187–195. [Google Scholar] [CrossRef]
- Klein, T. Eco-Physiology of Water Use in Pinus halepensis: From Leaf to Forest Scale. Ph.D. Thesis, Scientific Council of the Weizmann Institute of Science, Rehovot, Israel, 2012. [Google Scholar]
- Hover, A.; Buissart, F.; Caraglio, Y.; Heinz, C.; Pailler, F.; Ramel, M.; Vennetier, M.; Prévosto, B.; Sabatier, S. Growth phenology in Pinus halepensis Mill.: Apical shoot bud content and shoot elongation. Ann. For. Sci. 2017, 74. [Google Scholar] [CrossRef]
- Pardos, M.; Climent, J.; Gil, L.; Pardos, J.A. Shoot growth components and flowering phenology in grafted Pinus halepensis Mill. Trees Struct. Funct. 2003, 17, 442–450. [Google Scholar] [CrossRef]
- Prislan, P.; Gričar, J.; de Luis, M.; Novak, K.; Del Castillo, E.M.; Schmitt, U.; Koch, G.; Štrus, J.; Mrak, P.; Žnidarič, M.T.; et al. Annual cambial rhythm in Pinus halepensis and Pinus sylvestris as indicator for climate adaptation. Front. Plant. Sci. 2016, 7, 1923. [Google Scholar] [CrossRef] [Green Version]
- Baquedano, F.J.; Valladares, F.; Castillo, F.J. Phenotypic plasticity blurs ecotypic divergence in the response of Quercus coccifera and Pinus halepensis to water stress. Eur. J. For. Res. 2008, 127, 495–506. [Google Scholar] [CrossRef]
- Camarero, J.J.; Olano, J.M.; Parras, A. Plastic bimodal xylogenesis in conifers from continental Mediterranean climates. New Phytol. 2010, 185, 471–480. [Google Scholar] [CrossRef] [PubMed]
- Campelo, F.; Nabais, C.; Freitas, H.; Gutiérrez, E. Climatic significance of tree-ring width and intra-annual density fluctuations in Pinus pinea from a dry Mediterranean area in Portugal. Ann. For. Sci. 2007, 64, 229–238. [Google Scholar] [CrossRef] [Green Version]
- Zalloni, E.; de Luis, M.; Campelo, F.; Novak, K.; De Micco, V.; Di Filippo, A.; Vieira, J.; Nabais, C.; Rozas, V.; Battipaglia, G. Climatic signals from intra-annual density fluctuation frequency in mediterranean pines at a regional scale. Front. Plant. Sci. 2016, 7, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Novak, K.; de Luis, M.; Saz, M.A.; Longares, L.A.; Serrano-Notivoli, R.; Raventós, J.; Čufar, K.; Gričar, J.; Di Filippo, A.; Piovesan, G.; et al. Missing rings in Pinus halepensis—The missing link to relate the tree-ring record to extreme climatic events. Front. Plant. Sci. 2016, 7, 727. [Google Scholar] [CrossRef] [PubMed]
- Fritts, H.C.; Swetnam, T.W. Dendroecology: A tool for evaluating variations in past and present forest environments. Adv. Ecol. Res. 1989, 19, 111–188. [Google Scholar]
- Piermattei, A.; Crivellaro, A.; Carrer, M.; Urbinati, C. The “blue ring”: Anatomy and formation hypothesis of a new tree-ring anomaly in conifers. Trees Struct. Funct. 2015, 29, 613–620. [Google Scholar] [CrossRef]
- Montwé, D.; Isaac-Renton, M.; Hamann, A.; Spiecker, H. Cold adaptation recorded in tree rings highlights risks associated with climate change and assisted migration. Nat. Commun. 2018, 9, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Barinov, V.V.; Myglan, V.S.; Taynik, A.V.; Oydupa, O.C.; Vaganov, E.A. Extreme climatic events in the Republic of Tuva according to tree-ring analysis. Contemp. Probl. Ecol. 2015, 8, 414–422. [Google Scholar] [CrossRef]
- Pedrotti, F. Notizia di ricerche fitosociologiche sulle pinete a Pino d’Aleppo della valle del serra (Terni). Mitt. Ostalp. Pflanz. Arb. 1967, 7, 139–142. [Google Scholar]
- Caudullo, G.; Welk, E.; San-Miguel-Ayanz, J. Chorological maps for the main European woody species. Data Br. 2017, 12, 662–666. [Google Scholar] [CrossRef]
- Novak, K.; Čufar, K.; De Luis, M.; Sánchez, M.A.S.; Raventós, J. Age, climate and intra-annual density fluctuations in Pinus halepensis in Spain. IAWA J. 2013, 34, 459–474. [Google Scholar] [CrossRef] [Green Version]
- Di Filippo, A.; Biondi, F.; Maugeri, M.; Schirone, B.; Piovesan, G. Bioclimate and growth history affect beech lifespan in the Italian Alps and Apennines. Glob. Chang. Biol. 2012, 18, 960–972. [Google Scholar] [CrossRef]
- Bunn, A.; Korpela, M.; Biondi, F.; Campelo, F.; Mérian, P.; Qeadan, F.; Zang, C.; Buras, A.; Cecile, J.; Mudelsee, M.; et al. (Eds.) Package “dplR”, Version 1.7.1. In Dendrochronology Program Library in R; R Foundation: Vienna, Austria, 2015. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020. [Google Scholar]
- Di Filippo, A.; Biondi, F.; Čufar, K.; De Luis, M.; Grabner, M.; Maugeri, M.; Presutti Saba, E.; Schirone, B.; Piovesan, G. Bioclimatology of beech (Fagus sylvatica L.) in the Eastern Alps: Spatial and altitudinal climatic signals identified through a tree-ring network. J. Biogeogr. 2007, 34, 1873–1892. [Google Scholar] [CrossRef]
- Brunetti, M.; Maugeri, M.; Nanni, T.; Simolo, C.; Spinoni, J. High-resolution temperature climatology for Italy: Interpolation method intercomparison. Int. J. Climatol. 2014, 34, 1278–1296. [Google Scholar] [CrossRef] [Green Version]
- Louis, J.; Debaecker, V.; Pflug, B.; Main-Knorn, M.; Bieniarz, J.; Mueller-Wilm, U.; Cadau, E.; Gascon, F. Sentinel-2 SEN2COR: L2A processor for users. Eur. Sp. Agency (Spec. Publ.) ESA SP 2016, SP-740, 9–13. [Google Scholar]
- Gorelick, N.; Hancher, M.; Dixon, M.; Ilyushchenko, S.; Thau, D.; Moore, R. Google Earth Engine: Planetary-scale geospatial analysis for everyone. Remote Sens. Environ. 2017, 202, 18–27. [Google Scholar] [CrossRef]
- Aybar, C.; Wu, Q.; Bautista, L.; Yali, R.; Barja, A. rgee: An R package for interacting with Google Earth Engine. J. Open Source Softw. 2020, 5, 2272. [Google Scholar] [CrossRef]
- Huete, A.R.; Liu, H.Q.; Batchily, K.; van Leeuwen, W. A comparison of vegetation indices over a global set of TM images for EOS-MODIS. Remote Sens. Environ. 1997, 59, 440–451. [Google Scholar] [CrossRef]
- Huete, A.; Didan, K.; Miura, T.; Rodriguez, E.P.; Gao, X.; Ferreira, L.G. Overview of the radiometric and biophysical performance of the MODIS vegetation indices. Remote Sens. Environ. 2002, 83, 195–213. [Google Scholar] [CrossRef]
- Cai, Z.; Jönsson, P.; Jin, H.; Eklundh, L. Performance of smoothing methods for reconstructing NDVI time-series and estimating vegetation phenology from MODIS data. Remote Sens. 2017, 9, 1271. [Google Scholar] [CrossRef] [Green Version]
- Golub, G.H.; Heath, M.; Wahba, G. Generalized cross-validation as a method for choosing a good ridge parameter. Technometrics 1979, 21, 215–223. [Google Scholar] [CrossRef]
- Aragones, D.; Rodriguez-Galiano, V.F.; Caparros-Santiago, J.A.; Navarro-Cerrillo, R.M. Could land surface phenology be used to discriminate Mediterranean pine species? Int. J. Appl. Earth Obs. Geoinf. 2019, 78, 281–294. [Google Scholar] [CrossRef]
- Karamihalaki, M.; Stagakis, S.; Sykioti, O.; Kyparissis, A.; Parcharidis, I. Monitoring drought effects on mediteranean conifer forests using spot-vegetation NDVI and NDWI timeseries. Eur. Sp. Agency (Spec. Publ.) ESA SP 2016, SP-740, 603. [Google Scholar]
- Maselli, F. Monitoring forest conditions in a protected Mediterranean coastal area by the analysis of multiyear NDVI data. Remote Sens. Environ. 2004, 89, 423–433. [Google Scholar] [CrossRef]
- Zeng, L.; Wardlow, B.D.; Xiang, D.; Hu, S.; Li, D. A review of vegetation phenological metrics extraction using time-series, multispectral satellite data. Remote Sens. Environ. 2020, 237, 111511. [Google Scholar] [CrossRef]
- Chen, R.; Yin, G.; Liu, G.; Li, J.; Verger, A. Evaluation and normalization of topographic effects on vegetation indices. Remote Sens. 2020, 12, 2290. [Google Scholar] [CrossRef]
- Di Filippo, A.; Pederson, N.; Baliva, M.; Brunetti, M.; Dinella, A.; Kitamura, K.; Knapp, H.D.; Schirone, B.; Piovesan, G. The longevity of broadleaf deciduous trees in Northern Hemisphere temperate forests: Insights from tree-ring series. Front. Ecol. Evol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Piovesan, G.; Biondi, F.; Baliva, M.; Dinella, A.; Di Fiore, L.; Marchiano, V.; Presutti Saba, E.; De Vivo, G.; Schettino, A.; Di Filippo, A. Tree growth patterns associated with extreme longevity: Implications for the ecology and conservation of primeval trees in Mediterranean mountains. Anthropocene 2019, 26, 100199. [Google Scholar] [CrossRef]
- Brienen, R.J.W.; Caldwell, L.; Duchesne, L.; Voelker, S.; Barichivich, J.; Baliva, M.; Ceccantini, G.; Di Filippo, A.; Helama, S.; Locosselli, G.M.; et al. Forest carbon sink neutralized by pervasive growth-lifespan trade-offs. Nat. Commun. 2020, 11, 4241. [Google Scholar] [CrossRef]
- Linares, J.C.; Delgado-Huertas, A.; Carreira, J.A. Climatic trends and different drought adaptive capacity and vulnerability in a mixed Abies pinsapo-Pinus halepensis forest. Clim. Chang. 2011, 105, 67–90. [Google Scholar] [CrossRef]
- Stephenson, N.L.; Das, A.J.; Condit, R.; Russo, S.E.; Baker, P.J.; Beckman, N.G.; Coomes, D.A.; Lines, E.R.; Morris, W.K.; Rüger, N.; et al. Rate of tree carbon accumulation increases continuously with tree size. Nature 2014, 507, 90–93. [Google Scholar] [CrossRef] [PubMed]
- Rowland, L.; Da Costa, A.C.L.; Galbraith, D.R.; Oliveira, R.S.; Binks, O.J.; Oliveira, A.A.R.; Pullen, A.M.; Doughty, C.E.; Metcalfe, D.B.; Vasconcelos, S.S.; et al. Death from drought in tropical forests is triggered by hydraulics not carbon starvation. Nature 2015, 528, 119–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Piovesan, G.; Biondi, F.; Di Filippo, A.; Alessandrini, A.; Maugeri, M. Drought-driven growth reduction in old beech (Fagus sylvatica L.) forests of the central Apennines, Italy. Glob. Chang. Biol. 2008, 14, 1265–1281. [Google Scholar] [CrossRef]
- Camarero, J.J.; Gazol, A.; Sangüesa-Barreda, G.; Oliva, J.; Vicente-Serrano, S.M. To die or not to die: Early warnings of tree dieback in response to a severe drought. J. Ecol. 2015, 103, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Rathgeber, C.B.K.; Misson, L.; Nicault, A.; Guiot, J. Bioclimatic model of tree radial growth: Application to the French Mediterranean Aleppo pine forests. Trees Struct. Funct. 2005, 19, 162–176. [Google Scholar] [CrossRef]
- Trenberth, K.E.; Fasullo, J.T. An apparent hiatus in global warming? Earth’s Future 2013, 1, 19–32. [Google Scholar] [CrossRef]
- Galli, M.; Guadalupi, M.; Nanni, T.; Ruggiero, L.; Zuanni, F. Ravenna pine trees as monitors of winter severity in N-E Italy. Theor. Appl. Climatol. 1992, 45, 217–224. [Google Scholar] [CrossRef]
- Hadad, M.; Tardif, J.C.; Conciatori, F.; Waito, J.; Westwood, A. Climate and atmospheric circulation related to frost-ring formation in Picea mariana trees from the Boreal Plains, interior North America. Weather Clim. Extrem. 2020, 29, 100264. [Google Scholar] [CrossRef]
- Calamassi, R.; Paoletti, E.; Strati, S. Frost hardening and resistance in three Aleppo pine (Pinus halepensis Mill.) provenances. Isr. J. Plant. Sci. 2001, 49, 179–186. [Google Scholar] [CrossRef]
- Camuffo, D.; Bertolin, C.; Craievich, A.; Granziero, R.; Enzi, S. When the Lagoon was frozen over in Venice from A.D. 604 to 2012: Evidence from written documentary sources, visual arts and instrumental readings. Méditerranée. Rev. Géographique Des Pays Méditerranéens/J. Mediterr. Geogr. 2017. Available online: https://journals.openedition.org/mediterranee/7983 (accessed on 4 March 2021).
- Pignatti, S. I Boschi d'Italia. Sinecologia e Biodiversità; UTET: Torino, Italy, 1998. [Google Scholar]
- Mensing, S.; Tunno, I.; Cifani, G.; Passigli, S.; Noble, P.; Archer, C.; Piovesan, G. Human and climatically induced environmental change in the Mediterranean during the Medieval Climate Anomaly and Little Ice Age: A case from central Italy. Anthropocene 2016, 15, 49–59. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | Samples | Latitude (°) | Longitude (°) | Elevation Range (m a.s.l.) | Slope (°) | Aspect |
|---|---|---|---|---|---|---|
| Val Serra | 35 | 42.6448 | 12.6565 | 490–730 | 26.8 | SW |
| Monte Barone | 23 | 41.7450 | 16.1303 | 200–300 | 12.9 | NE |
| Site | N Samples | DBH 1 (cm) | Age 1 (yr) | BAI 1 (cm²/yr) | Earlywood Width ¹ (mm/yr) | Latewood Width ¹ (mm/yr) | EPS > 0.85 | N Trees 2 |
|---|---|---|---|---|---|---|---|---|
| Val Serra | 35 | 44 ± 10 (25–62) | 79 ± 24 (35–122) | 16.1 ± 3.2 (7.0–42.1) | 1.13 ± 0.70 (0.10–7.26) | 0.39 ± 0.24 (0.02–3.12) | 1920–2016 | 10 |
| Monte Barone | 23 | 45 ± 4 (37–52) | 64 ± 12 (43–97) | 23.7 ± 6.6 (12.0–35.8) | 1.79 ± 0.69 (0.15–9.68) | 0.56 ± 0.22 (0.06–4.53) | 1952–2019 | 10 |
| Year Preceding Growth | Year of Growth | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | JAN | FEB | MAR | APR | MAY | JUN | JUL | AUG | SEP | OCT | NOV | DEC | |
| VAL SERRA | ||||||||||||||||||||
| P | 0.10 | −0.03 | −0.14 | −0.30 | −0.06 | 0.13 | −0.05 | 0.08 | −0.11 | −0.09 | −0.13 | 0.05 | 0.10 | 0.29 | 0.21 | 0.24 | 0.18 | 0.06 | −0.05 | 0.09 |
| Tmax | 0.05 | 0.08 | 0.34 | 0.36 | −0.02 | 0.02 | −0.21 | −0.14 | 0.05 | 0.28 | 0.27 | 0.20 | −0.05 | −0.22 | −0.19 | −0.15 | −0.13 | −0.01 | 0.23 | 0.09 |
| Tmin | 0.03 | 0.13 | 0.30 | 0.30 | 0.02 | 0.10 | −0.12 | −0.30 | −0.03 | 0.19 | 0.18 | 0.24 | 0.04 | −0.07 | −0.07 | −0.05 | −0.04 | 0.05 | 0.14 | 0.21 |
| MONTE BARONE | ||||||||||||||||||||
| P | −0.07 | −0.09 | −0.08 | 0.08 | −0.07 | −0.07 | 0.20 | 0.16 | 0.25 | 0.06 | 0.07 | 0.24 | 0.17 | 0.25 | 0.30 | 0.46 | 0.10 | 0.13 | −0.18 | −0.01 |
| Tmax | 0.16 | 0.05 | 0.05 | 0.02 | 0.05 | −0.10 | 0.00 | 0.09 | −0.09 | −0.07 | −0.27 | −0.20 | −0.09 | −0.09 | −0.20 | −0.41 | −0.27 | −0.11 | 0.06 | 0.17 |
| Tmin | 0.09 | 0.10 | 0.14 | 0.10 | 0.08 | −0.15 | 0.12 | 0.22 | 0.09 | 0.04 | −0.21 | 0.00 | −0.08 | −0.05 | −0.05 | −0.31 | −0.11 | −0.12 | −0.04 | 0.23 |
| Year Preceding Growth | Year of Growth | |||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| May | Jun | Jul | Aug | Sep | Oct | Nov | Dec | JAN | FEB | MAR | APR | MAY | JUN | JUL | AUG | SEP | OCT | NOV | DEC | |
| VAL SERRA | ||||||||||||||||||||
| (a) | ||||||||||||||||||||
| P | 0.08 | 0.00 | −0.13 | −0.12 | 0.06 | 0.02 | −0.04 | 0.05 | −0. 09 | −0.14 | −0.14 | 0.06 | 0.11 | 0.33 | 0.06 | 0.07 | 0.00 | 0.11 | −0.10 | −0.05 |
| Tmax | −0.02 | 0.07 | 0.27 | 0.22 | −0.09 | 0.01 | −0.14 | −0.05 | 0.15 | 0.37 | 0.29 | 0.11 | −0.09 | −0.26 | −0.20 | −0.04 | 0.02 | −0.08 | 0.08 | 0.01 |
| Tmin | −0.01 | 0.16 | 0.25 | 0.19 | 0.03 | 0.11 | −0.09 | −0.16 | 0.12 | 0.29 | 0.20 | 0.15 | 0.07 | −0.06 | −0.13 | 0.04 | 0.05 | 0.03 | −0.05 | 0.05 |
| (b) | ||||||||||||||||||||
| P | 0.15 | 0.05 | −0.08 | −0.27 | −0.09 | 0.08 | −0.01 | 0.03 | −0.07 | −0.01 | −0.17 | 0.00 | 0.03 | 0.11 | 0.38 | 0.49 | 0.27 | −0.07 | −0.03 | 0.19 |
| Tmax | 0.05 | −0.11 | 0.19 | 0.33 | −0.08 | 0.04 | −0.26 | −0.22 | −0.13 | 0.22 | 0.24 | 0.08 | −0.10 | −0.09 | −0.30 | −0.43 | −0.34 | 0.06 | 0.25 | 0.21 |
| Tmin | 0.11 | −0.04 | 0.18 | 0.30 | −0.06 | 0.06 | −0.20 | −0.30 | −0.19 | 0.17 | 0.08 | 0.08 | 0.00 | −0.02 | −0.08 | −0.26 | −0.11 | 0.11 | 0.22 | 0.44 |
| MONTE BARONE | ||||||||||||||||||||
| (a) | ||||||||||||||||||||
| P | −0.15 | −0.07 | −0.05 | 0.09 | −0.06 | −0.01 | 0.20 | 0.13 | 0.23 | 0.16 | 0.10 | 0.26 | 0.18 | 0.19 | 0.09 | 0.26 | −0.10 | −0.01 | −0.06 | −0.03 |
| Tmax | 0.22 | 0.03 | 0.03 | 0.00 | 0.04 | −0.16 | 0.08 | 0.09 | −0.09 | −0.13 | −0.34 | −0.21 | −0.17 | −0.16 | −0.08 | −0.22 | −0.05 | 0.01 | 0.02 | 0.08 |
| Tmin | 0.08 | 0.11 | 0.09 | 0.03 | 0.09 | −0.17 | 0.18 | 0.19 | 0.04 | −0.02 | −0.24 | −0.04 | −0.13 | −0.09 | −0.03 | −0.20 | −0.01 | −0.03 | −0.08 | 0.12 |
| (b) | ||||||||||||||||||||
| P | −0.01 | −0.04 | 0.01 | 0.18 | 0.05 | −0.04 | 0.09 | 0.08 | 0.18 | 0.08 | 0.11 | 0.08 | 0.03 | 0.25 | 0.35 | 0.48 | 0.33 | 0.27 | −0.21 | 0.08 |
| Tmax | 0.08 | −0.02 | 0.05 | −0.07 | −0.05 | −0.06 | 0.01 | 0.02 | −0.09 | −0.01 | −0.27 | −0.10 | 0.11 | 0 | −0.31 | −0.41 | −0.39 | −0.32 | 0.15 | 0.11 |
| Tmin | 0.02 | 0 | 0.10 | 0 | 0.01 | −0.15 | 0 | 0.19 | 0.10 | 0.13 | −0.19 | 0.03 | 0.10 | 0.07 | −0.11 | 0.30 | −0.13 | −0.21 | 0.05 | 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Di Filippo, A.; Baliva, M.; Brunetti, M.; Di Fiore, L. Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy. Forests 2021, 12, 305. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030305
Di Filippo A, Baliva M, Brunetti M, Di Fiore L. Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy. Forests. 2021; 12(3):305. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030305
Chicago/Turabian StyleDi Filippo, Alfredo, Michele Baliva, Michele Brunetti, and Luca Di Fiore. 2021. "Long-Term Tree-Ring Response to Drought and Frost in Two Pinus halepensis Populations Growing under Contrasting Environmental Conditions in Peninsular Italy" Forests 12, no. 3: 305. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030305