
Comparison of Morphological Indexes and the Pathogenicity of Bursaphelenchus xylophilus in Northern and Southern China


1
Co-Innovation Center for Sustainable Forestry in Southern China, College of Forestry, Nanjing Forestry University, Nanjing 210037, China
2
Southern Forestry Collaborative Innovation Center, College of Forestry, Nanjing Forestry University, Nanjing 210037, China
*
Author to whom correspondence should be addressed.
Forests 2021, 12(3), 310; https://0-doi-org.brum.beds.ac.uk/10.3390/f12030310
Submission received: 31 October 2020
/
Revised: 18 February 2021
/
Accepted: 3 March 2021
/
Published: 7 March 2021
(This article belongs to the Special Issue Biological and Bio-Based Management of Forest Pests and Pathogens)
Abstract
:The pine wood nematode (PWN) Bursaphelenchus xylophilus is recognized as a major invasive species in many countries and causes widespread mortality in pine trees. Pine wood nematode disease (PWD) has spread northward from southern China to several areas of Liaoning Province, which has temperatures outside of the optimal range for this disease. To determine whether obvious variations in the population adaptability of PWN are involved in its rapid spread from southern to northern China, this study compared the differences in morphology of eight southern strains and eight northern strains and the pathogenicity of the 16 strains to Pinus thunbergii, the pine species that is the most susceptible to PWD in China, and to P. tabuliformis, the main PWN host in northern Liaoning Province. The southern-strain females were smaller than the northern-strain females, except for strain GD32. The size differences between the males of the different strains were not significant. The difference in pathogenicity between the northern and southern strains to P. tabuliformis was more significant than the difference in their pathogenicity to P. thunbergii. The pathogenicity differentiation among northern strains was lower than that among southern strains, and the northern strains showed stronger pathogenicity to P. tabuliformis. The P. tabuliformis inoculation experiment showed that the pathogenicity of GD32, JS27, FJ14, LN13, and LN06 was significantly higher than that of FJ13. The results suggest that some PWN populations in the southern region, which are better adapted to P. tabuliformis, were likely directly transmitted to the northern region, resulting in the spread of PWD in the northern region. The spread of PWN from the south did not necessarily require a process of adaptation to the host or to the northern climate.
1. Introduction
Pine wood nematode disease (PWD) is a devastating forest disease caused by pine wood nematode (PWN). The stem pests of pine trees are responsible for the short-distance natural transmission medium of the disease, and human economic and logistics activities are the main factors associated with the long-distance transmission and spread of the disease [1]. Symptoms of the PWN disease were first discovered in Japan in 1905 [2], but the PWN was identified as the causal agent behind the disease only in 1971. In the 1980s, the disease was introduced to China and South Korea and then to Portugal and Spain, gradually becoming one of the most dangerous tree diseases worldwide [3,4,5,6,7]. Since the first discovery of PWD in China in 1982, the disease has developed rapidly in just a few decades. Its rapid spread in tropical and subtropical regions has caused serious damage. In recent years, it has gradually invaded the warm temperate zone; its range has crossed the 10 °C average annual temperature boundary and expanded to mid-temperate zones such as Liaoning Province and high-altitude areas such as the Qinling Mountains [8,9]. The reason why the population of PWN that has invaded can expand to high altitude and high-latitude low-temperature regions is associated with the long-distance transport of man-made logistics and economic activities, and the inheritance of low-temperature adaptation of PWN itself [10]. In China, 60 million hectares of pine forest face the threat of a PWD epidemic [11]. In 2019, PWD continued to spread, and the area south of the Yangtze River experienced a breakout. The epidemic developed from a single location into an area with an irregular circular shape and continued to spread rapidly from west to north [12]. According to the 2020 PWN disease epidemic area announcement issued by the Chinese State Forestry and Grassland Administration (No. 4, 2020), the disease has spread to 666 county-level administrative regions in 18 provinces (regions, cities) in China.
The disease can harm more than 70 species, including 58 species of pinaceae trees [13]. PWD is currently known to naturally infect 25 species of pinaceae trees in China, including P. bungeana, P. pinaster, P. densiflora, P. densiflora var. umbraculifera, P. thunbergii, P. massoniana, P. taeda, P. elliottii, P. thunbergii × P. massoniana, P. luchuensis, P. taiwanensis, P. caribaea, P. kesiya, P. yunnan ensis, P. tabuliformis, P. armandii, P. koraiensis, P. virginiana, P. palustris, P. greggii, P. strobus var. chiapensis, Abies holophylla, Larix olgensis, L. kaempferi, and L. principis-rupprechtii [14,15,16,17,18]. These pinaceae trees grow in large numbers from southern to northern China, and most of them are highly or moderately susceptible to PWN. P. tabuliformis is one of the main afforestation tree species in China’s “Three-North” region [19]. Currently, the total area of P. tabuliformis in Liaoning Province is approximately 700 km2, with a total stock volume of approximately 36 million m3. In areas of P. tabuliformis afforestation in Liaoning Province, soil and water conservation, landscaping, timber supplies and the underforest economy have played a substantial role in the development of the ecological environment of the forests [20].
The adaptation of PWN pathogenicity to different pine species is one of the key factors affecting the spread of the disease. A large number of domestic and foreign studies have shown that there are differences in the pathogenicity of PWN from different geographic sources [21,22], and PWN isolates from different host sources exhibit different levels of virulence to different tree species. Siliang (2013) [23] found through inoculation experiments on 18 species of PWN strains that the pathogenicity of different host source strains is different. Overall, the strains of P. massoniana, P. kesiya, P. thunbergii, and P. elliottii are highly pathogenic to P. massoniana. P. elliottii and P. tabuliformis are moderately pathogenic, while P. caribaea is weakly pathogenic. Xuelian (2007) [24] studied the population morphology and pathogenic variation in PWN in China and showed that even strains from the same geographic origin are not completely consistent in their morphology and pathogenicity. Inoculation experiments on two-year-old Pine. thunbergii showed that 90 PWN strains had extremely significant differences in pathogenicity; of these strains, AMA3 from Anhui had the strongest pathogenicity.
Due to the short outbreak of PWN in northern China, there are fewer studies on the pathogenicity of PWN in Liaoning Province. There is also a lack of relevant experimental data on the pathogenicity experiment of PWN artificially inoculated with P. tabuliformis. More research and experimentation are needed to determine whether PWN strains in northern and southern China exhibit differences in morphology and pathogenicity in different hosts and climate regions. This study intends to compare the morphology and pathogenicity of eight strains from southern China and eight strains from Liaoning Province in order to provide further evidence explaining the cause of the outbreak of PWNs in northern China.
2. Materials and Methods
2.1. Pine Seedlings for Testing
Two-year-old P. thunbergia and P. tabuliformis seedlings (purchased in a Dianzhuang flower and seedling shop and originating from Suqian city, Jiangsu Province, China) in pots were subjected to inoculation. The seedlings were kept in the greenhouse of Xiashu Forest Farm at Nanjing Forestry University, watered regularly and managed in the manner way. See Table 1 for more details regarding the tested seedlings.
2.2. Pine Wood Nematodes for Testing
2.3. Activation and Cultivation of Nematodes
Botrytis cinerea is a fungus belonging to Sclerotiniaceae and Botryotinia, and it can be used as a source of nutrients for PWN. B. cinerea was inoculated onto potato dextrose agar (PDA) medium and placed in an incubator at 25 °C for 4–7 days. After the mycelium covered the culture dish, the suspensions of 16 strains isolated in the laboratory were removed from refrigeration at 4 °C and inoculated onto the B. cinerea at an ultraclean workbench. The culture dishes were sealed with sealing film and placed into a constant-temperature incubator at 25 °C. When the B. cinerea had been eaten by the nematodes, the nematodes were separated by the Baermann funnel method to obtain a fresh nematode suspension.
2.4. Morphological Measurement Indexes for the Nematodes (the De Man Formula)
A total of 10 μL of the mixed nematode suspension was dropped onto a glass slide, with the bottom of the slide heated to kill the nematode. Then, a cover slip was placed gently over the slide, which was then viewed under the microscope for observation and measurement. Thirty nematodes of each strain were randomly selected for morphological measurement of the following indicators. N is the number of specimens measured, L is the body length (μm), LW is the maximum body width (μm), a is the body length/maximum body width ratio, b is the body length/tail length ratio, V is the distance from the vulva to the top of the head × 100/Body length, WMB is the width of the middle oesophageal bulb, Stylet is the length of the stylus (μm), Spi is the length of the spicule (μm), and Tail is the length of the tail (μm).
2.5. Determination of Pathogenicity
The propagative nematodes separated with the Baermann funnel method were collected in a 10 mL centrifuge tube and washed with sterile water 3 times to avoid the influence of nematode or fungal secretions on the inoculation result. Inoculation method: The artificial tree bark inoculation method was used for the pine trees. A sterile scalpel blade was used to cut a 0.3 cm deep incision into the tree bark. A small amount of sterile cotton was inserted into the incision. The incision with the cotton inside was wrapped with parafilm, leaving room for the spout of a funnel, and an appropriate amount of sterile water was added through the funnel. After checking for water leakage, the nematode solution (3000 nematodes per pine seedling) was added with a micro-injector. Each pine wood nematode strain was used to inoculate 5 P. thunbergia and 5 P. tabuliformis seedlings, and the control seedlings were injected with sterile water. The inoculations were performed on 22 June 2019. The inoculated pine seedlings were placed in a greenhouse at 30 °C with 18 h of light for cultivation. The presence of pine trees was observed daily, and the earliest time of the appearance of wilting symptoms of pine seedlings and the number of wilting pine seedlings caused by each strain were recorded.
2.6. Pathogenicity Indicators
According to the method of Fuyuan et al. (1998) [25] and Qi et al. (2020) [26], the disease incidence level of individual pine trees was recorded as follows: level 0, normal; level 1, fewer than half of the needles exhibited chlorosis, fewer than 1/4 of the needles exhibited yellowing; level 2, more than half of the needles exhibited chlorosis, 1/4~3/4 of the needles exhibited yellowing; level 3, more than 3/4 of the needles exhibited yellowing, fewer than half of the needles had turned red; and level 4 (number of disease symptom stage is 0–4), more than half of the needles had turned red, the plant is dying or has died.
2.7. Data Collection and Statistical Analysis
The nematodes were observed and counted under a Leica DM500 (Leica microsystems Gmbh, Wetzlar, Germany) microscope, and the nematodes were photographed and measured using the Carl Zeiss M2 imager (Carl Zeiss Microlmaging Gmbh, d-37081, Gottingen, Germany). The whole and part of the nematodes were photographed under 10×/0.30, 20×/0.50, and 40×/0.75 objective lenses (eyepiece is 10×) and corresponding scales were added to the photos, then we measured the length of the morphological index through the line segment to obtain the data. The morphological indexes included the body length and maximum width, the middle esophageal bulb width, the stylus length, the female distance from the vulva to the top of the head, the female tail length and the male spicule length. Each indicator was measured 30 times. SPSS Statistics 25.0 software was used to perform statistical analysis. Single-factor analysis of variance and Duncan multiple comparison analysis of variance were performed on the experimental results. Prism software was used for the mapping.
3. Results
3.1. Comparison of Morphological Indexes of PWN Strains
The morphological indicators of 16 strains from northern and southern China were measured, and the picture showed the overall and partial morphology of pine wood nematodes (Figure 2). According to the measurement results (Table 3 and Table 4), the body length and body width of the male PWNs were smaller than those of the females. There was a significant difference in the body length and the body width among the 16 strains (p < 0.05), but the significant difference between the males was not greater than the females. The females of all strains from the south except GD32 had shorter bodies than the female PWNs from the north, and the body length was between 898.3 μm and 964.5 μm. However, the minimum body length of northern females was 1005.1 μm. The female body width of LN18, LN16, LN14, LN06 and LN04 was significantly higher than that of FJ13, FJ14, GX08, GD29, JS27, JS58 and AH21 (p < 0.05). The female body width of the northern strain was between 29.1 μm and 30.5 μm. Except for GD32, the body width of females of southern strains was between 27.4 μm and 29.5 μm. The average values of the tail length of the northern-strain females were mostly larger than those of the southern-strain females. The tail length of LN18 females reached 40.3 μm, and even LN03 with the shortest tail length among the northern strains also had 35.2 μm. The females of the eight southern strains had tails between 30.7 μm and 37.2 μm. The females of the 16 strains had significant differences in the length of the stylus (p < 0.05), and among them, the stylus length of GX08, LN16, LN18, LN13, LN14, LN11, LN06, FJ13 and AH21 was significantly higher than that of LN04, LN03, FJ14, GD29, DG32, JS27 and JS58 (p < 0.05).
3.2. Comparison of the Pathogenicity of PWN Strains from Southern and Northern China to P. thunbergii
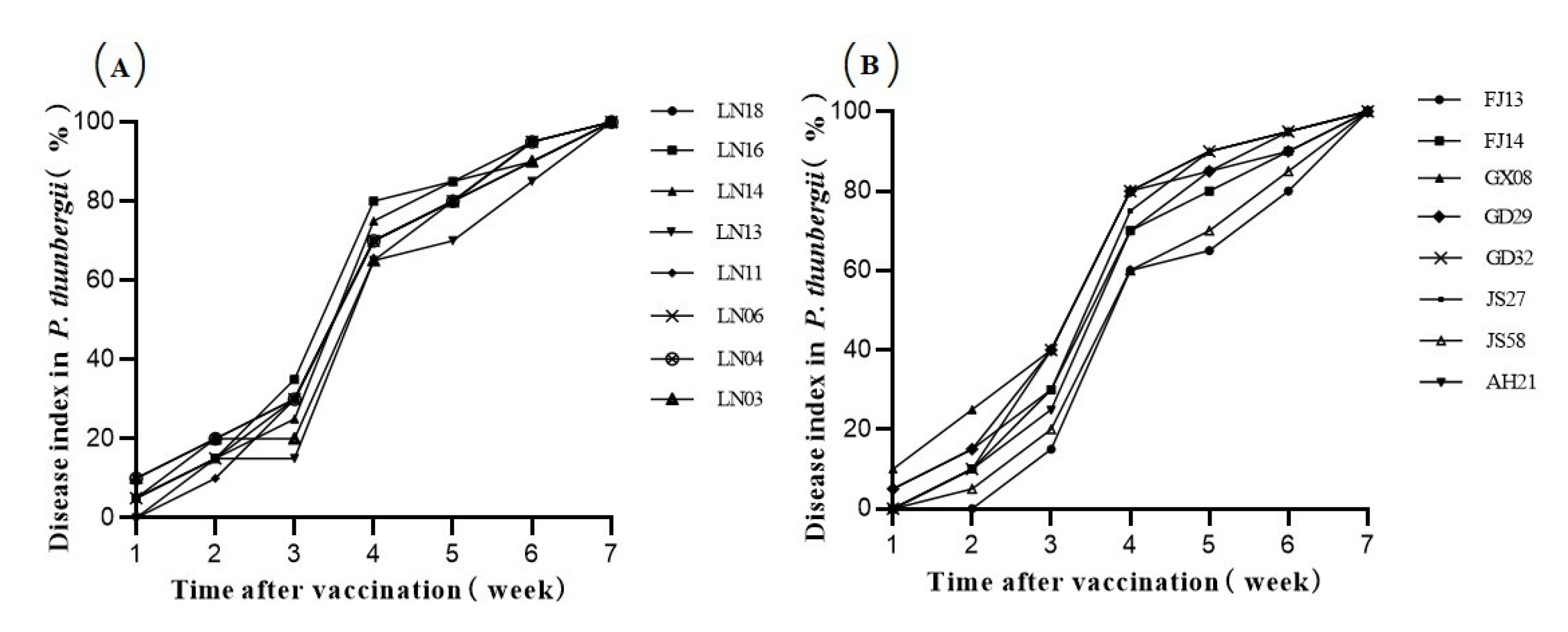
The 16 PWN strains from northern and southern China were inoculated into potted 2-year-old P. thunbergii seedlings. One week after inoculation, some of the pine seedlings showed needle chlorosis. At 3–4 weeks after inoculation, most of the pine seedlings reached the middle stage of disease progression; approximately 1/2 of the needles of the diseased plants exhibited chlorosis and browning, but the needles did not drop. Regarding the symptoms of disease in P. thunbergii after inoculation, the pine needles near the inoculation site first turned light green and yellow; these signs of disease in the pine needles progressed upward from the inoculation point; the pine needles near the inoculation point gradually turned from yellow to reddish brown; and this color change progressed upward until the entire plant was affected. At this point, the pine tree wilted and died, and the pine needles dropped. Thirty-two days after inoculation, one pine seedling had died in each of the treatment groups with LN18, LN16, LN14, LN11, LN04, FJ14, GX08, GD29, GD32 or AH21. In the 6th week after inoculation, the lethality rates of all strains except for LN13, FJ13, and JS58 were 60% or higher (Table 5). By the 7th week, all the pine seedlings had wilted and died (Figure 3). The 16 strains all showed strong pathogenicity to P. thunbergii, and the disease progression exhibited slow progress in the early stage, rapid expansion in the middle stage, and slow progression in the late stage. In the experiments, the difference in pathogenicity among the northern strains on P. thunbergii was very small, the difference in pathogenicity among the southern strains on P. thunbergii was greater than that of the northern strains, and GD32 had the strongest pathogenicity (Figure 4).
3.3. Comparison of the Pathogenicity of PWN Strains from Southern and Northern China to P. tabuliformis
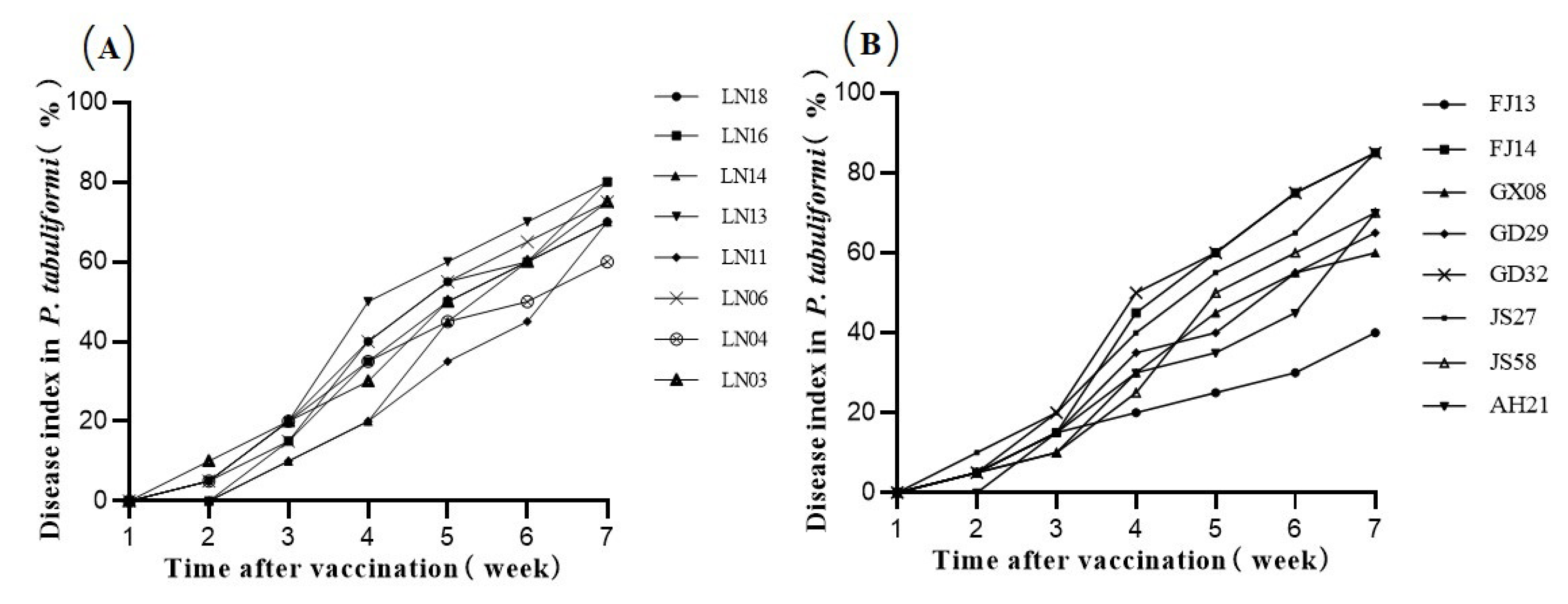
The 16 PWN strains were inoculated into 2-year-old P. tabuliformis potted seedlings. After inoculation, symptoms of pine disease appeared: the bases of the needles throughout the pine seedlings began to turn yellow, and the color change gradually spread to the needle tips. Then, the color of the needles deepened to reddish brown as the disease progressed. The P. tabuliformis seedlings wilted and died, but the needles rarely dropped when the seedlings died. The first onset of the disease in the pine seedlings typically occurred around the second week, and the onset time in the seedlings inoculated with the southern strain was generally earlier than that in seedlings inoculated with the northern strain. In the 4th to 5th weeks after inoculation, the inoculated pine seedlings entered the middle stage of disease progression. In the 10th week, the mortality rate of P. tabuliformis inoculated with GD32 was 100%, while FJ13 was only 0% (Table 6). After 38 days, three pine seedlings, one of each inoculated with LN03, LN04, or FJ14, had died. At the 10th week after inoculation, the average mortality rate of the pine seedlings inoculated with northern strains was 62.5% ± 26.1, and the average mortality rate of the pine seedlings inoculated with southern strains was 55% ± 33.4 (Table 7). By the 11th week, almost all the pine seedlings had wilted and died except GD29 (Figure 5). The differences in pathogenicity among the southern strains were greater than those among the northern strains, and the pathogenicity of LN13, LN06, and GD32 was significantly higher than that of FJ13 (Figure 6).
4. Discussion
According to previous studies, differences in pathogenicity among PWN strains may be related to the host [23,27], geographic location [21], environment [28,29], associated bacterial species [30], and strain fecundity [31,32]. This study showed that the same strain exhibits different levels of pathogenicity to different host pine trees. For instance, AH21 was more pathogenic to P. thunbergii than JS58, but its pathogenicity to P. tabuliformis was not as strong as that of JS58. This result suggested that PWN showed host specialization. The pathogenicity of strains isolated from different hosts to different pine also varied. Strains isolated from the same host showed similar pathogenicity. For instance, the strains LN18, LN16, and LN14 isolated from L. gmelinii exhibited similar virulence and showed strong virulence against P. thunbergii and P. tabuliformis. Siliang (2013) [23] pointed out that the strains with the same host source and inoculated target were more pathogenic. However, this correlation was not obvious in our study, and the pathogenicity of the strain still depended on the strain itself.
The pathogenicity of strains isolated from the same host source but different geographical populations may be different. The results of this experiment showed that the pathogenicity of the nematodes isolated from P. massoniana was highly differentiated. This is because P. massoniana itself has a large distribution area in southern China, and its disease resistance in different provenances varies [25,27]. Peigen et al. (1995) [33] used pine wood nematode strains from China and Japan to inoculate P. massoniana, P. thunbergii, Cedrus deodara, P. elliottii and P. taeda, respectively, and the study showed that there were certain differences in the pathogenicity of different hosts in different regions. In addition, the two strains from China and Japan, respectively, also differed in host specificity. The strains from China had strong pathogenicity to P. thunbergii and basically no pathogenicity to C. deodara, while strains from Japan had strong pathogenicity to C. deodara and P. elliottii. Generally, the PWN isolated from P. thunbergii had weaker pathogenicity than that isolated from P. massoniana. However, our results showed that the pathogenicity of strains isolated on P. thunbergii is not always stronger than that of P. massoniana. This illustrated that the geographical origin factor is generally, but not always, stronger than the host source factor. Nevertheless, the pathogenicity differences between strains with different geographical origins did not show absolute regularity. Zhiyu et al. (2002) [28] measured the pathogenicity of pine wood nematode populations in China, Japan, and Canada to 3–4-year-old P. thunbergii, and found that their pathogenicity was significantly differentiated. The study also showed there were both strong and weakly pathogenic groups in Nanjing. Thus, different strains of PWN had different pathogenicity levels, but the results had little correlation with geographical factors.
A comparison of the morphology and pathogenicity of two PWN strains in China by Ruocheng et al. (2019) [34] showed that the female body length of the northern strain FCBX was longer than that of strain AMA3 in the south. The results of the inoculation experiment on P. thunbergii showed that the strain FCBX is more pathogenic than the strain AMA3. However, it was not sufficient to only study these two individual strains, and P. thunbergii was considered to be the most susceptible pine species for PWN. The inoculation experiment on P. thunbergi cannot adequately explain the pathogenicity relationship between these two strains. Based on the experimental results, the female body length of GD32 was longer than all northern strains except LN04, and pathogenicity among the eight strains of Liaoning Province was similar and maintained at a high level, while the pathogenicity difference between the eight southern strains was greater. This pathogenic differentiation was more remarkable in the P. tabuliformis inoculation experiment. There were not only highly pathogenic GD32 and JS27, but also moderately pathogenic GD29, JS58, GX08, FJ14, AH21 and weakly pathogenic FJ13 among the eight southern strains. Thus, there was no absolute northern strain whose pathogenicity is stronger than southern strains. Whether in the P. thunbergii inoculation experiment or the P. tabuliformis inoculation experiment, the southern strain GD32 was more pathogenic than all eight northern strains. From this, we can speculate that if the southern strain GD32 is introduced and established in the northern regions, it would quickly spread and cause damage in the north.
5. Conclusions
This study showed that the females of the northern strains were longer and wider than those of the southern strains, but there were no significant differences in the morphological indicators of the males. The southern strains all showed strong pathogenicity to P. thunbergii, but their pathogenicity to P. tabuliformis was weaker. There was a certain degree of virulence differentiation between southern strains, but the pathogenicity of all northern strains was relatively strong. The results of this study have an important reference value and may support the evaluation of whether pine wood nematode disease will continue to cause major damage and further spread to the north of Liaoning Province in China.
Author Contributions
Q.-Q.K.: designed the study, conducted the experiment, performed the data analysis and wrote the article; X.-L.D.: guided the article writing and data analysis; Y.-F.C.: collected the samples and performed data analysis; J.-R.Y.: guarantor of the integrity of the entire study and approved the final version of the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the National Key Research and Development Program of China (No. 2018YFD0600203) and the National Natural Science Foundation of China 31800543 (X.D.).
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Xinyao, Z.; Youqing, L. Major Forest Diseases and Insect Pests in China; China Forestry Press: Hong Kong, China, 2003. [Google Scholar]
- Yano, S. Investigation on pine death in Nagasaki prefecture. Sanrin Kouhou 1913, 4, 1–4. (In Japanese) [Google Scholar]
- Ryoji, S.; Hironobu, M.; Yuko, T.; Kazuyoshi, F.; Mitsuyoshi, U. Making headway in understanding pine wilt disease: What do we perceive in the postgenomic era? J. Biosci. Bioeng. 2013, 116, 1–8. [Google Scholar]
- Mamiya, Y.; Kiyohara, T. Description of Bursaphelenchus Lignicolus N. Sp. (Nematoda: Aphelenchoididae) From Pine Wood and Histopathology of Nematode-Infested Trees. Nematologica 1972, 18, 120–124. [Google Scholar] [CrossRef]
- Cheng, H. The occurrence of a pine wilting disease caused by a nematode found in Nanjing. For. Pest Dis. 1983, 4, 1–5. [Google Scholar]
- Yi, C.; Byun, B.H.; Park, J.D.; Yang, S.I.; Chang, K.H. First finding of the pine wood nematode, Bursaphelenchus xylophilus (Steiner & Buhrer) Nickle and its insect vector in Korea. Res. Rep. For. Res. Inst. 1989, 38, 141–149. [Google Scholar]
- Mota, M.M.; Braasch, H.; Bravo, M.A.; Penas, A.C.; Burgermeister, W.; Metge, K.; Sousa, E. First report of Bursaphelenchus xylophilus in Portugal and in Europe. Nematology 1999, 1, 727–734. [Google Scholar] [CrossRef]
- Yu, H.; Wu, H.; Zhang, X.; Wang, L.; Zhang, X.; Song, Y. Preliminary study on Larix spp. infected by Bursaphelenchus xylophilus in natural environment. For. Pest Dis. 2019, 38, 7–10. [Google Scholar]
- Yu, H.; Wu, H.; Huang, R.; Wang, J.; Zhang, R.; Song, Y. Separation and identification of Bursaphelenchus xylophilus from Pinus sylvestris var. mongolica in FuShun city. For. Pest Dis. 2020, 39, 6–10. [Google Scholar]
- Futai, K.; Kanzaki, N. A PCR primer set for determination of phylogenetic relationships of Bursaphelenchus species within the xylophilus group. Nematology 2002, 4, 35–41. [Google Scholar]
- Yongxia, L.; Xingyao, Z. High Risk of Invasion and Expansion of Bursaphelenchus xylophilus in Middle Temperate Zone of China. For. Labor Saf. 2018, 1, 3–6. [Google Scholar]
- Xiaodong, L.; Yue, W.; Yantao, Z.; Hong, S. The occurrence of major forestry pests nationwide in 2019 and the forecast of their occurrence in 2020. For. Pest Dis. 2020, 39, 44–48. [Google Scholar]
- Baojun, Y.; Hongyang, P.; Jian, T. Pine Wilt Disease; China Forestry Press: Hong Kong, China, 2003. [Google Scholar]
- Zhengmei, Y.; Jiajin, T. Species and geographical distribution of parasitic nematode in pine in China. For. Pest Dis. 2019, 38, 38–42. [Google Scholar]
- Wang, X.Z.; Wang, L.F.; Wang, Y.; Huang, Y.Q.; Ding, Z.G.; Zhou, J.; Gou, D.P. Identification and genetic analysis of the pine wood nematode Bursaphelenchus xy-lophilus from Pinus yunnanensis. For. Pathol. 2015, 45, 388–399. [Google Scholar] [CrossRef]
- Juan, L.; Hanwen, Y. Current status and management countermeasures of Bursaphelenchus xylophilus disease areas in my country. For. Pest Dis. 2019, 38, 45–46. [Google Scholar]
- Jiajin, T.; Jianren, Y. List of nematodes in pine in China. For. Pest Dis. 2009, 28, 24–26. [Google Scholar]
- Xiupeng, L.; Yijiao, W. Pinus strobus var. chiapensis and Other Exotic Pines:Investigation and Analysis on Natural Nematode Infections and Death. J. Zhejiang A&F Univ. 1997, 3, 63–66. [Google Scholar]
- Lu, Z. Current Situation and Countermeasures of the Ecological Public Welfare Forest Resources of Pinus tabuliformis in Northwestern Liaoning. LiaoNing For. Sci. Technol. 2015, 1, 36–38. [Google Scholar]
- Ping, L.; Jincheng, H.; Lei, Y.; Xin, J.; Yutao, W. Biomass of Pinus tabuliformis Plantation in Mountainous Area of Eastern Liaoning. J. Shenyang Agric. Univ. 2019, 50, 740–746. [Google Scholar]
- Jiajin, T.; Rongzhen, Y.; Huiping, W. Differences in pathogenicity of Bursaphelenchus xylophilus from different geographic populations to Masson pine. Plant Quar. 2000, 6, 324–325. [Google Scholar]
- Lin, H.; Jianren, Y.; Xiaoqin, W. Nematode families establishment and pathogenicity evaluation of Bursaphelenchus xylophilus and B. mucronatus Strains. J. For. Eng. 2011, 25, 40–42. [Google Scholar]
- Siliang, W. Bursaphelenchus xylophilus from China Naturally Infected Host Populations Pathogenic to Pine Masson Study. Ph.D. Thesis, Northeast Forestry University, Harbin, China, 2013. [Google Scholar]
- Xuelian, L. Analysis of the morphologic and pathogenic variation of Bursaphelenchus xylophilus in China Abstract. Ph.D. Thesis, Nanjing Forestry University, 2007. Ph.D. Thesis, Nanjing Forestry University, Nanjing, China, 2007. [Google Scholar]
- Fuyuan, X.; Minghong, G.; Qiming, W.; Pei, Z.; Kegong, Z.; Julin, Z.; Xu, D.; Minmin, W. Study on the Masson pine provenances resistance to pine wood nematode (PWN) disease in China. J. Nanjing For. Univ. 1998, 2, 31–35. [Google Scholar]
- Que, X.; Wu, X.Q.; Wu, F.; Ye, J.R. Transcriptome Analysis of Bursaphelenchus xylophilus Uncovers the Impact of Steno-trophomonas maltophilia on Nematode and Pine Wilt Disease. Forests 2020, 11, 908. [Google Scholar] [CrossRef]
- Liu, Q.H.; Zhou, Z.C.; Xu, L.Y.; Chen, X.L.; Ning, L.U.O. Identification of Candidate Constitutive Expressed Resistant Genes of Pine Wilt Disease in Pinus massoniana Based on High-throughput Transcriptome Sequencing. For. Res. 2019, 32, 1–10. [Google Scholar]
- Zhiyu, Z.; Keyun, Z.; Maosong, L.; Hongwei, L.; Fuyuan, X. Pathogenicity determination of Bursaphelenchus xylophilus isolates to Pine thunbergii. J. Nanjing Agric. Univ. 2002, 25, 43–46. [Google Scholar]
- Kiyohara, T.; Bolla, R.I. Pathogenic variability among populations of the pinewood nematode, Bursaphelenchus Xylophilus. For. Sci. 1990, 36, 1061–1076. [Google Scholar]
- Yingdi, H.; Boguang, Z.; Yue, C.; Zhengmin, H. Pathogenicity of Bacteria Carried by Pine Wood Nematodes. J. Nanjing Agric. Univ. 2003, 27, 45–48. [Google Scholar]
- Kikuchi, T.; Aikawa, T. Estimation of virulence of Bursaphelenchus xylophilus (Nematoda: Aphelenchoididae) based on its reproductive ability. Nematology 2007, 9, 371–377. [Google Scholar] [CrossRef]
- Joung, A.S.; Yil-Sung, M. Migrations and multiplications of Bursaphelenchus xylophilus and B. mucronatus in Pinus thumbergii in relation to their pathogenicity. Agric. Biol. Chem. 2013, 29, 116–122. [Google Scholar]
- Peigen, S.; Guoyao, J.; Hongmei, L. Difference in pathogenicity of pine wood nematode between Nanjing and Japan. For. Pest Dis. 1995, 4, 1–2. [Google Scholar]
- Ruocheng, S.; Min, L.; Jun, C.; Zhijian, G.; Shouhui, S.; Jianren, Y.; Fengmao, C. Comparison of morphological index and pathogenicity of two isolates of Bursaphelenchus xylophilus in southern and northern in China. J. Nanjing For. Univ. 2019, 43, 18–24. [Google Scholar]
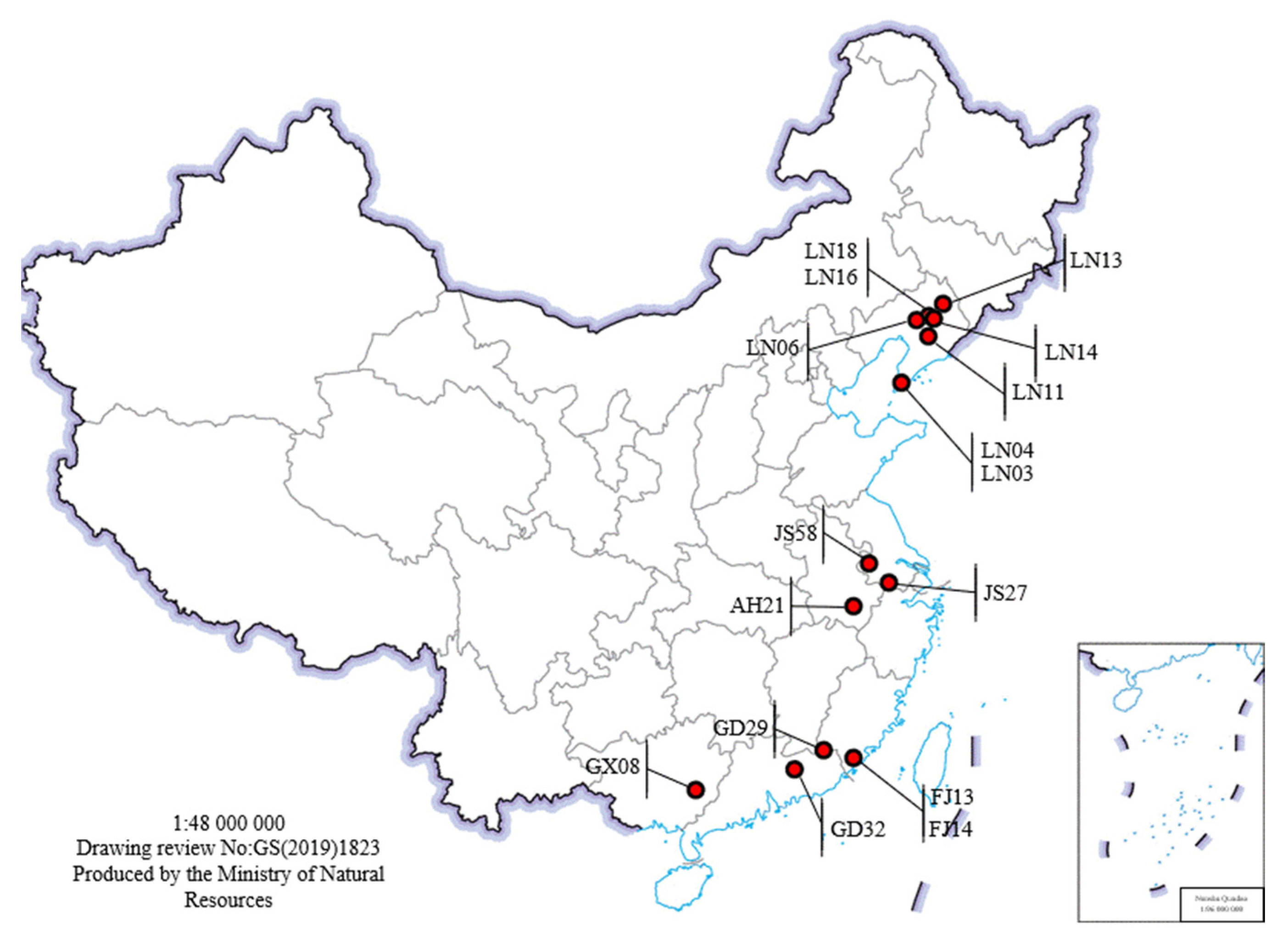
Figure 1.
Sampling locations for 16 strains. The red dot on the map represents the sampling location of the corresponding strain. The sampling locations of LN16 and LN18, LN03 and LN04, FJ13 and FJ14 are very close. All strains were isolated from different pine trees and different forest stands.
Figure 1.
Sampling locations for 16 strains. The red dot on the map represents the sampling location of the corresponding strain. The sampling locations of LN16 and LN18, LN03 and LN04, FJ13 and FJ14 are very close. All strains were isolated from different pine trees and different forest stands.

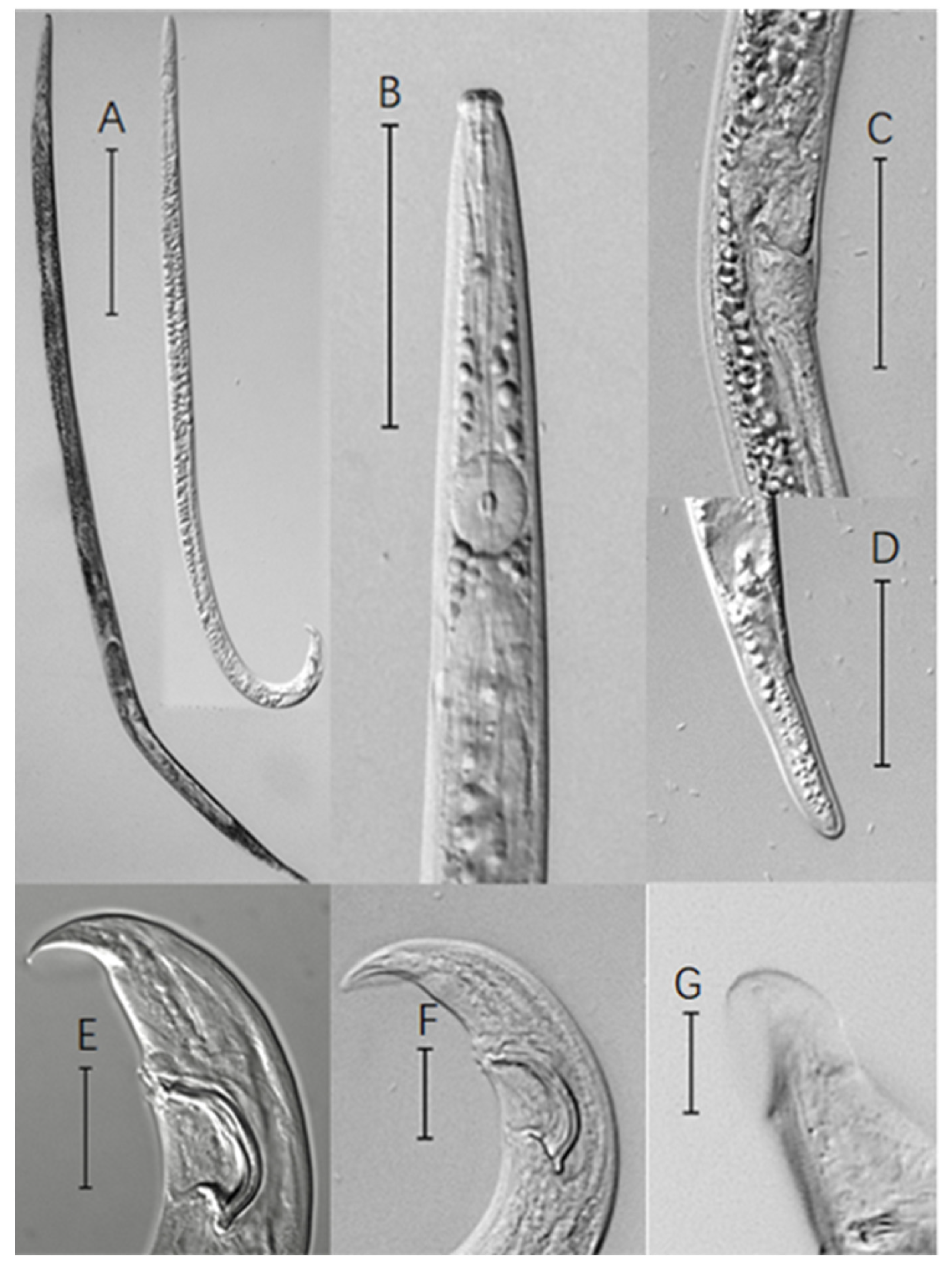
Figure 2.
Photomicrographs of pine wood nematode (PWN). (A): Entire view of female (left) and male (right); (B): Anterior region of female; (C): Vulva region; (D): Female tail; (E,F): Ventral view of male tail; (G): Bursa of the adult male tail. (Scale bars: (A) = 200 μm; (B–D) = 50 μm; (E–G) = 20 μm).
Figure 2.
Photomicrographs of pine wood nematode (PWN). (A): Entire view of female (left) and male (right); (B): Anterior region of female; (C): Vulva region; (D): Female tail; (E,F): Ventral view of male tail; (G): Bursa of the adult male tail. (Scale bars: (A) = 200 μm; (B–D) = 50 μm; (E–G) = 20 μm).

Figure 3.
Symptoms of P. thunbergii after artificial inoculation with 16 strains of PWN.

Figure 4.
The pine wood nematode (PWN) Bursaphelenchus xylophilus disease index in P. thunbergii after inoculation with PWN strains. Sixteen reference numbers represent sixteen PWN isolates from different regions. (A) Disease index of the 8 northern strains. (B) Disease index of the 8 southern strains.
Figure 4.
The pine wood nematode (PWN) Bursaphelenchus xylophilus disease index in P. thunbergii after inoculation with PWN strains. Sixteen reference numbers represent sixteen PWN isolates from different regions. (A) Disease index of the 8 northern strains. (B) Disease index of the 8 southern strains.

Figure 5.
Symptoms of P. tabuliformis after artificial inoculation with 16 strains of PWN.

Figure 6.
The pine wood nematode (PWN) Bursaphelenchus xylophilus disease index in P. tabuliformis after inoculation with PWN strains. Sixteen reference numbers represent sixteen PWN isolates from different regions. (A) Disease index of the 8 northern strains. (B) Disease index of the 8 southern strains.
Figure 6.
The pine wood nematode (PWN) Bursaphelenchus xylophilus disease index in P. tabuliformis after inoculation with PWN strains. Sixteen reference numbers represent sixteen PWN isolates from different regions. (A) Disease index of the 8 northern strains. (B) Disease index of the 8 southern strains.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Species, height and ground diameter of pine seedlings.
| Pine Seedling Species | Height/cm | Ground Diameter/mm |
|---|---|---|
| P. thunbergia | 50 | 7.5 |
| P. tabuliformis | 40 | 9.0 |
Table 2.
Sources and codes of the pine wood nematode (PWN) isolates used in the study.
| No. | Code | Strain Source | Collection Time (YYYY/MM/DD) |
|---|---|---|---|
| 1 | LN18 | Fushun city, Liaoning Province (Larix gmelinii) | 2018/9/25 |
| 2 | LN16 | Fushun city, Liaoning Province (L. gmelinii) | 2018/9/25 |
| 3 | LN14 | Fushun city, Liaoning Province (L. gmelinii) | 2018/9/25 |
| 4 | LN13 | Dandong city, Liaoning Province (P. tabuliformis) | 2017/11/30 |
| 5 | LN11 | Benxi city, Liaoning Province (P. massoniana) | 2017/8/16 |
| 6 | LN06 | Shenyang city, Liaoning Province (P. tabuliformis) | 2017/8/20 |
| 7 | LN04 | Dalian city, Liaoning Province (P. thunbergii) | 2016/10/20 |
| 8 | LN03 | Dalian city, Liaoning Province (P. thunbergii) | 2016/10/20 |
| 9 | FJ13 | Zhangzhou city, Fujian Province (P. massoniana) | 2019/4/6 |
| 10 | FJ14 | Zhangzhou city, Fujian Province (P. massoniana) | 2019/4/6 |
| 11 | GX08 | Guigang city, Guangxi Province (P. massoniana) | 2015/1/19 |
| 12 | GD29 | Meizhou city, Guangdong Province (P. massoniana) | 2017/8/6 |
| 13 | GD32 | Heyuan city, Guangdong Province (P. massoniana) | 2017/8/8 |
| 14 | JS27 | Nanjing, Jiangsu Province (P. massoniana) | 2017/10/24 |
| 15 | JS58 | Wuxi city, Jiangsu Province (P. massoniana) | 2017/11/1 |
| 16 | AH21 | Huangshan city, Anhui Province (P. massoniana) | 2018/10/31 |
Table 3.
Morphometrics of female nematodes in north and south China. L is the body length (μm); LW is the maximum body width (μm); a is the body length/maximum body width ratio; b is the body length/tail length ratio; V is the distance from the vulva to the top of the head × 100/Body length; WMB is the width of the middle esophageal bulb (μm); Stylet is the length of the stylus (μm); Tail is the length of the tail (μm).
Table 3.
Morphometrics of female nematodes in north and south China. L is the body length (μm); LW is the maximum body width (μm); a is the body length/maximum body width ratio; b is the body length/tail length ratio; V is the distance from the vulva to the top of the head × 100/Body length; WMB is the width of the middle esophageal bulb (μm); Stylet is the length of the stylus (μm); Tail is the length of the tail (μm).
| No. | Code | n | L (μm) | LW (μm) | a | b | V | WMB (μm) | Stylet (μm) | Tail (μm) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LN18 | 30 | 1005.1 ± 67.0 c | 30.5 ± 3.3 b | 34.3 ± 2.1 b | 29.3 ± 2.2 a | 75.2 ± 0.7 a | 15.0 ± 1.0 a | 14.9 ± 0.6 b | 40.3 ± 1.2 a |
| (907.4–1190.8) | (27.5–37.1) | (30.6–39.3) | (26.0–34.6) | (73.7–77.9) | (14.2–16.0) | (13.1–15.9) | (38.0–44.2) | |||
| 2 | LN16 | 30 | 1076 ± 152.4 b | 30.1 ± 3.7 c | 37.5 ± 5.4 a | 31.8 ± 3.7 a | 74.8 ± 1.5 a | 14.7 ± 1.8 a | 15.2 ± 0.6 a | 35.2 ± 3.4 a |
| (846.5–1399.9) | (28.2–38.3) | (27.0–51.8) | (23.7–40.5) | (72.1–79.1) | (12.1–20.5) | (14.3–15.9) | (33.9–37.5) | |||
| 3 | LN14 | 30 | 1056 ± 102.8 b | 30.5 ± 2.7 b | 35.6 ± 4.0 b | 31.5 ± 2.5 a | 74.8 ± 1.4 a | 14.5 ± 1.1 a | 14.8 ± 0.7 b | 36.2 ± 2.5 a |
| (867.7–1302.4) | (27.2–38.3) | (27.8–41.8) | (25.8–41.6) | (72.1–79.1) | (13.1–21.0) | (14.2–15.5) | (35.5–40.3) | |||
| 4 | LN13 | 30 | 1012.4 ± 68.7 c | 29.6 ± 1.0 d | 39.2 ± 1.1 a | 31.0 ± 0.6 a | 74.1 ± 0.8 a | 14.8 ± 0.2 a | 14.8 ± 0.2 b | 36.2 ± 0.7 a |
| (875.5–1239.0) | (28.0–35.1) | (33.4–43.7) | (28.0–34.1) | (73.5–76.1) | (13.9–16.5) | (14.1–16.2) | (35.2–39.3) | |||
| 5 | LN11 | 30 | 1047.6 ± 87.5 b | 29.4 ± 0.6 d | 36.7 ± 1.3 a | 29.9 ± 1.0 a | 73.7 ± 1.2 a | 14.6 ± 0.2 a | 14.8 ± 0.3 b | 38.0 ± 0.7 a |
| (842.8–1192.6) | (27.3–32.1) | (28.9–43.0) | (25.9–37.3) | (73.4–75.7) | (13.9–16.6) | (13.8–17.2) | (35.5–39.2) | |||
| 6 | LN06 | 30 | 1055.1 ± 73.3 b | 30.2 ± 1.0 c | 34.3 ± 1.2 b | 30.6 ± 1.1 a | 74.6 ± 0.3 a | 14.5 ± 0.2 a | 14.7 ± 0.4 bc | 39.3 ± 0.6 a |
| (863.5–1192.0) | (26.7–34.6) | (29.3–41.5) | (26.5–37.3) | (72.6–76.6) | (13.8–15.8) | (13.9–15.2) | (37.6–40.5) | |||
| 7 | LN04 | 30 | 1123.4 ± 123.8 a | 30.8 ± 1.5 a | 36.4 ± 3.2 a | 31.3 ± 3.2 a | 74.8 ± 1.5 a | 14.6 ± 0.9 a | 13.7 ± 0.7 d | 36.2 ± 1.4 a |
| (987.6–1309.2) | (29.2–31.3) | (30.7–40.6) | (30.0–41.2) | (73.1–79.4) | (12.1–17.5) | (12.0–14.9) | (34.9–37.8) | |||
| 8 | LN03 | 30 | 1080.2 ± 102.4 b | 29.1 ± 4.7 f | 37.5 ± 3.0 a | 32.1 ± 2.2 a | 73.6 ± 1.0 a | 14.3 ± 0.8 a | 13.7 ± 0.7 e | 36.3 ± 0.8 a |
| (846.5–1299.9) | (26.2–38.3) | (31.4–43.8) | (24.7–40.5) | (72.5–74.5) | (12.8–16.2) | (12.0–14.9) | (34.7–38.3) | |||
| 9 | FJ13 | 30 | 918.8 ± 29.0 f | 29.5 ± 0.7 d | 32.3 ± 0.9 b | 29.9 ± 0.8 a | 74.6 ± 0.3 a | 14.6 ± 0.4 a | 14.8 ± 0.2 b | 30.7 ± 0.8 b |
| (710.8–990.8) | (26.1–34.5) | (26.6–39.3) | (29.9–43.1) | (73.1–76.2) | (12.5–16.4) | (13.9–16.5) | (28.5–33.0) | |||
| 10 | FJ14 | 30 | 913.0 ± 58.5 f | 29.1 ± 3.9 f | 36.1 ± 2.4 a | 25.6 ± 2.3 b | 73.8 ± 0.8 a | 14.3 ± 1.2 a | 14.1 ± 0.5 d | 37.2 ± 1.1 a |
| (839.5–1020.5) | (23.9–35.7) | (33.4–40.7) | (22.4–31.8) | (72.3–76.4) | (12.9–15.4) | (11.2–13.8) | (35.2–38.8) | |||
| 11 | GX08 | 30 | 906.0 ± 69.7 f | 29.4 ± 3.0 d | 37.2 ± 3.2 a | 24.2 ± 2.8 b | 74.8 ± 1.3 a | 14.4 ± 1.5 a | 15.2 ± 1.0 a | 34.2 ± 3.1 a |
| (862.3–1004.2) | (25.2–36.3) | (33.5–41.2) | (23.8–30.8) | (72.1–75.1) | (13.1–16.5) | (13.4–14.9) | (31.3–39.2) | |||
| 12 | GD29 | 30 | 898.3 ± 78.3 g | 27.4 ± 5.5 h | 34.2 ± 2.1 b | 26.7 ± 3.2 a | 75.8 ± 2.0 a | 14.9 ± 1.4 a | 13.7 ± 0.6 e | 34.8 ± 1.1 a |
| (718.4–978.7) | (23.9–35.7) | (33.4–40.7) | (24.4–31.8) | (72.3–76.4) | (12.9–15.4) | (12.2–13.8) | (32.3–37.8) | |||
| 13 | GD32 | 30 | 1120.6 ± 73.4 b | 30.8 ± 1.5 a | 37.5 ± 3.0 a | 31.8 ± 3.2 a | 74.8 ± 1.5 a | 14.7 ± 1.5 a | 14.1 ± 0.7 d | 36.2 ± 1.0 a |
| (1037.5–1176.8) | (29.2–31.3) | (31.4–43.8) | (24.7–40.5) | (72.1–79.1) | (12.1–17.5) | (13.0–14.9) | (34.9–37.8) | |||
| 14 | JS27 | 30 | 924.0 ± 68.5 e | 29.1 ± 3.9 f | 36.1 ± 2.4 a | 26.6 ± 2.3 a | 73.8 ± 0.8 a | 14.3 ± 1.2 a | 14.1 ± 0.5 d | 37.2 ± 1.1 a |
| (839.5–1010.5) | (25.2–36.3) | (33.5–41.2) | (24.8–30.8) | (72.1–75.1) | (13.1–16.5) | (13.4–14.9) | (36.3–40.2) | |||
| 15 | JS58 | 30 | 964.5 ± 113.2 d | 28.2 ± 3.7 g | 34.3 ± 2.9 b | 30.2 ± 4.2 a | 75.4 ± 4.5 a | 14.5 ± 0.7 a | 14.0 ± 1.0 d | 32.2 ± 1.8 b |
| (730.0–1071.5) | (22.6–31.7) | (31.5–39.9) | (22.6–35.2) | (71.1–87.1) | (12.1–13.0) | (13.4–14.9) | (28.8–35.0) | |||
| 16 | AH21 | 30 | 955.1 ± 33.3 e | 28.5 ± 4.0 g | 34.3 ± 1.2 b | 28.9 ± 1.0 a | 74.8 ± 0.3 a | 14.7 ± 1.2 a | 14.7 ± 0.1 bc | 31.3 ± 0.6 b |
| (763.5–1112.0) | (25.6–37.2) | (29.3–41.5) | (24.0–34.5) | (73.5–76.4) | (13.5~16.2) | (14.3–15.0) | (29.6–35.5) |
Note: The measured value in the form (μm) = mean ± standard error (minimum-maximum) Values (μm) in the form = mean ± SD (range). Different letters after the same column of values indicate significant differences (Duncan multiple comparisons, significance level p < 0.05), One way ANOVA analysis results: F (7, 120) = 2434, p < 0.0001.
Table 4.
Morphometrics of male nematodes in north and south China. L is the body length (μm); LW is the maximum body width (μm); a is the body length/maximum body width ratio; b is the body length/tail length ratio; WMB is the width of the middle esophageal bulb (μm); Stylet is the length of the stylus (μm); Spi is the length of the spicule (μm); Tail is the length of the tail (μm).
Table 4.
Morphometrics of male nematodes in north and south China. L is the body length (μm); LW is the maximum body width (μm); a is the body length/maximum body width ratio; b is the body length/tail length ratio; WMB is the width of the middle esophageal bulb (μm); Stylet is the length of the stylus (μm); Spi is the length of the spicule (μm); Tail is the length of the tail (μm).
| No. | Code | n | L (μm) | LW (μm) | a | b | WMB (μm) | Stylet (μm) | Tail (μm) | Spi (μm) |
|---|---|---|---|---|---|---|---|---|---|---|
| 1 | LN18 | 30 | 975.1 ± 56.1 b | 29.4 ± 3.4 a | 34.3 ± 1.2 b | 27.9 ± 0.6 a | 14.8 ± 1.0 a | 14.4 ± 0.6 a | 36.3 ± 1.3 a | 29.3 ± 0.4 a |
| (879.3–1001.2) | (25.4–33.2) | (29.3–41.5) | (25.1–31.3) | (14.1–15.0) | (13.6–15.0) | (34.3–39.5) | (26.3–30.3) | |||
| 2 | LN16 | 30 | 953.0 ± 96.9 b | 28.9 ± 3.3 b | 39.2 ± 1.1 a | 27.7 ± 0.7 a | 14.3 ± 1.6 a | 14.0 ± 0.5 a | 32.2 ± 2.6 a | 28.7 ± 0.5 a |
| (808.6–1103.3) | (26.2–32.7) | (33.4–43.7) | (23.5–31.1) | (13.2–16.5) | (13.5–14.9) | (30.4–36.5) | (26.6–30.0) | |||
| 3 | LN14 | 30 | 937 ± 100.3 b | 29.8 ± 3.0 a | 36.7 ± 1.3 a | 28.1 ± 0.6 a | 14.0 ± 1.6 b | 14.1 ± 0.6 a | 34.2 ± 2.3 a | 31.2 ± 0.5 a |
| (873.1–1012.2) | (27.2–36.2) | (28.9–43.0) | (25.3–30.5) | (13.4–17.0) | (13.8–14.8) | (32.8–39.5) | (28.0–32.2) | |||
| 4 | LN13 | 30 | 904.5 ± 88.8 c | 29.0 ± 1.1 b | 35.1 ± 0.9 b | 28.3 ± 0.7 a | 14.4 ± 0.4 a | 14.2 ± 0.4 a | 34.7 ± 0.8 a | 29.1 ± 0.6 a |
| (875.5–1239.0) | (27.2–34.7) | (30.3–40.9) | (23.6–31.3) | (13.9–15.8) | (13.8–15.2) | (33.7–37.3) | (27.8–29.8) | |||
| 5 | LN11 | 30 | 963.6 ± 87.5 b | 28.2 ± 1.6 c | 32.7 ± 1.3 b | 25.5 ± 0.6 b | 14.1 ± 0.4 a | 14.3 ± 0.2 a | 34.8 ± 0.7 a | 30.4 ± 0.6 a |
| (892.8–1102.5) | (26.2–31.5) | (25.0–39.1) | (22.3–28.9) | (13.9–15.8) | (13.8–15.2) | (32.5–35.2) | (28.3–31.6) | |||
| 6 | LN06 | 30 | 978.4 ± 84.6 b | 29.1 ± 1.3 a | 34.0 ± 1.1 b | 27.3 ± 0.8 a | 14.0 ± 0.6 b | 14.2 ± 0.5 a | 35.6 ± 0.8 a | 29.3 ± 0.4 a |
| (823.3–1094.9) | (26.2–33.0) | (29.9–40.1) | (23.0–31.3) | (13.1–15.4) | (13.9–14.7) | (32.6–36.6) | (26.4–31.1) | |||
| 7 | LN04 | 30 | 1040.3 ± 93.2 a | 28.6 ± 2.5 b | 34.6 ± 0.9 b | 26.8 ± 0.8 a | 14.3 ± 1.0 a | 13.2 ± 0.6 b | 33.0 ± 1.1 a | 28.9 ± 0.5 a |
| (890.6–1124.9) | (26.8–30.3) | (30.7–40.4) | (20.8–32.2) | (12.3–14.9) | (12.6–14.1) | (30.9–34.8) | (27.0–29.6) | |||
| 8 | LN03 | 30 | 951.7 ± 113.7 b | 28.0 ± 3.8 d | 32.3 ± 0.9 b | 27.2 ± 0.4 a | 13.9 ± 1.0 b | 13.0 ± 0.7 b | 33.7 ± 1.0 a | 29.3 ± 0.3 a |
| (876.6–1109.9) | (26.2–38.3) | (27.3–37.0) | (24.7–30.0) | (12.6–15.7) | (12.0–13.3) | (31.8–36.2) | (27.1–30.2) | |||
| 9 | FJ13 | 30 | 901.2 ± 36.3 c | 29.0 ± 1.7 a | 34.5 ± 1.2 b | 29.2 ± 0.8 a | 13.8 ± 0.4 b | 14.0 ± 0.3 a | 28.4 ± 1.2 b | 28.9 ± 0.4 a |
| (699.3–972.1) | (25.7–34.3) | (29.0–41.8) | (23.3–32.8) | (13.0–14.3) | (13.9–14.5) | (26.5–30.0) | (26.2–29.8) | |||
| 10 | FJ14 | 30 | 908.2 ± 38.8 c | 28.6 ± 3.1 c | 35.0 ± 1.3 b | 26.4 ± 0.7 a | 14.0 ± 0.8 b | 13.4 ± 0.5 b | 33.2 ± 1.4 a | 30.2 ± 0.5 a |
| (812.7–988.8) | (23.5–35.4) | (28.6–40.1) | (22.7–30.7) | (13.1–14.4) | (11.8–13.8) | (31.5–36.0) | (28.5–31.6) | |||
| 11 | GX08 | 30 | 911.2 ± 72.5 c | 27.8 ± 3.3 d | 36.5 ± 1.1 a | 28.5 ± 0.5 a | 14.2 ± 1.0 a | 14.1 ± 0.6 a | 32.6 ± 2.6 a | 27.8 ± 0.6 a |
| (832.8–987.4) | (24.2–32.7) | (30.8–41.6) | (25.2–32.3) | (13.1–15.2) | (13.4–14.9) | (34.5–37.1) | (24.0–30.3) | |||
| 12 | GD29 | 30 | 821.3 ± 38.3 d | 28.7 ± 3.5 b | 36.4 ± 1.5 a | 25.5 ± 0.6 b | 14.5 ± 1.2 a | 13.4 ± 0.6 a | 32.1 ± 1.5 a | 28.3 ± 0.3 a |
| (700.4–889.2) | (26.9–34.5) | (31.3–48.3) | (22.4–29.4) | (12.9–15.4) | (12.5–13.8) | (30.1–34.6) | (26.6–29.7) | |||
| 13 | GD32 | 30 | 987.6 ± 66.6 b | 27.8 ± 2.6 d | 39.2 ± 0.7 a | 28.4 ± 0.4 a | 14.5 ± 1.5 a | 13.3 ± 0.8 b | 33.7 ± 1.7 a | 27.5 ± 0.5 a |
| (892.7–1054.3) | (26.3–31.3) | (35.6–43.3) | (25.2–30.5) | (12.4–16.0) | (12.7–13.9) | (31.4–36.2) | (24.7–29.7) | |||
| 14 | JS27 | 30 | 848.0 ± 62.8 d | 28.7 ± 3.7 b | 35.9 ± 1.2 a | 26.4 ± 0.6 a | 14.0 ± 1.0 b | 13.5 ± 0.6 b | 34.2 ± 0.8 a | 29.0 ± 0.7 a |
| (789.6–935.6) | (25.5–35.2) | (30.7–44.0) | (23.6–29.9) | (13.1–15.5) | (13.4–14.1) | (33.7–35.7) | (22.9–32.0) | |||
| 15 | JS58 | 30 | 922.5 ± 83.7 b | 28.3 ± 4.1 c | 39.0 ± 1.1 a | 27.5 ± 0.5 a | 14.1 ± 0.8 a | 13.2 ± 0.6 b | 30.2 ± 1.3 b | 28.6 ± 1.0 a |
| (803.2–989.4) | (24.6–33.2) | (31.5–44.2) | (24.9–30.4) | (13.1–14.8) | (12.4–13.9) | (27.8–31.8) | (24.6–35.5) | |||
| 16 | AH21 | 30 | 913.7 ± 43.3 c | 28.2 ± 4.1 c | 34.5 ± 1.1 b | 26.9 ± 0.5 a | 14.2 ± 1.2 a | 14.0 ± 0.8 a | 30.1 ± 1.0 b | 28.4 ± 0.6 a |
| (799.5–972.2) | (25.2–31.1) | (29.2–41.3) | (23.7–30.0) | (13.3~15.6) | (13.9–14.6) | (28.4–33.2) | (24.0–31.7) |
Note: The measured value in the form (μm) = mean ± standard error (minimum-maximum) Values (μm) in the form = mean ± SD (range). Different letters after the same column of values indicate significant differences (Duncan multiple comparisons, significance level p < 0.05), One way ANOVA analysis results: F (7, 120) = 4557, p < 0.0001.
Table 5.
The disease level and index and the mortality rate of P. thunbergii after artificial inoculation with different PWN strains.
Table 5.
The disease level and index and the mortality rate of P. thunbergii after artificial inoculation with different PWN strains.
| No. | Code | n | One Week after Inoculation | Three Weeks after Inoculation | Six Weeks after Inoculation | ||||
|---|---|---|---|---|---|---|---|---|---|
| Disease Level | Disease Index | Disease Level | Disease Index | Disease Level | Disease Index | Mortality Rate (%) | |||
| 1 | LN18 | 5 | 0.2 | 5 | 2.2 | 55 | 3.8 | 95 | 80 |
| 2 | LN16 | 5 | 0.2 | 5 | 2 | 50 | 3.8 | 95 | 80 |
| 3 | LN14 | 5 | 0.2 | 5 | 1.8 | 45 | 3.4 | 95 | 60 |
| 4 | LN13 | 5 | 0 | 0 | 1.4 | 35 | 3 | 80 | 20 |
| 5 | LN11 | 5 | 0.2 | 5 | 2 | 50 | 3.4 | 85 | 40 |
| 6 | LN06 | 5 | 0.2 | 5 | 2.2 | 55 | 3.8 | 95 | 80 |
| 7 | LN04 | 5 | 0.4 | 10 | 2 | 50 | 3.6 | 90 | 60 |
| 8 | LN03 | 5 | 0.2 | 5 | 1.6 | 40 | 3.6 | 90 | 60 |
| 9 | FJ13 | 5 | 0 | 0 | 1 | 25 | 3 | 80 | 20 |
| 10 | FJ14 | 5 | 0 | 0 | 2 | 50 | 3.6 | 90 | 60 |
| 11 | GX08 | 5 | 0.2 | 5 | 2.2 | 55 | 4 | 100 | 100 |
| 12 | GD29 | 5 | 0 | 0 | 1.8 | 45 | 3.4 | 85 | 40 |
| 13 | GD32 | 5 | 0.4 | 10 | 2.4 | 60 | 4 | 100 | 100 |
| 14 | JS27 | 5 | 0.2 | 5 | 1.8 | 45 | 3.8 | 95 | 80 |
| 15 | JS58 | 5 | 0 | 0 | 2 | 50 | 3.4 | 85 | 20 |
| 16 | AH21 | 5 | 0 | 0 | 1.6 | 40 | 3.8 | 95 | 60 |
Table 6.
The disease level and index and the mortality rate of P. tabuliformis after artificial inoculation with southern and northern PWN strains.
Table 6.
The disease level and index and the mortality rate of P. tabuliformis after artificial inoculation with southern and northern PWN strains.
| No. | Code | n | Two Weeks after Inoculation | Five Weeks after Inoculation | Ten Weeks after Inoculation | ||||
|---|---|---|---|---|---|---|---|---|---|
| Disease Level | Disease Index | Disease Level | Disease Index | Disease Level | Disease Index | Mortality Rate (%) | |||
| 1 | LN18 | 5 | 0.2 | 5 | 1.8 | 45 | 3.4 | 85 | 40 |
| 2 | LN16 | 5 | 0 | 0 | 2 | 50 | 3.4 | 85 | 40 |
| 3 | LN14 | 5 | 0 | 0 | 1.6 | 40 | 3.4 | 85 | 60 |
| 4 | LN13 | 5 | 0.4 | 10 | 2.4 | 60 | 3.8 | 95 | 80 |
| 5 | LN11 | 5 | 0.2 | 5 | 1.4 | 35 | 3.8 | 95 | 80 |
| 6 | LN06 | 5 | 0.6 | 15 | 2.2 | 55 | 3.6 | 90 | 80 |
| 7 | LN04 | 5 | 0.8 | 20 | 1.6 | 40 | 3.6 | 90 | 20 |
| 8 | LN03 | 5 | 0.6 | 15 | 1.8 | 45 | 2.8 | 70 | 20 |
| 9 | FJ13 | 5 | 0.4 | 10 | 0.8 | 20 | 2.2 | 55 | 0 |
| 10 | FJ14 | 5 | 0.2 | 5 | 2.4 | 60 | 3.6 | 90 | 60 |
| 11 | GX08 | 5 | 0.4 | 10 | 1.4 | 35 | 3.4 | 85 | 40 |
| 12 | GD29 | 5 | 0.6 | 15 | 1.4 | 35 | 3.2 | 80 | 20 |
| 13 | GD32 | 5 | 0.6 | 15 | 2.8 | 65 | 4 | 100 | 100 |
| 14 | JS27 | 5 | 0.8 | 20 | 3 | 70 | 3.8 | 95 | 80 |
| 15 | JS58 | 5 | 0.4 | 10 | 1.8 | 45 | 3.2 | 80 | 60 |
| 16 | AH21 | 5 | 0.2 | 5 | 1.4 | 35 | 3.5 | 85 | 80 |
Table 7.
Disease index and mortality of P. tabuliformis inoculated with northern and southern PWN strains.
Table 7.
Disease index and mortality of P. tabuliformis inoculated with northern and southern PWN strains.
| Inoculum | n | Disease Index | Mortality Rate % | ||
|---|---|---|---|---|---|
| 14 days | 35 days | 70 days | |||
| Northern strain | 8 | 8.8 ± 7.4 | 46.2 ± 8.3 | 86.9 ± 8.0 | 62.5 ± 26.1 |
| Southern strain | 8 | 11.2 ± 5.1 | 44.3 ± 16.3 | 83.8 ± 13.6 | 55.0 ± 33.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kong, Q.-Q.; Ding, X.-L.; Chen, Y.-F.; Ye, J.-R. Comparison of Morphological Indexes and the Pathogenicity of Bursaphelenchus xylophilus in Northern and Southern China. Forests 2021, 12, 310. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030310
AMA Style
Kong Q-Q, Ding X-L, Chen Y-F, Ye J-R. Comparison of Morphological Indexes and the Pathogenicity of Bursaphelenchus xylophilus in Northern and Southern China. Forests. 2021; 12(3):310. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030310
Chicago/Turabian StyleKong, Qing-Qing, Xiao-Lei Ding, Yi-Fan Chen, and Jian-Ren Ye. 2021. "Comparison of Morphological Indexes and the Pathogenicity of Bursaphelenchus xylophilus in Northern and Southern China" Forests 12, no. 3: 310. https://0-doi-org.brum.beds.ac.uk/10.3390/f12030310
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.