
Crown Structure Explains the Discrepancy in Leaf Phenology Metrics Derived from Ground- and UAV-Based Observations in a Japanese Cool Temperate Deciduous Forest


Abstract
:1. Introduction
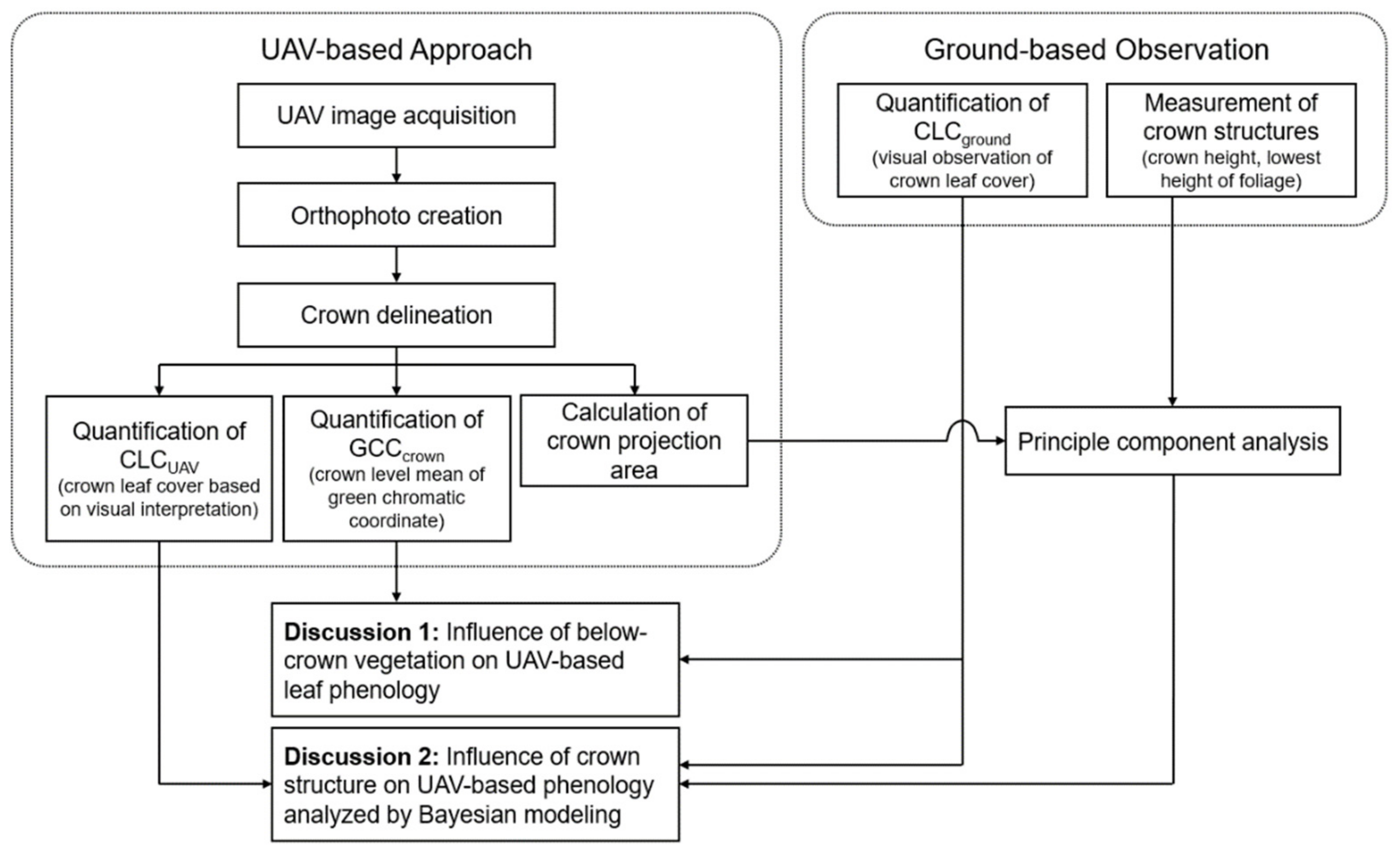
2. Materials and Methods
2.1. Study Site
2.2. Measurements of Crown Leaf Phenology
2.2.1. UAV-Based Observation
2.2.2. Ground-Based Observations
2.3. Crown Structural Traits
2.4. Data Analysis
3. Results
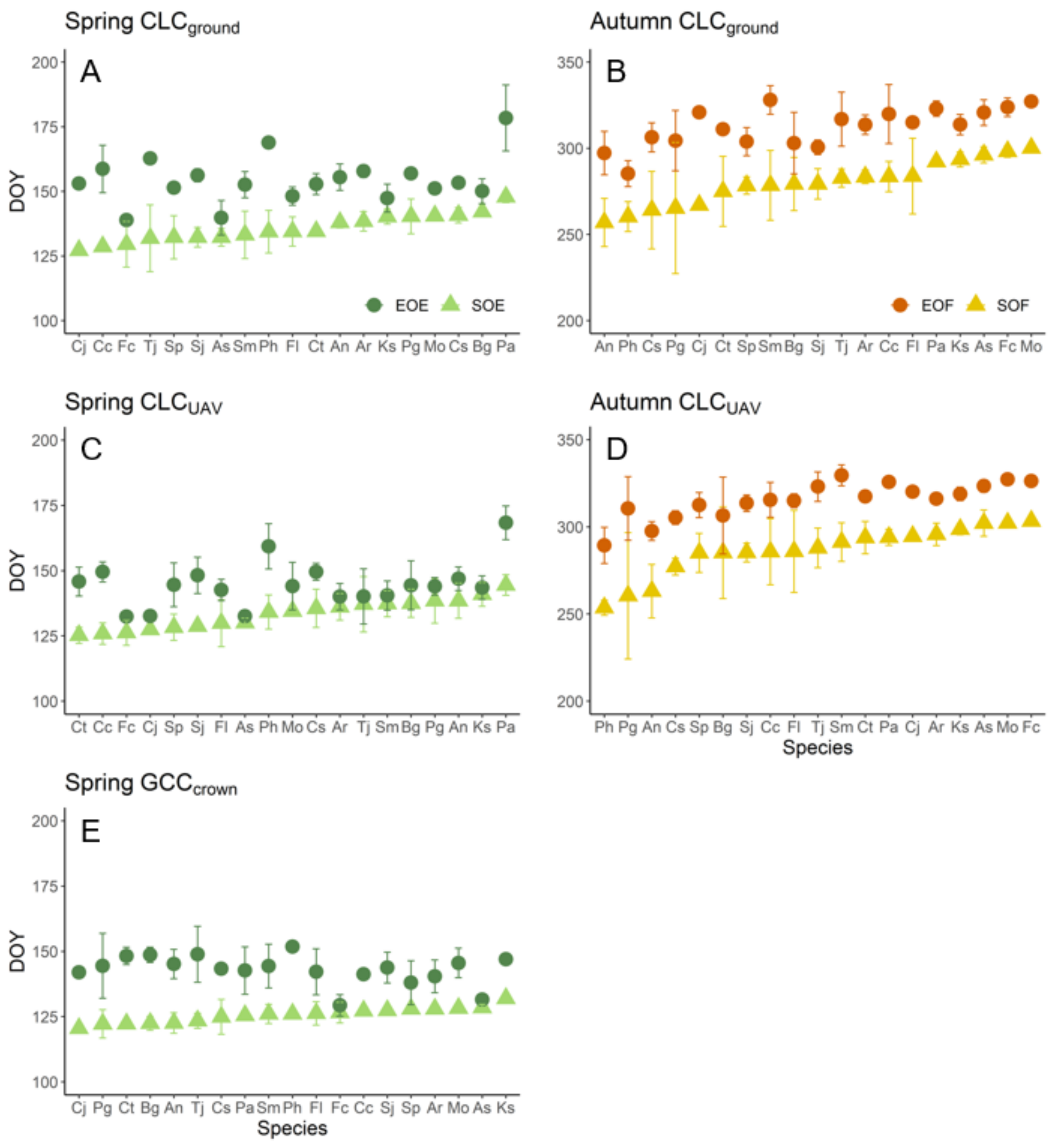
3.1. Inter-Species Differences in the Phenological Transition Dates Derived from the Ground Observations and UAV Images
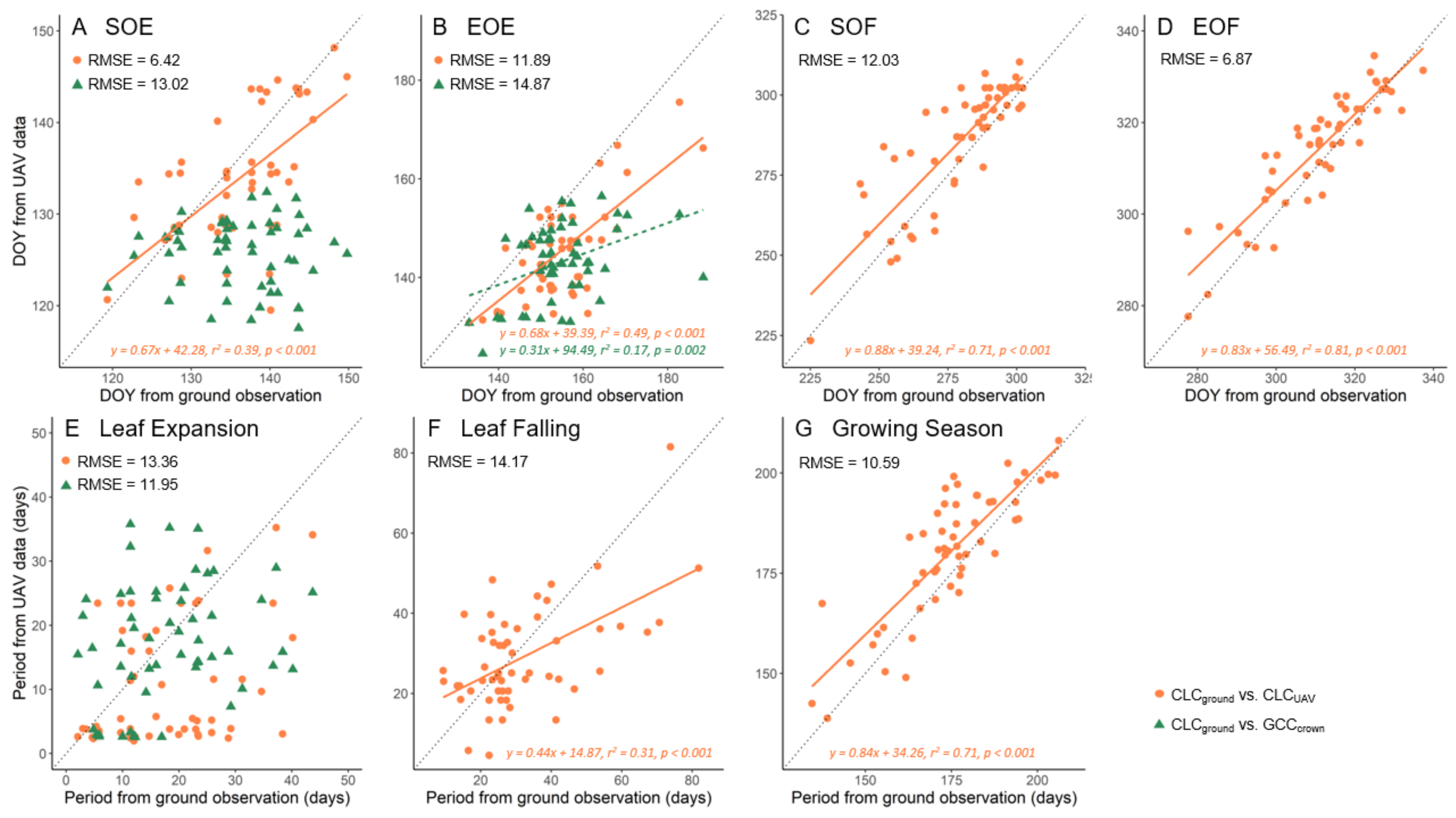
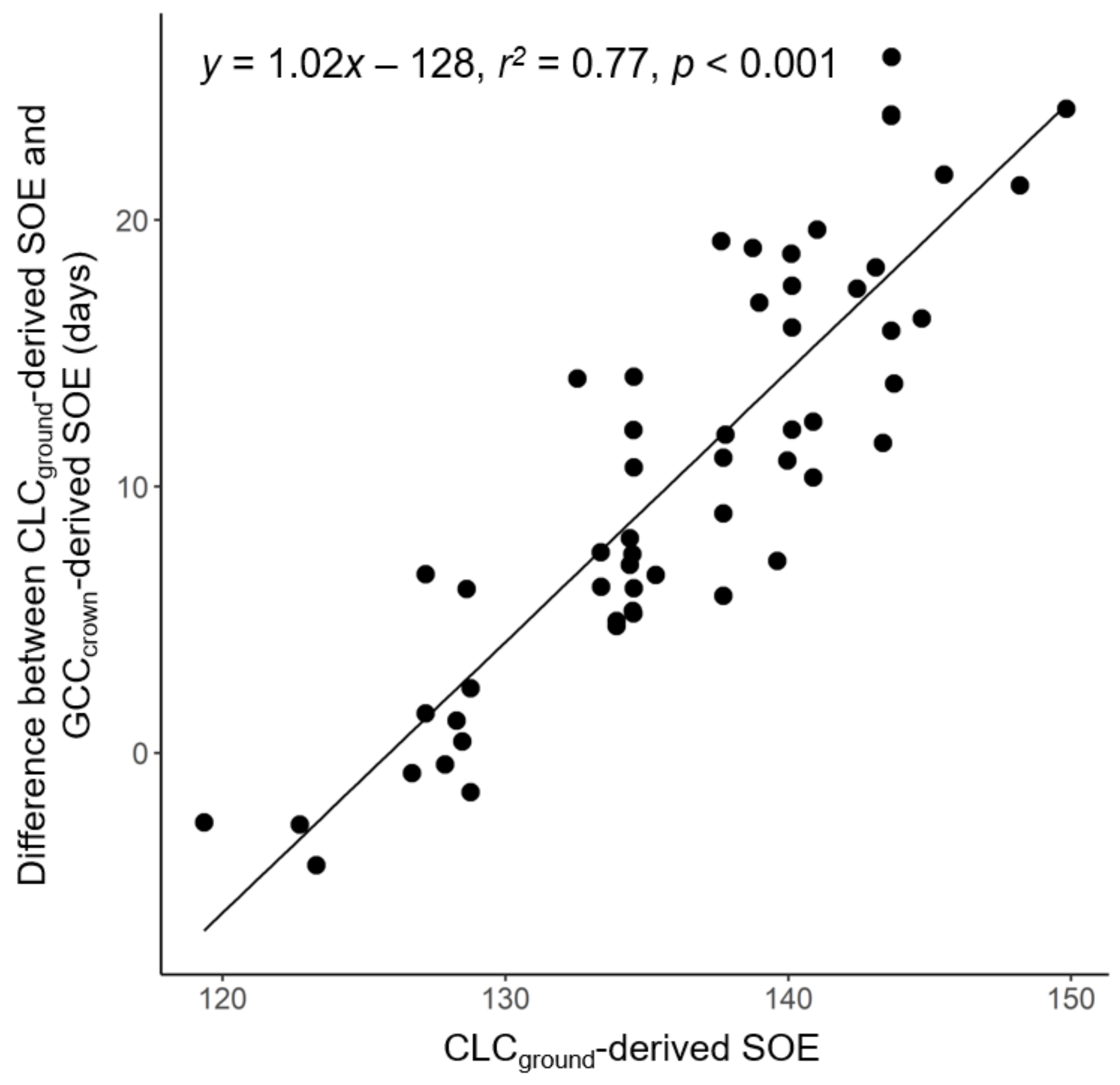
3.2. Direct Comparison of the Phenological Metrics Derived from UAV Data and Ground Observations
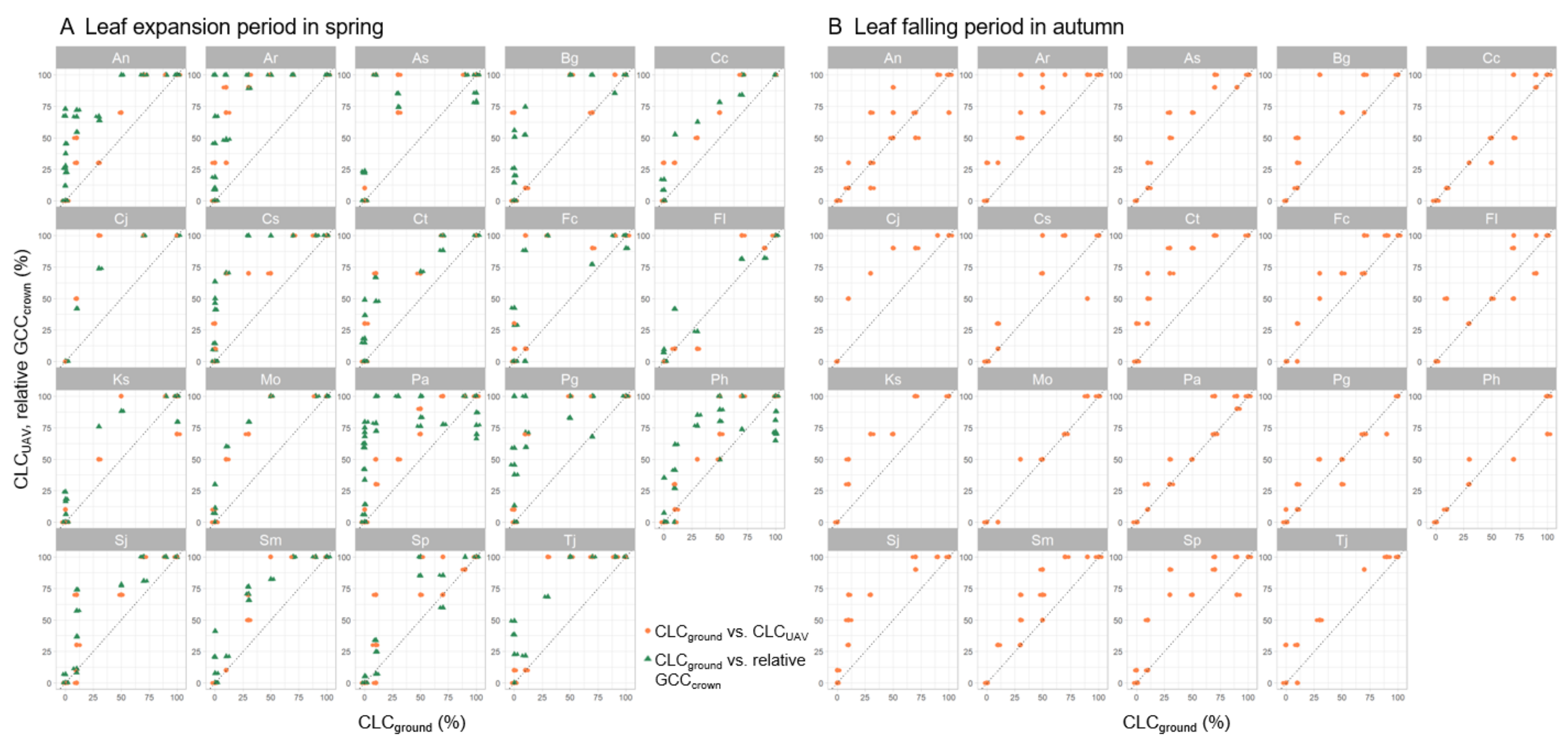
3.3. Contribution of Crown Structure to the Discrepancy between CLCUAV and CLCground
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| CA | Crown area |
| CA/CL | Ratio of CA to crown length |
| CH | Crown height |
| CL | Crown length |
| CV | Crown volume |
| CLCground | Crown leaf cover determined from ground observation |
| CLCUAV | Crown leaf cover determined from visual interpretation of UAV images |
| EOE | End date of leaf expansion |
| EOF | End date of leaf falling |
| GCCcrown | Mean crown-level green chromatic coordinate derived from UAV image |
| LAI | Leaf area index |
| SOE | Start date of leaf expansion |
| SOF | Start date of leaf falling |
| UAV | Unmanned aerial vehicle |
References
- Richardson, A.D.; Keenan, T.F.; Migliavacca, M.; Ryu, Y.; Sonnentag, O.; Toomey, M. Climate change, phenology, and phenological control of vegetation feedbacks to the climate system. Agric. For. Meteorol. 2013, 169, 156–173. [Google Scholar] [CrossRef]
- Klosterman, S.; Richardson, A.D. Observing spring and fall phenology in a deciduous forest with aerial drone imagery. Sensors 2017, 17, 2852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.Y.; Muller-Landau, H.C.; Lichstein, J.W.; Rifai, S.W.; Dandois, J.P.; Bohlman, S.A. Quantifying leaf phenology of individual trees and species in a tropical forest using unmanned aerial vehicle (UAV) images. Remote Sens. 2019, 11, 1534. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Braswell, B.H.; Hollinger, D.Y.; Jenkins, J.P.; Ollinger, S.V. Near-surface remote sensing of spatial and temporal variation in canopy phenology. Ecol. Appl. 2009, 19, 1417–1428. [Google Scholar] [CrossRef] [PubMed]
- Sonnentag, O.; Hufkens, K.; Teshera-Sterne, C.; Young, A.M.; Friedl, M.; Braswell, B.H.; Milliman, T.; O’Keefe, J.; Richardson, A.D. Digital repeat photography for phenological research in forest ecosystems. Agric. For. Meteorol. 2012, 152, 159–177. [Google Scholar] [CrossRef]
- Ahrends, H.E.; Bräugger, R.; Stöckli, R.; Schenk, J.; Michna, P.; Jeanneret, F.; Wanner, H.; Eugster, W. Quantitative phenological observations of a mixed beech forest in northern Switzerland with digital photography. J. Geophys. Res. Biogeosci. 2008, 113, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Richardson, A.D.; Hollinger, D.Y.; Dail, D.B.; Lee, J.T.; Munger, J.W.; O’Keefe, J. Influence of spring phenology on seasonal and annual carbon balance in two contrasting New England forests. Tree Physiol. 2009, 29, 321–331. [Google Scholar] [CrossRef]
- Migliavacca, M.; Sonnentag, O.; Keenan, T.F.; Cescatti, A.; O’Keefe, J.; Richardson, A.D. On the uncertainty of phenological responses to climate change, and implications for a terrestrial biosphere model. Biogeosciences 2012, 9, 2063–2083. [Google Scholar] [CrossRef] [Green Version]
- Hufkens, K.; Friedl, M.; Sonnentag, O.; Braswell, B.H.; Milliman, T.; Richardson, A.D. Linking near-surface and satellite remote sensing measurements of deciduous broadleaf forest phenology. Remote Sens. Environ. 2012, 117, 307–321. [Google Scholar] [CrossRef]
- Keenan, T.F.; Darby, B.; Felts, E.; Sonnentag, O.; Friedl, M.A.; Hufkens, K.; O’Keefe, J.; Klosterman, S.; Munger, J.W.; Toomey, M.; et al. Tracking forest phenology and seasonal physiology using digital repeat photography: A critical assessment. Ecol. Appl. 2014, 24, 1478–1489. [Google Scholar] [CrossRef] [Green Version]
- Mizunuma, T.; Wilkinson, M.; Eaton, E.L.; Mencuccini, M.; Morison, J.I.L.; Grace, J. The relationship between carbon dioxide uptake and canopy colour from two camera systems in a deciduous forest in southern England. Funct. Ecol. 2013, 27, 196–207. [Google Scholar] [CrossRef]
- Seiwa, K. Changes in leaf phenology are dependent on tree height in Acer mono, a deciduous broad-leaved tree. Ann. Bot. 1999, 83, 355–361. [Google Scholar] [CrossRef] [Green Version]
- Augspurger, C.K.; Cheeseman, J.M.; Salk, C.F. Light gains and physiological capacity of understorey woody plants during phenological avoidance of canopy shade. Funct. Ecol. 2005, 19, 537–546. [Google Scholar] [CrossRef]
- Lopez, O.R.; Farris-Lopez, K.; Montgomery, R.A.; Givnish, T.J. Leaf phenology in relation to canopy closure in southern Appalachian trees. Am. J. Bot. 2008, 95, 1395–1407. [Google Scholar] [CrossRef]
- Richardson, A.D.; O’Keefe, J. Phenological Differences between Understory and Overstory: A Case Study Using the Long-Term Harvard Forest Records. In Phenology of Ecosystem Processes: Applications in Global Change Research; Noormets, A., Ed.; Springer: New York, NY, USA, 2009; pp. 87–117. ISBN 978-1-4419-0025-8. [Google Scholar]
- Gressler, E.; Jochner, S.; Capdevielle-Vargas, R.M.; Morellato, L.P.C.; Menzel, A. Vertical variation in autumn leaf phenology of Fagus sylvatica L. in southern Germany. Agric. For. Meteorol. 2015, 201, 176–186. [Google Scholar] [CrossRef]
- Berra, E.F.; Gaulton, R.; Barr, S. Assessing spring phenology of a temperate woodland: A multiscale comparison of ground, unmanned aerial vehicle and Landsat satellite observations. Remote Sens. Environ. 2019, 223, 229–242. [Google Scholar] [CrossRef]
- Klosterman, S.T.; Hufkens, K.; Gray, J.M.; Melaas, E.; Sonnentag, O.; Lavine, I.; Mitchell, L.; Norman, R.; Friedl, M.A.; Richardson, A.D. Evaluating remote sensing of deciduous forest phenology at multiple spatial scales using PhenoCam imagery. Biogeosciences 2014, 11, 4305–4320. [Google Scholar] [CrossRef] [Green Version]
- Xie, Y.; Civco, D.L.; Silander, J.A. Species-specific spring and autumn leaf phenology captured by time-lapse digital cameras. Ecosphere 2018, 9, 1–21. [Google Scholar] [CrossRef] [Green Version]
- Valladares, F.; Niinemets, Ü. Shade Tolerance, a Key Plant Feature of Complex Nature and Consequences. Annu. Rev. Ecol. Evol. Syst. 2008, 39, 237–257. [Google Scholar] [CrossRef] [Green Version]
- Niinemets, Ü. A review of light interception in plant stands from leaf to canopy in different plant functional types and in species with varying shade tolerance. Ecol. Res. 2010, 25, 693–714. [Google Scholar] [CrossRef]
- Mizunaga, H.; Fujii, K. Is foliage within crowns of Cryptomeria japonica more heterogeneous and clumpy with age? J. Sustain. For. 2013, 32, 266–285. [Google Scholar] [CrossRef]
- Niinemets, Ü.; Valladares, F. Tolerance to shade, drought, and waterlogging of temperate northern hemisphere trees and shrubs. Ecol. Monogr. 2006, 76, 521–547. [Google Scholar] [CrossRef]
- Zhang, X.; Friedl, M.A.; Schaaf, C.B.; Strahler, A.H.; Hodges, J.C.F.; Gao, F.; Reed, B.C.; Huete, A. Monitoring vegetation phenology using MODIS. Remote Sens. Environ. 2003, 84, 471–475. [Google Scholar] [CrossRef]
- Kikuzawa, K. Leaf survival of woody plants in deciduous broad-leaved forests. 1. Tall trees. Can. J. Bot. 1983, 61, 2133–2139. [Google Scholar] [CrossRef]
- Aoki, Y.; Hashimoto, R. Leaf phenology of woody plant species in a cool-temperate secondary forest of Quercus serrata. Bull. Iwate Univ. For. 1995, 26, 29–41. [Google Scholar]
- Kato, S.; Yamamoto, M.; Komiyama, A. Leaf phenology of over-and understory trees in a deciduous broad-leaved forest: An observation at the Mumai plot in 1997. Japan J. For. Environ. 1999, 41, 39–44. [Google Scholar]
- Aikawa, T.; Tateno, R.; Takeda, H. Leaf phenology along a slope in a cool temperate deciduous forest. For. Res. Kyoto 2002, 74, 21–33. [Google Scholar]
- Vitasse, Y. Ontogenic changes rather than difference in temperature cause understory trees to leaf out earlier. New Phytol. 2013, 198, 149–155. [Google Scholar] [CrossRef] [PubMed]
- Larcher, W. Physiological Plant Ecology: Ecophysiology and Stress Physiology of Functional Groups, 4th ed.; Springer: Berlin/Heidelberg, Germany, 2003. [Google Scholar]
- Hagemeier, M.; Leuschner, C. Functional crown architecture of five temperate broadleaf tree species: Vertical gradients in leaf morphology, leaf angle, and leaf area density. Forests 2019, 10, 265. [Google Scholar] [CrossRef] [Green Version]
- Koike, T. Autumn coloring, photosynthetic performance and leaf development of deciduous broad-leaved trees in relation to forest succession. Tree Physiol. 1991, 7, 21–32. [Google Scholar] [CrossRef] [PubMed]
- Brown, L.A.; Dash, J.; Ogutu, B.O.; Richardson, A.D. On the relationship between continuous measures of canopy greenness derived using near-surface remote sensing and satellite-derived vegetation products. Agric. For. Meteorol. 2017, 247, 280–292. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.D.; Hanes, J.M.; Liang, L. Comparing carbon flux and high-resolution spring phenological measurements in a northern mixed forest. Agric. For. Meteorol. 2013, 169, 136–147. [Google Scholar] [CrossRef]
- Hall, R.J.; Fernandes, R.A.; Hogg, E.H.; Brandt, J.P.; Butson, C.; Case, B.S.; Leblanc, S.G. Relating aspen defoliation to changes in leaf area derived from field and satellite remote sensing data. Can. J. Remote Sens. 2003, 29, 299–313. [Google Scholar] [CrossRef]
- Hufkens, K.; Friedl, M.A.; Keenan, T.F.; Sonnentag, O.; Bailey, A.; O’Keefe, J.; Richardson, A.D. Ecological impacts of a widespread frost event following early spring leaf-out. Glob. Change Biol. 2012, 18, 2365–2377. [Google Scholar] [CrossRef]
- Iio, A. Masting changes canopy structure, light interception, and photosynthesis in Fagus crenata. In Proceedings of the 7th International Conference on Functional-Structural Plant Models, Saariselkä, Finland, 9–14 June 2013; p. 174. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Species Name | Species Code | Number of Samples | DBH (cm) | H (m) | CA (m2) | CL (m) | Successional Type |
|---|---|---|---|---|---|---|---|---|
| 1. | Acer nipponicum | An | 5 | 20.13–53.81 | 14.75–20.40 | 14.63–80.90 | 4.93–10.90 | Unknown |
| 2. | Acer rufinerve | Ar | 4 | 41.80–52.87 | 14.25–20.85 | 20.30–51.54 | 4.85–11.85 | Mid |
| 3. | Acer shirasawanum | As | 3 | 42.44–76.60 | 17.15–21.10 | 53.10–83.15 | 9.40–10.50 | Late |
| 4. | Betula grossa | Bg | 3 | 31.80–34.90 | 17.35–18.97 | 37.44–81.59 | 8.30–11.17 | Early |
| 5. | Carpinus japonica | Cj | 1 | 26.75 | 15.45 | 27.67 | 6.5 | Mid |
| 6. | Carpinus tschonoskii | Ct | 3 | 30.78–37.48 | 14.35–27.8 | 24.29–45.44 | 5.00–13.10 | Mid |
| 7. | Chengiopanax sciadophylloides | Cs | 3 | 34.62–41.50 | 16.65–19.90 | 16.51–39.82 | 7.30–9.15 | Mid |
| 8. | Cornus controversa | Cc | 2 | 22.63–55.21 | 13.50–20.95 | 5.55–111.77 | 3.20–11.35 | Mid |
| 9. | Fagus crenata | Fc | 3 | 45.75–59.46 | 23.00–32.35 | 71.35–132.09 | 14.10–23.10 | Late |
| 10. | Fraxinus lanuginosa | Fl | 3 | 23.22–32.47 | 14.35–16.05 | 27.73–52.09 | 4.05–6.45 | Mid |
| 11. | Kalopanax septemlobus | Ks | 3 | 29.39–37.44 | 12.95–17.73 | 21.62–45.32 | 3.95–8.83 | Mid |
| 12. | Magnolia obovata | Mo | 2 | 49.14–59.78 | 14.65–17.15 | 29.45–53.36 | 6.85–10.10 | Unknown |
| 13. | Phellodendron amurense | Pa | 3 | 24.90–39.39 | 11.80–22.60 | 11.89–65.37 | 3.65–9.07 | Early |
| 14. | Prunus grayana | Pg | 3 | 35.24–49.86 | 13.55–22.00 | 40.37–90.07 | 4.40–11.15 | Mid |
| 15. | Pterostyrax hispidus | Ph | 3 | 21.83–31.34 | 10.00–12.20 | 13.58–30.59 | 5.10–6.40 | Early |
| 16. | Stewartia monadelpha | Sm | 3 | 21.62–29.16 | 14.70–17.43 | 5.87–21.66 | 7.20–7.80 | Unknown |
| 17. | Stewartia pseudocamellia | Sp | 3 | 22.04–28.70 | 15.70–16.65 | 6.74–15.53 | 6.40–6.85 | Unknown |
| 18. | Styrax japonicus | Sj | 3 | 18.03–30.57 | 10.80–18.65 | 6.06–25.90 | 4.40–9.40 | Mid |
| 19. | Tilia japonica | Tj | 2 | 40.22–58.90 | 22.70–23.90 | 36.44–131.33 | 12.85–13.65 | Late |
| No. | Season | a | b | WAIC | ||||
|---|---|---|---|---|---|---|---|---|
| c | d | e | f | g | h | |||
| 1 | Spring | −0.05 | 1.16 | 0.42 | 2.55 | 4.036 | ||
| Autumn | 0.07 | 1.07 | −0.13 | 1.31 | ||||
| 2 | Spring | −0.04 | 1.17 | 0.43 | −0.20 | 2.55 | 4.038 | |
| Autumn | 0.06 | 1.08 | −0.11 | −0.13 | 1.32 | |||
| 3 | Spring | −0.05 | −0.02 | 1.17 | 0.42 | 2.55 | 4.038 | |
| Autumn | 0.08 | −0.09 | 1.07 | −0.13 | 1.31 | |||
| 4 | Spring | −0.01 | 1.16 | 0.45 | 2.55 | 4.043 | ||
| Autumn | −0.08 | 1.07 | −0.13 | 1.32 | ||||
| 5 | Spring | −0.04 | −0.04 | 1.17 | 0.45 | −0.21 | 2.55 | 4.045 |
| Autumn | 0.08 | −0.09 | 1.07 | −0.13 | −0.15 | 1.32 | ||
| 6 | Spring | −0.05 | 1.03–1.19 | 0.24 | −0.23 | 1.50–3.35 | 4.089 | |
| Autumn | 0.06 | 1.05–1.17 | −0.13 | 0.00 | 0.27–2.42 | |||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Budianti, N.; Mizunaga, H.; Iio, A. Crown Structure Explains the Discrepancy in Leaf Phenology Metrics Derived from Ground- and UAV-Based Observations in a Japanese Cool Temperate Deciduous Forest. Forests 2021, 12, 425. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040425
Budianti N, Mizunaga H, Iio A. Crown Structure Explains the Discrepancy in Leaf Phenology Metrics Derived from Ground- and UAV-Based Observations in a Japanese Cool Temperate Deciduous Forest. Forests. 2021; 12(4):425. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040425
Chicago/Turabian StyleBudianti, Noviana, Hiromi Mizunaga, and Atsuhiro Iio. 2021. "Crown Structure Explains the Discrepancy in Leaf Phenology Metrics Derived from Ground- and UAV-Based Observations in a Japanese Cool Temperate Deciduous Forest" Forests 12, no. 4: 425. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040425