
Effect of Temperature and Exposure Time on Cambium Cell Viability In Vitro for Eucalyptus Species


School of Ecosystem and Forest Sciences, The University of Melbourne, Water Street, Creswick, VIC 3363, Australia
*
Author to whom correspondence should be addressed.
Forests 2021, 12(4), 445; https://0-doi-org.brum.beds.ac.uk/10.3390/f12040445
Submission received: 12 February 2021
/
Revised: 25 March 2021
/
Accepted: 4 April 2021
/
Published: 6 April 2021
(This article belongs to the Section Forest Ecophysiology and Biology)
Abstract
:(1) Research Highlights: Thermal damage to cambium cells of Eucalyptus held in vitro was recorded at sublethal temperatures (40 °C–50 °C) when the duration of exposure extends beyond 2.5 min up to 5 min. (2) Background and Objectives: During a forest fire, heat can be transferred through tree bark potentially impacting viability of vascular cambium cells and the perennial growth of the tree. With the increased temperature of the cambium, cells are known to lose viability at temperatures exceeding 60 °C. However, it is possible that extended exposure to temperatures below 60 °C may also impair cell viability. This study aimed to identify the effect of the temperature and exposure time interaction on the cambium cell viability of Eucalyptus, a genus widely distributed in natural forests and commercial plantations globally. (3) Methods: Excised cambium-phloem tissue sections from three Eucalyptus species (Messmate–E. obliqua L’Hér., Narrow-leaf peppermint–E. radiata Sieber ex DC. and Swamp gum–E. ovata Labill.) were exposed in vitro to a series of temperature–time treatments (40 °C, 50 °C, 60 °C, 70 °C for 1 min, 2.5 min, and 5 min) and tested for cell viability using a tetrazolium reduction method. (4) Results: Cell viability of cambium cells decreased with increased temperature and exposure times for all three Eucalyptus species. Longer exposure to sublethal temperatures of 40 °C to 50 °C showed statistically similar results to shorter exposure to lethal temperatures (>50 °C). (5) Conclusions: Longer exposure to sublethal temperatures (40 °C–50 °C) caused irreversible thermal damage to cambium cells of Eucalyptus when tested in vitro, further refining our understanding of raised temperature on cell viability.
1. Introduction
Vascular cambium regulates the secondary growth of a tree by producing secondary xylem and phloem [1,2]. During a forest fire, trees are exposed to heat from direct flame or post-frontal radiation that may penetrate the bark to raise the temperature of the vascular cambium [3,4,5,6]. If the temperature of the cambium reaches 60 °C, cells lose their viability due to the denaturation of proteins of the plasma membrane and the cytoplasm and disrupting enzyme-mediated cell metabolism. Due to some combination of temperature and duration of heat exposure, irreversible thermal damage occurs, ceasing the production of xylem and phloem cells by cambium and potentially leading to tree death [7,8].
The traditional methodology for determining cell death following heat exposure has been to define a critical temperature at which cells lose viability, which in most of the previous studies has been demonstrated at 60 °C [9,10,11,12], including for Eucalyptus species [13]. However, while the 60 °C critical temperature may be useful in predicting cell death, several studies have emphasized that duration of heat exposure also plays a key role in determining the magnitude of heat damage to the cells [14,15,16,17,18,19,20]. According to Hare [15] and Kayll [16], prolonged exposure to temperatures below the 60 °C critical temperature can also cause irreversible thermal damage to cambium cells because the rate of cellular acclimation and repair processes are insignificant compared to the rate of thermal damage. Therefore, to develop a better understanding of heat impacts on cambium cell viability, it is essential to describe the interaction between temperature and time of exposure to raised temperature. Evidence for the decline in viability of cells with extended exposure to temperatures less than 60 °C comes from studies by Nielsen and Todd [21], who identified 43 °C–45 °C as the sublethal temperature for potato tubers (Solanum tuberosum L.), and Daniell et al. [22], who identified temperatures from 43 °C to 52 °C as the sublethal range (and temperature above 52 °C as the lethal range) for leaf cells of Soybean (Glycine max L.) and Elodea (Elodea canadensis Michx.). For the purpose of this study, 40 °C to 50 °C was considered the sublethal temperature range and temperatures above 50 °C were considered as lethal for cambium-phloem tissue samples.
Given the importance of fire in the ecology of many angiosperm-dominated forests, it is surprising that there are so few detailed studies of cambium cell damage associated with duration of heating. While Subasinghe Achchige et al. [13] identified 60 °C as the critical temperature in vitro for cambium-phloem tissues of Eucalyptus treated for one minute, their study did not encompass varying the duration of heating beyond a minute at temperatures less than 60 °C. Only the two studies of Dickinson et al. [19,20] have attempted to identify the duration of heating effects in woody tree species (Acer rubrum L., Quercus prinus L., Pseudotsuga menziesii Mirbel and Pinus ponderosa Douglas ex Lawson and C.Lawson) by quantifying cambium cell viability according to cell damage following heat treatments. As far as we are aware, there are no published studies of the role of heat duration on critical temperature for cambium cells of Eucalyptus species.
To determine any loss of cell viability with cambial heating below 60 °C requires varying the duration of heating (exposure time) over a range of fixed temperatures from 40 °C to 60 °C. Any loss of cell function following these temperature-exposure time treatments can be quantified as the cell viability index (CVI), typically by setting a CVI ≥ 0.37 as the threshold value for viable cells [20]. This approach offers the ability to identify critical temperature-exposure time interactions.
Prior to field experiments with species of different bark types (in vivo heating of the cambium), in vitro testing of cambium cells is required to identify the critical temperature-exposure time treatments for cambium cell viability. It is essential that method for measuring cell viability following temperature-exposure time treatments is rapid and of minimum interference with the cell viability of the tissue.
This study aimed to determine the interacting effects of temperature and duration of exposure on the cambium cell viability of three Eucalyptus species (E. obliqua, E. radiata, and E. ovata), and if these species respond differently to temperature–time treatments. Using the method optimized by Subasinghe Achchige et al. [13] and based on results of Dickinson et al. [20], a loss of 63.2% or more of cell viability (CVI ≥ 0.37) was chosen as the criterion for determining lethal temperature-exposure time treatments.
2. Materials and Methods
Reduction of 2,3,5-triphenyl tetrazolium chloride (TTC), described in Subasinghe Achchige et al. [13], was used to test cell viability following the ‘’exposure time-temperature’’ treatment as described below. A Neutral Red method described by the same study was used to cross validate the results of the TTC method.
2.1. Sample Collection and Preparation
Mature trees of E. obliqua (Messmate), E. radiata (Narrow-leaf peppermint), and E. ovata (Swamp gum) grown in Central Victoria, Australia were randomly selected for this study (Supplementary Materials Table S1) (Figure 1a–c). A circular bark window (Figure 1d,e) of 8.0 cm diameter was removed from the stem at 1.3 m height on the east side of the tree with a drill fitted with a cutting tool. To limit cell dehydration, paraffin film was applied immediately to the inner face of bark samples prior to transport to the laboratory and cooling to 4 °C.
To obtain tissue samples comprised of cambial cells and immediate phloem cells [13], the outer bark was first removed and a clean cuboid (0.5 cm width, 0.5 cm height, 5.0 cm length, Figure 1f) of cambium-phloem cells was cut with a sharp scalpel blade. Thin slices of the optimum size (0.5 cm length × 0.5 cm width × 0.1 cm thickness-average 0.01 g dry weight) were obtained from the cuboids (Figure 1g, bottom). Slices were immediately transferred to 0.05 mol/L potassium phosphate buffer solutions of pH 7.4 kept at 20 °C and held in plastic vials.
Cross sections (ideally uni-cellular or bi-cellular layers) were also obtained from the same cuboid (Figure 1g, top; Figure S1) to test for CVI with the Neutral Red method (see Appendix A). Cross sections were immediately transferred to 0.05 mol/L potassium phosphate buffer solutions of 7.2 pH and kept at 20 °C.
2.2. Exposure Time-Temperature Treatments
The range of temperature (40–70 °C) and exposure time (1 min–5 min) treatments tested are shown in Table 1. The tissue samples were placed in a hot water bath (Figure 1h) at a set temperature of either 1 min, 2.5 min or 5 min. The control samples were kept in the buffer solution at 20 °C for 2.5 min. The temperature of water in the water bath and of the buffer solution in the vials was measured simultaneously to ensure consistent heating. A timer was used to measure the exposure time. In the TTC method five tissue slices were tested for each treatment comprised of a combination of temperature and exposure time. For the cross validation with results from the Neutral Red method, three cross sections from each species were treated at 60 °C for 1 min, 2.5 min or 5 min.
2.3. Application of the TTC Method
Following treatment, slices were placed in vials containing 1.5 mL of 0.8% tetrazolium chloride (TTC) solution (Figure 1i) prepared in 0.05 M potassium phosphate buffer (pH 7.4). The samples were vacuum-infiltrated for six cycles of 15 s each and held at room temperature (20 °C) for 20–22 h in the dark. Slices were then removed from the TTC solution and rinsed twice with deionized water, blot dried, and macerated using a mortar and pestle. Constituents were then transferred to Falcon® conical tubes (Corning Inc., Corning, NY, USA) containing 4 mL of 95% ethyl alcohol, vortexed for 30 s, and kept in 95% ethyl alcohol for 12–16 h to allow formazan extraction (Figure 1j). The supernatant was then centrifuged at 3000 rpm for 5 min, and the absorbance at 485 nm measured using a UV-1800 spectrophotometer (SHIMADZUTM, Kyoto, Japan) (Figure 1k). Macerated tissue in each Falcon® tube was oven dried at 70 °C for 12 h and weighed (Figure 1l). The formazan concentration was calculated from the standard curve (Supplementary Materials, Figure S2) and the mass of formazan reported on a sample dry weight basis.
The cell viability index (CVI) was calculated by dividing the formazan concentration of a treated sample by the mean formazan concentration of the control (20 °C) samples (Equation (1)).
A CVI of 36.8% was applied as the critical threshold for cambium-phloem cell viability of Eucalyptus, after the study of Dickinson et al. [20].
2.4. Statistical Analysis
A linear mixed model (LMM) fitted by restricted maximum likelihood (REML) was applied to test the effect of species, temperature, time, and their interaction on the CVI. The fixed factors were “Species”, “Temperature”, and “Time”; “Species/Tree” (Tree nested within Species) was used as the random part of the model. Type III ANOVA with Satterthwaite’s Approximation was used to test the significance of the model parameters. Post hoc tests assessed the significance between the temperatures and the exposure time. The differences were considered significant at 5% level. Based on the results of the LMM, for the data visualization purpose, One-Way ANOVA was applied to test effect of 13 temperature and time treatment combinations (e.g., 20-2.5, 40-1, 40-2.5) on CVI.
For each analysis, normality and heterogeneity of variance were tested graphically using histograms of residuals and fitted value plots. Based on visual assessments, log transformation of CVI values was required to overcome heteroscedasticity prior the analysis. The analyses were completed in the R statistical environment, version 3.5.0 (R Core Team: Vienna, Austria, 2018), using the packages lme4 v.1.1-21 [23] and lmerTest v.3.1-1 [24].
3. Results
The three Eucalyptus species responded similarly to the temperature and time treatments (Table 2; p = 0.237), thus the results from all species were grouped and analyzed as one. Both increasing temperature and increasing exposure time reduced the cell viability index (Table 2; p < 0.001; Figure 2).
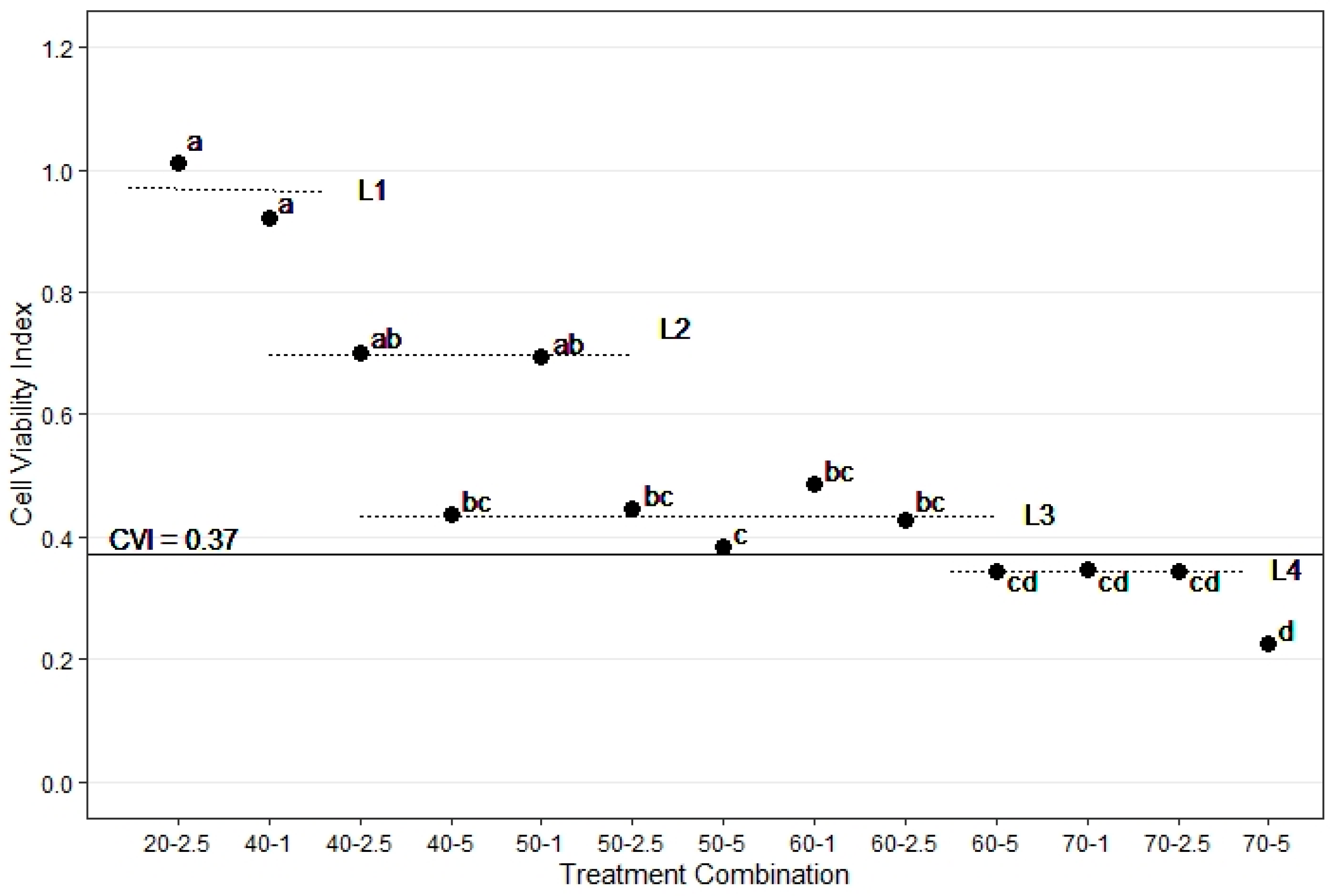
Up to 2.5 min exposure to sublethal temperatures (i.e., 1 min or 2.5 min at 40 °C and 1 min at 50 °C) did not significantly affect CVI, which remained comparable to the control samples (Figure 2, L2). Yet a fifty percent decrease in cell viability was observed when samples were exposed for 2.5 min or longer to sublethal temperatures (from a CVI of 92% at 40 °C for 1 min down to 44% at 5 min). The CVI of the samples treated at 40 °C for 5 min (0.435 ± 0.048), 50 °C for 2.5 min (0.445 ± 0.046), and 60 °C for 1 min (0.487 ± 0.080) to 2.5 min (0.425 ± 0.038) were statistically similar (Figure 2, L3). Longer exposure to temperatures below 60 °C and a short exposure to ≥70 °C further decreased cell viability to about 35% (Figure 2, L4). Exposure of cambium cells for 5 min to 70 °C decreased the CVI to 0.23. Overall, the CVI was less than the 0.37 threshold for each of four exposure time-temperature treatments (60-5, 70-1, 70-2.5, and 70-5; Figure 2).
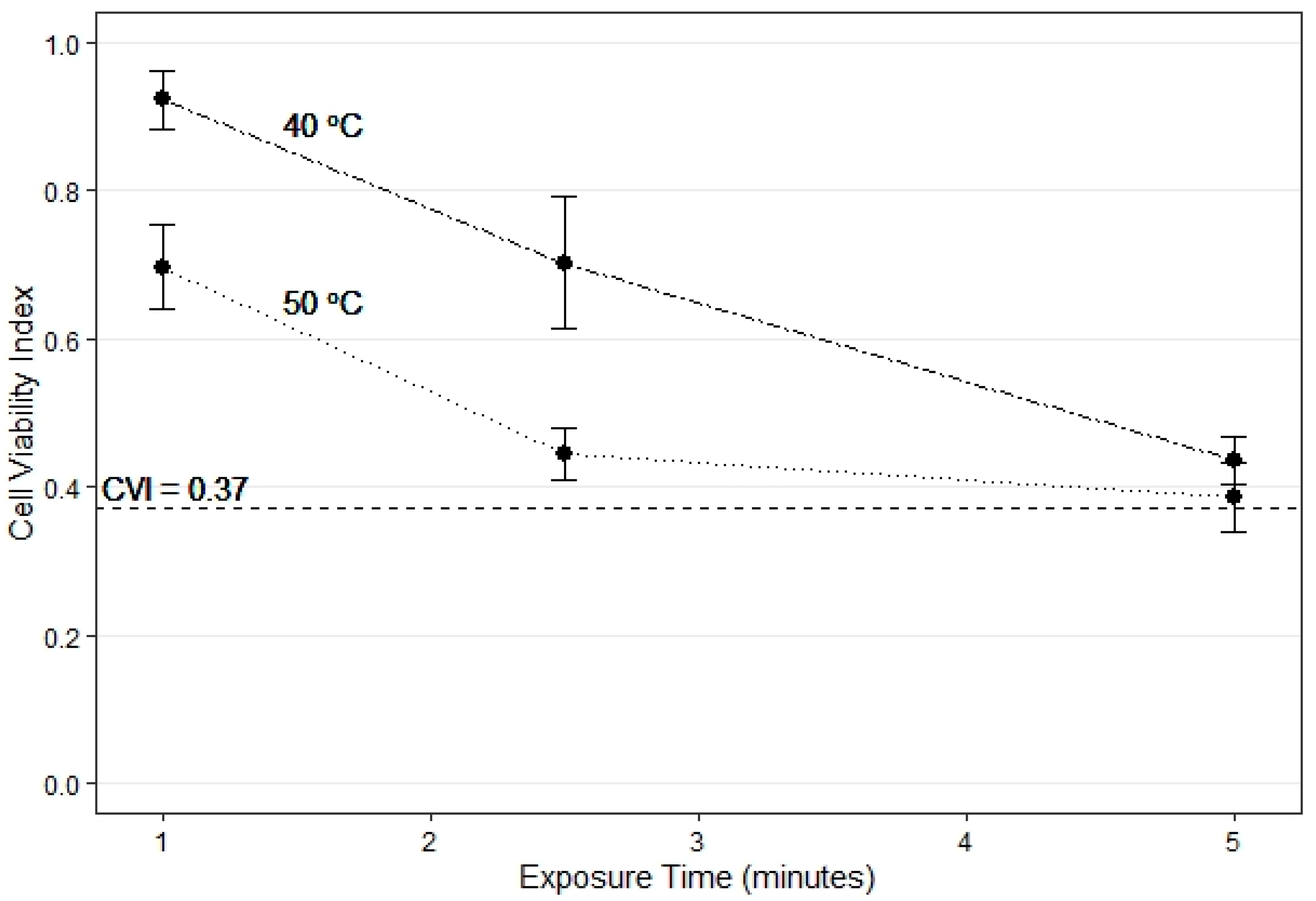
At the sublethal temperatures of 40–50 °C, the decrease in CVI was more rapid from 1 to 2.5 min than from 2.5 to 5 min, indicating that most cell damage occurs in this exposure range (Figure 3). Further increase in exposure time to 5 min showed a decrease in CVI towards the critical threshold of 0.37 (Figure 3).
The results for CVI assessed with the Neutral Red method were consistent with results from the TTC method, with all species responding similarly (p = 0.281) to temperature and time treatments. Therefore, the results of the Neutral Red method confirmed that cell viability decreases with increased duration of exposure. For example, results from the Neutral Red method show that at 60 °C the CVI declines from 0.43 ± 0.01 at 1 min. 1 to 0.26 ± 0.04 at 5 min (p = 0.002; Figure S4).
4. Discussion
This study of Eucalyptus cambium cells in vitro shows that increasing temperature and exposure time reduces cell viability, with longer exposure to sublethal temperatures of 40 °C and 50 °C resulting in cells reaching the critical CVI. The observed decrease in cell viability with increasing temperature in this study confirms that Eucalyptus is similar to results from vascular cambium of red maple (Acer rubrum L.) and chestnut oak (Quercus prinus L.) trees [19], and also to results from cucumber (Cucumis sativus L.) [18] and crop plants (leaves of tomato and common bean) [25].
Some studies consider that thermal damage to cells in the sublethal temperature range is reversible [7,8]. According to Daniell et al. [22], sublethal temperature treatments caused cessation of cytoplasm streaming and enlargement of chloroplasts, but cytoplasm resumed streaming 6 to 8 h after the heat treatments. In contrast, Dickinson et al. [20] reported that prolonged exposure to sublethal temperature may result in irreversible thermal damage to the cells. Results of the current study support Dickinson et al. [20] with the CVI of the Eucalyptus cells treated at 40 °C to 5 min and 50 °C to 2.5 min being statistically similar to CVI of cells treated at 60 °C to 1 min, suggesting that prolonged exposure to sublethal temperatures is similar to short exposure to the lethal temperature. The CVI values of the 40-5 and 60-5 treatment combinations were statistically similar (Figure 2), implying that 5-min exposure to sublethal and lethal temperatures may result in similar irreversible thermal damage to cambium-phloem tissue of Eucalyptus when treated in vitro.
The results of this study reveal the shortcomings of relying solely on explicit critical temperature as an indicator of cell mortality, because longer exposure to sublethal temperature will also cause cambium cell death. Relying on measurement of whether heated tissues reach the critical temperature (60 °C) is not sufficient for accurately quantifying cell impairment and for predicting likely tree mortality. Therefore, the best approach for investigating the temperature and time effect on cambium cell viability is to test impacted tissues with the TTC method to determine a CVI, rather than just measuring the temperature of the cambium, to build a picture of the heat tolerance capacity of cambium cells for a given species. Furthermore, because the cambium cells from the three Eucalyptus species responded in a similar way to temperature–time treatments, investigation of the effectiveness of thermal insulation properties of the bark can assume there are no heat tolerance differences at a cellular level. With this knowledge, future studies (in vivo) can focus on bark properties such as thickness, density, and moisture content that regulate the rate of heat transfer and heat residence time (duration of heat exposure) inside the cambium [4,26,27,28,29,30]. When assessing such insulating properties of the bark, heat from an external heat source is applied while the bark is intact in the log or standing tree. Subsequently, heat insulation of the bark is quantified by measuring the time required for heat to transfer through the bark towards the vascular cambium potentially raising the cambium temperature to 60 °C [6,12,20,21,29,30]. However, when assessing the thermal damage to cambium cells, instead of only measuring the heat residence time for which the cambium was exposed to 60 °C, the heat residence time of cambium exposure to temperatures above >40 °C should also be considered.
Depending on the difference in heat-insulating properties of the bark among species, heat residence time inside the bark may differ [4,26,27,28,29,30]. Thus, the results of this study will provide rich opportunities for future studies to compare the effect of different bark types of Eucalyptus species on tree mortality following fire. Ultimately, this revised knowledge of the heat tolerance regime of Eucalyptus may assist land managers in a practical way to maintain a non-lethal maximum fire intensity during prescribed and regeneration burning to ensure tree survival [12].
5. Conclusions
Both the degree of heating (temperature maximum) and exposure time (duration of raised temperature) determined the cambium cell viability following heat exposure. Short exposure to the lethal temperatures (≥60 °C), and longer exposure to sublethal temperatures, causes irreversible thermal damage to cambium cells of Eucalyptus species. Knowledge of the loss of cell viability during minutes long exposure to 40 °C or more demonstrates that so-called sublethal temperatures can be lethal for cambium cells. Furthermore, studies of the protective properties of bark can now proceed with improved knowledge of the survival envelope of the temperature duration of heating impacts on cells. This will allow for more confident prediction of the likelihood of tree mortality, based on bark condition and fuel parameters, following fires of increasing severity.
Supplementary Materials
The following are available online at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f12040445/s1, Figure S1: Microscopic image of a cross section of a stained secondary phloem tissue of E. obliqua. CM—cambium, DC—dead cell, FC—false live cell, LC—live cell, PF—phloem fibres, PS—phloem sieve tube; Figure S2: Standard absorbance curve for 1,3,5 triphenyl tetrazolium formazan at 20 °C; Figure S3: Closed circles indicate the average cell viability index (CVI) values of all species for each treatment combination. Error bar indicates the standard error of mean at 95% confidence interval. Different lower case (in bold) letters adjacent to each closed circle indicates the significant difference tested for the average of log transformed CVI values of each treatment combination; Figure S4: Closed circles indicate the average cell viability index (CVI) values of all species treated at 60 °C for 1 min, 2.5 min, and 5 min in the Neutral Red method. Error bar indicates the standard error of mean at 95% confidence interval. Different lower case (in bold) letters adjacent to each closed circle indicates the significant difference tested for the average of log transformed CVI values of each treatment combination; Table S1: Tree data of the three Eucalyptus species.
Author Contributions
Conceptualization, Y.M.S.A., L.V. and C.J.W.; methodology, Y.M.S.A., L.V. and C.J.W.; software, Y.M.S.A.; formal analysis, Y.M.S.A., L.V. and C.J.W.; investigation, Y.M.S.A., L.V. and C.J.W.; resources, Y.M.S.A., L.V. and C.J.W.; data curation, Y.M.S.A.; writing—original draft preparation, Y.M.S.A., L.V. and C.J.W.; writing—review and editing, Y.M.S.A., L.V. and C.J.W.; validation, Y.M.S.A., L.V. and C.J.W.; project administration, Y.M.S.A., L.V. and C.J.W. All authors have read and agreed to the published version of the manuscript.
Funding
Authors received no external funding for this study.
Data Availability Statement
The data presented in this study are available on request from the corresponding author. The data are not publicly available due to this study being a part of a PhD thesis in submission.
Acknowledgments
Authors acknowledge Vincent White and Leon Holt from VicForests for lending their assistance in obtaining the Forest Produce License, and Leon Holt for his assistance in selecting field sites. We thank Steve Smith for his technical support during the sample collection. We also thank Philip Polglase, Fedor Torgovnikov, and Rajitha Ratnayake for their assistance during sample collection and handling. Yasika Subasinghe Achchige acknowledges the Melbourne Research Scholarship from The University of Melbourne.
Conflicts of Interest
The authors declare no conflict of interest.
Appendix A
- Neutral Red Method
Following the temperature-exposure time treatments the cross sections were transferred to freshly prepared neutral red solution (20 ppm), made up in a 0.05 mol/L potassium phosphate buffer (pH 7.2) and vacuum infiltrated for 30 s for consistent staining [19]. Tissue sections were steeped in the stain for 1 hour and then washed with distilled water to remove background residual stain prior to microscopy. Cell staining was observed with slides of tissue sample mounted in a compound light microscope set at magnification of × 40 (XSP-103a, Xiamen Phio Scientific Instruments Co., Ltd., Xiamen, China). Three field view counts were made over the entire cross section and later scaled up to 100. Cells stained as light red were counted as alive (true live cells) and unstained cells were counted as “dead cells” and cells appeared in dark red colour due to the presence of tannin were counted as “False live cells” [13]. The Cell Viability Index for each field view was calculated by dividing the number of live cells of the each filed view by the mean number of live cells in the samples exposed to 20 °C treatment (Equation (A1)).
References
- Evert, R.F. Vascular cambium. In Esau’s Plant Anatomy, 3rd ed.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2006; pp. 323–355. [Google Scholar]
- Beck, C.B. An Introduction to Plant Structure and Development. Plant Anatomy for the Twenty-First Century; Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Gill, A.M.; Ashton, D.H. The role of bark type in relative tolerance to fire of three central Victorian eucalypts. Aust. J. Bot. 1968, 16, 491–498. [Google Scholar] [CrossRef]
- Vines, R. Heat transfer through bark, and the resistance of trees to fire. Aust. J. Bot. 1968, 16, 499–514. [Google Scholar] [CrossRef]
- Costa, J.; Oliveira, L.; Viegas, D.; Neto, L. On the temperature distribution inside a tree under fire conditions. Int. J. Wildland Fire 1991, 1, 87–96. [Google Scholar] [CrossRef]
- Bär, A.; Michaletz, S.T.; Mayr, S. Fire effects on tree physiology. New Phytol. 2019, 223, 1728–1741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, G.N.; Gill, A.M.; Weber, R.O. A time dependent model of fire impact on seed survival in woody fruits. Aust. J. Bot. 1994, 42, 71–81. [Google Scholar] [CrossRef]
- Mercer, G.N.; Weber, R.O. Fire plumes. In Forest Fires: Behavior and Ecological Effects; Johnson, E., Miyanishi, K., Eds.; Academic Press: New York, NY, USA, 2001; pp. 225–255. [Google Scholar]
- Brown, J.K.; DeByle, N.V. Fire damage, mortality, and suckering in aspen. Can. J. For. Res. 1987, 17, 1100–1109. [Google Scholar] [CrossRef]
- Gutsell, S.L.; Johnson, E.A. How fire scars are formed: Coupling a disturbance process to its ecological effect. Can. J. For. Res. 1996, 26, 166–174. [Google Scholar] [CrossRef]
- Bauer, G.; Speck, T.; Blomer, J.; Bertling, J.; Speck, O. Insulation capability of the bark of trees with different fire adaptation. J. Mater. Sci. 2010, 45, 5950–5959. [Google Scholar] [CrossRef]
- Espinosa, J.; Rodríguez de Rivera, O.; Madrigal, J.; Guijarro, M.; Hernando, C. Predicting potential cambium damage and fire resistance in pinus nigra arn. Ssp. Salzmannii. For. Ecol. Manag. 2020, 474, 118372. [Google Scholar] [CrossRef]
- Subasinghe Achchige, Y.M.; Volkova, L.; Drinnan, A.; Weston, C.J. A quantitative test for heat-induced cell necrosis in vascular cambium of Eucalyptus obliqua stems. J. Plant. Ecol. 2020, 14, 160–169. [Google Scholar] [CrossRef]
- Lorenz, R.W. High temperature tolerance of forest trees. Univ. Minnesota Agric. Exp. Sta. Tech. Bull. 1939, 141, 25. [Google Scholar]
- Hare, R.C. Heat Effects on Living Plants; Occasional Paper 183; USDA Forest Service, Southern Forest Experiment Station: New Orleans, LA, USA, 1961; pp. 1–32. [Google Scholar]
- Kayll, A.J.; Canada Department of Forestry. Heat Tolerance of Scots Pine Seedling Cambium Using Tetrazolium Chloride to Test Viability. Available online: books.google.ro/books?id=xeYsAQAAMAAJ (accessed on 2 January 2021).
- Martin, R.E. A basic approach to fire injury of tree stems. In Proceedings of the Second Annual Tall Timbers Fire Ecology Conference, Tallahassee, FL, USA, 14–15 March 1963; pp. 151–162. [Google Scholar]
- Caldwell, C.R. Estimation and analysis of cucumber (Cucumis sativus L.) leaf cellular heat sensitivity. Plant Physiol. 1993, 101, 939–945. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dickinson, M.B.; Johnson, E.A. Temperature-dependent rate models of vascular cambium cell mortality. Can. J. For. Res. 2004, 34, 546–559. [Google Scholar] [CrossRef]
- Dickinson, M.B.; Jolliff, J.; Bova, A. Vascular cambium necrosis in forest fires: Using hyperbolic temperature regimes to estimate parameters of a tissue-response model. Aust. J. Bot. 2004, 52, 757–763. [Google Scholar] [CrossRef]
- Nielsen, L.W.; Todd, F.A. Bacterial soft rot of irish potatoes as influenced by sublethal temperatures. Am. Potato J. 1946, 23, 73–87. [Google Scholar] [CrossRef]
- Daniell, J.W.; Chappell, W.E.; Couch, H.B. Effect of sublethal and lethal temperature on plant cells. Plant Physiol. 1969, 44, 1684–1689. [Google Scholar] [CrossRef]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Lme4: Linear Mixed-Effects Models Using Eigen and s4. R package version 1.1-10. Available online: http://CRAN.R-project.org/package=lme4 (accessed on 2 January 2021).
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B. lmerTest package: Tests in linear mixed effects models. J. Stat. Softw. 2017, 82, 1–26. [Google Scholar] [CrossRef] [Green Version]
- Chen, H.H.; Shen, Z.Y.; Li, P. Adaptability of crop plants to high temperatures stress. Crop Sci. 1982, 22, 719–725. [Google Scholar] [CrossRef]
- Uhl, C.; Kauffman, J.B. Deforestration, fire susceptibility and potential tree responses to fire in the eastern Amazon. Ecology 1990, 71, 437–449. [Google Scholar] [CrossRef]
- Hengst, G.E.; Dawson, J.O. Bark properties and fire resistance of selected tree species from the central hardwood region of North America. Can. J. For. Res. 1994, 24, 688–696. [Google Scholar] [CrossRef]
- Lawes, M.J.; Richards, A.; Dathe, J.; Midgley, J.J. Bark thickness determines fire resistance of selected tree species from fire-prone tropical savanna in north Australia. Plant Ecol. 2011, 212, 2057–2069. [Google Scholar] [CrossRef]
- Wesolowski, A.; Adams, M.A.; Pfautsch, S. Insulation capacity of three bark types of temperate eucalyptus species. For. Ecol. Manag. 2014, 313, 224–232. [Google Scholar] [CrossRef]
- Bär, A.; Mayr, S. Bark insulation: Ten Central Alpine tree species compared. For. Ecol. Manag. 2020, 474, 118361. [Google Scholar] [CrossRef]
Figure 1.
(a–c) Eucalyptus species used for the study, (d) circular bark window removed from the tree stem, (e) circular bark window, (f) cuboid containing both phloem and cambium, (g) cross sections (top), slices (bottom), (h) samples treated in a hot water bath, (i) treated samples transferred to TTC solution, (j) formazan diffused into 95% ethyl alcohol from the macerated samples, (k) absorbance measured using a spectrophotometer, and (l) macerated dry tissue weighted in an analytical balance.
Figure 1.
(a–c) Eucalyptus species used for the study, (d) circular bark window removed from the tree stem, (e) circular bark window, (f) cuboid containing both phloem and cambium, (g) cross sections (top), slices (bottom), (h) samples treated in a hot water bath, (i) treated samples transferred to TTC solution, (j) formazan diffused into 95% ethyl alcohol from the macerated samples, (k) absorbance measured using a spectrophotometer, and (l) macerated dry tissue weighted in an analytical balance.

Figure 2.
The average cell viability index (CVI; closed circles) of all Eucalyptus species grouped together, for each treatment combination. Lower-case letters indicate significantly different treatments, based on the average of log transformed CVI. For the purpose of discussing the results, the horizontal dotted lines (L1, L2, L3, and L4) group neighboring overlapping CVI treatments. The horizontal solid line shows the critical CVI (0.37) below which cells are considered dead. See Figure S3 for 95% CI range.
Figure 2.
The average cell viability index (CVI; closed circles) of all Eucalyptus species grouped together, for each treatment combination. Lower-case letters indicate significantly different treatments, based on the average of log transformed CVI. For the purpose of discussing the results, the horizontal dotted lines (L1, L2, L3, and L4) group neighboring overlapping CVI treatments. The horizontal solid line shows the critical CVI (0.37) below which cells are considered dead. See Figure S3 for 95% CI range.

Figure 3.
Average cell viability index (CVI) for all Eucalyptus species grouped together (closed circles) for exposure time up to 5 min. for the 40 °C (dashed) and 50 °C (dotted) temperature treatments. The horizontal dashed line is the critical CVI of 0.37.
Figure 3.
Average cell viability index (CVI) for all Eucalyptus species grouped together (closed circles) for exposure time up to 5 min. for the 40 °C (dashed) and 50 °C (dotted) temperature treatments. The horizontal dashed line is the critical CVI of 0.37.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Matrix of the temperature–time treatment combinations that were applied to cambium-phloem cells of the three Eucalyptus species.
Table 1.
Matrix of the temperature–time treatment combinations that were applied to cambium-phloem cells of the three Eucalyptus species.
| Time (min) | 1 | 2.5 | 5 | |
|---|---|---|---|---|
| Temperature (°C) | ||||
| 40 | 40-1 | 40-2.5 | 40-5 | |
| 50 | 50-1 | 50-2.5 | 50-5 | |
| 60 | 60-1 | 60-2.5 | 60-5 | |
| 70 | 70-1 | 70-2.5 | 70-5 | |
Table 2.
Summary of the statistical analysis of the cell viability index (CVI) of Eucalyptus according to species, temperature, and time (duration of exposure) treatments.
Table 2.
Summary of the statistical analysis of the cell viability index (CVI) of Eucalyptus according to species, temperature, and time (duration of exposure) treatments.
| Parameter | Method | p-Value |
|---|---|---|
| Species | LMM | 0.237 |
| Temperature | LMM | <0.001 |
| Time (duration of exposure) | LMM | <0.001 |
| Temperature × Time | LMM | 0.069 |
| Treatment combination | One-way ANOVA | 0.002 |
LMM-Linear Mixed Model.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Subasinghe Achchige, Y.M.; Volkova, L.; Weston, C.J. Effect of Temperature and Exposure Time on Cambium Cell Viability In Vitro for Eucalyptus Species. Forests 2021, 12, 445. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040445
AMA Style
Subasinghe Achchige YM, Volkova L, Weston CJ. Effect of Temperature and Exposure Time on Cambium Cell Viability In Vitro for Eucalyptus Species. Forests. 2021; 12(4):445. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040445
Chicago/Turabian StyleSubasinghe Achchige, Yasika Medhavi, Liubov Volkova, and Christopher J. Weston. 2021. "Effect of Temperature and Exposure Time on Cambium Cell Viability In Vitro for Eucalyptus Species" Forests 12, no. 4: 445. https://0-doi-org.brum.beds.ac.uk/10.3390/f12040445
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.