
Genetic Diversity and Population Genetic Structure of Ancient Platycladus orientalis L. (Cupressaceae) in the Middle Reaches of the Yellow River by Chloroplast Microsatellite Markers


Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. DNA Extraction and cpSSR Analysis
2.3. Statistical Analysis
3. Results
3.1. Microsatellite Analysis
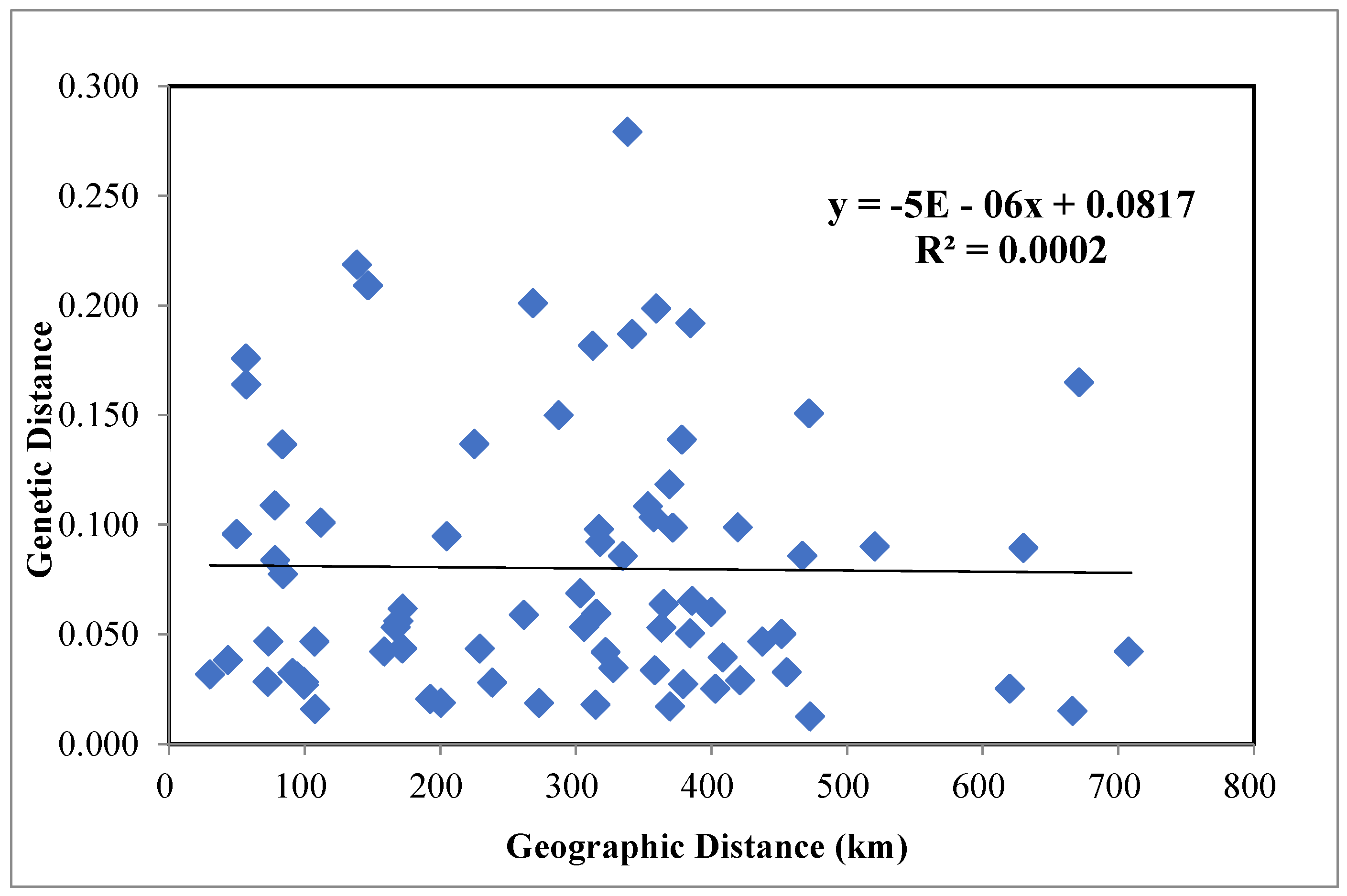
3.2. Genetic Diversity and Mantel Test Results in P. orientalis
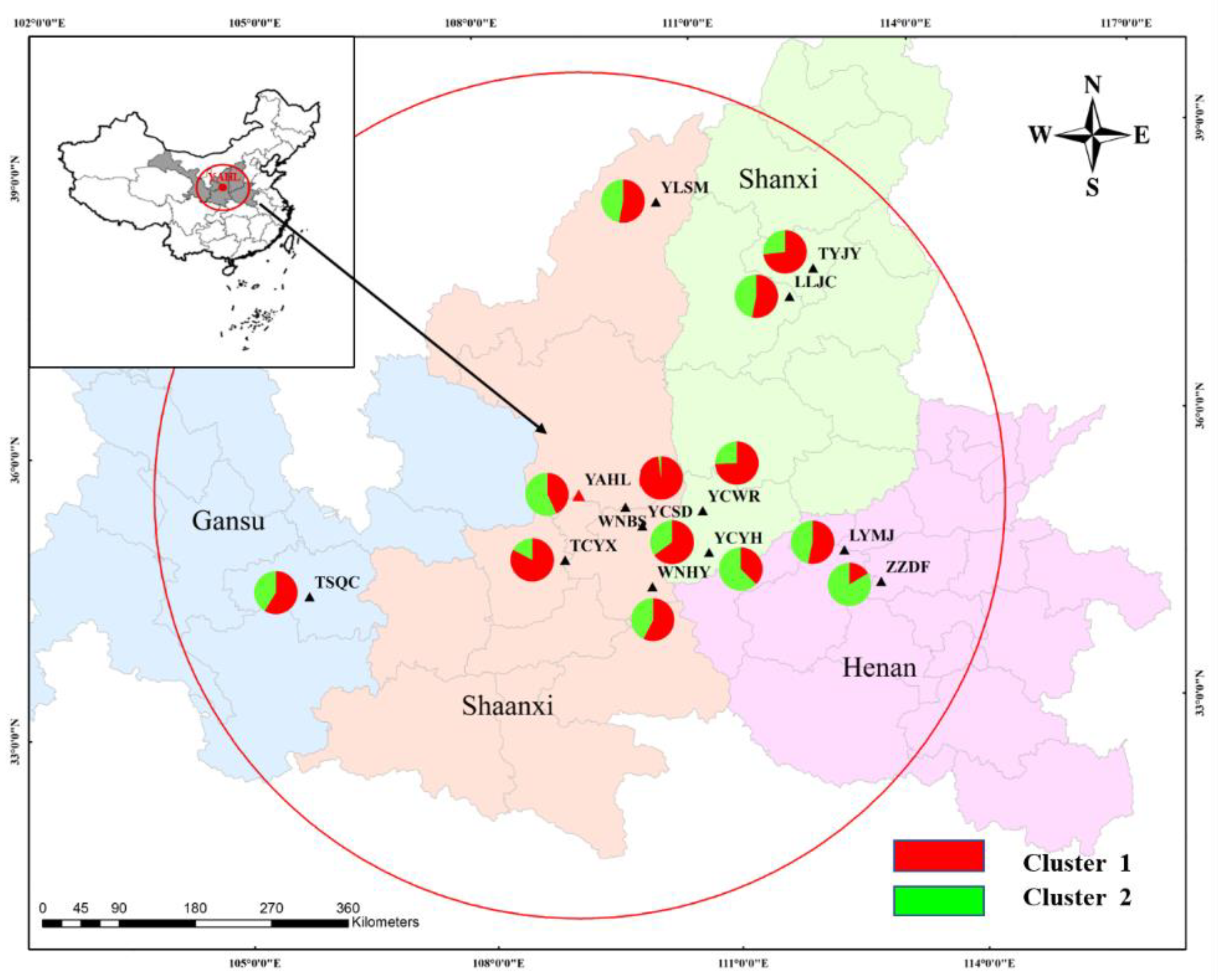
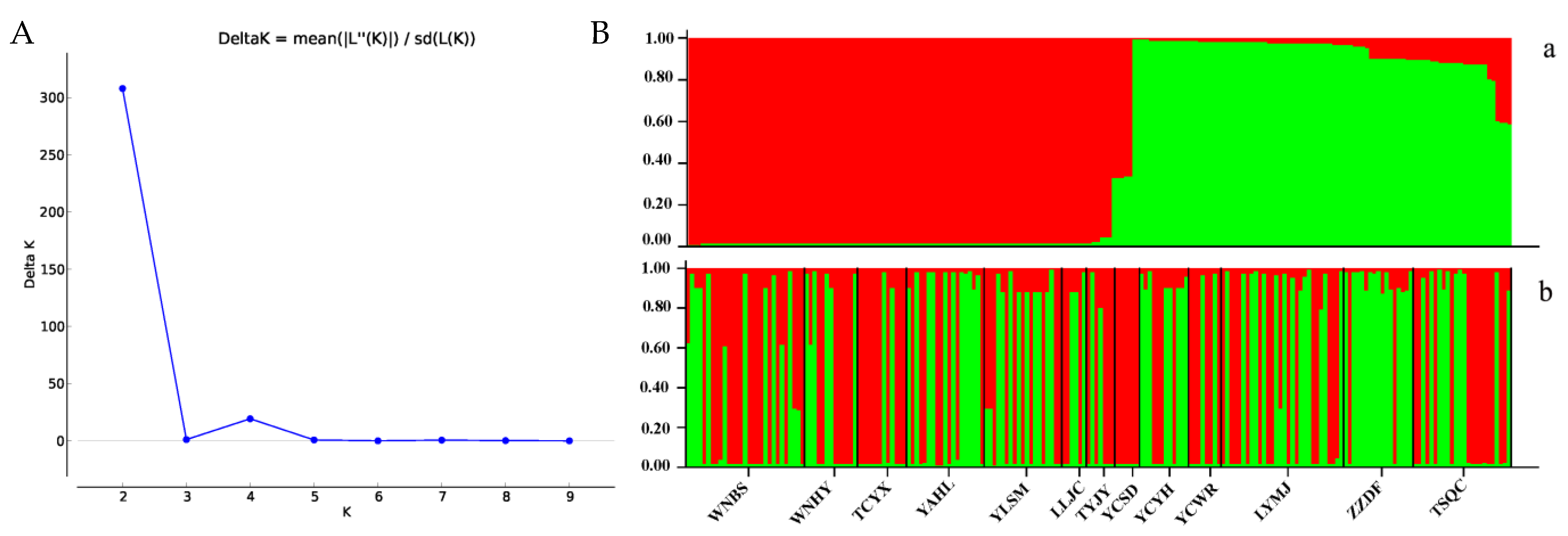
3.3. Population Clustering and Genetic Structure of P. orientalis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhou, Q.Y.; Jiang, Z.H.; Zhang, X.; Zhang, T.; Zhu, H.L.; Cui, B.; Li, Y.M.; Zhao, F.; Zhao, Z. Leaf anatomy and ultrastructure in senescing Ancient tree, Platycladus orientalis L. (Cupressaceae). PeerJ 2019, 7, e6766. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.M.; Yang, X.H.; Liu, J.L. Distribution and rejuvenation technology of ancient and famous tree in Beijing. Int. J. Med. Plant. Res. 2013, 26, 188. [Google Scholar] [CrossRef]
- Neale, D.B.; Kremer, A. Forest tree genomics: Growing resources and applications. Nat. Rev. Genet. 2011, 12, 111–122. [Google Scholar] [CrossRef]
- Seyed, M.H.; Seyed, A.S. Chemical constituents and toxicity of essential oils of Oriental Arborvitae, Platycladus orientalis (L.) Franco, against three stored-product beetles. Chil. J. Agric. Res. 2012, 72, 188–194. [Google Scholar]
- Wang, F.; Wu, D.J.; Yamamoto, H.; Xing, S.Y.; Zang, L.P. Digital image analysis of different crown shape of Platycladus orientalis. Ecol. Inform. 2016, 34, 146–152. [Google Scholar] [CrossRef]
- Hu, X.G.; Wang, T.L.; Liu, S.S.; Jiao, S.Q.; Jia, K.H.; Zhou, S.S.; Jin, Y.Q.; Li, Y.; Yousry, A.E.-K.; Mao, J.F. Predicting future seed sourcing of Platycladus orientalis (L.) for future climates using climate niche models. Forests 2017, 8, 471. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.Y.; Jiang, Z.H.; Li, Y.M.; Zhang, T.; Zhao, Z. Mesophyll protoplast isolation technique and flow cytometry analysis of ancient Platycladus orientalis (Cupressaceae). Turk. J. Agric. 2019, 43, 275–287. [Google Scholar] [CrossRef]
- Zhou, Q.Y.; Jiang, Z.H.; Zhang, X.; Lai, Q.; Zhao, Z. Tree age did not affect the leaf anatomical structure or ultrastructure of Platycladus orientalis L. (Cupressaceae). PeerJ 2019, 7, e7938. [Google Scholar] [CrossRef] [Green Version]
- Chang, E.M.; Tian, Y.X.; Wang, C.Y.; Deng, N.; Liu, C.X. Exploring the Phylogeography of Ancient Platycladus orientalis in China by Specific-Locus Amplified Fragment Sequencing. Int. J. Mol. Sci. 2019, 20, 3871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rosen, A.M.; Lee, J.; Li, M.; Wright, J.; Wright, H.T.; Fang, H. The Anthropocene and the landscape of Confucius: A historical ecology of landscape changes in northern and eastern China during the middle to late-Holocene. Holocene 2015, 25, 75–97. [Google Scholar] [CrossRef]
- Hua, S.Z.; Mou, J.Z. Harnessing and development of the Loess Plateau in the middle reaches of the Yellow River. In Taming the Yellow River: Silt and Floods: Proceedings of a Bilateral Seminar on Problems in the Lower Reaches of the Yellow River, China; Brush, L.M., Wolman, M.G., Bing-Wei, H., Eds.; Springer: Dordrecht, The Netherlands, 1989; pp. 517–528. [Google Scholar]
- Kai, L.; Wu, Z.-Y.; Raven, P.H. Flora of China 4. Cycadaceae through Fagaceae. Nord. J. Bot. 1999, 19, 580. [Google Scholar]
- Xie, C.Y.; Dancik, B.P.; Yeh, F.C. Genetic structure of Thuja orientalis. Biochem. Sys. Ecol. 1992, 20, 433–441. [Google Scholar] [CrossRef]
- Zhu, L.; Lou, A.R. Old-growth Platycladus orientalis as a resource for reproductive capacity and genetic diversity. PLoS ONE 2013, 8, e56489. [Google Scholar] [CrossRef] [PubMed]
- Martin, M.A.; Mattioni, C.; Lusini, I.; Molina, J.R.; Cherubini, M.; Drake, F.; Herrera, M.A.; Villani, F.; Martin, L.M. New insights into the genetic structure of Araucaria araucana forests based on molecular and historic evidences. Tree Genet. Genomes 2014, 10, 839–851. [Google Scholar] [CrossRef]
- Van Vuuren, B. Introduction to Conservation Genetics (Book); Cambridge University Press: Cambridge, UK, 2010. [Google Scholar]
- Cato, S.A.; Richardson, T.E. Inter- and intraspecific polymorphism at chloroplast SSR loci and the inheritance of plastids in Pinus radiata D. Don. Theor. Appl. Genet. 1996, 93, 587–592. [Google Scholar] [CrossRef] [PubMed]
- Morgante, M.; Pfeiffer, A.; Costacurta, A.; Olivieri, A.M. Molecular tools for population and ecological genetics in coniferous trees. Ann. Rei Bot. 1996, 36, 129–138. [Google Scholar]
- Cheng, Y.J.; Guo, W.W.; Deng, X.X. cpSSR: A new tool to analyse chloroplast genome of Citrus somatic hybrids. Acta. Bot. Sin. 2003, 45, 906–909. [Google Scholar]
- Rai, K.C.; Ginwal, H.S. Microsatellite analysis to study genetic diversity in Khasi Pine (Pinus Kesiya Royle Ex. Gordon) using chloroplast SSR markers. Silvae Genet. 2018, 67, 99–105. [Google Scholar] [CrossRef] [Green Version]
- Ebert, D.; Peakall, R. Chloroplast simple sequence repeats (cpSSRs): Technical resources and recommendations for expanding cpSSR discovery and applications to a wide array of plant species. Mol. Ecol. Resour. 2010, 9, 673–690. [Google Scholar] [CrossRef] [PubMed]
- Shota, S.; Tsumura, Y.; Crisp, M.D.; Bowman, D.M.J.S.; Isagi, Y. Genetic evidence for paternal inheritance of the chloroplast in four Australian Callitris species (Cupressaceae). J. For. Res. 2014, 19, 244–248. [Google Scholar]
- Vendramin, G.G.; Lelli, L.; Rossi, P.; Morgante, M. A set of primers for the amplification of 20 chloroplast microsatellites in Pinaceae. Mol. Ecol. 2010, 5, 595–598. [Google Scholar] [CrossRef]
- Wang, Y.S.; Xing, S.Y.; Tang, H.X.; Feng, D.Q. Genetic diversity of Platycladus orientalis provenances. Sci. Silvae Sin. 2011, 47, 91–96. [Google Scholar]
- Zhu, L.; Lou, A.R. Development and characterization of nine highly polymorphic microsatellite primers for Platycladus orientalis (Cupressaceae). Am. J. Bot. 2012, 99, e280–e282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, Y.Q.; Ma, Y.Q.; Wang, S.; Hu, X.G.; Huang, L.S.; Li, Y.; Wang, X.R.; Mao, J.F. Genetic evaluation of the breeding population of a valuable reforestation conifer Platycladus orientalis (Cupressaceae). Sci. Rep. 2016, 6, 34821. [Google Scholar] [CrossRef] [Green Version]
- Huang, L.S.; Sun, Y.; Jin, Y.Q.; Gao, Q.; Hu, X.G.; Gao, F.L.; Yang, X.L.; Zhu, J.; El-Kassaby, Y.A.; Mao, J.F. Development of high transferability cpSSR markers for individual identification and genetic investigation in Cupressaceae species. Ecol. Evol. 2018, 8, 4967–4977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, L.S.; Song, J.; Sun, Y.Q.; Gao, Q.; Jiao, S.Q.; Zhou, S.S.; Jin, Y.; Yang, X.L.; Zhu, J.J.; Gao, F.L. Pollination dynamics in a Platycladus orientalis seed orchard as revealed by partial pedigree reconstruction. Can. J. For. Res. 2018, 48, 952–957. [Google Scholar] [CrossRef]
- Richards, C.M.; Antolin, M.F.; Reilley, A.; Poole, J.; Walters, C. Capturing genetic diversity of wild populations for ex situ conservation: Texas wild rice (Zizania texana) as a model. Genet. Resour. Crop. Evol. 2007, 54, 837–848. [Google Scholar] [CrossRef]
- Yeh, F.C.; Yang, R.C.; Boyle, T. POPGENE Version 1.32 Microsoft Windows-Based Freeware for Populations Genetic Analysis; University of Alberta: Edmonton, AB, Canada, 1999. [Google Scholar]
- Peakall, R.; Smouse, P.E. genalex 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Goudet, J. FSTAT (version 1.2): A computer program to calculate Fstatistics. J. Hered. 1995, 86, 485–486. [Google Scholar] [CrossRef]
- Nagy, S.; Poczai, P.; Cernák, I.; Gorji, A.M.; Hegedűs, G.; Taller, J. PICcalc: An online program to calculate polymorphic information content for molecular genetic studies. Biochem. Genet. 2012, 50, 670–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raymond, M.; Rousset, F. GENEPOP (Version 1.2): Population genetics software for exact tests and ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Cristescu, R.; Sherwin, W.B.; Handasyde, K.; Cahill, V.; Cooper, D.W. Detecting bottlenecks using BOTTLENECK 1.2.02 in wild populations: The importance of the microsatellite structure. Conserv. Gene. 2010, 11, 1043–1049. [Google Scholar] [CrossRef]
- Excoffier, L.; Laval, G.; Schneider, S. Arlequin (version 3.0): An integrated software package for population genetics data analysis. Evol. Bioinf. Online 2005, 1, 47–50. [Google Scholar] [CrossRef] [Green Version]
- Legendre, P.; FORTIN, M.-J. Comparison of the Mantel test and alternative approaches for detecting complex multivariate relationships in the spatial analysis of genetic data. Mol. Ecol. Resour. 2010, 10, 831–844. [Google Scholar] [CrossRef]
- Naoko, T.; Masatoshi, N.; Koichiro, T. POPTREE2: Software for constructing population trees from allele frequency data and computing other population statistics with windows interface. Mol. Biol. Evol. 2010, 27, 747–752. [Google Scholar]
- Pritchard, J.; Mj, S.; Donnelly, P.J. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef] [PubMed]
- Earl, D.A.; Vonholdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Yang, A.; Li, Z.; Zhang, H.; Liu, L.; Wu, Z.; Li, Y.; Liu, T.; Xu, M.; Yu, F. Genetic diversity and population genetic structure of Cinnamomum camphora in South China revealed by EST-SSR markers. Forests 2019, 10, 1019. [Google Scholar] [CrossRef] [Green Version]
- Nowakowska, J.A.; Hsiang, T.; Patynek, P.; Stere´nczak, K.; Olejarski, I.; Oszako, T. Health Assessment and Genetic Structure of Monumental Norway Spruce Trees during A Bark Beetle (Ips typographus L.) Outbreak in the Białowie˙za Forest District, Poland. Forests 2020, 11, 647. [Google Scholar] [CrossRef]
- Frankham, R.; Ballou, J.D.; Briscoe, D.A. Introduction to conservation genetics. Genet. Res. 2002, 83, 1–617. [Google Scholar]
- Vanden-Broeck, A.; Gruwez, R.; Cox, K.; Adriaenssens, S.; Michalczyk, I.M.; Verheyen, K. Genetic structure and seed-mediated dispersal rates of an endangered shrub in a fragmented landscape: A case study for Juniperus communis in northwestern Europe. BMC Genet. 2011, 12, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.M.; Zhao, A.M.; Kang, X.Y.; Zhou, S.L.; López-Pujol, J. Genetic diversity, population structure, and conservation of Sophora moorcroftiana (Fabaceae), a shrub endemic to the Tibetan Plateau. Plant Biol. 2006, 8, 81–92. [Google Scholar] [CrossRef]
- Wang, H.Z.; Kang, J.Y.; Liu, J.J.; Qiu, S.; Chen, Z. Dendrobium SSR Markers play a good role in genetic diversity and phylogenetic analysis of Orchidaceae species. Sci. Hortic. 2015, 183, 160–166. [Google Scholar]
- Yue, X.Y.; Zheng, X.Y.; Zong, Y.; Jiang, S.; Hu, C.Y.; Yu, P.Y.; Liu, G.Q.; Cao, Y.F.; Hu, H.J.; Teng, Y.W. Combined analyses of chloroplast DNA haplotypes and microsatellite markers reveal new insights into the origin and dissemination route of cultivated pears native to East Asia. Front. Plant Sci. 2018, 9, 591. [Google Scholar] [CrossRef] [PubMed]
- Powell, W.; Morgante, M.; Andre, C.; Mcnicol, J.W.; Rafalski, J.A. Hypervariable microsatellites provide a general source of polymorphic DNA markers for the chloroplast genome. Curr. Biol. 1995, 5, 1023–1029. [Google Scholar] [CrossRef] [Green Version]
- Powell, W.; Machray, G.C.; Provan, J. Polymorphism revealed by simple sequence repeats. Trends Plant Sci. 1996, 1, 215–222. [Google Scholar] [CrossRef]
- Dow, B.D.; Ashley, M.V. High levels of gene flow in bur oak revealed by paternity analysis using microsatellites. J. Hered. 2000, 69, 62–70. [Google Scholar] [CrossRef] [Green Version]
- Gerber, S.; Mariette, S.; Streiff, R.; Bodenes, C.; Kremer, A. Comparison of microsatellites and amplified fragment length polymorphism markers for parentage analysis. Mol. Ecol. 2000, 9, 1037–1048. [Google Scholar] [CrossRef]
- Selkoe, K.A.; Toonen, R.J. Microsatellites for ecologists: A practical guide to using and evaluating microsatellite markers. Ecol. Lett. 2010, 9, 615–629. [Google Scholar] [CrossRef]
- Adelalu, K.F.; Zhang, X.; Qu, X.J.; Landis, J.B.; Wang, H. Plastome Phylogenomic and Biogeographical Study on Thuja (Cupressaceae). Biomed. Res. Int. 2020, 26, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Ferrão, L.F.V.; Caixeta, E.T.; Pena, G.; Zambolim, E.M.; Cruz, C.D.; Zambolim, L.; Ferrao, M.A.G.; Sakiyama, N.S. New EST–SSR markers of Coffea arabica: Transferability and application to studies of molecular characterization and genetic mapping. Mol. Breed. 2015, 35, 31. [Google Scholar] [CrossRef] [Green Version]
- Reisch, C.; Mayer, F.; Ruether, C.; Nelle, O. Forest history affects genetic diversity–molecular variation of Dryopteris dilatata (Dryopteridaceae) in ancient and recent forests. Nord. J. Bot. 2007, 25, 366–371. [Google Scholar] [CrossRef]
- Jacquemart, A.L.; Buyens, C.; Delescaille, L.M.; Rossum, F.V. Using genetic evaluation to guide conservation of remnant Juniperus communis (Cupressaceae) populations. Plant Biol. 2020, 23, 193–204. [Google Scholar] [CrossRef]
- Loveless, M.D.; Hamrick, J.L. Ecological determinants of genetic structure in plant populations. Annu. Rev. Ecol. Syst. 1984, 15, 65–95. [Google Scholar] [CrossRef]
- SJakob, S.; Burgi, L. Patterns of variation of a common fern (Athyrium filix-femina; Woodsiaceae): Population structure along and between altitudinal gradients. Am. J. Bot. 2007, 94, 965–971. [Google Scholar]
- Bradshaw, R.H.W. Past anthropogenic influence on European forests and some possible genetic consequences. For. Ecol. Manag. 2004, 197, 203–212. [Google Scholar] [CrossRef]
- Patino, J.; Werner, O.; González-Mancebo, J.M. The impact of forest disturbance on the genetic diversity and population structure of a late-successional moss. Trans. Br. Bryol. Soc. 2010, 32, 220–231. [Google Scholar] [CrossRef]
- Ellstrand, N.C.; Elam, D.R. Population genetic consequences of small population size: Implications for plant conservation. Annu. Rev. Ecol. Syst. 1993, 24, 217–242. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Populations | Sample Size | Locations | Latitude (N) | Longitude (E) | Elevation (m) | Average Age (years) | Na | Ne | AR | Apriv | I | TPM | SMM |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WNBS | 29 | Weinan, Shaanxi | 35°22′ | 109°41′ | 816 | 2314 | 2.750 | 1.436 | 2.123 | 0.250 | 0.510 | 0.290 | 0.253 |
| WNHY | 13 | Weinan, Shaanxi | 34°34′ | 110°06′ | 315 | 1518 | 2.250 | 1.611 | 2.180 | 0.000 | 0.546 | 0.562 | 0.384 |
| TCYX | 12 | Tongchuan, Shaanxi | 34°54′ | 108°59′ | 717 | 1238 | 1.750 | 1.148 | 1.571 | 0.250 | 0.213 | 0.241 | 0.217 |
| YAHL | 19 | Yan’an, Shaanxi | 35°35′ | 109°16′ | 865 | 2711 | 2.250 | 1.689 | 2.063 | 0.250 | 0.588 | 0.686 | 0.680 |
| YLSM | 19 | Yulin, Shaanxi | 38°40′ | 110°25′ | 1172 | 1668 | 2.750 | 1.583 | 2.294 | 0.250 | 0.610 | 0.229 | 0.200 |
| LLJC | 6 | Lvliang, Shanxi | 37°34′ | 112°07′ | 884 | 1200 | 1.500 | 1.346 | 1.500 | 0.000 | 0.286 | 0.739 | 0.715 |
| TYJY | 7 | Taiyuan, Shanxi | 37°42′ | 112°26′ | 816 | 1871 | 2.000 | 1.366 | 1.989 | 0.000 | 0.404 | 0.081 | 0.063 |
| YCSD | 6 | Yuncheng, Shanxi | 35°07′ | 110°54′ | 398 | 2417 | 1.250 | 1.096 | 1.250 | 0.000 | 0.113 | 0.516 | 0.477 |
| YCYH | 12 | Yuncheng, Shanxi | 34°54′ | 110°50′ | 325 | 1308 | 2.000 | 1.414 | 1.869 | 0.000 | 0.411 | 0.444 | 0.481 |
| YCWR | 8 | Yuncheng, Shanxi | 35°21′ | 110°48′ | 770 | 2250 | 1.750 | 1.321 | 1.725 | 0.000 | 0.324 | 0.661 | 0.713 |
| LYMJ | 30 | Luoyang, Henan | 34°50′ | 112°35′ | 124 | 1500 | 2.750 | 1.693 | 2.330 | 0.000 | 0.668 | 0.634 | 0.562 |
| ZZDF | 17 | Zhengzhou, Henan | 34°27′ | 113°04′ | 350 | 1435 | 2.500 | 1.858 | 2.275 | 0.000 | 0.659 | 0.658 | 0.629 |
| TSQC | 24 | Tianshui, Gansu | 34°34′ | 105°42′ | 1173 | 929 | 2.750 | 1.709 | 2.371 | 0.250 | 0.678 | 0.634 | 0.577 |
| Mean | 15.538 | - | - | - | - | 1720 | 2.173 | 1.482 | 1.964 | 0.096 | 0.462 | 0.490 | 0.457 |
| ID | Primer Sequence (5’–3’) | Repeat Motif | Expected PCR Product Size (bp) | Location | Tm (°C) | Species |
|---|---|---|---|---|---|---|
| N1 | TTCTAGCTCGCACCCAAACT TTGTTTCGCCGATATGTTCA | (AC)6 | 260 | rbcL/accD IR | 56 | Cupressus gigantean |
| N2 | TGGTCATACCATTGCTGTTCA TGGGCTACTCTACGTGCTTT | (AT)5 | 395 | rps19/rp122 IR | 56 | Cupressus gigantean |
| N6 | GGGAACAACCAGAATTGGAA GCCACTTTTATGGCACGACT | (TA)5 | 360 | ycf1 | 56 | Cupressus gigantean |
| N33 | CTGTTCCCCTGTGCATCATA AGGAGGAAAATCCGTTGGTT | (TCT)5 | 400 | trnF-GAA/trnL-UAA IR | 56 | Juniperus scopulorum |
| ID | Na | Ne | I | PIC | H | Fst | Nm |
|---|---|---|---|---|---|---|---|
| N1 | 4 | 1.833 | 0.793 | 0.397 | 0.455 | 0.210 | 0.938 |
| N2 | 6 | 1.5339 | 0.761 | 0.332 | 0.350 | 0.115 | 1.919 |
| N6 | 2 | 1.669 | 0.590 | 0.320 | 0.400 | 0.170 | 0.221 |
| N33 | 4 | 1.348 | 0.548 | 0.245 | 0.258 | 0.114 | 1.935 |
| Mean | 4 | 1.597 | 0.673 | 0.324 | 0.367 | 0.153 | 1.503 |
| Source of Variation | d.f. | Sum of Squares | Variance Components | Total Variation (%) | p Value |
|---|---|---|---|---|---|
| Among populations | 9 | 38.979 | 0.062 | 8 | <0.001 |
| Among individuals within populations | 189 | 256.635 | 0.679 | 92 | |
| Total | 403 | 295.614 | 0.741 | 100 |
| PC | WNBS | WNHY | TCYX | YAHL | YLSM | LLJC | TYJY | YCSD | YCYH | YCWR | LYMJ | ZZDF | TSQC |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| WNBS | - | 0.981 | 0.980 | 0.906 | 0.951 | 0.918 | 0.983 | 0.975 | 0.914 | 0.968 | 0.948 | 0.820 | 0.954 |
| WNHY | 0.027 | - | 0.962 | 0.902 | 0.938 | 0.918 | 0.982 | 0.959 | 0.870 | 0.971 | 0.967 | 0.825 | 0.951 |
| TCYX | 0.031 | 0.047 | - | 0.907 | 0.959 | 0.948 | 0.981 | 0.985 | 0.897 | 0.973 | 0.942 | 0.829 | 0.961 |
| YAHL | 0.176 | 0.219 | 0.084 | - | 0.925 | 0.959 | 0.909 | 0.860 | 0.910 | 0.897 | 0.904 | 0.948 | 0.945 |
| YLSM | 0.053 | 0.033 | 0.047 | 0.199 | - | 0.972 | 0.954 | 0.941 | 0.861 | 0.972 | 0.969 | 0.872 | 0.972 |
| LLJC | 0.060 | 0.027 | 0.040 | 0.187 | 0.021 | - | 0.940 | 0.914 | 0.849 | 0.957 | 0.943 | 0.912 | 0.966 |
| TYJY | 0.034 | 0.029 | 0.050 | 0.192 | 0.019 | 0.038 | - | 0.987 | 0.872 | 0.984 | 0.968 | 0.811 | 0.957 |
| YCSD | 0.032 | 0.028 | 0.028 | 0.137 | 0.051 | 0.042 | 0.064 | - | 0.848 | 0.975 | 0.937 | 0.756 | 0.934 |
| YCYH | 0.101 | 0.109 | 0.056 | 0.053 | 0.099 | 0.098 | 0.103 | 0.078 | - | 0.818 | 0.834 | 0.888 | 0.906 |
| YCWR | 0.033 | 0.016 | 0.044 | 0.209 | 0.017 | 0.019 | 0.018 | 0.047 | 0.096 | - | 0.973 | 0.804 | 0.954 |
| LYMJ | 0.059 | 0.044 | 0.035 | 0.092 | 0.086 | 0.054 | 0.086 | 0.028 | 0.042 | 0.062 | - | 0.839 | 0.970 |
| ZZDF | 0.182 | 0.201 | 0.099 | 0.118 | 0.090 | 0.108 | 0.139 | 0.150 | 0.095 | 0.137 | 0.164 | - | 0.920 |
| TSQC | 0.065 | 0.025 | 0.069 | 0.279 | 0.025 | 0.015 | 0.042 | 0.060 | 0.151 | 0.013 | 0.090 | 0.165 | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, B.; Deng, P.; Zhang, S.; Zhao, Z. Genetic Diversity and Population Genetic Structure of Ancient Platycladus orientalis L. (Cupressaceae) in the Middle Reaches of the Yellow River by Chloroplast Microsatellite Markers. Forests 2021, 12, 592. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050592
Cui B, Deng P, Zhang S, Zhao Z. Genetic Diversity and Population Genetic Structure of Ancient Platycladus orientalis L. (Cupressaceae) in the Middle Reaches of the Yellow River by Chloroplast Microsatellite Markers. Forests. 2021; 12(5):592. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050592
Chicago/Turabian StyleCui, Bei, Ping Deng, Sheng Zhang, and Zhong Zhao. 2021. "Genetic Diversity and Population Genetic Structure of Ancient Platycladus orientalis L. (Cupressaceae) in the Middle Reaches of the Yellow River by Chloroplast Microsatellite Markers" Forests 12, no. 5: 592. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050592