
Spatial Distribution and Structural Characteristics for Haloxylon ammodendron Plantation on the Southwestern Edge of the Gurbantünggüt Desert


 ,
,
Abstract
:1. Introduction
2. Materials and Methods
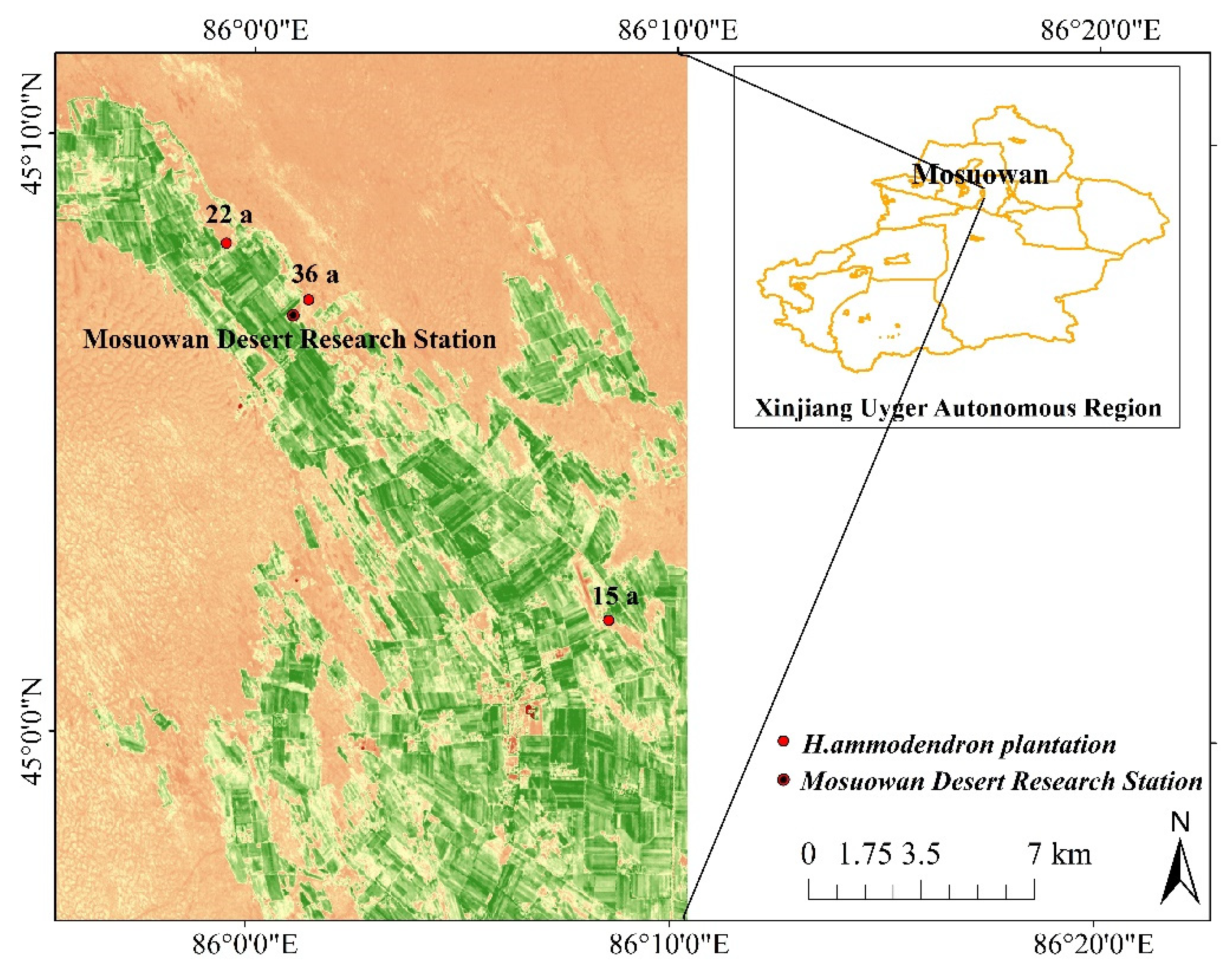
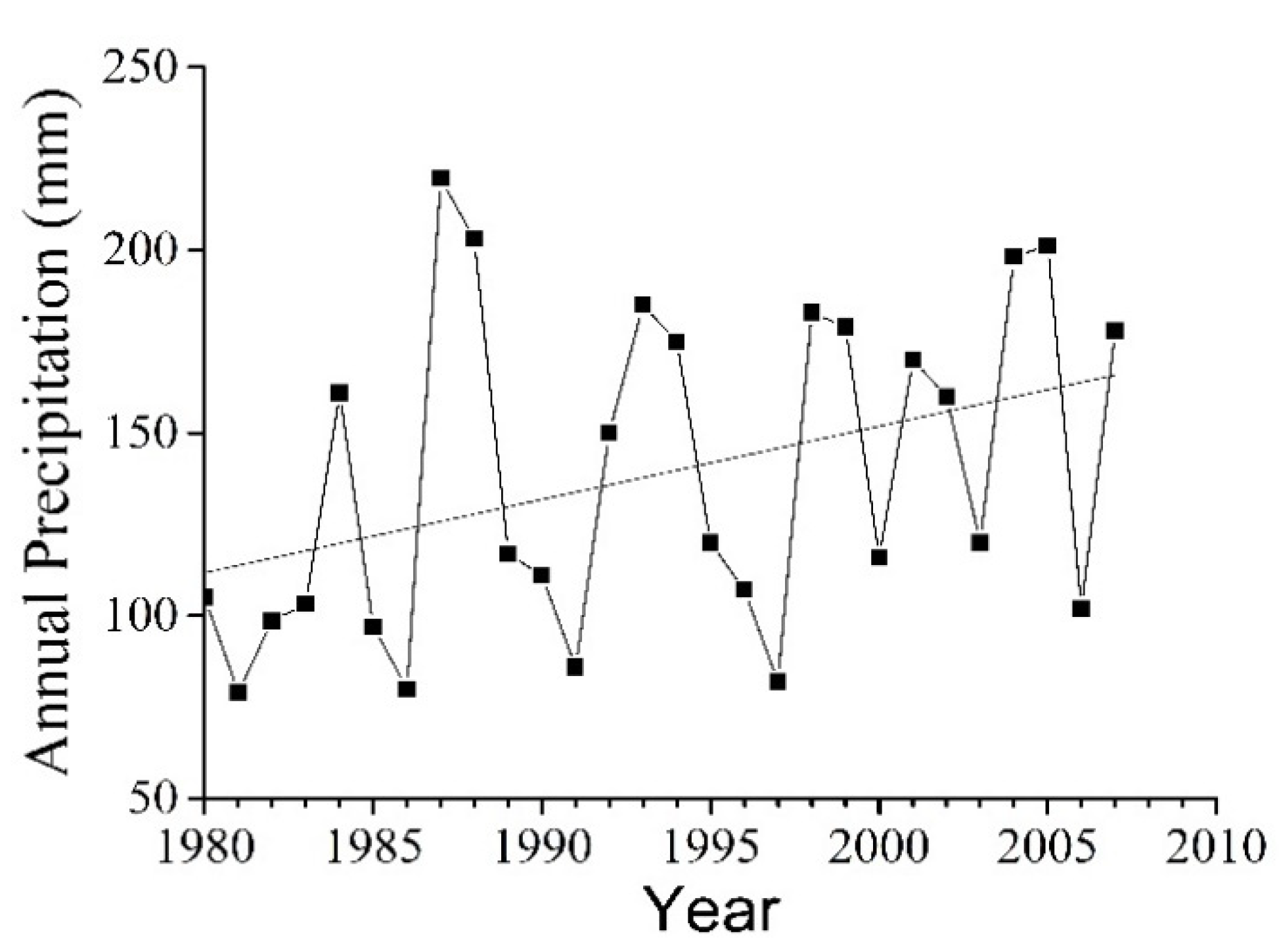
2.1. Study Area
2.2. Field Investigation
2.3. Data Analysis
3. Results
3.1. Basic Characteristics of Haloxylon ammodendron Plantations
3.2. Structural Characteristics of the Main Forest Layer
3.3. Structural Characteristics of the Regeneration Layer
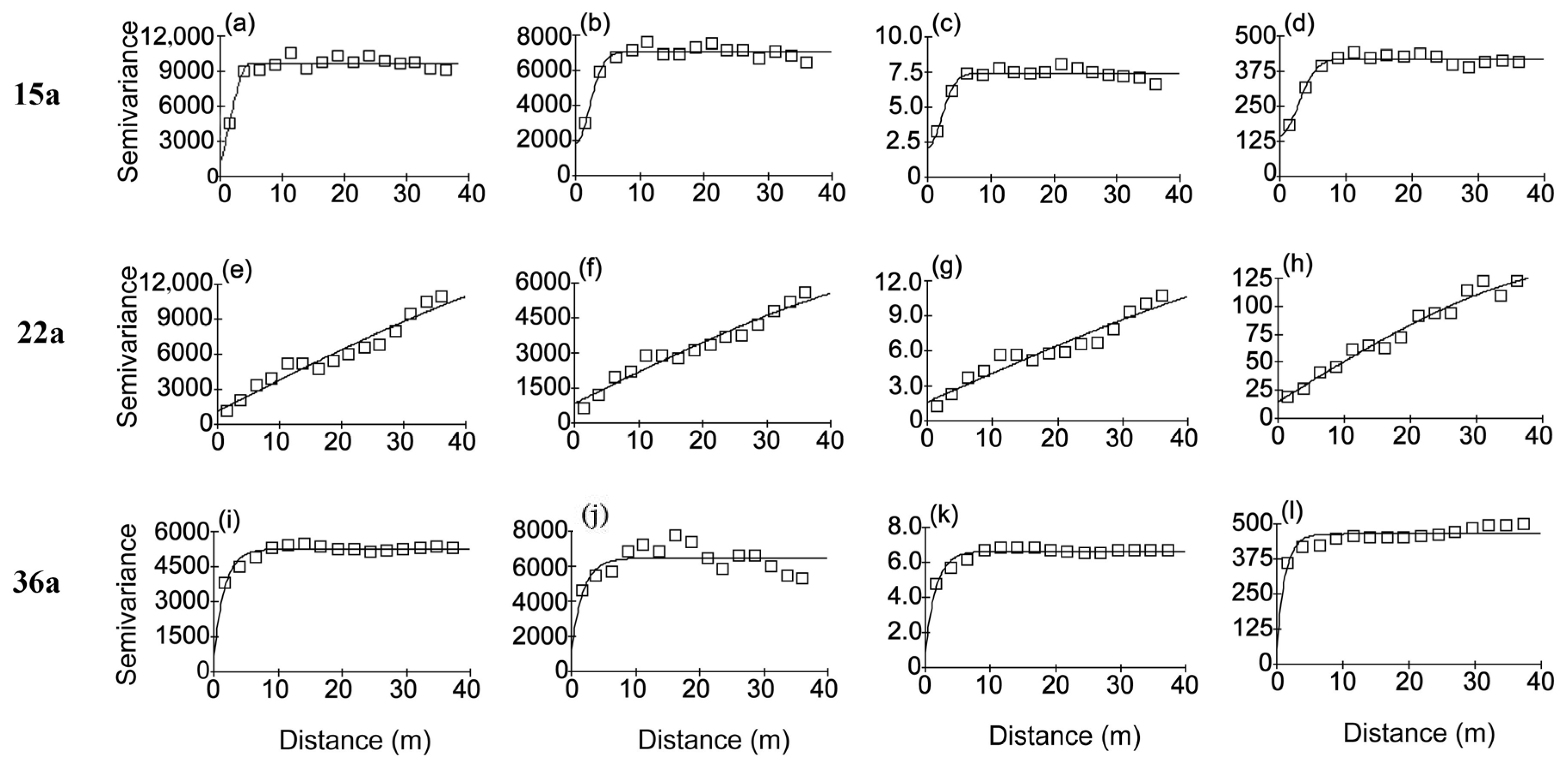
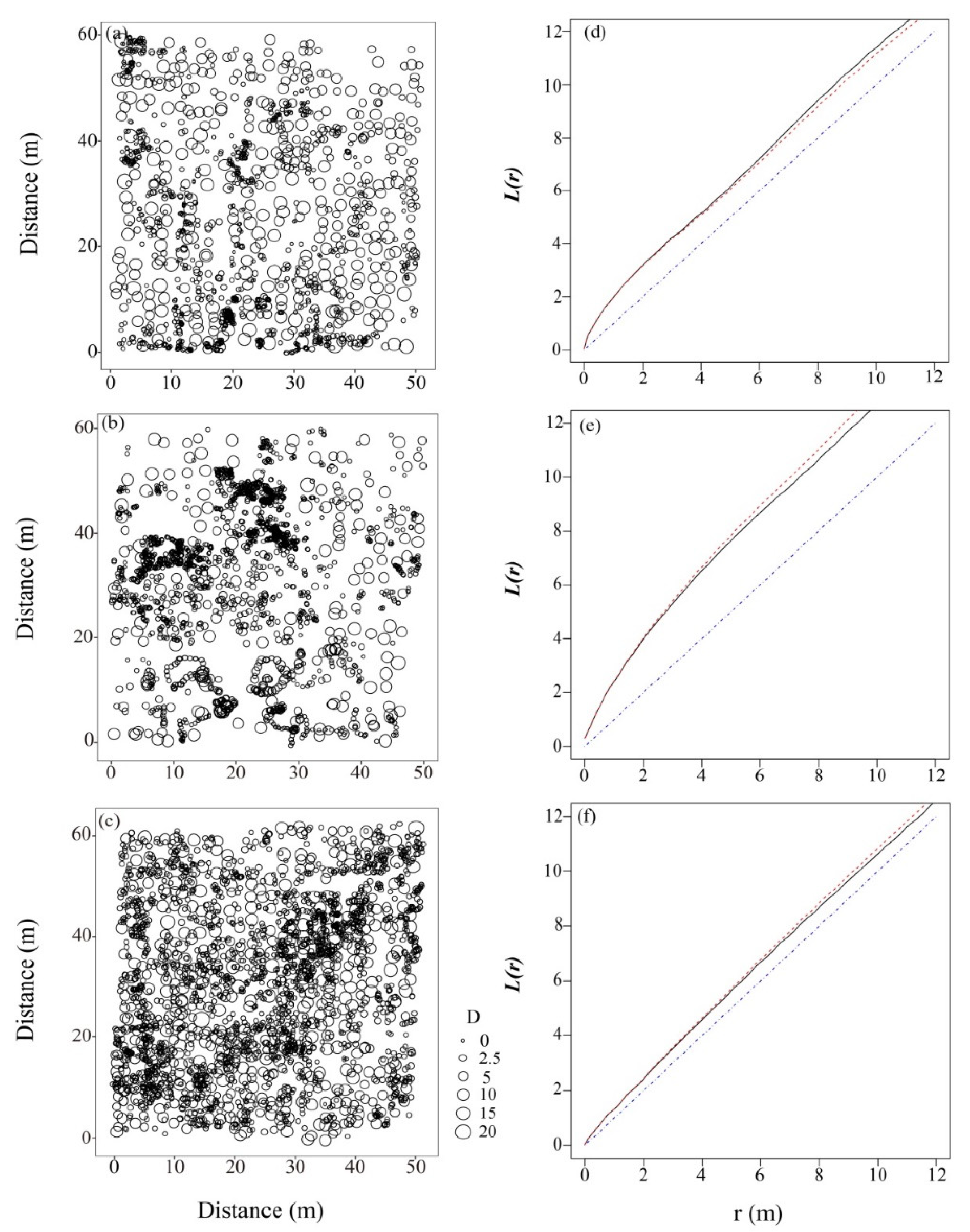
3.4. Spatial Patterns of Haloxylon ammodendron Individuals at Different Forest Ages
4. Discussion
4.1. Growth Dynamics of the Haloxylon ammodendron Plantations
4.2. Spatial Patterns of the Haloxylon ammodendron Plantations at Different Stand Ages
4.3. Main Factors Affecting the Spatial Patterns
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Adeel, Z. Findings of the global desertification assessment by the Millennium Ecosystem Assessment—A perspective for better managing scientific knowledge. In The Future of Drylands; Lee, C., Schaaf, T., Eds.; Springer: Dordrecht, The Netherlands, 2008; pp. 677–685. [Google Scholar]
- Hufkens, K.; Scheunders, P.; Ceulemans, R. Ecotones in vegetation ecology: Methodologies and definitions revisited. Ecol. Res. 2009, 24, 977–986. [Google Scholar] [CrossRef]
- Wang, X.Q.; Zhao, C.J. Sand surface change and natural species entrance in straw barrier system in Gurbantunggut Desert, Xinjiang, China. Arid Land Geogr. 2002, 25, 201–207. (In Chinese) [Google Scholar]
- Xu, J.; Gu, H.; Meng, Q.; Cheng, J.; Liu, Y.; Jiang, P.; Sheng, J.; Deng, J.; Bai, X. Spatial pattern analysis of Haloxylon ammodendron using UAV imagery—A case study in the Gurbantunggut Desert. Int. J. Appl. Earth Obs. Geoinf. 2019, 83, 101891. [Google Scholar] [CrossRef]
- Huang, P.Z.; Liu, Z.J.; Cui, W.C. A preliminary study on water harvesting afforestation of Haloxylon ammodendron. Xinjiang Agric. Sci. 1985, 6, 23–25. (In Chinese) [Google Scholar]
- Li, C.J.; Lei, J.Q.; Zhao, Y. Effect of saline water irrigation on soil development and plant growth in the Takllimakan Desert Highway Shelterbelt. Soil. Till. Res. 2015, 146, 99–107. [Google Scholar] [CrossRef]
- Zhou, Z.B.; Xu, X.W. Absorption characteristics of ions in three species of shrubs of the artificially-planted greenbelts in the hinterland of the Taklamakan Desert. Arid Zone Res. 2002, 19, 49–52. (In Chinese) [Google Scholar]
- Li, C.; Shi, X.; Mohamad, O.A.; Gao, J.; Xu, X.; Xie, Y. Moderate irrigation intervals facilitate establishment of two desert shrubs in the Taklimakan Desert Highway Shelterbelt. PLoS ONE 2017, 12, e0180875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.Q.; Wang, T.; Jiang, J. On the sand surface stability in the southern part of the Gurbantunggut Desert. Sci. China Ser. D Earth Sci. 2005, 48, 778–785. [Google Scholar] [CrossRef]
- Xiao, D.Z.; Hu, Y.K. A preliminary study on the runoff and its utilization of the crevasse crack ground (A case study of Mo-suowan pilot zone). Arid Land Geogr. 1985, 8, 28–34. (In Chinese) [Google Scholar]
- Li, Y.F.; Yang, G. Studies on the moisture Balance of Haloxylon ammodendron sand-break Forest: The Moisture State of Haloxylon ammodendron autumn irrigated sand-break forest. Arid Zone Res. 1996, 13, 57–62. (In Chinese) [Google Scholar]
- Chen, Q.M.; Luo, Q.H.; Ning, H.S.; Zhao, C.Y.; Duan, W.B. Characteristics of main stand layer and regeneration layer of Haloxylon ammodendron plantations at different ages on the southern edge of the Gurbantunggut Desert, Northwest China. Chin. J. Appl. Ecol. 2017, 28, 739–747. [Google Scholar]
- Greig-Smith, P. Pattern in Vegetation. J. Ecol. 1979, 67, 755. [Google Scholar] [CrossRef]
- Bagchi, R.; Henrys, P.A.; Brown, P.E.; Burslem, D.F.R.P.; Diggle, P.; Gunatilleke, C.V.S.; Gunatilleke, I.A.U.N.; Kassim, A.R.; Law, R.; Noor, S.; et al. Spatial patterns reveal negative density dependence and habitat associations in tropical trees. Ecology 2011, 92, 1723–1729. [Google Scholar] [CrossRef] [PubMed]
- Velázquez, E.; Paine, C.E.T.; May, F.; Wiegand, T. Linking trait similarity to interspecific spatial associations in a moist tropical forest. J. Veg. Sci. 2015, 26, 1068–1079. [Google Scholar] [CrossRef]
- Zhang, J.T. Analysis of spatial point pattern for plant species. Acta Phytoecol. Sin. 1998, 22, 344–349. [Google Scholar]
- Li, C.-J.; Lei, J.-Q.; Shi, X.; Liu, R. Scale Dependence of Soil Spatial Variation in a Temperate Desert. Pedosphere 2014, 24, 417–426. [Google Scholar] [CrossRef]
- Couteron, P.; Kokou, K. Woody vegetation spatial patterns in a semi-arid savanna of Burkina Faso, West Africa. Plant Ecol. 1997, 132, 211–227. [Google Scholar] [CrossRef]
- Barot, S.; Gignoux, J.; Menaut, J. Demography of a savanna palm tree: Predictions from comprehensive spatial pattern analyses. Ecology 1999, 80, 1987–2005. [Google Scholar] [CrossRef]
- Stoll, P.; Bergius, E. Pattern and process: Competition causes regular spacing of individuals within plant populations. J. Ecol. 2005, 93, 395–403. [Google Scholar] [CrossRef]
- Zheng, Y.; Zhao, W.Z.; Zhang, G.F. Spatial analysis of a Haloxylon ammodendron plantation in an oasis-desert ecotone in the Hexi corridor, northwestern China. Forests 2017, 8, 200. [Google Scholar] [CrossRef] [Green Version]
- Malkinson, D.; Tielbörger, K. What does the stress-gradient hypothesis predict? Resolving the discrepancies. Oikos 2010, 119, 1546–1552. [Google Scholar] [CrossRef]
- Hao, H.-M.; Huang, Z.; Lu, R.; Jia, C.; Liu, Y.; Liu, B.-R.; Wu, G.-L. Patches structure succession based on spatial point pattern features in semi-arid ecosystems of the water-wind erosion crisscross region. Glob. Ecol. Conserv. 2017, 12, 158–165. [Google Scholar] [CrossRef]
- Gross, K. Positive interactions among competitors can produce species-rich communities. Ecol. Lett. 2008, 11, 929–936. [Google Scholar] [CrossRef] [PubMed]
- Konings, A.G.; Dekker, S.C.; Rietkerk, M.; Katul, G.G. Drought sensitivity of patterned vegetation determined by rainfall-Land surface feedbacks. J. Geophys. Res. 2011, 116, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Bestelmeyer, B.T.; Duniway, M.C.; James, D.K.; Burkett, L.M.; Havstad, K.M. A test of critical thresholds and their indicators in a desertification-prone ecosystem: More resilience than we thought. Ecol. Lett. 2012, 16, 339–345. [Google Scholar] [CrossRef]
- Kulasova, A.; Blazkova, S.; Beven, K.; Řezáčová, D.; Cajthaml, J. Vegetation pattern as an indicator of saturated areas in a Czech headwater catchment. Hydrol. Process. 2014, 28, 5297–5308. [Google Scholar] [CrossRef]
- Schleicher, J.; Wiegand, K.; Ward, D. Changes of woody plant interaction and spatial distribution between rocky and sandy soil areas in a semi-arid savanna, South Africa. J. Arid. Environ. 2011, 75, 270–278. [Google Scholar] [CrossRef]
- Song, Y.Y.; Li, Y.Y.; Zhang, W.H. Analysis of spatial pattern and spatial association of Haloxylon ammodendron population in different developmental stages. Acta Ecol. Sin. 2010, 30, 4317–4327. (In Chinese) [Google Scholar]
- Browning, D.M.; Franklin, J.; Archer, S.R.; Gillan, J.K.; Guertin, D.P. Spatial patterns of grassland–shrubland state transitions: A 74-year record on grazed and protected areas. Ecol. Appl. 2014, 24, 1421–1433. [Google Scholar] [CrossRef]
- Li, Y.; Xu, H. Water and Carbon banlances of Haloxylon ammodendron: Integrated study and physiological, plant abd community level. Arid Land Geogr. 2008, 31, 313–323. [Google Scholar]
- Field, C.B.; Barros, V.R.; Dokken, D.J. Intergovernmental Panel on Climate Change. Summary for Policymakers//Climate Change 2014-Impacts, Adaptation and Vulnerability. Part A: Global and Sectoral Aspects; Cambridge University Press: Cambridge, UK, 2014; pp. 1–32. [Google Scholar]
- Wang, X.Q.; Zhang, Y.M.; Jiang, J. Variation pattern of soil water content in longitudinal dune in the southern part of Gurbantünggüt desert: How snow melt and frozen soil change affect the soil moisture. J. Glaciol. Geocryol. 2006, 28, 262–268. (In Chinese) [Google Scholar]
- Dai, Y.; Zheng, X.-J.; Tang, L.-S.; Li, Y. Stable oxygen isotopes reveal distinct water use patterns of two Haloxylon species in the Gurbantonggut Desert. Plant Soil 2015, 389, 73–87. [Google Scholar] [CrossRef]
- Cheng, C.D.; Zhang, L.Y.; Hu, W.K. The basic characteristics of plant communities, flora and their distribution in the sandy district of Gurbantungut. Acta Phytoecol. Geobot. Sin. 1983, 7, 89–98. (In Chinese) [Google Scholar]
- Ning, H.S.; Luo, Q.H.; Ji, X.M.; Lei, C.Y. Assessment on ecosystem service value of Haloxylon ammodendron forest in Xinjiang. Ecol. Sci. 2017, 36, 74–81. (In Chinese) [Google Scholar]
- Luo, Q.H.; Chen, Q.M.; Ning, H.S.; Zhao, C.Y. Chronosequence-based population structure and natural regeneration of Haloxylon ammodendron plantation in the southern edge of the Gurbantunggut Desert, Northwestern China. Russ. J. Ecol. 2017, 48, 364–371. [Google Scholar] [CrossRef]
- Shao, Y.Y.; Zhang, Y.Q.; Wu, X.Q.; Charles, P.A.; Zhang, J.T.; Qin, S.G.; Wu, B. Relating historical vegetation cover to aridity patterns in the greater desert region of northern China: Implications to planned and existing restoration projects. Ecol. Indic. 2018, 89, 528–537. [Google Scholar] [CrossRef]
- Chen, F.L.; Zheng, X.R.; He, X.L.; Yang, G.; Liu, B. Change of groundwater depth and impact factors in Mosuowan irrigation district during 1998–2007. Eng. J. Wuhan Univ. 2011, 44, 317–320. (In Chinese) [Google Scholar]
- Jiang, L. Analysis on groundwater characteristics at Mosuowan irrigation district of the Manas river basin. Ground water 2016, 38, 1–3. (In Chinese) [Google Scholar]
- Li, C.J.; Li, Y.; Ma, J. Spatial heterogeneity of soil chemical properties at fine scales induced by Haloxylon ammodendron (Chenopodiaceae) plants in a sandy desert. Ecol. Res. 2011, 26, 385–394. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, Y.; Xie, J.-B. Fixed allocation patterns, rather than plasticity, benefit recruitment and recovery from drought in seedlings of a desert shrub. AoB Plants 2016, 8, 1–11. [Google Scholar] [CrossRef]
- Dale, M.R.T.; Fortin, M.J. Spatial Analysis: A Guide for Ecologists, 2nd ed.; Cambridge University Press: Cambridge, UK, 2014; p. 382. [Google Scholar]
- Zuo, X.A.; Zhao, X.Y.; Zhao, H.L. Spatial heterogeneity of vegetation characteristics in the processes of degraded vegetation restoration in Horqin Sandy Land, northern China. Ecol. Environ. Sci. 2010, 19, 1513–1518. (In Chinese) [Google Scholar]
- Rundel, P.W. Community Structure and Stability in the Giant Sequoia Groves of the Sierra Nevada, California. Am. Midl. Nat. 1971, 85, 478. [Google Scholar] [CrossRef]
- Jia, Z.Q.; Lu, Q. Haloxylon Ammodendron; China Environmental Science Press: Beijing, China, 2005. [Google Scholar]
- Pillay, T.; Ward, D. Spatial pattern analysis and competition between Acacia karroo trees in humid savannas. Plant Ecol. 2012, 213, 1609–1619. [Google Scholar] [CrossRef]
- Diggle, P.J. Statistical Analysis of Spatial and Spatio-Temporal Point Patternss, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2013; pp. 1–5. [Google Scholar]
- Ramon, P.; De La Cruz, M.; Zavala, I.; Zavala, M.A. Factors influencing the dispersion ofArceuthobium oxycedriin Central Spain: Evaluation with a new null model for marked point patterns. For. Pathol. 2016, 46, 610–621. [Google Scholar] [CrossRef] [Green Version]
- Besag, J. Contribution to the discussion of Dr. Ripley’s paper. J. R. Stat. Soc. Ser. B 1977, 39, 193–195. [Google Scholar]
- Ripley, B.D. Modelling Spatial Patterns. J. R. Stat. Soc. Ser. B 1977, 39, 172–192. [Google Scholar] [CrossRef]
- Goreaud, F.; Pélissier, R. On explicit formulas of edge effect correction for Ripley’s K-function. J. Veg. Sci. 1999, 10, 433–438. [Google Scholar] [CrossRef]
- Wiegand, T.; Moloney, K.A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos 2004, 104, 209–229. [Google Scholar] [CrossRef]
- Duncan, R.P.; Stewart, G.H. The temporal and spatial analysis of tree age distributions. Can. J. For. Res. 1991, 21, 1703–1710. [Google Scholar] [CrossRef]
- Baddeley, A.; Turner, R.; Hazelton, J.M. Residual analysis for spatial point processes. J. R. Stat. Soc. 2005, 67, 617–666. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zheng, X.-J.; Li, Y.; Xu, G.-Q. Varying responses of two Haloxylon species to extreme drought and groundwater depth. Environ. Exp. Bot. 2019, 158, 63–72. [Google Scholar] [CrossRef]
- Li, J.J.; Xue, B.R.; Chai, Z.Z. Quantity dynamics and distribution pattern for Haloxylon ammodendron population of the desert-oasis ecotone in Minqin county of Gansu province, china. J. Northeast For. Univ. 2013, 41, 27–31. [Google Scholar]
- Wu, X.; Zheng, X.-J.; Yin, X.-W.; Yue, Y.-M.; Liu, R.; Xu, G.-Q.; Li, Y. Seasonal variation in the groundwater dependency of two dominant woody species in a desert region of Central Asia. Plant Soil 2019, 444, 39–55. [Google Scholar] [CrossRef]
- Zheng, X.J.; Li, Y.; Wu, X.; Xu, G.Q. Deepening Rooting Depths Improve Plant Water and Carbon Status of A Xeric Tree during Summer Drough. Forests 2019, 10, 592. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Xie, J.B.; Li, Y. Effects of increasing root carbon investment on the mortality and resprouting of Haloxylon ammodendron seedlings under drought. Plant Biol. 2016, 19, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Ponge, J.F.; Andre, J.; Zackrisson, O.; Bernier, N.; Nilsson, M.C.; Gallet, C. The forest regeneration puzzle. BioScience 1998, 48, 523–530. [Google Scholar]
- Tirado, R.; Pugnaire, F.I. Shrub spatial aggregation and consequences for reproductive success. Oecologia 2003, 136, 296–301. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.B.; Song, Y.Y. Influence of different longitudinal dune positions in the Gurbantunggut Desert on the reproduction of Haloxylon ammodendron seedlings. Appl. Ecol. Environ. Res. 2015, 13, 99–113. (In Chinese) [Google Scholar]
- Halik, Ü.; Aishan, T.; Betz, F.; Kurban, A.; Rouzi, A. Effectiveness and challenges of ecological engineering for desert riparian forest restoration along China’s largest inland river. Ecol. Eng. 2019, 127, 11–22. [Google Scholar] [CrossRef]
- Song, C.W.; Jiang, J.; Fu, H.F.; Chen, J.J. Soil moisture in plantation area and its affecting factors in the Gurbantonggut desert. Arid Land Geogr. 2009, 32, 704–710. (In Chinese) [Google Scholar]
- Xu, H.; Li, Y.; Xu, G.; Zou, T. Ecophysiological response and morphological adjustment of two Central Asian desert shrubs towards variation in summer precipitation. Plant Cell Environ. 2007, 30, 399–409. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.-Q.; McDowell, N.G.; Li, Y. A possible link between life and death of a xeric tree in desert. J. Plant Physiol. 2016, 194, 35–44. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | 15a | 22a | 36a |
|---|---|---|---|
| Initial Column and Row Spacing (m): 1 × 3 | Initial Column and Row Spacing (m): 1 × 3 | Initial Column and Row Spacing (m): 1 × 3 | |
| Number of current individuals | 304 | 177 | 559 |
| Density/individual (m2) | 0.104 | 0.059 | 0.186 |
| Survival rate (%) | 30.4 | 17.7 | 55.9 |
| Height (cm) | 235.49 ± 78.91 | 289.44 ± 91.79 | 200.66 ± 52.32 |
| Crown width (m2) | 3.41 ± 0.61 | 4.51 ± 0.69 | 2.59 ± 0.39 |
| Base stem diameter (cm) | 5.85 ± 2.61 | 8.77 ± 3.42 | 6.17 ± 2.46 |
| Deadwood proportion (%) | 29.70 ± 25.65 | 15.65 ± 15.76 | 41.15 ± 23.79 |
| Traits | 15a | 22a | 36a | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Seedling | Juvenile | Mature | Seedling | Juvenile | Mature | Seedling | Juvenile | Mature | |
| Survival | 946 | 22 | / | 1952 | 170 | 14 | 2227 | 21 | / |
| Frequency (%) | 97.73 | 2.27 | 0 | 91.39 | 7.96 | 0.65 | 99.07 | 0.93 | 0 |
| Density (m2) | 0.315 | 0.007 | / | 0.651 | 0.057 | 0.005 | 0.742 | 0.007 | |
| Height (cm) | 20.79 ± 12.11 | 91.36 ± 27.18 | / | 21.29 ± 15.86 | 142.79 ± 50.11 | 211.07 ± 36.38 | 32.14 ± 13.56 | 73.52 ± 30.36 | / |
| Crown width (m2) | 0.015 ± 0.015 | 0.47 ± 0.06 | / | 0.013 ± 0.014 | 0.64 ± 0.13 | 0.94 ± 0.16 | 0.039 ± 0.014 | 0.19 ± 0.049 | / |
| Base diameter (cm) | 0.28 ± 0.21 | 1.42 ± 0.34 | / | 0.27 ± 0.19 | 2.58 ± 1.42 | 10.04 ± 3.74 | 0.39 ± 0.18 | 1.67 ± 0.52 | / |
| Number of Individual Trees | 15a | 22a | 36a |
|---|---|---|---|
| Main forest layer | 304 | 177 | 559 |
| Regeneration layer (Juvenile) | 22 | 170 | 21 |
| Regeneration layer (Mature) | 0 | 14 | 0 |
| Total in plot | 326 | 354 | 580 |
| Initial plant density (plant ha−1) | 3333 | 3333 | 3333 |
| Preservation Density (plant ha−1) | 1086 | 1180 | 1933 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Song, C.; Li, C.; Halik, Ü.; Xu, X.; Lei, J.; Zhou, Z.; Fan, J. Spatial Distribution and Structural Characteristics for Haloxylon ammodendron Plantation on the Southwestern Edge of the Gurbantünggüt Desert. Forests 2021, 12, 633. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050633
Song C, Li C, Halik Ü, Xu X, Lei J, Zhou Z, Fan J. Spatial Distribution and Structural Characteristics for Haloxylon ammodendron Plantation on the Southwestern Edge of the Gurbantünggüt Desert. Forests. 2021; 12(5):633. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050633
Chicago/Turabian StyleSong, Chunwu, Congjuan Li, Ümüt Halik, Xinwen Xu, Jiaqiang Lei, Zhibin Zhou, and Jinglong Fan. 2021. "Spatial Distribution and Structural Characteristics for Haloxylon ammodendron Plantation on the Southwestern Edge of the Gurbantünggüt Desert" Forests 12, no. 5: 633. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050633