
Variations in Wood Density, Annual Ring Width and Vessel Properties of Quercus brantii Affected by Crown Dieback


 and
and
Abstract
:1. Introduction
2. Material and Methods
2.1. Study Areas
2.2. Sampling Method
2.3. Wood Density
2.4. Preparation for Macroscopic Measurement
2.5. Sample Preparation for Sectioning
2.6. Sectioning
2.7. Statistical Analysis
3. Results
3.1. Physical-Anatomical Variables
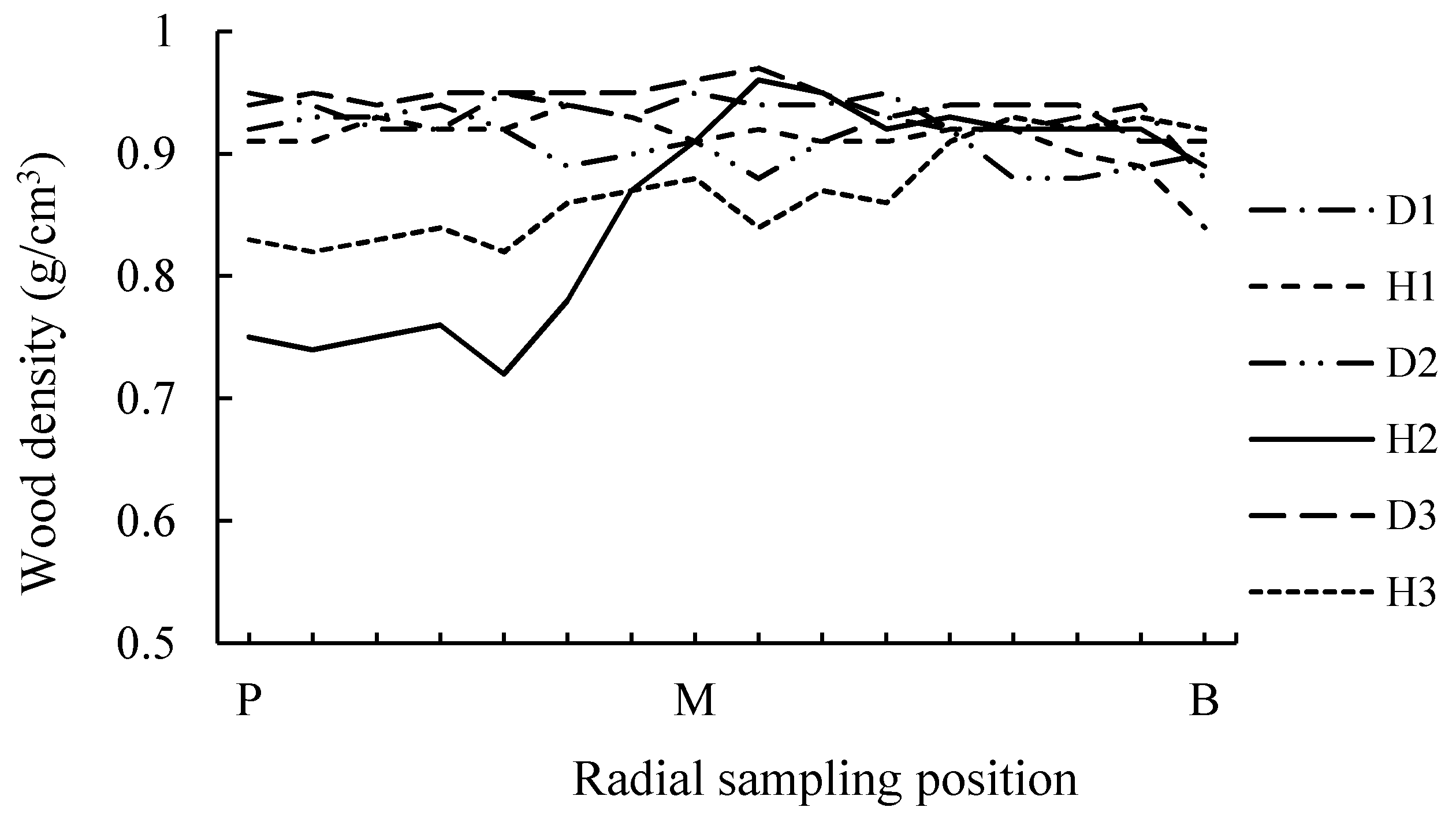
3.1.1. Wood Density (WD)
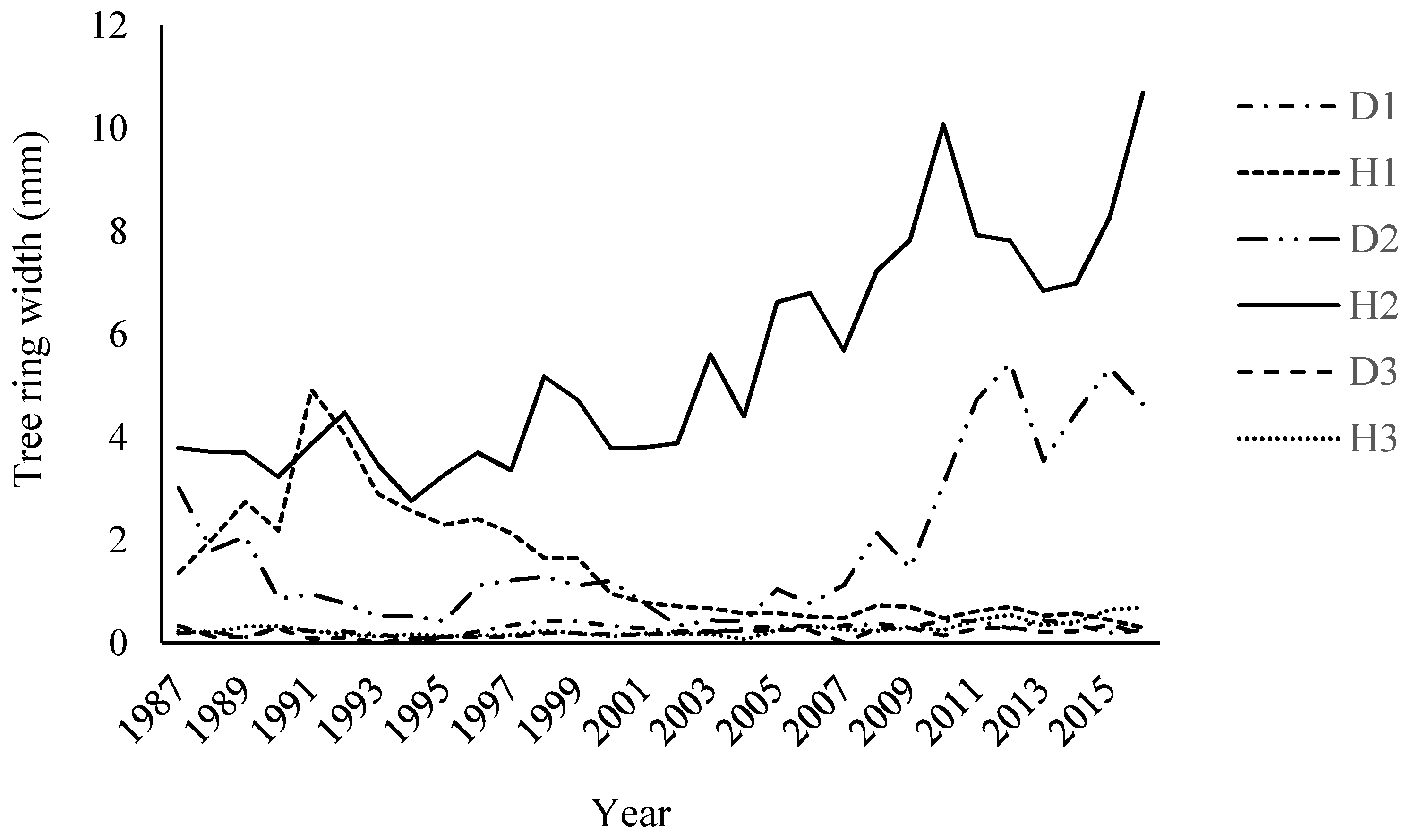
3.1.2. Annual Ring Width (ARW)
3.2. Wood Anatomical Features
3.2.1. Diameter of Earlywood Vessels (DEV)
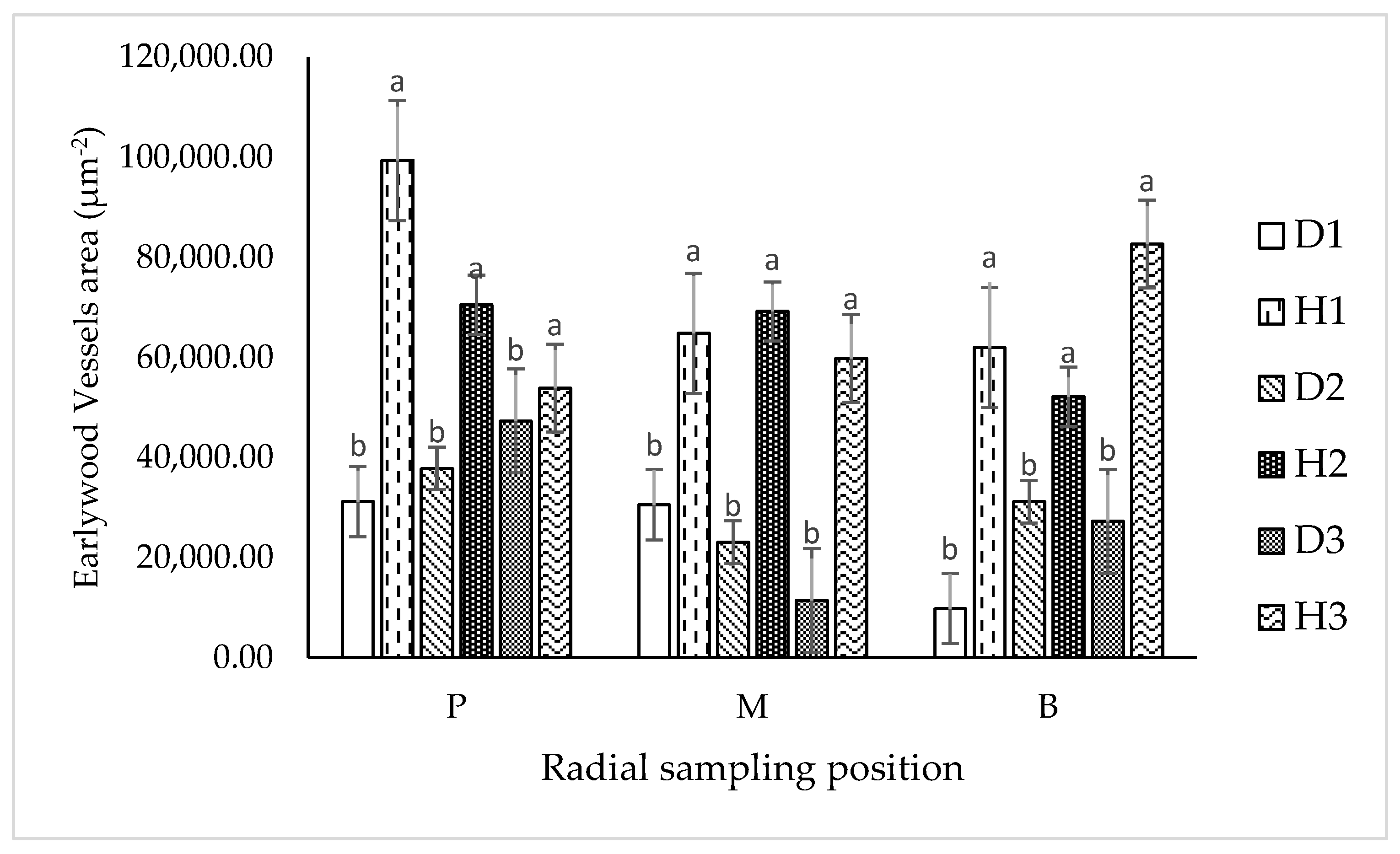
3.2.2. Earlywood Vessels’ Area (EVA)
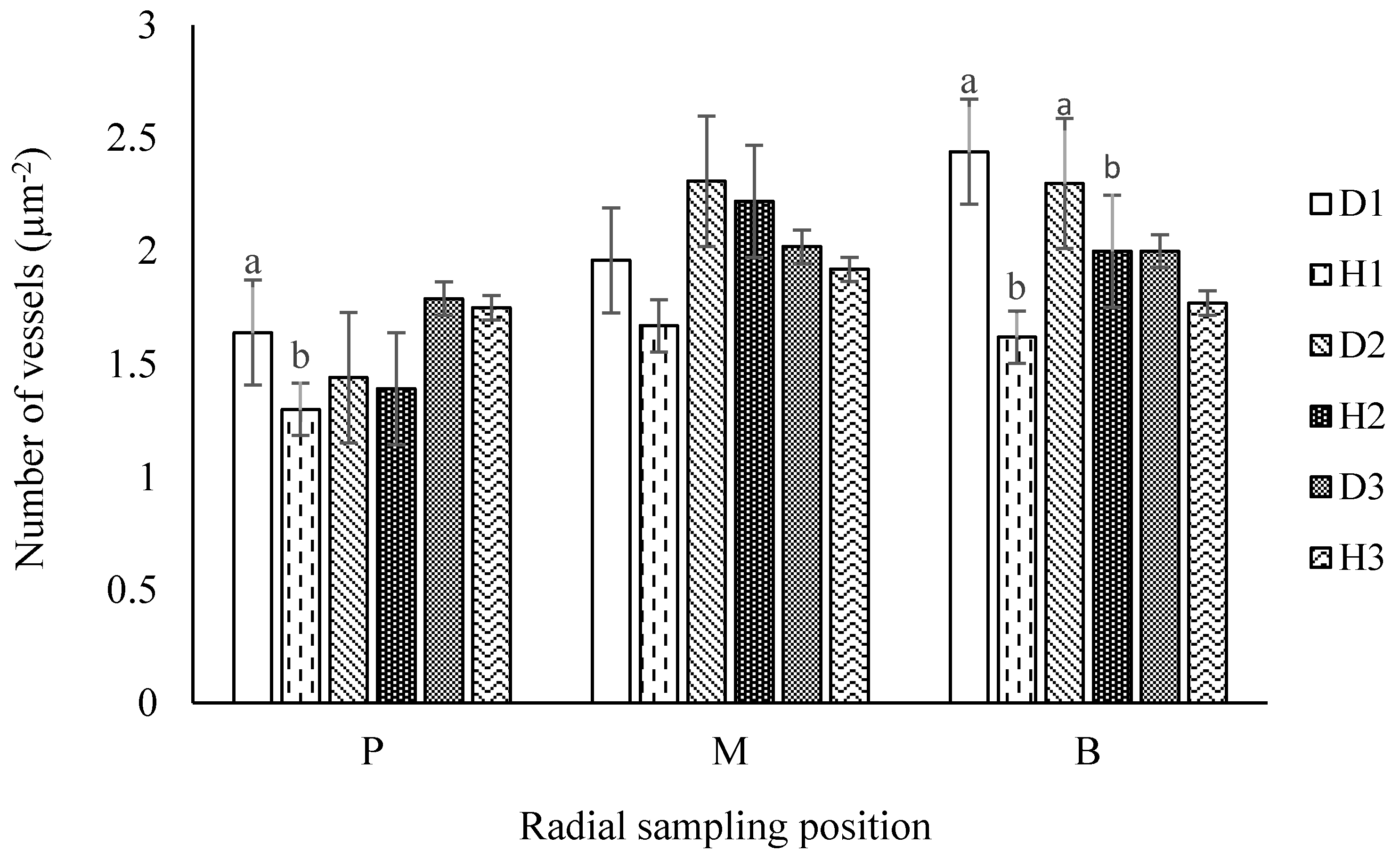
3.2.3. Number of Vessels (NV)
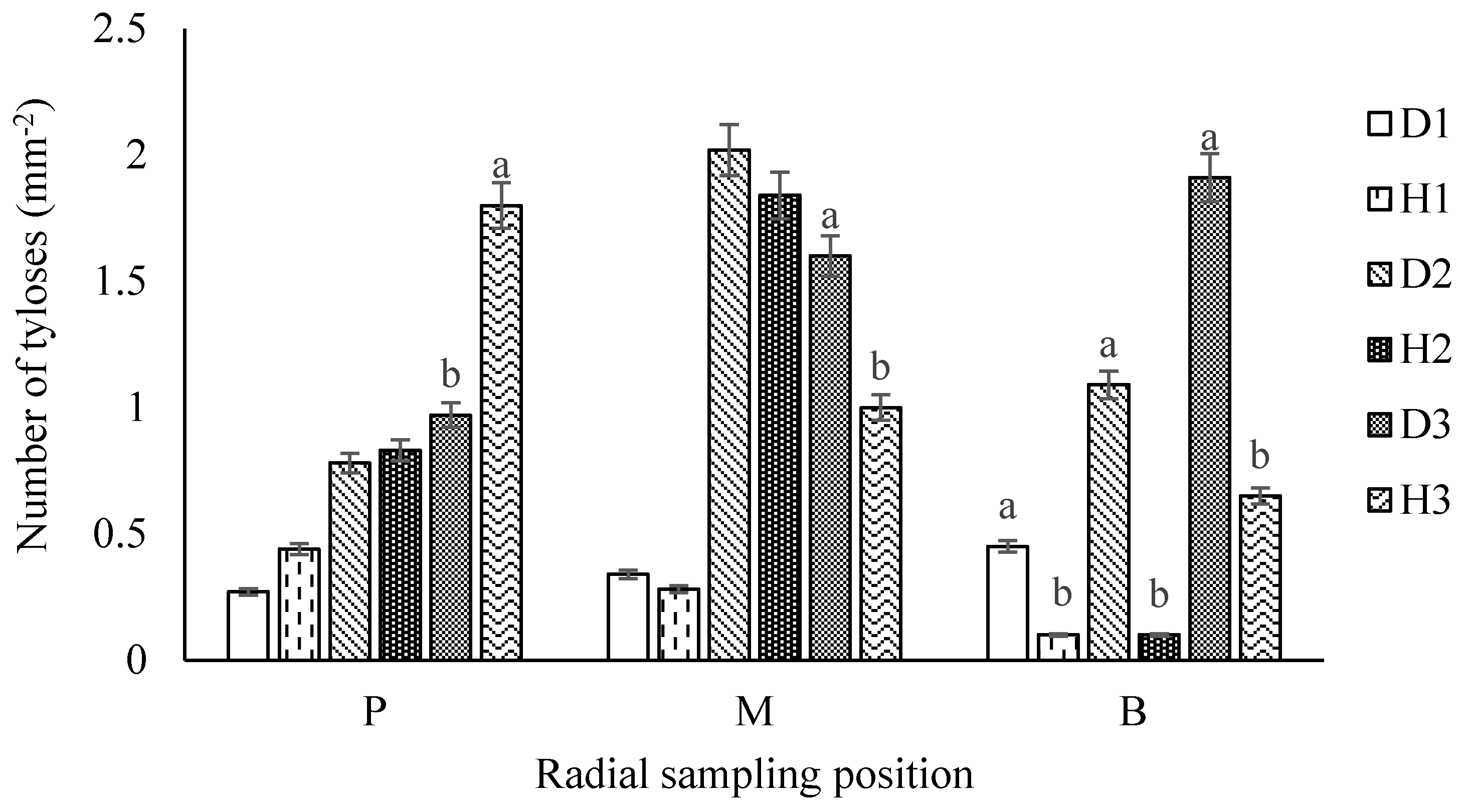
3.2.4. Number of Tyloses (NT)
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- McLaughlin, S.B.; Downing, D.J.; Blasing, T.J.; Cook, E.R.; Adams, H.S. An analysis of climate and competition as contributors to decline of red spruce in high elevation Appalachian forests of the eastern United States. Oecologia 1987, 72, 487–501. [Google Scholar] [CrossRef] [PubMed]
- Kabrick, J.M.; Dey, D.C.; Randy, R.G.; Wallendorf, M. The role of environmental factors in oak decline and mortality in the Ozark Highlands. For. Ecol. Manag. 2008, 255, 1409–1417. [Google Scholar] [CrossRef] [Green Version]
- Philip, M.; David, R.; Leon, A. Oak Decline. Forest Insect and Disease Leaflet 165; U.S. Department of Agriculture, Forest Service, Northern Area State & Private Forestry: Broomall, PA, USA, 1983; pp. 1–8.
- Nourinejad, J.; Rostami, A. Investigation of oak decline and its relation to physiographic factors in the forests of West of Iran. JBES 2014, 5, 201–207. [Google Scholar]
- Sepahvand, T.; Zandebasiri, M. Evaluation of Oak decline with local resident, opinions in Zagros forests, Iran. SJAS 2014, 4, 231–234. [Google Scholar]
- Evert, R.F.; Eichhorn, S.E. Esau’s Plant Anatomy: Meristems, Cells, and Tissues of the Plant Body: Their Structure, Function, and Development; John Wiley and Sons: Hoboken, NJ, USA, 2006. [Google Scholar]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar]
- Pedersen, B.S. The role of stress in the mortality of midwestern oaks as indicated by growth prior to death. Ecology 1998, 79, 79–93. [Google Scholar] [CrossRef]
- Kopabayeva, A.; Mazarzhanova, K.; Köse, N.; Akkemik, U. Tree-ring chronologies of Pinus sylvestris from Burabai Region (Kazakhstan) and their response to climate change. Dendrobiology 2017, 78, 96–110. [Google Scholar] [CrossRef] [Green Version]
- Agusti, J.; Greb, T. Going with the wind adaptive dynamics of plant secondary meristems. Mech. Dev. 2013, 130, 34–44. [Google Scholar] [CrossRef]
- Balducci, L.; Cuny, H.E.; Rathgeber, C.B.K.; Deslauriers, A.; Giovannelli, A.; Rossi, S. Compensatory mechanisms mitigate the effect of warming and drought on wood formation. Plant Cell Environ. 2016, 39, 1338–1352. [Google Scholar] [CrossRef] [PubMed]
- Cuny, H.E.; Rathgeber, C.B.K.; Kiess’e, T.S.; Hartmann, F.P.; Barbeito, I.; Fournier, M. Generalized additive models reveal the intrinsic complexity of wood formation dynamics. J. Exp. Bot. 2013, 64, 1983–1994. [Google Scholar] [CrossRef] [Green Version]
- Brodribb, T.J.; Cochard, H. Hydraulic failure defines the recovery and point of death inwater-stressed conifers. Plant Physiol. 2009, 149, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eilmann, B.; Zweifel, R.; Buchmann, N.; Fonti, P.; Rigling, A. Drought-induced adaptation of the xylem in Scots pine and pubescent oak. Tree Physiol. 2009, 29, 1011–1020. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campelo, F.; Nabais, C.; Gutierrez, E.; Freitas, H.; Garcia-Gonzalez, I. Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees Struct. Funct. 2010, 24, 463–470. [Google Scholar] [CrossRef]
- Sperry, J.S. Evolution of water transport and xylem structure. Int. J. Plant Sci. 2003, 164, S115–S127. [Google Scholar] [CrossRef] [Green Version]
- Anderegg, W.R.L.; Hicke, J.A.; Fisher, R.A.; Allen, C.D.; Aukema, J.; Bentz, B.; Hood, S.; Lichstein, J.W.; Macalady, A.K.; McDowell, N.; et al. Tree mortality from drought, insects, and their interactions in a changing climate. New Phytol. 2015, 208, 674–683. [Google Scholar] [CrossRef]
- Giagli, K.; Gričar, J.; Vavrčik, H.; Menšik, L.; Gryc, V. The effects of drought on wood formation in Fagus sylvatica during two contrasting years. IAWA J. 2016, 37, 332–348. [Google Scholar] [CrossRef]
- Naji, H.R.; Taher Pour, M. The effect of simulated dust storm on wood development and leaf stomata in Quercus brantii L. Desert 2019, 24, 43–49. [Google Scholar]
- Aloni, R.; Zimmermann, M.H.Z. The control of vessel size and density along the plant axis. A new hypothesis. Differentiation 1983, 24, 203–208. [Google Scholar] [CrossRef]
- ISO. Wood-Determination of Density for Physical and Mechanical Tests; ISO 3131; International Organization for Standardization: Geneva, Switzerland, 1975. [Google Scholar]
- Radmehr, A.; Soosani, J.; Balapour, S.; Hosseini Ghale Bahmani, S.M. Effects of climate variables (temperature and precipitation) on the width of Rings-growth in Persian coppice oak in the central Zagros (Case study: Khoramabad). J. Wood For. Sci. Technol. 2014, 22, 93–110. (In Persian) [Google Scholar]
- Camargo, M.A.B.; Marenco, R.A. Density, size and distribution of stomata in 35 rainforest tree species in Central Amazonia. Acta Amaz. 2011, 41, 205–212. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.V.; Aebischer, D.P.; Denne, M.P. Latewood density in relation to wood fibre diameter, wall thickness, and fibre and vessel percentages in Quercus robur L. IAWA J. 1997, 18, 127–138. [Google Scholar] [CrossRef]
- Santiago, L.S.; Goldstein, G.; Meinzer, F.C.; Fisher, J.B.; Machado, K.; Woodruff, D.; Jones, T. Leaf photosynthetic traits scale with hydraulic conductivity and wood density in Panamian forest canopy trees. Ecophysiology 2004, 140, 543–550. [Google Scholar]
- Baas, P.; Ewers, F.W.; Davis, S.D.; Wheeler, E.A. Evolution of xylem physiology. In The Evolution of Plant Physiology; Hemsley, A.R., Poole, I., Eds.; Elsevier Academic Press: London, UK, 2004; pp. 273–295. [Google Scholar]
- Zobel, B.J.; Van Buijtenen, J.P. Wood Variation, Its Causes and Control; Springer: Berlin, Germany, 1989; p. 363. [Google Scholar]
- Preston, K.A.; Cornwell, W.K.; DeNoyer, J.L. Wood density and vessel traits as distinct correlates of ecological strategy in 51 California coast range angiosperms. New Phytol. 2006, 170, 807–818. [Google Scholar] [CrossRef]
- Roque, R.M.; Tomazello-Filho, M. Relationships between anatomical features and intra-ring wood density profiles in Gmelina arborea applying X-ray densitometry. Cerne 2007, 13, 384–392. [Google Scholar]
- Pourtahmasi, K.; Lotfiomaran, N.; Brauning, A.; Parsapajouh, D. Tree-ring width and vessel characteristics of oriental beech (Fagus orientalis) along an altitudinal gradient in the Caspian forests, Northern Iran. IAWA J. 2011, 32, 461–473. [Google Scholar] [CrossRef]
- Fritts, H.C. Tree Ring and Climate; Academic Press: London, UK, 1976; p. 567. [Google Scholar]
- Mirabolfathy, M.; Groenewald, J.; Crous, P. The occurrence of charcoal disease caused by Biscogniauxia mediterranea on chestnut-leaved oak (Quercus castaneifolia) in the Golestan Forests of Iran. Plant Dis. 2013, 95, 3539. [Google Scholar] [CrossRef] [PubMed]
- Reyes-Santamaría, I.; Terrazas, T.; Barrientos-Priego, A.F.; Trejo, C. Xylem conductivity and vulnerability in cultivars and races of avocado. Sci. Hortic. 2002, 92, 97–105. [Google Scholar] [CrossRef]
- Bayramzadeh, V.; Funada, R.; Kubo, T. Relationships between vessel element anatomy and physiological as well as morphological traits of leaves in Fagus crenata seedlings originating from different provenances. Trees 2008, 22, 217–224. [Google Scholar] [CrossRef]
- Tulik, M. The anatomical traits of trunk wood and their relevance to oak (Quercus robur L.) vitality. Eur. J. For. Res. 2014, 133, 845–855. [Google Scholar] [CrossRef] [Green Version]
- Oladi, R.; Bräuning, A.; Pourtahmasi, K. “Plastic” and “static” behavior of vessel-anatomical features in Oriental beech (Fagus orientalis Lipsky) in view of xylem hydraulic conductivity. Trees 2014, 28, 493–502. [Google Scholar] [CrossRef]
- Levanic, T.; Cater, M.; McDowell, N.G. Associations between growth, wood anatomy, carbon isotope discrimination and mortality in a Quercus robur forest. Tree Physiol. 2011, 31, 298–308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nabeshima, E.; Kubo, T.; Yasue, K.; Hiura, T.; Funada, R. Changes in radial growth of earlywood in Quercus crispula between 1970 and 2004 reflect climate change. Trees 2015, 29, 1273–1281. [Google Scholar] [CrossRef]
- Oladi, R.; Nasiriani, S.; Danekar, A.; Pourtahmasi, K. Inter-relations between tree-ring width and vessel features in black alder (Alnus glutinosa). Iran. J. Wood Pap. Sci. 2016, 30, 278–288. [Google Scholar]
- Kulkarni, M.; Schneider, B.; Raveh, E.; Tel-Zur, A.N. Leaf anatomical characteristics and physiological responses to short-term drought in Ziziphus mauritiana (Lamk.). Sci. Hortic. 2010, 124, 316–322. [Google Scholar] [CrossRef]
- Plavcova, L.; Hacke, U.G. Phenotypic and developmental plasticity of xylem in hybrid poplar saplings subjected to experimental drought, nitrogen fertilization, and shading. J. Exp. Bot. 2012, 63, 6481–6491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sperry, J.S.; Nichols, K.L.; Sullivan, J.E.M.; Eastlack, S.E. Xylem embolism in ring-porous, diffuse-porous, and coniferous trees of northern Utah and interior Alaska. Ecology 1994, 75, 1736–1752. [Google Scholar] [CrossRef]
- Fink, S. Pathological and regenerative plant anatomy. In Encyclopedia of Plant Anatomy, Part 6; Gebrüder Borntraeger: Berlin, Germany, 1999; Volume 14, p. 1095. [Google Scholar]
- Tyree, M.T.; Zimmermann, M.H. Xylem Structure and the Ascent of Sap, 2nd ed.; Springer: Berlin, Germany, 2002. [Google Scholar]
- Cochard, H.; Peiffer, M.; Le Gall, K.; Granier, A. Developmental control of xylem hydraulic resistances and vulnerability to embolism in Fraxinus excelsior L.: Impacts of water relations. J. Exp. Bot. 1997, 48, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Baguskas, S.A.; Peterson, S.H.; Bookhagen, B.; Still, C.J. Evaluating spatial patterns of droughtinduced tree mortality in a coastal California pine forest. Forest Ecol. Manag. 2014, 315, 43–53. [Google Scholar] [CrossRef]
- Henriques, J.; Barrento, M.J.; Bonifácio, L.; Gomes, A.A.; Lima, A.; Sousa, E. Factors affecting the dispersion of Biscogniauxia mediterranea in Portuguese cork oak stands. Silva Lusit. 2014, 22, 83–97. [Google Scholar]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forest Area | Altitude (m) | Longitude | Latitude | Av. Precip. | Av. Temp. | Av. RH |
|---|---|---|---|---|---|---|
| mm | °C | % | ||||
| Dareh shahr (A) | 933 | 33°3′30″ N | 47°19′30″ E | 465.1 | 19.5 | 41 |
| Sheshdar (B) | 2230 | 33°38′55″ N | 46°30′37″ E | 582.2 | 16.9 | 40 |
| Chavar (C) | 1680 | 33°43′03″ N | 46°14′36″ E |
| Mean Square | |||
|---|---|---|---|
| Source of Variation | df | WD (g/cm3) | ARW (µm) |
| Decline | 1 | 0.57 ** | 115.408 ** |
| Areas | 2 | 0.13 ** | 201.184 ** |
| Decline × Areas | 2 | 0.005 * | 47.531 ** |
| Error | 594 | 0.002 | 1.439 |
| Attributes | Sampling Group | Mean | Difference | t-test |
|---|---|---|---|---|
| WD (g/cm3) | Declined | 0.92 ± 0.02 | 0.048 | 5.181 * |
| Healthy | 0.87 ± 0.06 | |||
| ARW (µm) | Declined | 0.78 ± 0.24 | 1.60 | 5.241 * |
| Healthy | 2.38 ± 0.54 |
| Mean Square | ||||||
|---|---|---|---|---|---|---|
| The Source of Variation | df | DEV (µm) | EVA (µm−2) | NV (mm−2) | NT (mm−2) | |
| Decline | Pith | 1 | 38.977 ** | 27.906 ** | 1.533 ns | 14.092** |
| Middle | 762.045 ** | 127.558 ** | 2.219 ns | 8.477 ** | ||
| Bark | 508.746 ** | 64.223 ** | 11.486 ** | 111.562 ** | ||
| Area | Pith | 2 | 1.571 ns | 1.71 ns | 3.633 * | 40.897 ** |
| Middle | 9.400 ** | 3.984 * | 6.040 ** | 96.150 ** | ||
| Bark | 1.125 ns | 4.442 * | 1.332 ns | 52.205 ** | ||
| Decline × Area | Pith | 2 | 13.842 ** | 6.941 ** | 0.722 ns | 6.764 ** |
| Middle | 32.288 ** | 1.330 ns | 0.367 ns | 2.898 ns | ||
| Bark | 67.281 ** | 4.221 * | 1.964 ns | 10.815 ** | ||
| Error | Pith | 594 | 12,267.234 | 2.622 × 1011 | 2.010 | 0.918 |
| Middle | 2808.444 | 2,165,248,861 | 1.730 | 1.387 | ||
| Bark | 3290.675 | 4,289,042,107 | 2.104 | 1.396 | ||
| Anatomical Sampling | Pith | Middle | Bark | |
|---|---|---|---|---|
| Group Features | ||||
| DEV (µm) | Declined Healthy | 228.43 ± 15.54 ** | 171.74 ± 6.72 ** | 173.85 ± 7.9 ** |
| 284.88 ± 10.00 | 291.19 ± 6.28 | 282.93 ± 7.18 | ||
| EVA (µm2) | Declined Healthy | 38,754 ± 3391.46 ** | 21,669 ± 2852.86 ** | 22,720 ± 2646.88 ** |
| 74,562.83 ± 13,280.79 | 64,579.56 ± 7088.08 | 65,573.28 ± 10,484.12 | ||
| NV (mm−2) | Declined Healthy | 1.62 ± 0.16 ns | 2.09 ± 0.16 ns | 2.24 ± 0.18 ** |
| 1.48 ± 0.14 | 1.93 ± 0.12 | 1.79 ± 0.18 | ||
| NT (mm−2) | Declined Healthy | 0.67 ± 0.12 ns | 1.32 ± 0.16 ns | 1.15 ± 0.16 ** |
| 1.02 ± 0.14 | 1.04 ± 0.12 | 0.28 ± 0.06 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soheili, F.; Woodward, S.; Almasi, I.; Abdul-Hamid, H.; Naji, H.R. Variations in Wood Density, Annual Ring Width and Vessel Properties of Quercus brantii Affected by Crown Dieback. Forests 2021, 12, 642. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050642
Soheili F, Woodward S, Almasi I, Abdul-Hamid H, Naji HR. Variations in Wood Density, Annual Ring Width and Vessel Properties of Quercus brantii Affected by Crown Dieback. Forests. 2021; 12(5):642. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050642
Chicago/Turabian StyleSoheili, Forough, Stephen Woodward, Isaac Almasi, Hazandy Abdul-Hamid, and Hamid Reza Naji. 2021. "Variations in Wood Density, Annual Ring Width and Vessel Properties of Quercus brantii Affected by Crown Dieback" Forests 12, no. 5: 642. https://0-doi-org.brum.beds.ac.uk/10.3390/f12050642