Developmental Dynamics of Gilbertiodendron dewevrei (Fabaceae) Drive Forest Structure and Biomass in the Eastern Congo Basin

 , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
1.1. Hypotheses Related to Forest Structure and Growth
1.2. Hypotheses Related to Tree Species Composition
1.3. Hypotheses Related to Development and Geographic Expansion
2. Materials and Methods
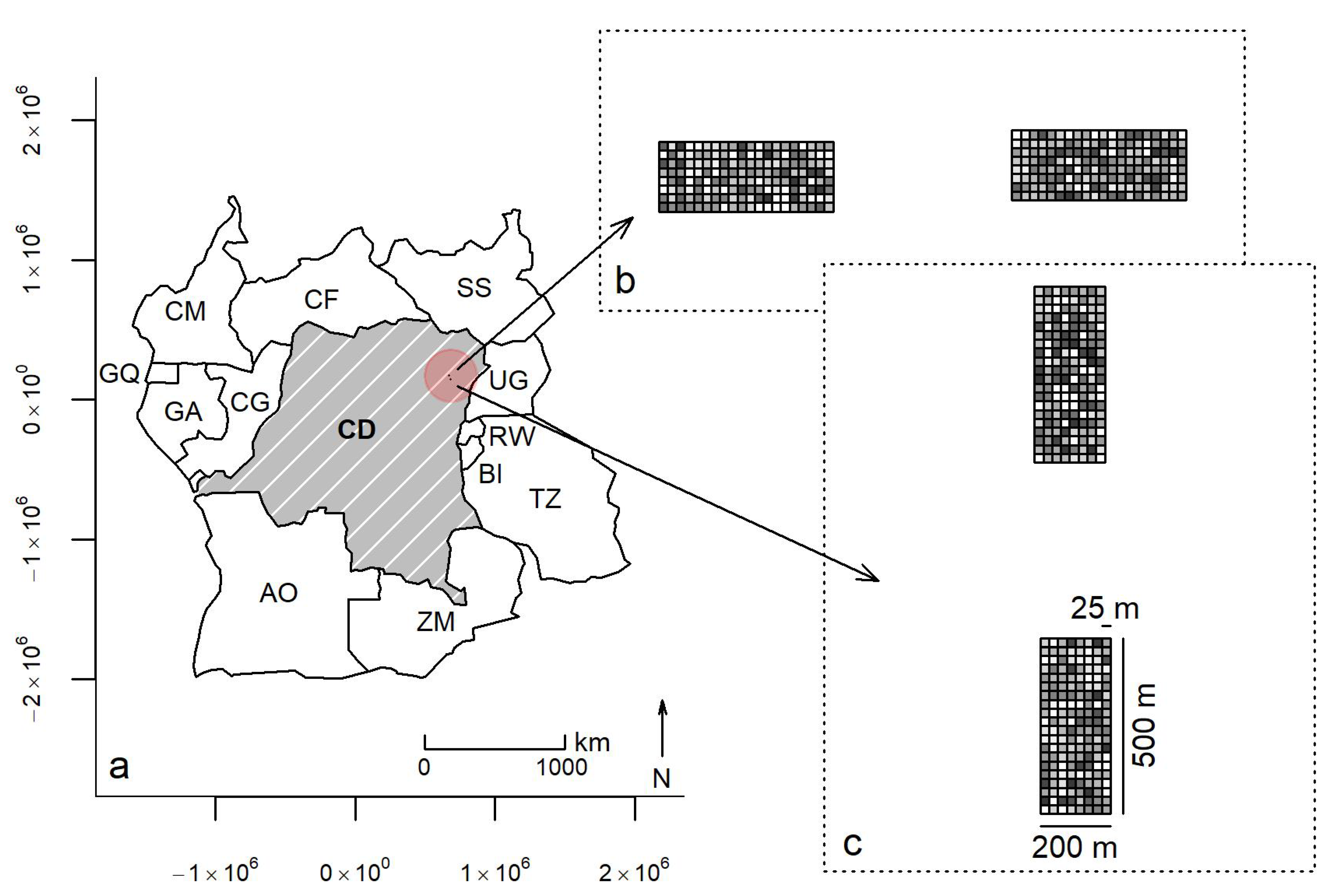
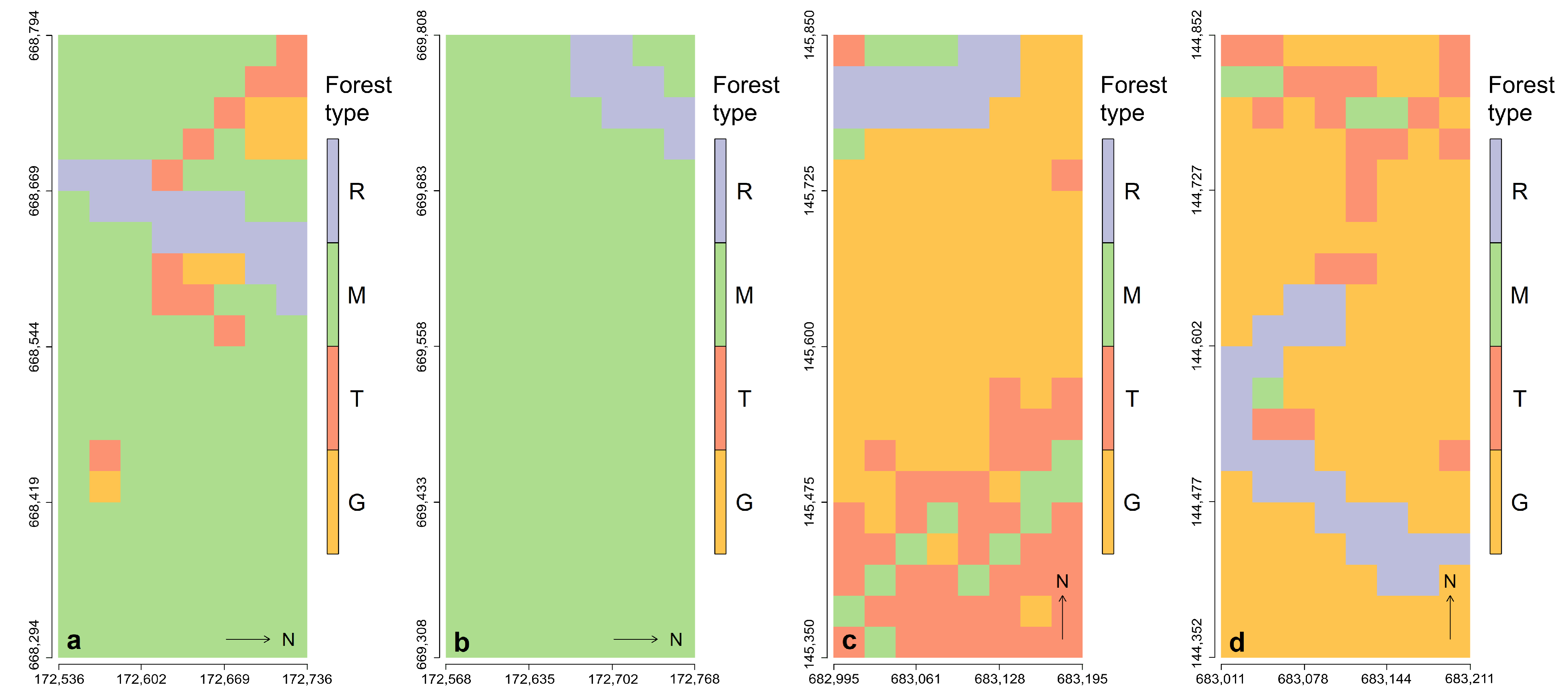
2.1. Field Plots and Data Structures
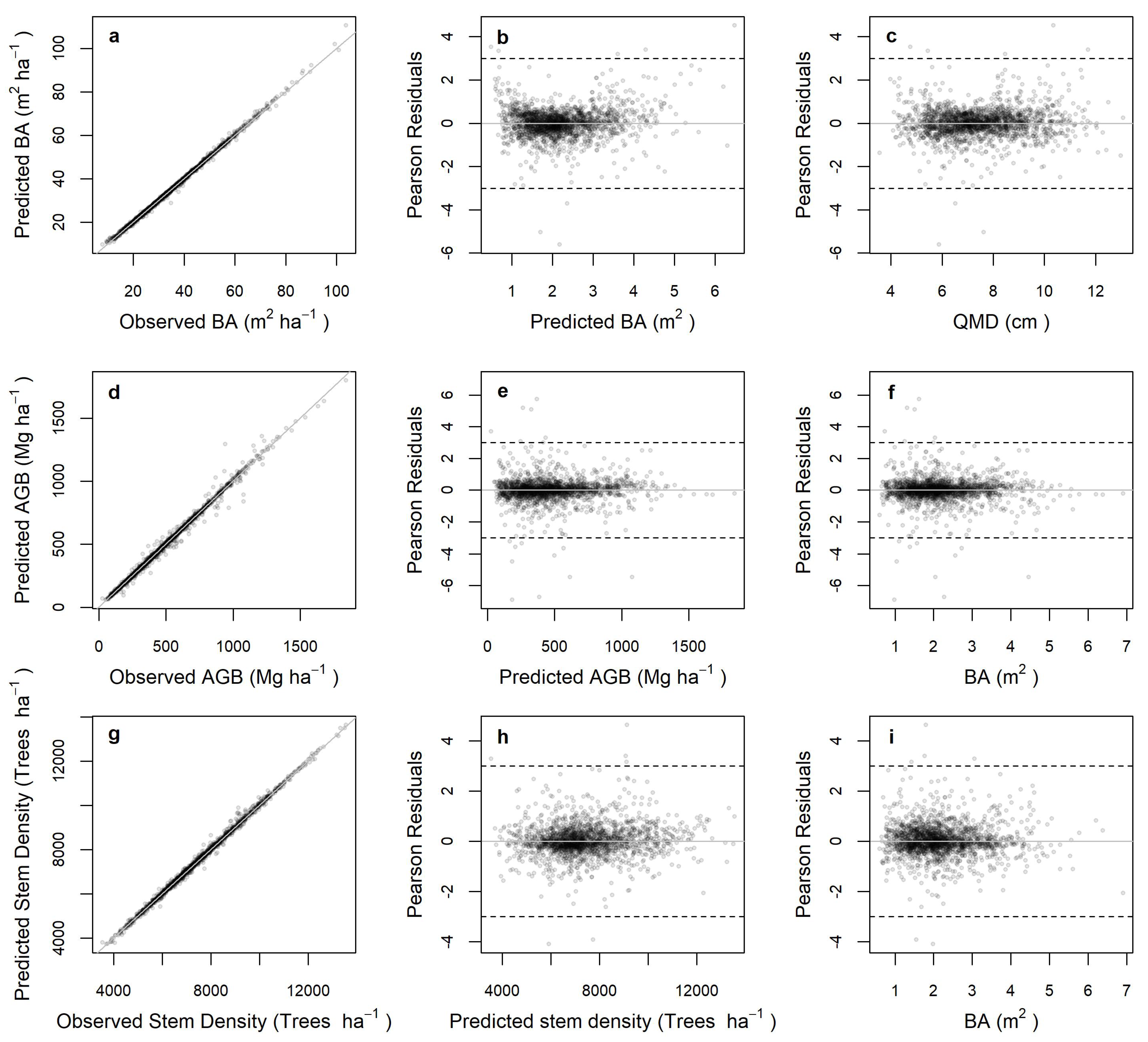
2.2. Statistical Analysis
3. Results
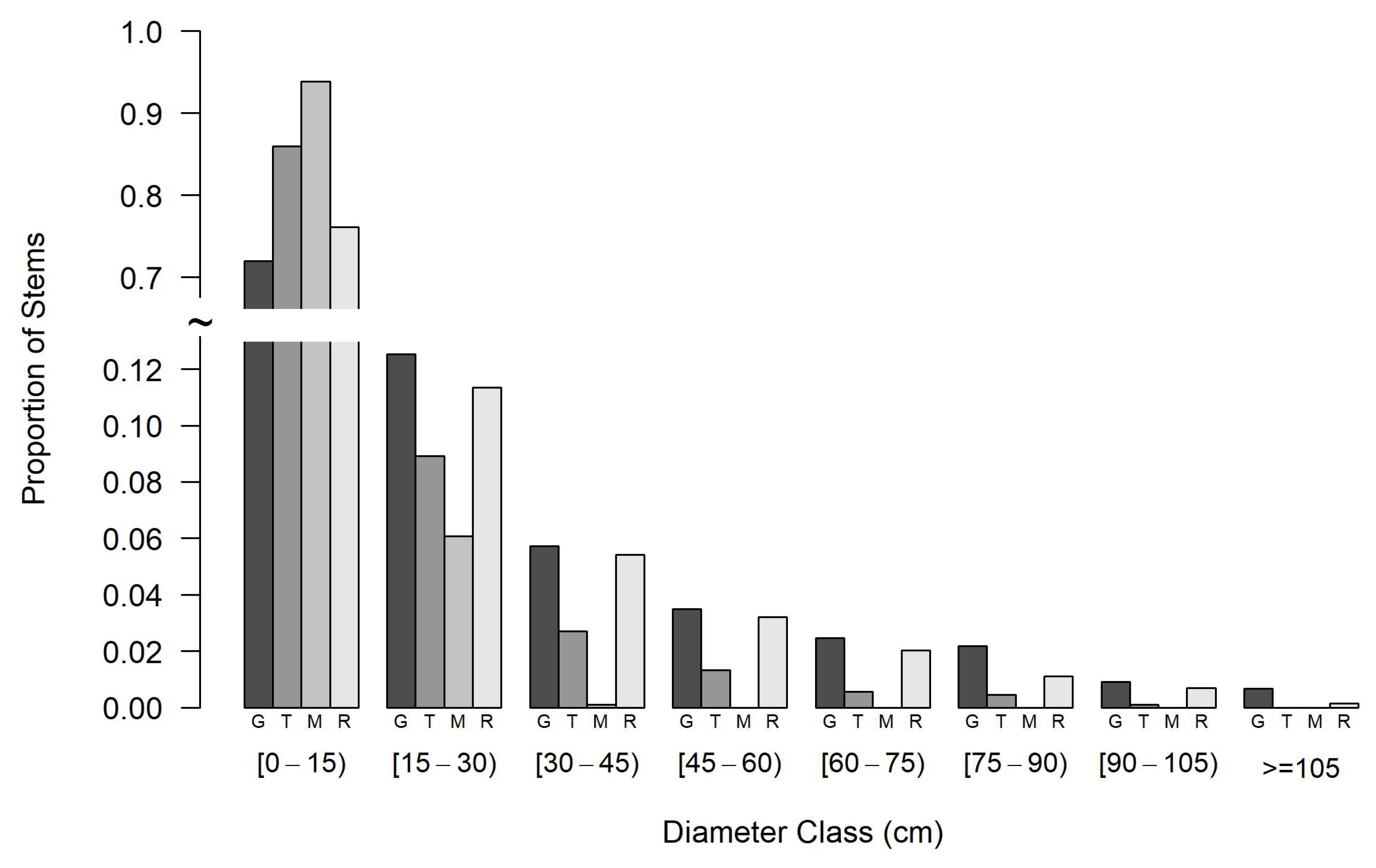
3.1. Overall Results
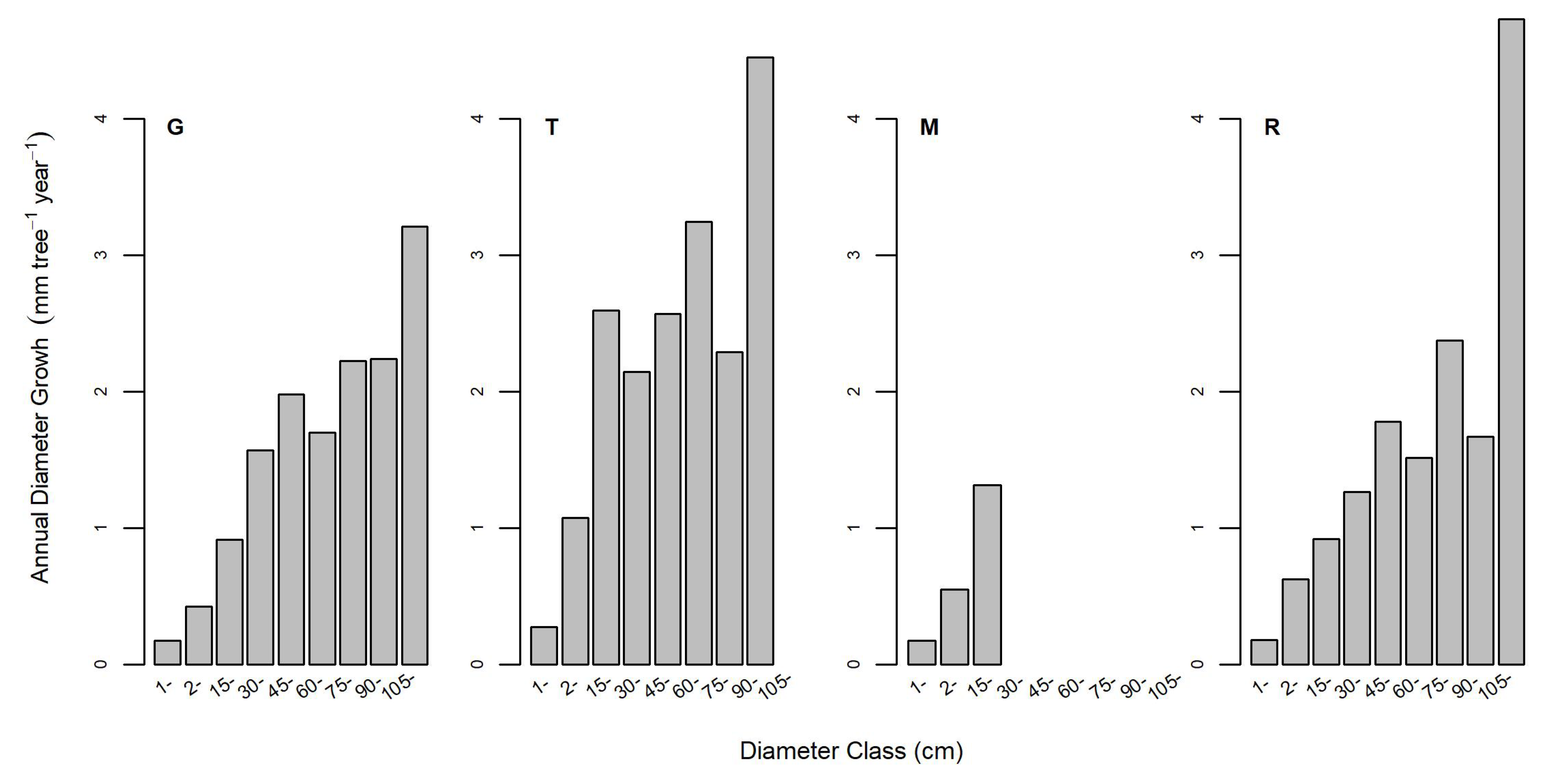
3.2. Hypotheses Related to Forest Structure and Growth
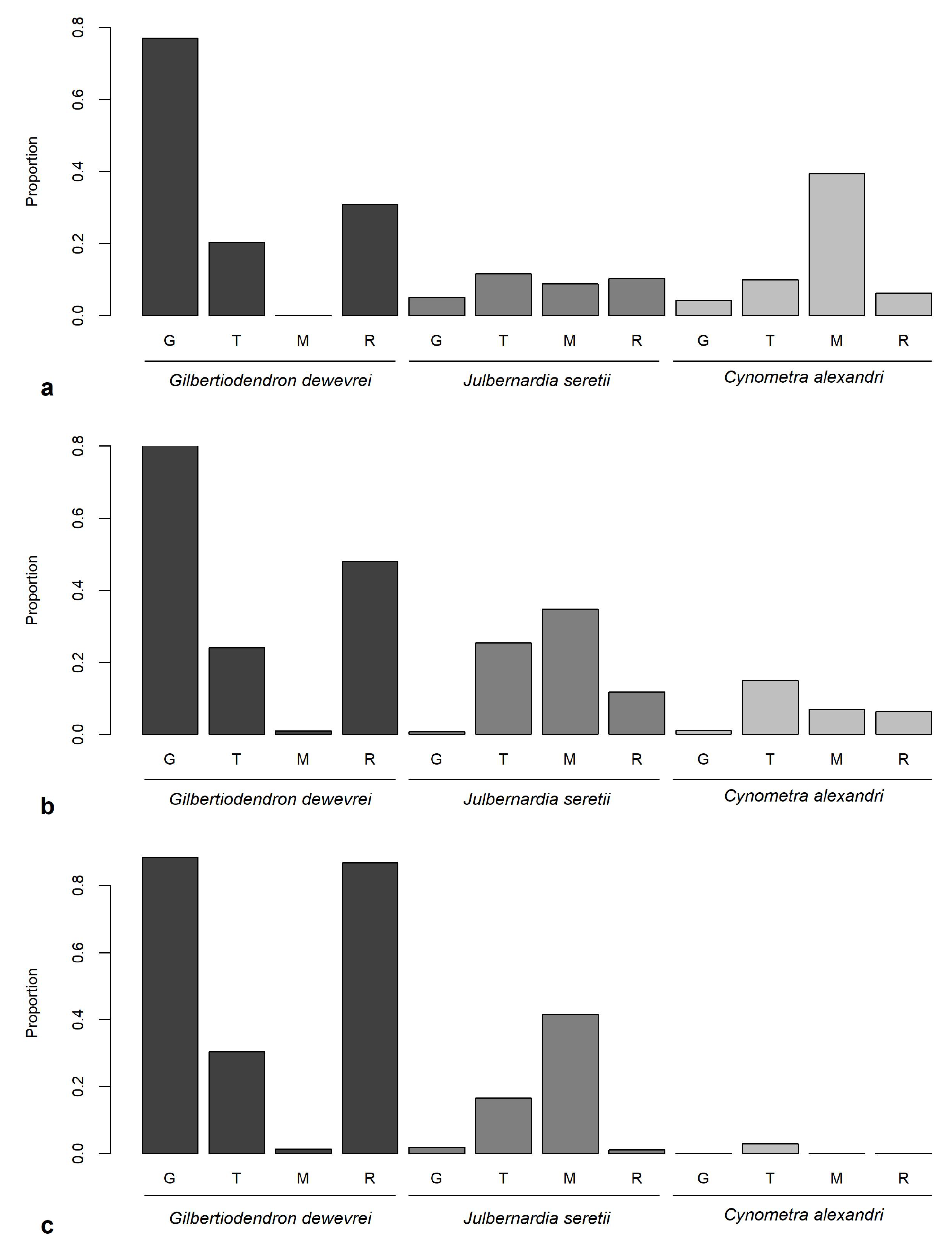
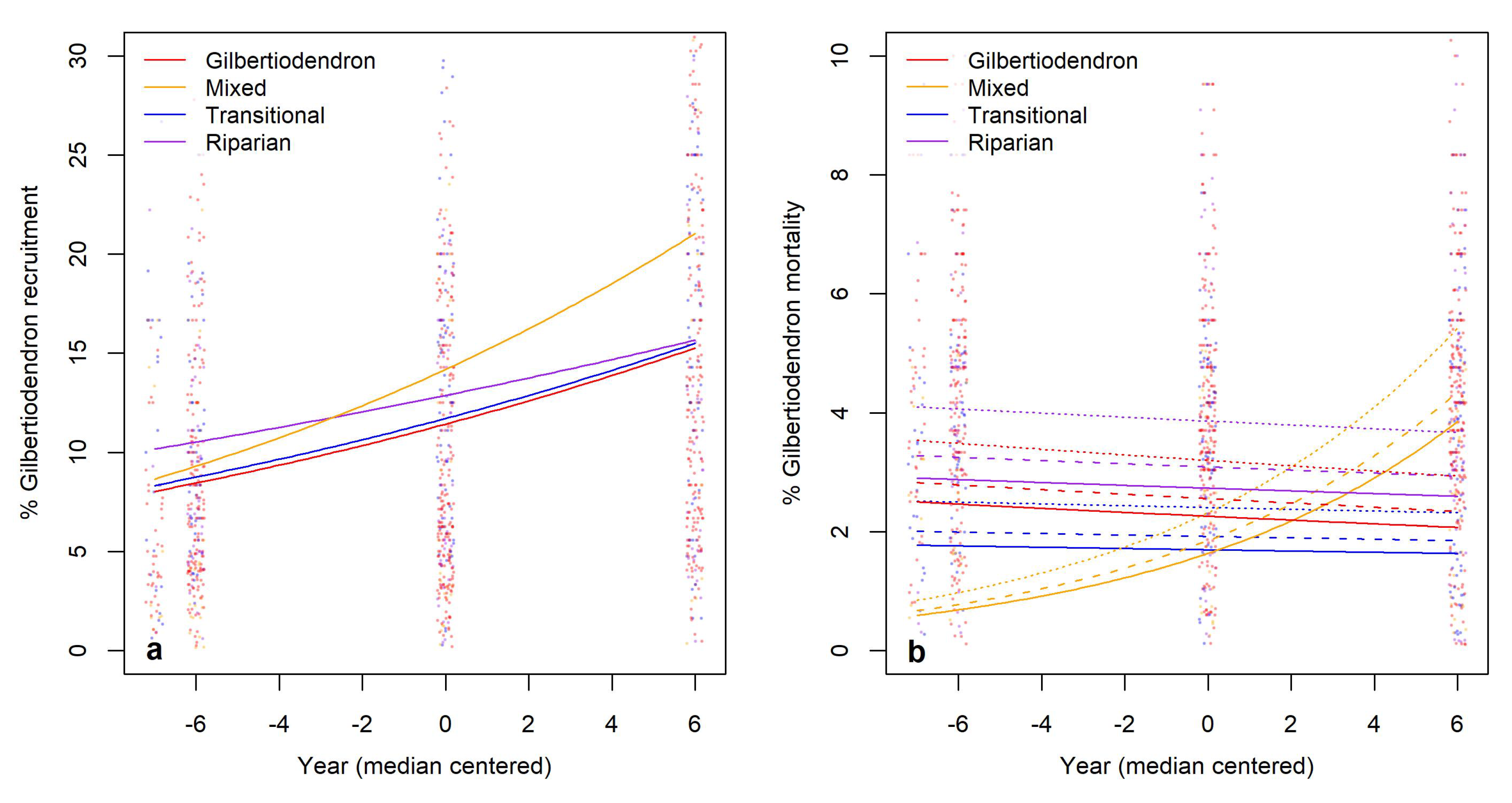
3.3. Hypotheses Related to Tree Species Composition
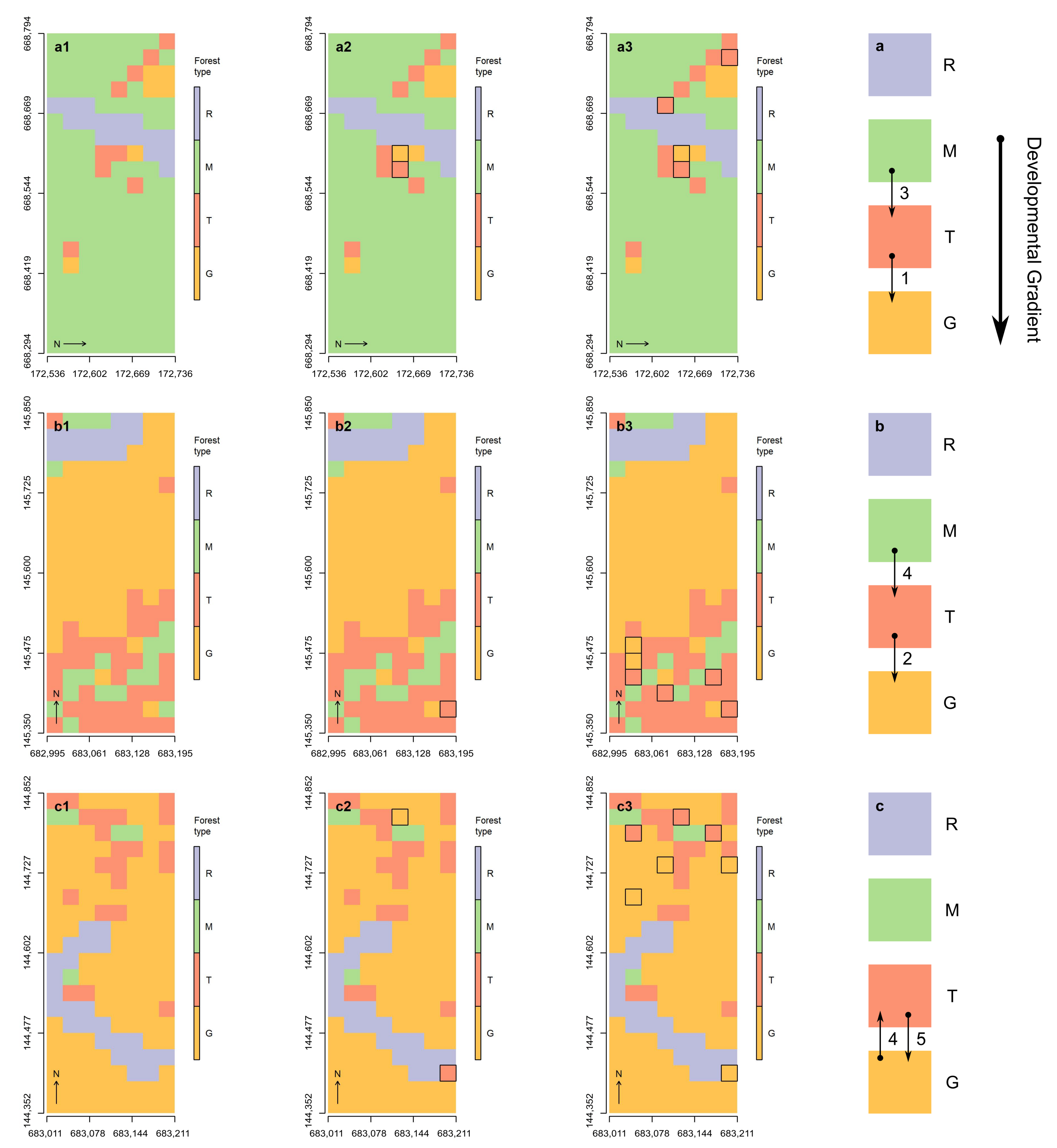
3.4. Hypotheses Related to Development and Geographic Expansion
4. Discussion
4.1. AGB and BA
4.2. Species Diversity and Stem Density
4.3. Succession and Geographic Expansion
4.4. Support for Existing Developmental Dynamics Frameworks
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Makana, J.R.; Ewango, C.N.; McMahon, S.M.; Thomas, S.C.; Hart, T.B.; Condit, R. Demography and biomass change in monodominant and mixed old-growth forest of the Congo. J. Trop. Ecol. 2011, 27, 447–461. [Google Scholar] [CrossRef]
- Lewis, S.L.; Lopez-Gonzalez, G.; Sonké, B.; Affum-Baffoe, K.; Baker, T.R.; Ojo, L.O.; Phillips, O.L.; Reitsma, J.M.; White, L.; Comiskey, J.A. Increasing carbon storage in intact African tropical forests. Nature 2009, 457, 1003. [Google Scholar] [CrossRef] [PubMed]
- Davis, T.; Richards, P.W. The vegetation of Moraballi Creek, British Guiana: An ecological study of a limited area of tropical rain forest. Part II. J. Ecol. 1934, 106–155. [Google Scholar] [CrossRef]
- Beard, J. The Mora forests of Trinidad, British West Indies. J. Ecol. 1946, 173–192. [Google Scholar] [CrossRef]
- Gérard, P. Etude Ècologique de la Forêt Dense à Gilbertiodendron dewevrei dans la Région de l’Uele; Institut national pour l’étude agronomique du Congo: Brussels, Belgium, 1960. [Google Scholar]
- Hart, T.B. THE Ecology of a Single-Species-Dominant Forest and of a Mixed Forest in Zaire, Africa. Ph.D. Thesis, Michigan State University, East Lansing, MI, USA, 1985. [Google Scholar]
- Makana, J.R.; Terese, B.; Hibbs, D.; Condit, R.; Losos, E.; Leigh, E. Stand structure and species diversity in the Ituri forest dynamics plots: A comparison of monodominant and mixed forest stands. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; University of Chicago Press: Chicago, IL, USA, 2004; Chapter 12; pp. 159–174. [Google Scholar]
- Day, M.; Baldauf, C.; Rutishauser, E.; Sunderland, T.C. Relationships between tree species diversity and above-ground biomass in central African rainforests: Implications for REDD. Environ. Conserv. 2013, 41, 64–72. [Google Scholar] [CrossRef]
- Lewis, S.L.; Malhi, Y.; Phillips, O.L. Fingerprinting the impacts of global change on tropical forests. Philos. Trans. R. Soc. London. Ser. B 2004, 359, 437–462. [Google Scholar] [CrossRef] [PubMed]
- Hart, T.B.; Hart, J.A.; Murphy, P.G. Monodominant and species-rich forests of the humid tropics: Causes for their co-occurrence. Am. Nat. 1989, 133, 613–633. [Google Scholar] [CrossRef]
- Peh, K.S.H.; Sonké, B.; Lloyd, J.; Quesada, C.A.; Lewis, S.L. Soil does not explain monodominance in a Central African tropical forest. PLoS ONE 2011, 6, e16996. [Google Scholar] [CrossRef]
- Lokonda, M.; Freycon, V.; Gourlet-Fleury, S.; Kombele, F. newblock Are soils under monodominant Gilbertiodendron dewevrei and under adjacent mixed forests similar? A case study in the Democratic Republic of Congo. J. Trop. Ecol. 2018, 34, 176–185. [Google Scholar] [CrossRef]
- Maley, J. A catastrophic destruction of African forests about 2500 years ago still exerts a major influence on present vegetation formations. IDS Bull. 2002, 33, 13–30. [Google Scholar] [CrossRef]
- Hart, T.B. Seed, seedling and sub-canopy survival in monodominant and mixed forests of the Ituri Forest, Africa. J. Trop. Ecol. 1995, 11, 443–459. [Google Scholar] [CrossRef]
- Torti, S.D.; Coley, P.D.; Kursar, T.A. Causes and consequences of monodominance in tropical lowland forests. Am. Nat. 2001, 157, 141–153. [Google Scholar] [CrossRef] [PubMed]
- Tovar, C.; Harris, D.J.; Breman, E.; Brncic, T.; Willis, K.J. Tropical monodominant forest resilience to climate change in Central Africa: A Gilbertiodendron dewevrei forest pollen record over the past 2700 years. J. Veg. Sci. 2019, 30, 575–586. [Google Scholar] [CrossRef]
- Connell, J.H.; Lowman, M.D. Low-diversity tropical rain forests: Some possible mechanisms for their existence. Am. Nat. 1989, 134, 88–119. [Google Scholar] [CrossRef]
- Hart, T.B. Monospecific dominance in tropical rain forests. Trends Ecol. Evol. 1990, 5, 6–11. [Google Scholar] [CrossRef]
- Hart, T.B.; Hart, J.A.; Dechamps, R.; Fournier, M.; Ataholo, M. Changes in forest composition over the last 4000 years in the Ituri basin, Zaire. In The Biodiversity of African Plants; van der Maesen, L.J.G., van der Burgt, X.M., van Medenbach de Rooy, J.M., Eds.; Springer: Wageningen, The Netherlands, 1996; pp. 545–560. [Google Scholar] [CrossRef]
- Read, J.; Hallam, P.; Cherrier, J.F. The anomaly of monodominant tropical rainforests: Some preliminary observations in the Nothofagus-dominated rainforests of New Caledonia. J. Trop. Ecol. 1995, 11, 359–389. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Muller-Landau, H.C.; Thomas, S.C.; Ashton, P.S.; Bunyavejchewin, S.; Dattaraja, H.S.; Davies, S.J.; Esufali, S.; Ewango, C.E. Assessing evidence for a pervasive alteration in tropical tree communities. PLoS Biol. 2008, 6, e45. [Google Scholar] [CrossRef]
- Nelson, B.W. Pervasive alteration of tree communities in undisturbed Amazonian forests. Biotropica 2005, 37, 158–159. [Google Scholar] [CrossRef]
- Wright, S.J. Tropical forests in a changing environment. Trends Ecol. Evol. 2005, 20, 553–560. [Google Scholar] [CrossRef]
- Sullivan, M.J.; Talbot, J.; Lewis, S.L.; Phillips, O.L.; Qie, L.; Begne, S.K.; Chave, J.; Cuni-Sanchez, A.; Hubau, W.; Lopez-Gonzalez, G. Diversity and carbon storage across the tropical forest biome. Sci. Rep. 2017, 7, 39102. [Google Scholar] [CrossRef]
- Lewis, S.L.; Sonké, B.; Sunderl, T.; Begne, S.K.; Lopez-Gonzalez, G.; Van Der Heijden, G.M.; Phillips, O.L.; Affum-Baffoe, K.; Baker, T.R.; Banin, L.; et al. Above-ground biomass and structure of 260 African tropical forests. Philos. Trans. R. Soc. B 2013, 368, 20120295. [Google Scholar] [CrossRef]
- Bauters, M.; Vercleyen, O.; Vanlauwe, B.; Six, J.; Bonyoma, B.; Badjoko, H.; Hubau, W.; Hoyt, A.; Boudin, M.; Verbeeck, H.; et al. Long-term recovery of the functional community assembly and carbon pools in an African tropical forest succession. Biotropica 2019, 51, 319–329. [Google Scholar] [CrossRef]
- Umunay, P.; Gregoire, T.; Ashton, M. Estimating biomass and carbon for Gilbertiodendron dewevrei (De Wild) Leonard, a dominant canopy tree of African tropical rainforest: Implications for policies on carbon sequestration. For. Ecol. Manag. 2017, 404, 31–44. [Google Scholar] [CrossRef]
- Hall, J.S.; Harris, D.J.; Saltonstall, K.; Medjibe, V.d.P.; Ashton, M.S.; Turner, B.L. Resource acquisition strategies facilitate Gilbertiodendron dewevrei monodominance in African lowland forests. J. Ecol. 2019. [Google Scholar] [CrossRef]
- Makana, J.R.; Thomas, S.C. Effects of Light Gaps and Litter Removal on the Seedling Performance of Six African Timber Species. Biotropica J. Biol. Conserv. 2005, 37, 227–237. [Google Scholar] [CrossRef]
- Djuikouo, M.N.K.; Peh, K.S.H.; Nguembou, C.K.; Doucet, J.L.; Lewis, S.L.; Sonké, B. Stand structure and species co-occurrence in mixed and monodominant Central African tropical forests. J. Trop. Ecol. 2014, 30, 447–455. [Google Scholar] [CrossRef]
- Chave, J.; Condit, R.; Lao, S.; Caspersen, J.P.; Foster, R.B.; Hubbell, S.P. Spatial and temporal variation of biomass in a tropical forest: Results from a large census plot in Panama. J. Ecol. 2003, 91, 240–252. [Google Scholar] [CrossRef]
- Makana, J.R.; Hart, T.; Liengola, I.; Ewango, C.; Hart, J.; Condit, R. Ituri forest dynamics plots, Democratic Republic of Congo. In Tropical Forest Diversity and Dynamism: Findings from a Large-Scale Plot Network; University of Chicago Press: Chicago, IL, USA, 2004; Chapter 28; pp. 492–505. [Google Scholar]
- Muller-Landau, H.C. Interspecific and inter-site variation in wood specific gravity of tropical trees. Biotropica 2004, 36, 20–32. [Google Scholar] [CrossRef]
- Condit, R. Tropical Forest Census Plots: Methods and Results from Barro Colorado Island, Panama and a Comparison with Other Plots; Springer: Berlin/Heidelberg, Germany, 1998. [Google Scholar]
- Kearsley, E.; Verbeeck, H.; Hufkens, K.; Van de Perre, F.; Doetterl, S.; Baert, G.; Beeckman, H.; Boeckx, P.; Huygens, D. Functional community structure of African monodominant Gilbertiodendron dewevrei forest influenced by local environmental filtering. Ecol. Evol. 2017, 7, 295–304. [Google Scholar] [CrossRef]
- Fitzmaurice, G.; Laird, N.; Ware, J. Applied Longitudinal Analysis; Wiley Series in Probability and Statistics; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2004. [Google Scholar]
- Jelinski, D.E.; Wu, J. The modifiable areal unit problem and implications for landscape ecology. Landsc. Ecol. 1996, 11, 129–140. [Google Scholar] [CrossRef]
- Kieschnick, R.; McCullough, B.D. Regression analysis of variates observed on (0, 1): Percentages, proportions and fractions. Stat. Model. 2003, 3, 193–213. [Google Scholar] [CrossRef]
- Ferrari, S.; Cribari-Neto, F. Beta regression for modelling rates and proportions. J. Appl. Stat. 2004, 31, 799–815. [Google Scholar] [CrossRef]
- Berger, S.; Graham, N.; Zeileis, A. Various versatile variances: An object-oriented implementation of clustered covariances in R. In Report, Working Papers in Economics and Statistics; University of Innsbruck: Innsbruck, Austria, 2017; Volume 12. [Google Scholar]
- Arellano, M. Practitioner’s Corner: Computing robust standard errors for within-groups estimators. Oxf. Bull. Econ. Stat. 1987, 49, 431–434. [Google Scholar] [CrossRef]
- Liang, K.Y.; Zeger, S.L. Longitudinal data analysis using generalized linear models. Biometrika 1986, 73, 13–22. [Google Scholar] [CrossRef]
- Cameron, A.C.; Gelbach, J.B.; Miller, D.L. Robust inference with multiway clustering. J. Bus. Econ. Stat. 2011, 29, 238–249. [Google Scholar] [CrossRef]
- Chave, J.; Andalo, C.; Brown, S.; Cairns, M.A.; Chambers, J.; Eamus, D.; Fölster, H.; Fromard, F.; Higuchi, N.; Kira, T. Tree allometry and improved estimation of carbon stocks and balance in tropical forests. Oecologia 2005, 145, 87–99. [Google Scholar] [CrossRef]
- Baskerville, G. Use of logarithmic regression in the estimation of plant biomass. Can. J. For. Res. 1972, 2, 49–53. [Google Scholar] [CrossRef]
- Millard, S.P. EnvStats: An R Package for Environmental Statistics; Springer: New York, NY, USA, 2013. [Google Scholar]
- Muller-Landau, H.C.; Condit, R.S.; Harms, K.E.; Marks, C.O.; Thomas, S.C.; Bunyavejchewin, S.; Chuyong, G.; Co, L.; Davies, S.; Foster, R.; et al. Comparing tropical forest tree size distributions with the predictions of metabolic ecology and equilibrium models. Ecol. Lett. 2006, 9, 589–602. [Google Scholar] [CrossRef]
- Oliver, C.D.; Larson, B.C. Forest Stand Dynamics; John Wiley & Sons Inc.: Hoboken, NJ, USA, 1996. [Google Scholar]
- Niklas, K.J.; Midgley, J.J.; Rand, R.H. Tree size frequency distributions, plant density, age and community disturbance. Ecol. Lett. 2003, 6, 405–411. [Google Scholar] [CrossRef]
- Fayolle, A.; Picard, N.; Doucet, J.L.; Swaine, M.; Bayol, N.; Bénédet, F.; Gourlet-Fleury, S. A new insight in the structure, composition and functioning of central African moist forests. For. Ecol. Manag. 2014, 329, 195–205. [Google Scholar] [CrossRef]
- Peh, K.S.H.; Sonké, B.; Séné, O.; Djuikouo, M.N.K.; Nguembou, C.K.; Taedoumg, H.; Begne, S.K.; Lewis, S.L. Mixed-forest species establishment in a monodominant forest in central Africa: Implications for tropical forest invasibility. PLoS ONE 2014, 9, e97585. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, S.I. Forest gap dynamics and tree regeneration. J. For. Res. 2000, 5, 223–229. [Google Scholar] [CrossRef]
- Cuni-Sanchez, A.; White, L.J.; Calders, K.; Jeffery, K.J.; Abernethy, K.; Burt, A.; Disney, M.; Gilpin, M.; Gomez-Dans, J.L.; Lewis, S.L. African savanna-forest boundary dynamics: A 20-year study. PLoS ONE 2016, 11, e0156934. [Google Scholar] [CrossRef] [PubMed]
- Djuikouo, M.N.K.; Doucet, J.L.; Nguembou, C.K.; Lewis, S.L.; Sonké, B. Diversity and aboveground biomass in three tropical forest types in the Dja Biosphere Reserve, Cameroon. Afr. J. Ecol. 2010, 48, 1053–1063. [Google Scholar] [CrossRef]
- Baker, T.R.; Phillips, O.L.; Malhi, Y.; Almeida, S.; Arroyo, L.; Di Fiore, A.; Erwin, T.; Killeen, T.J.; Laurance, S.G.; Laurance, W.F.; et al. Variation in wood density determines spatial patterns in Amazonian forest biomass. Glob. Chang. Biol. 2004, 10, 545–562. [Google Scholar] [CrossRef]
- Slik, J.; Aiba, S.I.; Brearley, F.Q.; Cannon, C.H.; Forshed, O.; Kitayama, K.; Nagamasu, H.; Nilus, R.; Payne, J.; Paoli, G.; et al. Environmental correlates of tree biomass, basal area, wood specific gravity and stem density gradients in Borneo’s tropical forests. Glob. Ecol. Biogeogr. 2010, 19, 50–60. [Google Scholar] [CrossRef]
- Favier, C.; De Namur, C.; Dubois, M.A. Forest progression modes in littoral Congo, central Atlantic Africa. J. Biogeogr. 2004, 31, 1445–1461. [Google Scholar] [CrossRef]
- Schwartz, D.; Lanfranchi, R.; Mariotti, A. Origine et évolution des savanes intramayombiennes (RP du Congo). I. Apports de la pédologie et de la biogéochimie isotopique (13C and 14C). In Paysages Quaternaires de l’Afrique Centrale Atlantique; ORSTOM: Paris, France, 1990; pp. 314–325. [Google Scholar]
- Schwartz, D.; De Foresta, H.; Mariotti, A.; Balesdent, J.; Massimba, J.; Girardin, C. Present dynamics of the savanna-forest boundary in the Congolese Mayombe: A pedological, botanical and isotopic (13C and 14C) study. Oecologia 1996, 106, 516–524. [Google Scholar] [CrossRef]
- Delègue, M.A.; Fuhr, M.; Schwartz, D.; Mariotti, A.; Nasi, R. Recent origin of a large part of the forest cover in the Gabon coastal area based on stable carbon isotope data. Oecologia 2001, 129, 106–113. [Google Scholar] [CrossRef]
- Mitchard, E.T.; Flintrop, C.M. Woody encroachment and forest degradation in sub-Saharan Africa’s woodlands and savannas 1982–2006. Philos. Trans. R. Soc. B 2013, 368, 20120406. [Google Scholar] [CrossRef]
- Letouzey, R. Notice de la Carte Phytogèographique du Cameroun au 1: 500,000 (map); Institut de la Recherche Agronomique (Herbier National): Yaoundeé, Cameroun, 1985. [Google Scholar]
- Egler, F.E. Vegetation science concepts I. Initial floristic composition, a factor in old-field vegetation development. Vegetatio 1954, 4, 412–417. [Google Scholar] [CrossRef]
- Wilson, J.B.; Gitay, H.; Roxburgh, S.H.; McG, W.; Tangney, R.S. Egler’s concept of ‘initial floristic composition’ in succession: Ecologists citing it don’t agree what it means. Oikos 1992, 64, 591–593. [Google Scholar] [CrossRef]
- Finegan, B. Forest succession. Nature 1984, 312, 109–114. [Google Scholar] [CrossRef]
- Kotar, J. Approaches to Ecologically Based Forest Management on Private Lands; Minnesota Extension Service; University of Minnesota: Minneapolis, MN, USA, 1997. [Google Scholar]
- Wilson, J.B. Does the Initial Floristic Composition model of succession really work? J. Veg. Sci. 2014, 25, 4–5. [Google Scholar] [CrossRef]
- Connell, J.H.; Slatyer, R.O. Mechanisms of succession in natural communities and their role in community stability and organization. Am. Nat. 1977, 111, 1119–1144. [Google Scholar] [CrossRef]
- Wierman, C.A.; Oliver, C.D. Crown stratification by species in even-aged mixed stands of Douglas-fir–western hemlock. Can. J. For. Res. 1979, 9, 1–9. [Google Scholar] [CrossRef]
- Larson, B.C. Development and growth of even-aged stands of Douglas-fir and grand fir. Can. J. For. Res. 1986, 16, 367–372. [Google Scholar] [CrossRef]
- McCarthy, J. Gap dynamics of forest trees: A review with particular attention to boreal forests. Environ. Rev. 2001, 9, 1–59. [Google Scholar] [CrossRef]
- Hubau, W.; De Mil, T.; Van den Bulcke, J.; Phillips, O.L.; Ilondea, B.A.; Van Acker, J.; Sullivan, M.J.; Nsenga, L.; Toirambe, B.; Couralet, C.; et al. The persistence of carbon in the African forest understory. Nat. Plants 2019, 5, 133–140. [Google Scholar] [CrossRef]
- Baker, P.J.; Bunyavejchewin, S.; Oliver, C.D.; Ashton, P.S. Disturbance history and historical stand dynamics of a seasonal tropical forest in western Thailand. Ecol. Monogr. 2005, 75, 317–343. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metric | Gilbertiodendron | Transitional | Mixed | Riparian |
|---|---|---|---|---|
| Number of quadrats (census 3) (%) | 208 (32.5) | 73 (11.4) | 297 (46.4) | 62 (9.7) |
| Fisher’s alpha (census 3) | ||||
| Stem density (ha, census 3) | ||||
| Stem density change (stems ha y) | ||||
| Basal area (mha) | ||||
| Basal area change (mhay) | ||||
| Live AGB (Mg ha) | ||||
| Live AGB change (Mg hay) | ||||
| DBH growth (mm treey) | ||||
| Recruitment change (% y at 2001) | ||||
| Recruitment change (Odds y) | ||||
| Mortality change (% y at 2001) | ||||
| Mortality change (Odds y) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glick, H.B.; Umunay, P.M.; Makana, J.-R.; Thomas, S.C.; Reuning-Scherer, J.D.; Gregoire, T.G. Developmental Dynamics of Gilbertiodendron dewevrei (Fabaceae) Drive Forest Structure and Biomass in the Eastern Congo Basin. Forests 2021, 12, 738. https://0-doi-org.brum.beds.ac.uk/10.3390/f12060738
Glick HB, Umunay PM, Makana J-R, Thomas SC, Reuning-Scherer JD, Gregoire TG. Developmental Dynamics of Gilbertiodendron dewevrei (Fabaceae) Drive Forest Structure and Biomass in the Eastern Congo Basin. Forests. 2021; 12(6):738. https://0-doi-org.brum.beds.ac.uk/10.3390/f12060738
Chicago/Turabian StyleGlick, Henry B., Peter M. Umunay, Jean-Remy Makana, Sean C. Thomas, Jonathan D. Reuning-Scherer, and Timothy G. Gregoire. 2021. "Developmental Dynamics of Gilbertiodendron dewevrei (Fabaceae) Drive Forest Structure and Biomass in the Eastern Congo Basin" Forests 12, no. 6: 738. https://0-doi-org.brum.beds.ac.uk/10.3390/f12060738