
Habitat-Related Differences in Winter Presence and Spring–Summer Activity of Roe Deer in Warsaw

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
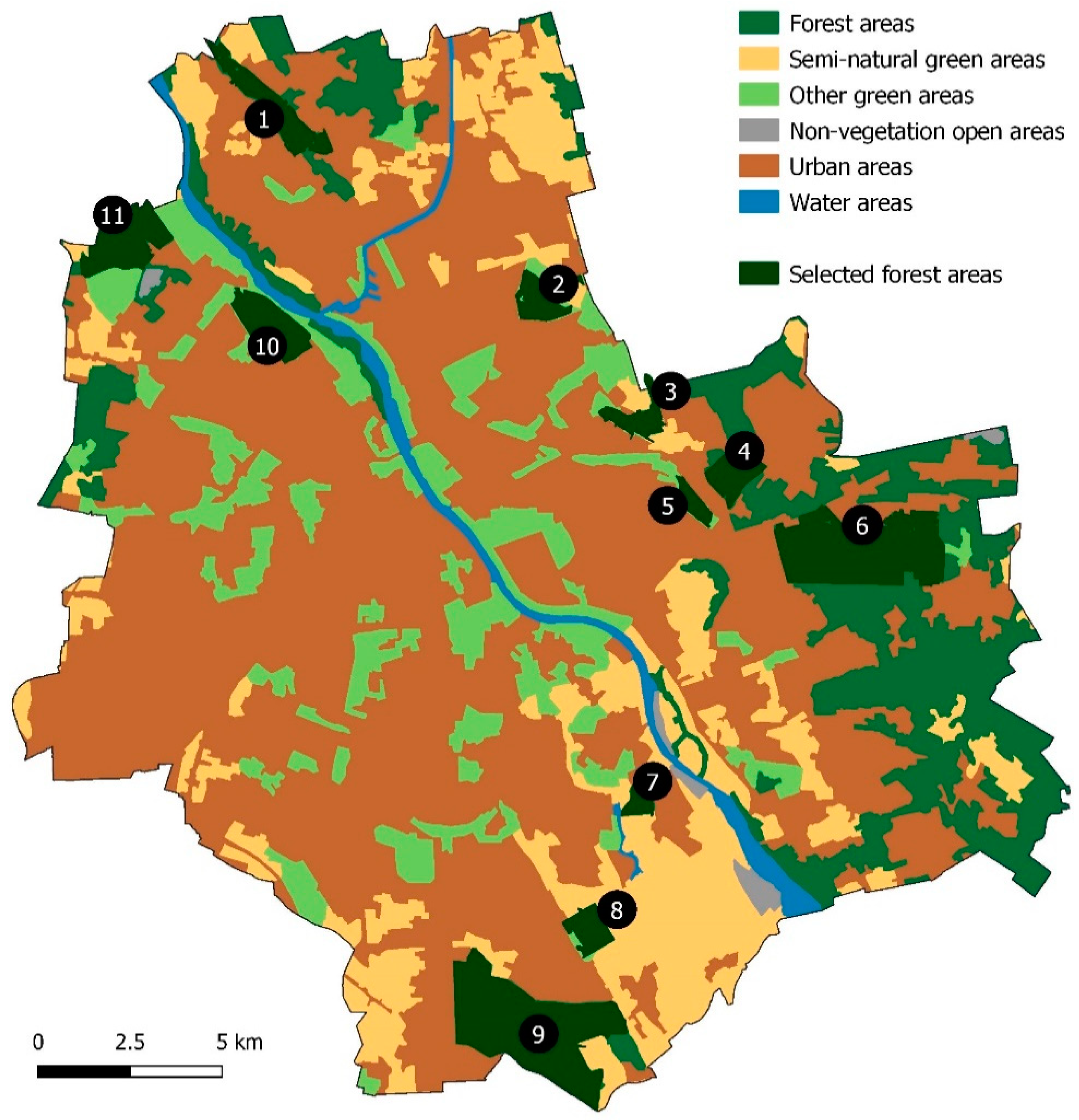
2.1. Study Area
2.2. Data Collection
2.3. Statistical Analysis
3. Results
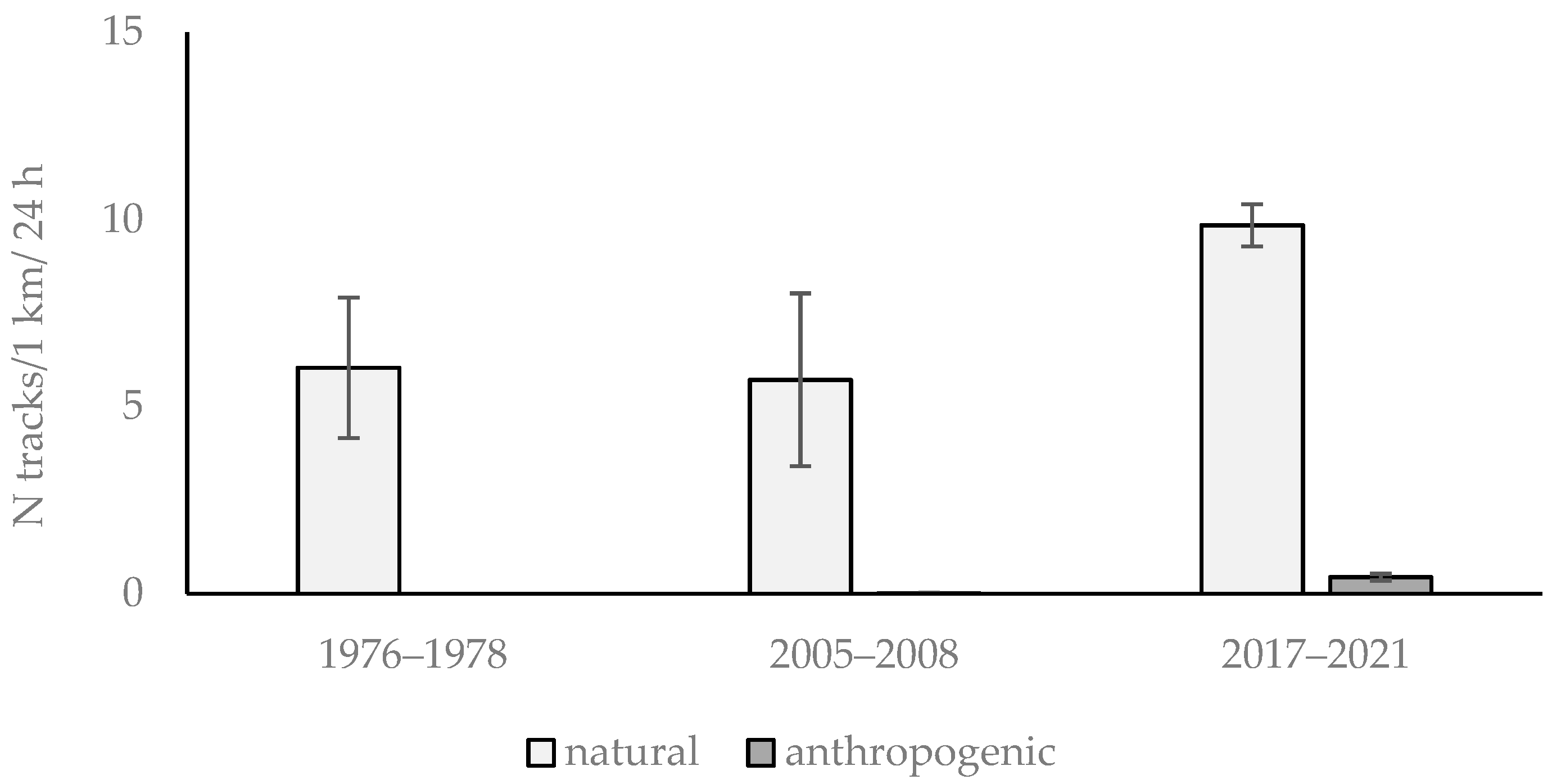
3.1. Abundance of Roe Deer in Different Habitats of Warsaw Using Snow Tracking Data
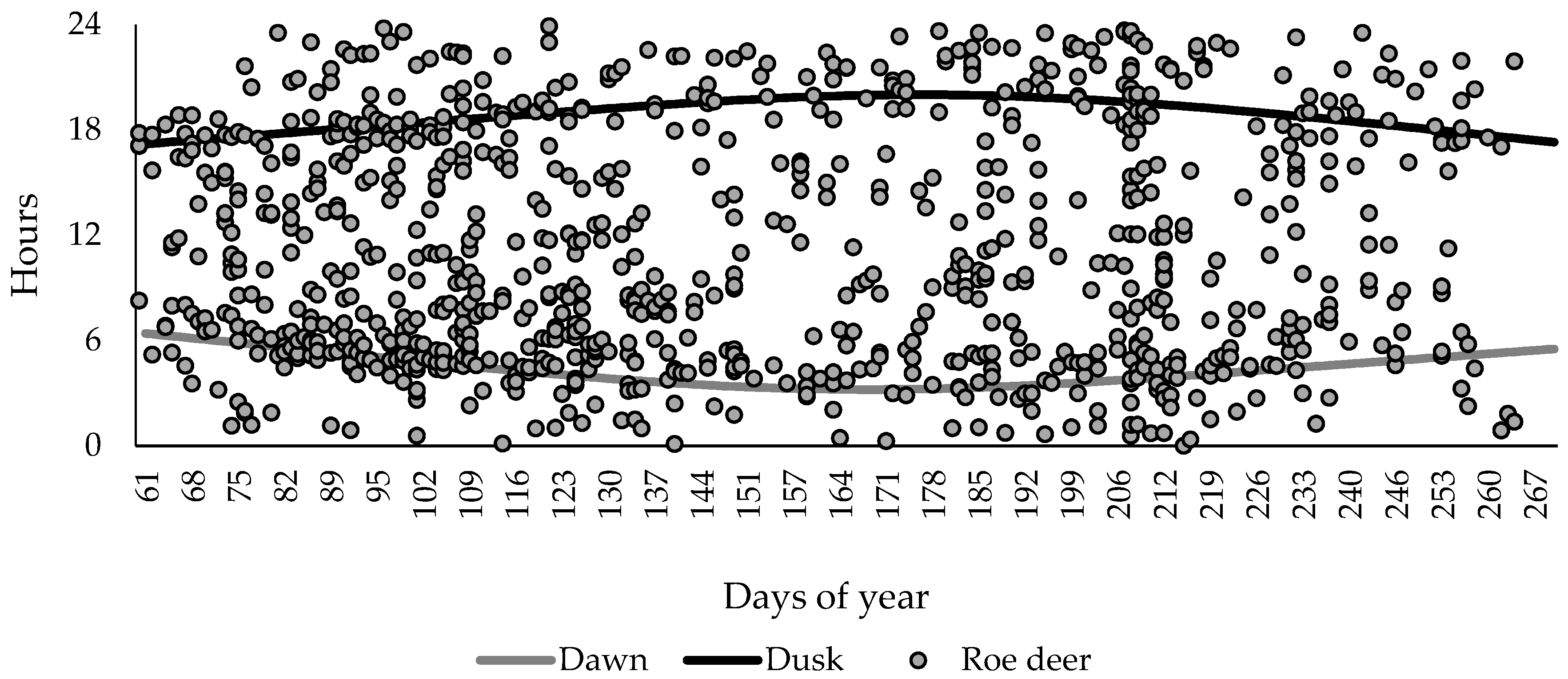
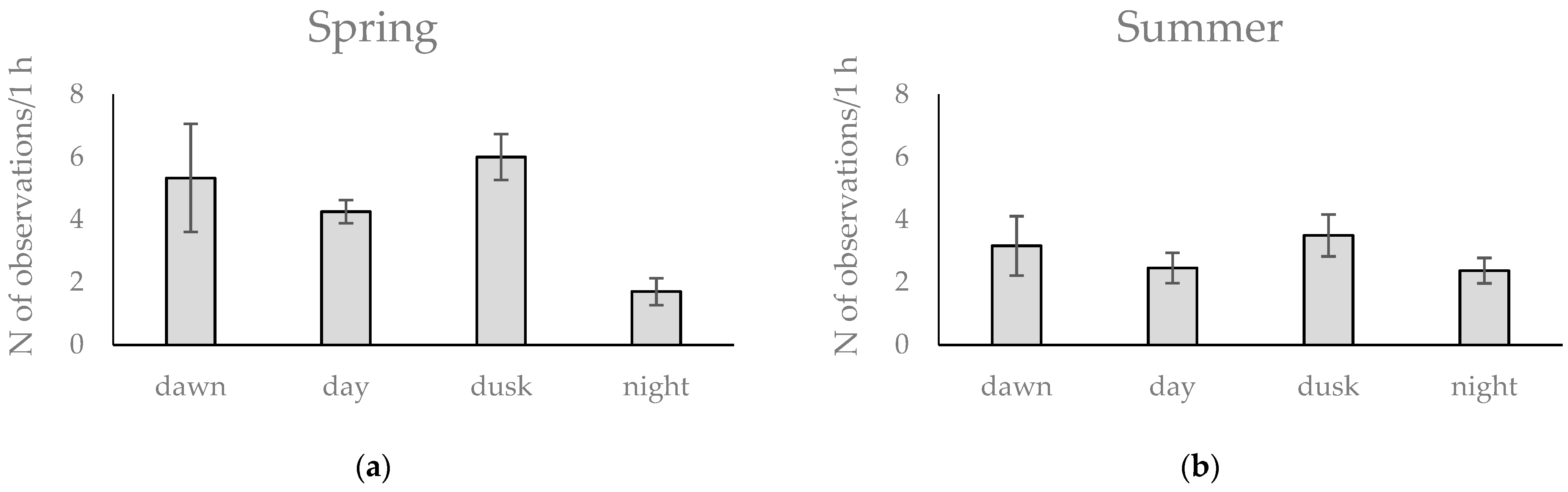
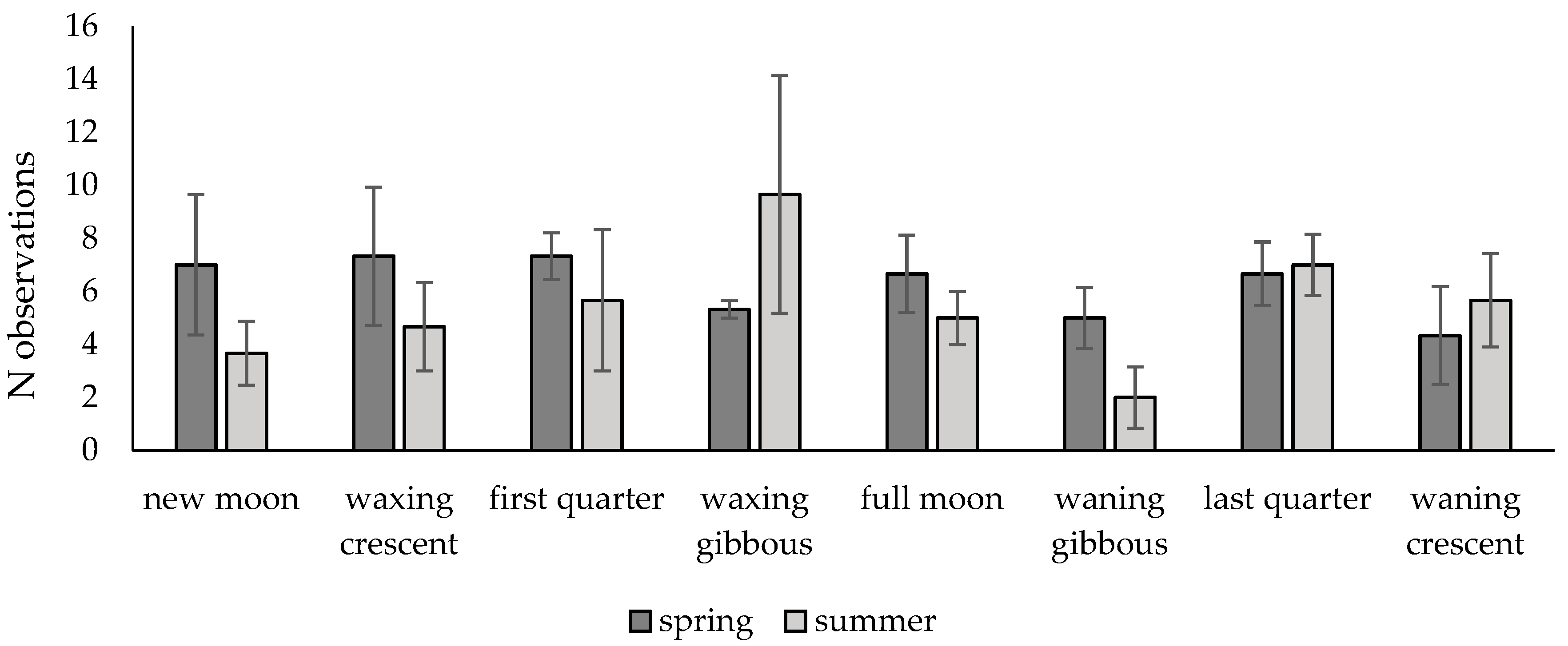
3.2. Activity Patterns of Roe Deer in Warsaw Urban Forests Using Camera Traps Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number of Urban Forest | Urban Forest | Forest Area (ha) | Number of Location | Latitude | Longitude | Dates of Camera Trap Exposition |
|---|---|---|---|---|---|---|
| 1 | Henryków i Dąbrówka Forest Park | 201.37 | 1 | 52°20.349′ N | 20°58.256′ E | 02.03.–27.06.2019 |
| 2 | 52°20.362′ N | 20°58.209′ E | 02.03.–06.05.2019 20.04.–13.07.2020 | |||
| 3 | 52°21.086′ N | 20°57.385′ E | 02.03.–01.05.2019 17.06.–29.09.2019 25.02.–04.04.2020 | |||
| 4 | 52°20.268′ N | 20°58.275′ E | 07.03.–20.04.2020 | |||
| 2 | Bródno Forest Park | 139.00 | 1 | 52°17.909′ N | 21°04.015′ E | 24.02.–20.05.2020 |
| 2 | 52°18.037′ N | 21°03.924′ E | 02.03.–07.05.2019 24.02.–14.07.2020 | |||
| 3 | 52°18.156′ N | 21°03.787′ E | 02.03.–07.05.2019 | |||
| 3 | Utrata Forest | 102.90 | 1 | 52°16.057′ N | 21°06.698′ E | 05.03.–04.06.2019 |
| 4 | Kawęczyn Nature Reserve | 123.18 | 1 | 52°15,249′ N | 21°08,733′ E | 05.03.–12.03.2019 23.02.–21.04.2020 |
| 2 | 52°15,242′ N | 21°08,285′ E | 10.03.–21.04.2020 | |||
| 5 | Olszynka Grochowska Nature Reserve | 58.91 | 1 | 52°14.896′ N | 21°07.301′ E | 18.03.–30.05.2019 18.06.–22.07.2019 |
| 2 | 52°14.873′ N | 21°07.328′ E | 12.05.–13.10.2020 | |||
| 6 | Sobieski Forest | 765.75 | 1 | 52°13.756′ N | 21°10.215′ E | 09.03.–01.06.2019 18.02.–07.07.2020 |
| 2 | 52°13.746′ N | 21°10.163′ E | 09.03.–07.05.2019 21.04.–07.07.2020 | |||
| 3 | 52°14.020′ N | 21°09.664′ E | 01.03.–17.07.2019 | |||
| 4 | 52°13.941′ N | 21°10.550′ E | 07.05.–04.10.2019 18.02.–03.03.2020 | |||
| 5 | 52°13.599′ N | 21°10.388′ E | 18.02.–01.03.2020 | |||
| 6 | 52°13.744′ N | 21°10.214′ E | 08.03.–21.04.2020 | |||
| 7 | Morysin Nature Reserve | 45.04 | 1 | 52°10.508′ N | 21°06.134′ E | 09.03.–25.04.2019 15.07.–05.09.2019 09.02.–23.02.2020 14.09.–22.10.2020 |
| 2 | 52°10.529′ N | 21°06.117′ E | 29.02.–11.05.2020 | |||
| 8 | Natolin Forest Nature Reserve | 104.16 | 1 | 52°08.459′ N | 21°05.092′ E | 11.03.–14.05.2019 22.07.–31.12.2019 10.02.–29.06.2020 |
| 2 | 52°08.611′ N | 21°04.223′ E | 27.02.–11.03.2019 29.04.–11.06.2019 09.03.–22.04.2020 | |||
| 3 | 52°08.421′ N | 21°05.018′ E | 29.04.–22.07.2019 | |||
| 4 | 52°08.379′ N | 21°05.206′ E | 05.06.–07.07.2020 | |||
| 9 | Kabaty Nature Reserve | 918.27 | 1 | 52°06.985′ N | 21°03.229′ E | 03.03.–27.04.2020 |
| 2 | 52°07.455′ N | 21°03.054′ E | 26.02.–11.04.2019 18.05.–01.09.2019 27.04.–24.10.2020 | |||
| 3 | 52°07.753′ N | 21°02.823′ E | 04.03.–11.03.2019 19.05.–08.07.2019 | |||
| 4 | 52°08.007′ N | 21°02.185′ E | 11.03.–07.04.2019 08.07.–15.08.2019 | |||
| 5 | 52°07.305′ N | 21°05.380′ E | 17.03.–31.12.2019 07.01.–08.05.2020 | |||
| 6 | 52°07.493′ N | 21°02.105′ E | 26.02.–04.03.2019 | |||
| 7 | 52°07.592′ N | 21°03.118′ E | 26.02.–17.06.2019 08.07.–28.09.2019 03.03.–24.10.2020 | |||
| 8 | 52°06.916′ N | 21°03.231′ E | 09.06.–24.10.2020 | |||
| 9 | 52°07.550′ N | 21°02.105′ E | 14.03.–19.05.2019 24.04.–26.10.2020 | |||
| 10 | 52°07.567′ N | 21°01.944′ E | 19.05.–28.09.2019 24.04.–08.06.2020 | |||
| 11 | 52°06.878′ N | 21°02.499′ E | 09.03.–24.04.2020 | |||
| 12 | 52°07.384′ N | 21°05.316′ E | 09.03.–24.04.2020 | |||
| 13 | 52°06.842′ N | 21°02.806′ E | 09.03.–24.04.2020 | |||
| 14 | 52°06.912′ N | 21°02.416′ E | 24.04.–09.06.2020 | |||
| 10 | Bielany Forest | 196.07 | 1 | 52°17.349′ N | 20°58.057′ E | 10.03.–22.09.2020 |
| 11 | Młociny (Nowa Warszawa) Forest | 293.64 | 1 | 52°18.864′ N | 20°54.201′ E | 02.03.–18.06.2019 02.03.–10.03.2020 |
| 2 | 52°19.114′ N | 20°54.025′ E | 02.03.–06.05.2019 |
Appendix B
| Urban Forest | Level of Light Pollution |
|---|---|
| Henryków i Dąbrówka Forest Park | 10.45 |
| Bródno Forest Park | 16.38 |
| Utrata Forest | 16.00 |
| Kawęczyn Nature Reserve | 12.63 |
| Olszynka Grochowska Nature Reserve | 9.88 |
| Sobieski Forest | 24.64 |
| Morysin Nature Reserve | 6.37 |
| Natolin Forest Nature Reserve | 15.22 |
| Kabaty Nature Reserve | 5.23 |
| Bielany Forest | 17.18 |
| Henryków i Dąbrówka Forest Park | 10.45 |
Appendix C
| Urban Forest | % Share in 250 m Buffer of ** | |||||
|---|---|---|---|---|---|---|
| Forest Areas | Semi-Natural Green Areas | Other Green Areas | Non-Vegetation Open Areas | Urban Areas | Water | |
| Henryków i Dąbrówka Forest Park | 19.5 | 7.0 | - | - | 73.5 | - |
| Bródno Forest Park | - | 15.5 | 25.3 | - | 59.2 | - |
| Utrata Forest | - | 25.9 | 20.9 | - | 53.2 | - |
| Kawęczyn Nature Reserve | 40.9 | - | - | - | 59.1 | - |
| Olszynka Grochowska Nature Reserve | - | - | 21.0 | - | 79.0 | - |
| Sobieski Forest | 49.4 | - | 1.3 | - | 49.3 | - |
| Morysin Nature Reserve | 5.6 | 39.8 | 1.6 | - | 41.8 | 11.2 |
| Natolin Forest Nature Reserve | - | 52.3 | 16.2 | - | 31.4 | - |
| Kabaty Nature Reserve | 8.7 | 34.5 | - | - | 56.8 | - |
| Bielany Forest | 1.8 | - | 37.4 | - | 48.8 | 12.0 |
| Młociny (Nowa Warszawa) Forest | 23.2 | 9.4 | 29.8 | 1.9 | 35.6 | - |
Appendix D
| Analysis | Parameter | r | p |
|---|---|---|---|
| share (%) in 250 m buffer around urban forests | Forest areas | 0.13 | 0.71 |
| Semi-natural green areas | 0.34 | 0.31 | |
| Other green areas | −0.43 | 0.19 | |
| Non-vegetation open areas | −0.44 | 0.18 | |
| Urban areas | −0.02 | 0.95 | |
| Water | −0.22 | 0.52 |
References
- McKinney, M.L. Urbanization as a major cause of biotic homogenization. Biol. Conserv. 2006, 127, 247–260. [Google Scholar] [CrossRef]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Natl. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [Green Version]
- Prugh, L.R.; Hodges, K.E.; Sinclair, A.R.E.; Brashares, J.S. Effect of habitat area and isolation on fragmented animal populations. Proc. Natl. Acad. Sci. USA 2008, 105, 20770–20775. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McDonnell, M.J.; Hahs, A.K. The use of gradient analysis studies in advancing our understanding of the ecology of urbanizing landscapes: Current status and future directions. Landsc. Ecol. 2008, 23, 1143–1155. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; Cadenasso, M.L.; Grove, J.M.; Boone, C.G.; Groffman, P.M.; Irwin, E.; Kaushal, S.S.; Marshall, V.; McGrath, B.P.; Nilon, C.H.; et al. Urban ecological systems: Scientific foundations and a decade of progress. J. Environ. Manag. 2011, 92, 331–362. [Google Scholar] [CrossRef] [PubMed]
- Forman, R.T. Urban Ecology: Science of Cities; Cambridge University Press: New York, NY, USA, 2014. [Google Scholar]
- Parris, K.M. Ecology of Urban Environments; Wiley-Blackwell: Chichester, UK, 2016. [Google Scholar]
- Hewison, A.J.M.; Vincent, J.P.; Joachim, J.; Angibault, J.M.; Cargnelutti, B.; Cibien, C. The effects of woodland fragmentation and human activity on roe deer distribution in agricultural landscapes. Can. J. Zool. 2001, 79, 679–689. [Google Scholar] [CrossRef]
- Acevedo, P.; Delibes-Mateos, M.; Escudero, M.A.; Vicente, J.; Marco, J.; Gortazar, C. Environmental constraints in the colonization sequence of roe deer (Capreolus capreolus Linnaeus, 1758) across the Iberian Mountains, Spain. J. Biogeogr. 2005, 32, 1671–1680. [Google Scholar] [CrossRef]
- Syphard, A.D.; Clarke, K.C.; Franklin, J.; Regan, H.M.; McGinnis, M. Forecasts of habitat loss and fragmentation due to urban growth are sensitive to source of input data. J. Environ. Manag. 2011, 92, 1882–1893. [Google Scholar] [CrossRef]
- Scolozzi, R.; Geneletti, D. A multi-scale qualitative approach to assess the impact of urbanization on natural habitats and their connectivity. Environ. Impact Assess. Rev. 2012, 36, 9–22. [Google Scholar] [CrossRef]
- van Strien, M.J.; Grêt-Regamey, A. How is habitat connectivity affected by settlement and road network configurations? Results from simulating coupled habitat and human networks. Ecol. Model. 2016, 342, 186–198. [Google Scholar] [CrossRef] [Green Version]
- van Strien, M.J.; Slager, C.T.J.; de Vries, B.; Gret-Regamey, A. An improved neutral landscape model for re-creating real landscapes and generating landscape series for spatial ecological simulations. Ecol. Evol. 2016, 6, 3808–3821. [Google Scholar] [CrossRef] [Green Version]
- Baker, P.J.; Dowding, C.V.; Molony, S.E.; White, P.C.L.; Harris, S. Activity patterns of urban red foxes (Vulpes vulpes) reduce the risk of traffic-induced mortality. Behav. Ecol. 2007, 35, 716–724. [Google Scholar] [CrossRef] [Green Version]
- Harris, S. Ecology of Urban Badger Meles meles: Distribution in Britain and Habitat Selection, Persecution, Food and Damage in the City of Bristol. Biol. Conserv. 1984, 28, 349–375. [Google Scholar] [CrossRef]
- Herr, J.; Schley, L.; Roper, T.J. Socio-spatial organization of urban stone martens. J. Zool. 2009, 277, 54–62. [Google Scholar] [CrossRef]
- Jackowiak, M.; Gryz, J.; Jasińska, K.; Brach, M.; Bolibok, L.; Kowal, P.; Krauze-Gryz, D. Colonization of Warsaw by the red fox Vulpes vulpes in the years 1976–2019. Sci. Rep. 2021, 11, 13931. [Google Scholar] [CrossRef]
- Csókás, A.; Schally, G.; Szabó, L.; Csányi, S.; Kovács, F.; Heltai, M. Space use of wild boar (Sus Scrofa) in Budapest: Are they resident or transient city dwellers? Biol. Futur. 2020, 71, 39–51. [Google Scholar] [CrossRef] [Green Version]
- Frid, A.; Dill, L.M. Human-caused disturbance stimuli as a form of predation risk. Conserv. Ecol. 2002, 6, 11. [Google Scholar] [CrossRef]
- Rich, C.; Longcore, T. Ecological Consequences of Artificial Night Lighting; Island Press: London, UK, 2006. [Google Scholar]
- LaPoint, S.; Balkenhol, N.; Hale, J.; Sadler, J.; van der Ree, R. Ecological connectivity research in urban areas. Funct. Ecol. 2015, 29, 868–878. [Google Scholar] [CrossRef]
- Ciuti, S.; Northrup, J.M.; Muhly, T.B.; Simi, S.; Musiani, M.; Pitt, J.A.; Boyce, M.S. Effects of humans on behaviour of wildlife exceed those of natural predators in a landscape of fear. PLoS ONE 2012, 7, e50611. [Google Scholar]
- Bonnot, N.; Morellet, N.; Verheyden, H. Habitat use under predation risk: Hunting, roads and human dwellings influence the spatial behavior of roe deer. Eur. J. Wildl. Res. 2013, 59, 185–193. [Google Scholar] [CrossRef]
- Bonnot, N.; Morellet, N.; Hewison, A.J.M.; Martin, J.-L.; Benhamou, S.; Chamaillé-Jammes, S. Black-tailed deer (Odocoileus hemionus sitkensis) adjust habitat selection and activity rhythm to the absence of predators. Can. J. Zool. 2016, 94, 385–394. [Google Scholar] [CrossRef]
- Wevers, J.; Fattebert, J.; Casaer, J.; Artois, T.; Beenaerts, N. Trading fear for food in the Anthropocene: How ungulates cope with human disturbance in a multi-use, suburban ecosystem. Sci. Total Environ. 2020, 741, 140369. [Google Scholar] [CrossRef] [PubMed]
- Bragina, E.V.; Ives, A.R.; Pidgeon, A.M.; Balčiauskas, L.; Csányi, S.; Khoyetskyy, P.; Radeloff, V.C. Wildlife population changes across Eastern Europe after the collapse of socialism. Front. Ecol. Environ. 2018, 16, 77–81. [Google Scholar] [CrossRef] [Green Version]
- Putman, R.J. Foraging by roe deer in agricultural areas and impact on arable crops. J. Appl. Ecol. 1986, 23, 91–99. [Google Scholar] [CrossRef]
- McLoughlin, P.D.; Gaillard, J.-M.; Boyce, M.S.; Bonenfant, C.; Messier, F.; Duncan, P.; Delorme, D.; Van Moorter, B.; Saïd, S.; Klein, F. Lifetime reproductive success and composition of the home range in a large herbivore. Ecology 2007, 88, 3192–3201. [Google Scholar] [CrossRef]
- Panzacchi, M.; Linnell, J.D.C.; Odden, M.; Odden, J.; Andersen, R. Habitat and roe deer fawn vulnerability to red fox predation. J. Anim. Ecol. 2009, 78, 1124–1133. [Google Scholar] [CrossRef]
- Tinoco Torres, R.; Carvahlo, J.C.; Panzacchi, M.; Linnell, J.D.C.; Fonseca, C. Comparative use of forest habitats by roe deer and moose in a human-modified landscape in southeastern Norway during winter. Ecol. Res. 2011, 26, 781–789. [Google Scholar] [CrossRef]
- Loro, M.; Ortega, E.; Arce, R.M.; Geneletti, D. Assessing landscape resistance to Roe deer dispersal using fuzzy set theory and multicriteria analysis: A case study in Central Spain. Landsc. Ecol. Eng. 2016, 12, 41–60. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, A.; Baker, A.; Rotherham, I. Urban-fringe deer management issues—A South Yorkshire case study. Br. Wildl. 1996, 8, 12–19. [Google Scholar]
- Statistics Poland. Statistical Yearbook of Warsaw; Zakład Wydawnictw Statystycznych: Warsaw, Poland, 2019. [Google Scholar]
- Climate-data.org. Available online: https://pl.climate-data.org/europa/polska/masovian-voivodeship/warszawa-4560/ (accessed on 29 October 2020).
- Luniak, M.; Kozłowski, P.; Nowicki, W. Magpie Pica pica in Warsaw—Abundance, distribution and changes in its population. Acta Ornithol. 1997, 32, 77–86. [Google Scholar]
- Büttner, G.; Kosztra, B.; Maucha, G.; Pataki, R.; Kleeschulte, S.; Hazeu, G.; Vittek, M.; Schröder, C.; Littkopf, A. Copernicus Land Monitoring Service CORINE Land Cover. User Manual. Copernicus Land Monitoring Service (CLMS). 2021. Available online: https://land.copernicus.eu/user-corner/technical-library/clc-product-user-manual?fbclid=IwAR2ZURZU-H1l5J45FSSZfGKSSHlp2AB0-i50pImfEB20ZR2iEUmFHvua7uc (accessed on 25 June 2021).
- Krop-Benesch, A.; Berger, A.; Hofer, H.; Heurich, M. Long-term measurement of roe deer (Capreolus capreolus) (Mammalia: Cervidae) activity using two-axis accelerometers in GPS-collars. Ital. J. Zool. 2013, 80, 69–81. [Google Scholar] [CrossRef] [Green Version]
- Dateandtime.info. Available online: https://dateandtime.info/pl/citysunrisesunset.php?id=756135&month=8&year=2019 (accessed on 29 October 2020).
- Caravaggi, A.; Gatta, M.; Vallely, M.; Hogg, K.; Freeman, M.; Fadaei, E.; Dick, J.T.A.; Montgomery, W.I.; Reid, N.; Tosh, D.G. Seasonal and predator-prey effects on circadian activity of free-ranging mammals revealed by camera traps. Peer J. 2018, 6, e5827. [Google Scholar] [CrossRef]
- Corsini, M.T.; Lovari, S.; Sonnino, S. Temporal activity patterns of crested porcupines Hystvix cvistata. J. Zool. 1995, 236, 43–54. [Google Scholar] [CrossRef]
- lightpollutionmap.info. Available online: https://www.lightpollutionmap.info/#zoom=4.00&lat=45.8720&lon=14.5470&layers=B0FFFFFTFFFFFFFFFF (accessed on 23 June 2021).
- Bielecka, E.; Jenerowicz, A. Intellectual structure of CORINE Land Cover research applications in Web of Science: A Europe-wide review. Remote Sens. 2019, 11, 2017. [Google Scholar] [CrossRef] [Green Version]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 4. [Google Scholar]
- Ciach, M.; Fröhlich, A. Ungulates in the city: Light pollution and open habitats predict the probability of roe deer occurring in an urban environment. Urban Ecosyst. 2019, 22, 513–523. [Google Scholar] [CrossRef] [Green Version]
- Bonnot, N.C.; Couriot, O.; Berger, A.; Cagnacci, F.; Ciuti, S.; de Groeve, J.E.; Gehr, B.; Heurich, M.; Kjellander, P.; Kröschel, M.; et al. Fear of the dark? Contrasting impacts of humans versus lynx on diel activity of roe deer across Europe. J. Anim. Ecol. 2020, 89, 132–145. [Google Scholar] [CrossRef]
- Laundré, J.W.; Hernández, L.; Altendorf, K.B. Wolves, elk, and bison: Reestablishing the "landscape of fear" in Yellowstone National Park, U.S.A. Can. J. Zool. 2001, 79, 1401–1409. [Google Scholar] [CrossRef]
- Laundré, J.W.; Hernández, L.; Ripple, W.J. The landscape of fear: Ecological implications of being afraid. Open Ecol. J. 2010, 3, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Benhaiem, S.; Delon, M.; Lourtet, B.; Cargnelutti, B.; Aulagnier, S.; Hewison, A.J.M.; Morellet, N.; Verheyden, H. Hunting increases vigilance levels in roe deer and modifies feeding site selection. Anim. Behav. 2008, 76, 611–618. [Google Scholar] [CrossRef]
- Bonnot, N.C.; Hewison, A.J.M.; Morellet, N.; Gaillard, J.-M.; Debeffe, L.; Couriot, O.; Cargnelutti, B.; Chaval, Y.; Lourtet, B.; Kjellander, P.; et al. Stick or twist: Roe deer adjust their flight behaviour to the perceived trade-off between risk and reward. Anim. Behav. 2017, 124, 35–46. [Google Scholar] [CrossRef]
- Mysterud, A.; Lian, L.-B.; Hjermann, D.Ø. Scale-dependent trade-offs in foraging by European roe deer (Capreolus capreolus) during winter. Can. J. Zool. 1999, 77, 1486–1493. [Google Scholar] [CrossRef] [Green Version]
- Jepsen, J.U.; Topping, C.J. Modelling roe deer (Capreolus capreolus) in a gradient of forest fragmentation: Behavioural plasticity and choice of cover. Can. J. Zool. 2004, 82, 1528–1541. [Google Scholar] [CrossRef]
- Lone, K.; Loe, L.E.; Gobakken, T.; Linnell, J.D.C.; Odden, J.; Remmen, J.; Mysterud, A. Living and dying in a multi-predator landscape of fear: Roe deer are squeezed by contrasting pattern of predation risk imposed by lynx and humans. Oikos 2014, 123, 641–651. [Google Scholar] [CrossRef]
- Kays, R.; Crofoot, M.C.; Jetz, W.; Wikelski, M. Terrestrial animal tracking as an eye on life and planet. Science 2015, 348, aaa2478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larson, C.L.; Reed, S.E.; Merenlender, A.M.; Crooks, K.R. Effects of recreation on animals revealed as widespread through a global systematic review. PLoS ONE 2016, 11, e0167259. [Google Scholar] [CrossRef] [Green Version]
- Oberosler, V.; Groff, C.; Iemma, A.; Pedrini, P.; Rovero, F. The influence of human disturbance on occupancy and activity patterns of mammals in the Italian Alps from systematic camera trapping. Mamm. Biol. 2017, 87, 50–61. [Google Scholar] [CrossRef]
- Pagon, N.; Grignolio, S.; Pipia, A.; Bongi, P.; Bertolucci, C.; Apollonio, M. Seasonal variation of activity patterns in roe deer in a temperate forested area. Chronobiol. Int. 2013, 30, 772–785. [Google Scholar] [CrossRef] [PubMed]
- Sönnichsen, L.; Bokje, M.; Marchal, J.; Hofer, H.; Jędrzejewska, B.; Kramer-Schadt, S.; Ortmann, S. Behavioural responses of european Roe deer to temporal variation in predation risk. Ethology 2013, 199, 233–243. [Google Scholar] [CrossRef]
- Ditchkoff, S.S.; Saalfeld, S.T.; Gibson, C.J. Animal behavior in urban ecosystems: Modifications due to human-induced stress. Urban Ecosyst. 2006, 9, 5–12. [Google Scholar] [CrossRef]
- Kotler, B.P.; Brown, J.; Mukherjee, S.; Berger-Tal, O.; Bouskila, A. Moonlight avoidance in gerbils reveals a sophisticated interplay among time allocation, vigilance and state-dependent foraging. Proc. R. Soc. B 2010, 277, 1469–1474. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Griffin, P.C.; Griffin, S.C.; Waroquiers, C.; Mills, L.S. Mortality by moonlight: Predation risk and the snowshoe hare. Behav. Ecol. 2005, 16, 938–944. [Google Scholar] [CrossRef] [Green Version]
- Haddock, J.K.; Threlfall, C.G.; Law, B.; Hochuli, D.F. Light pollution at the urban forest edge negatively impacts insectivorous bats. Biol. Conserv. 2019, 236, 17–28. [Google Scholar] [CrossRef]
- Prough, L.R.; Golden, C.D. Does moonlight increase predation risk? Meta-analysis reveals divergent responses of nocturnal mammals to lunar cycles. J. Anim. Ecol. 2014, 83, 504–514. [Google Scholar] [CrossRef] [PubMed]
- Kurt, F. Das Reh in der Kulturlandschaft: Sozialverhalten und Ökologie eines Anpassers Roe Deer in the Cultural Landscape: Social Behavior and Ecology of an Adaptor; Verlag Paul Parey: Hamburg/Berlin, Germany, 1991. [Google Scholar]





| Research Period | Length of Transect Routes in Habitats [km] | |
|---|---|---|
| Natural | Anthropogenic | |
| 1976–1978 | 96.2 | 79.5 |
| 2005–2008 | 39.0 | 180.8 |
| 2017–2021 | 146.0 | 139.7 |
| in total | 281.2 | 400.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jasińska, K.D.; Jackowiak, M.; Gryz, J.; Bijak, S.; Szyc, K.; Krauze-Gryz, D. Habitat-Related Differences in Winter Presence and Spring–Summer Activity of Roe Deer in Warsaw. Forests 2021, 12, 970. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080970
Jasińska KD, Jackowiak M, Gryz J, Bijak S, Szyc K, Krauze-Gryz D. Habitat-Related Differences in Winter Presence and Spring–Summer Activity of Roe Deer in Warsaw. Forests. 2021; 12(8):970. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080970
Chicago/Turabian StyleJasińska, Karolina D., Mateusz Jackowiak, Jakub Gryz, Szymon Bijak, Katarzyna Szyc, and Dagny Krauze-Gryz. 2021. "Habitat-Related Differences in Winter Presence and Spring–Summer Activity of Roe Deer in Warsaw" Forests 12, no. 8: 970. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080970