
Molecular-Based Reappraisal of a Historical Record of Dothistroma Needle Blight in the Centre of the Mediterranean Region

 , , , , and
, , , , and {kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Area and Plant Material
2.2. Isolation of Fungi
2.3. DNA Extraction from Mycelia and Pine Needles
2.4. LAMP on DNA Extracts from Needles
2.5. Conventional PCR and Sequencing on Mycelial DNA Extracts
3. Results
3.1. Symptoms, Signs and Conidial Micromorphology
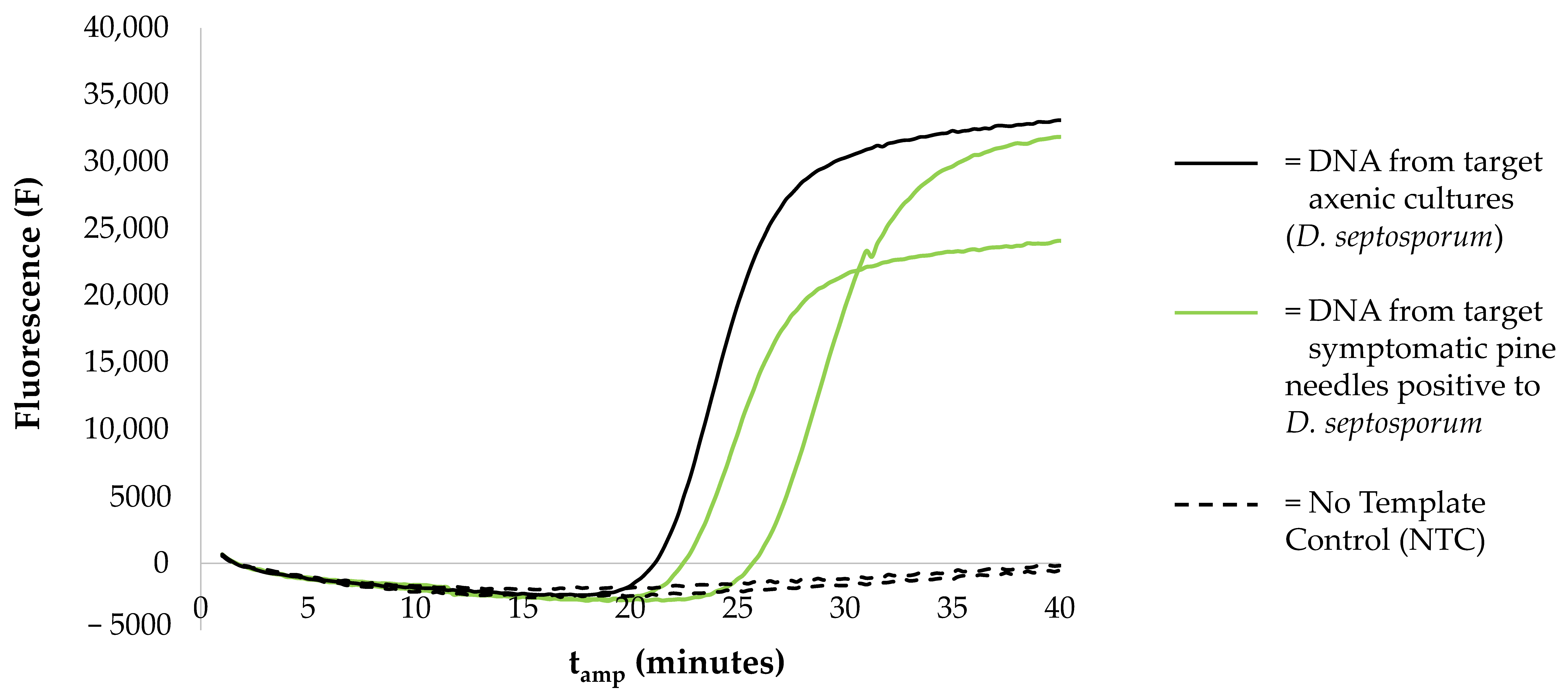
3.2. LAMP Assays
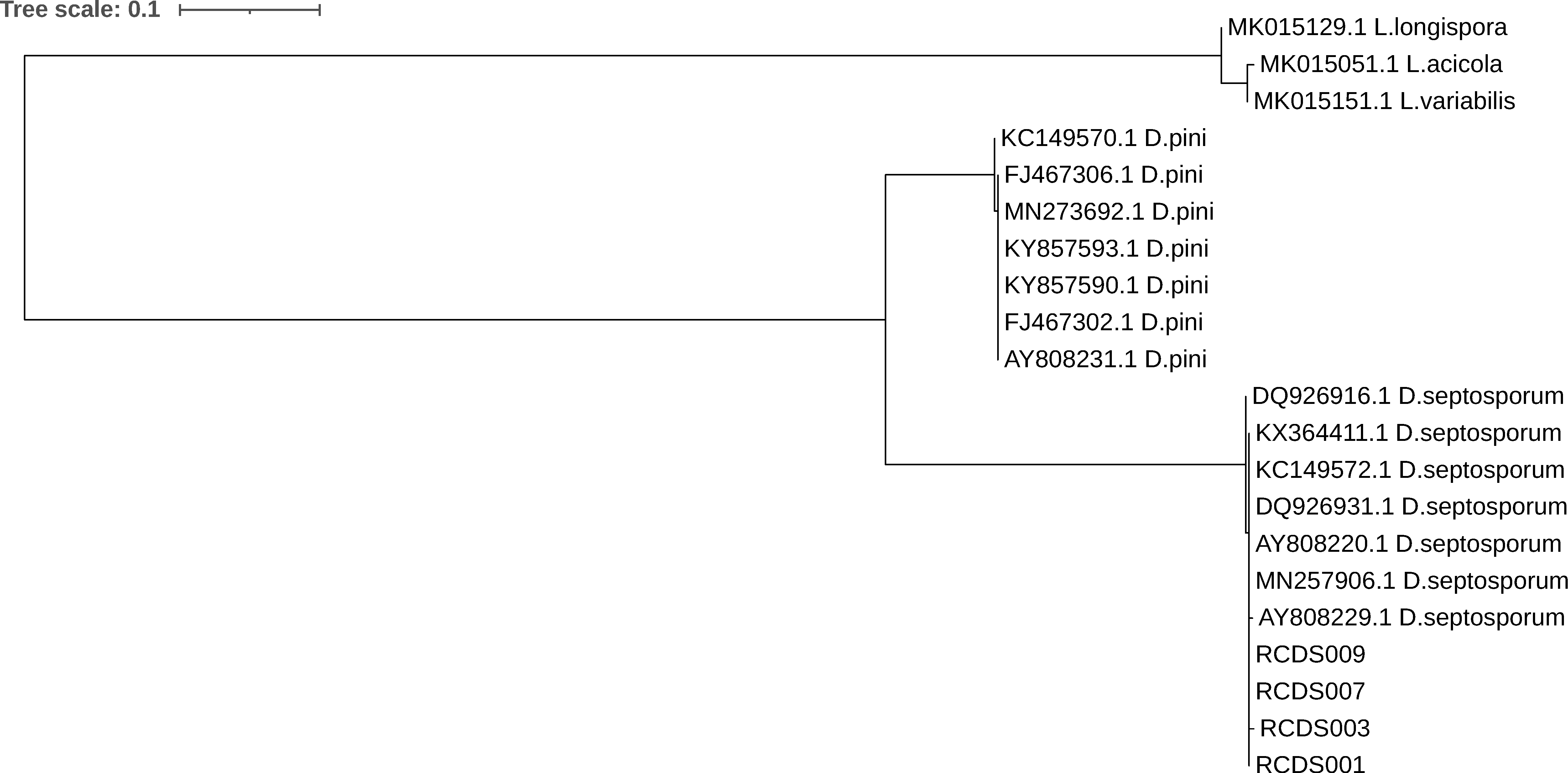
3.3. Isolation of Fungi and Identification by Sequencing
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Drenkhan, R.; Tomešová-Haataja, V.; Fraser, S.; Bradshaw, R.E.; Vahalik, P.; Mullett, M.S.; Martín-García, J.; Bulman, L.S.; Wingfield, M.J.; Kirisits, T.; et al. Global geographic distribution and host range of Dothistroma species: A comprehensive review. For. Pathol. 2016, 46, 408–442. [Google Scholar] [CrossRef]
- Mullett, M.S.; Adamson, K.; Bragança, H.; Bulgakov, T.S.; Georgieva, M.; Henriques, J.; Jürisoo, L.; Laas, M.; Drenkhan, R. New country and regional records of the pine needle blight pathogens Lecanosticta acicola, Dothistroma septosporum and Dothistroma pini. For. Pathol. 2018, 48, e12440. [Google Scholar] [CrossRef]
- Barnes, I.; van der Nest, A.; Mullett, M.S.; Crous, P.W.; Drenkhan, R.; Musolin, D.L.; Wingfield, M.J. Neotypification of Dothistroma septosporum and epitypification of D. pini, causal agents of Dothistroma needle blight of pine. For. Pathol. 2016, 46, 388–407. [Google Scholar] [CrossRef]
- Gibson, I.A.S. Dothistroma blight of Pinus radiata. Annu. Rev Phytopathol. 1972, 10, 51–72. [Google Scholar] [CrossRef]
- Woods, A.; Coates, K.D.; Hamann, A. Is an unprecedented Dothistroma needle blight epidemic related to climate change? BioScience 2005, 55, 761–769. [Google Scholar] [CrossRef] [Green Version]
- Barnes, I.; Kirisits, T.; Akulov, A.; Chhetri, D.B.; Wingfield, B.D.; Bulgakov, T.S.; Wingfield, M.J. New host and country records of the Dothistroma needle blight pathogens from Europe and Asia. For. Pathol 2008, 38, 178–195. [Google Scholar] [CrossRef]
- Watt, M.S.; Kriticos, D.J.; Alcaraz, S.; Brown, A.V.; Leriche, A. The hosts and potential geographic range of Dothistroma needle blight. For. Ecol Manag. 2009, 257, 1505–1519. [Google Scholar] [CrossRef]
- Morelet, M. Une maladie des pins, nouvelle pour la France, due à Scirrhia pini Funk et Parker et à son stade conidien Dothistroma pini Hulbary. Bulletin Mens. Soc. Linn. Lyon 1967, 36, 361–367. [Google Scholar] [CrossRef]
- Arthaud, J. Scirrhia pini (Ascomycetes, Dothideaceae) present in the southwest [of France]. Bull. Soc. Linn. Bordx. 1972, 2, 123–124. [Google Scholar]
- Fernandez de Ana Magan, F.J. Red Band Disease of Pinus Radiata; Comunicaciones I.N.I.A. Serie Proteccion Vegetal; Ministerio De Agricultura, Instituto Nacional de Investigaciones Agrarias: Madrid, Spain, 1975; Volume 3, p. 16. Available online: http://libros.inia.es/libros/product_info.php?products_id=1113 (accessed on 22 July 2021).
- Magnani, G. Presence of Dothistroma pini on Pinus radiata. Cellul. Carta 1977, 28, 37–42. [Google Scholar]
- Fratianni, S.; Acquaotta, F. The climate of Italy. In Landscapes and Landforms of Italy; Soldati, M., Marchetti, M., Eds.; World Geomorphological Landscapes; Springer International Publishing: Basel, Switzerland, 2017; pp. 29–38. [Google Scholar] [CrossRef]
- Barnes, I.; Crous, P.W.; Wingfield, B.D.; Wingfield, M.J. Multigene phylogenies reveal that red band needle blight of Pinus is caused by two distinct species of Dothistroma, D. septosporum and D. pini. Stud. Mycol. 2004, 50, 551–565. [Google Scholar]
- Ioos, R.; Fabre, B.; Saurat, C.; Fourrier, C.; Frey, P.; Marcais, B. Development, comparison, and validation of real-time and conventional PCR tools for the detection of the fungal pathogens causing brown spot and red band needle blights of pine. Phytopathology 2010, 100, 105–114. [Google Scholar] [CrossRef] [Green Version]
- Aglietti, C.; Meinecke, C.D.; Ghelardini, L.; Barnes, I.; van der Nest, A.; Villari, C. Rapid detection of pine pathogens Lecanosticta acicola, Dothistroma pini and D. septosporum on needles by probe-based LAMP assays. Forests 2021, 12, 479. [Google Scholar] [CrossRef]
- Marchi, G.; Ghelardini, L. Outbreak of Dothistroma septosporum on Corsican Pine in Southern Italy. J. Plant Pathol. 2017, 99, S53. Available online: https://iris.unito.it/retrieve/handle/2318/1651431/368356/Masino%20A.%20et%20al.%20-%20JPP%2099%20%28supp.%20XXIII%20convegno%29.pdf (accessed on 7 July 2021).
- Ghelardini, L.; Aglietti, C.; Loria, F.; Cerboneschi, M.; Gionni, A.; Goti, E.; Maresi, G.; Moricca, S.; Marchi, G. Dothistroma needle blight in protected pine forests in Italy. Manag. Biol. Invasions 2020, 11, 689–702. [Google Scholar] [CrossRef]
- Farjon, A. Pinus nigra subsp. laricio. IUCN Red List Threat. Species 2013, e.T20453493A20453502. [Google Scholar] [CrossRef]
- Bonavita, S.; Vendramin, G.G.; Bernardini, V.; Avolio, S.; Regina, T.M.R. The first SSR-based assessment of genetic variation and structure among Pinus laricio Poiret populations within their native area. Plant Biosyst. 2016, 150, 1271–1281. [Google Scholar] [CrossRef]
- Plutino, M.; Pollastrini, M.; Avolio, S.; Bernardini, V. Tree inventory data of Pinus nigra J.F.Arnold subsp. laricio (Poir.) Maire in southern Italy. Ann. Silv. Res. 2018, 42, 43–45. [Google Scholar] [CrossRef]
- Villebonne, D.; Maugard, F. Rapid development of Dothistroma needle blight (Scirrhia pini) on Corsican pine (Pinus nigra subsp. laricio) in France. In La Sante des Forets, Annual Report 1998; Les Cahiers du Département de la santé des forêts (DSF); DERF: Paris, France, 1999; Volume 1. [Google Scholar]
- Bulman, L.S.; Gadgil, P.D.; Kershaw, D.J.; Ray, J.W. Assessment and control of Dothistroma needle-blight. Res. Bull. 2004, 229, 1176–2632. [Google Scholar]
- Sturrock, R.N.; Frankel, S.J.; Brown, A.V.; Hennon, P.E.; Kliejunas, J.T.; Lewis, K.J.; Worrall, J.J.; Woods, A.J. Climate change and forest diseases: Climate change and forest diseases. Plant Pathol. 2011, 60, 133–149. [Google Scholar] [CrossRef]
- Fraser, S.; Brown, A.V.; Woodward, S. Intraspecific variation in susceptibility to Dothistroma needle blight within native Scottish Pinus sylvestris. Plant Pathol. 2015, 64, 864–870. [Google Scholar] [CrossRef]
- Fraser, S.; Woodward, S.; Brown, A.V. Inter- and intraspecific variation in susceptibility to Dothistroma needle blight in Britain. How susceptible are Pinus sylvestris and Pinus contorta? For. Pathol. 2016, 46, 534–546. [Google Scholar] [CrossRef]
- Mullett, M.; Peace, A.; Brown, A. Persistence of Dothistroma septosporum on abscised pine needles and its implications for disease management. Plant Dis. 2016, 100, 1271–1277. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pebesma, E. Simple Features for R: Standardized Support for Spatial Vector Data. R J. 2018, 10, 439–446. [Google Scholar] [CrossRef] [Green Version]
- Wickham, H. ggplot2: Elegant Graphics for Data Analysis, 2nd ed.; Springer: New York, NY, USA, 2016; p. 260. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: https://www.R-project.org/ (accessed on 8 July 2021).
- Mullett, C.M.; Barnes, I. Dothistroma Isolation and Molecular Identification Methods. In Detection and Diagnostics of Dothistroma; COST ACTION FP1102 Determining Invasiveness and Risk of Dothistroma, Training School May 2012, Brno, Czech Republic. Available online: https://www.forestresearch.gov.uk/documents/305/DIAROD_052012_Isolation_and_indentification_97fNCCI.pdf (accessed on 7 July 2021).
- Carter-House, D.; Stajich, J.E.; Unruh, S.; Kurbessoian, T. Fungal CTAB DNA Extraction. Protocols.io 2020. [Google Scholar] [CrossRef]
- Woudenberg, J.H.C.; Aveskamp, M.M.; de Gruyter, J.; Spiers, A.G.; Crous, P.W. Multiple Didymella teleomorphs are linked to the Phoma clematidina morphotype. Persoonia 2009, 22, 56–62. [Google Scholar] [CrossRef] [Green Version]
- Okonechnikov, K.; Golosova, O.; Fursov, M.; the UGENE Team. Unipro UGENE: A unified bioinformatics toolkit. Bioinformatics 2012, 28, 1166–1167. [Google Scholar] [CrossRef] [Green Version]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-coffee: A novel method for fast and accurate multiple sequence alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]
- Brown, N.P.; Leroy, C.; Sander, C. MView: A web-compatible database search or multiple alignment viewer. Bioinformatics 1998, 14, 380–381. [Google Scholar] [CrossRef]
- Madeira, F.; Park, Y.M.; Lee, J.; Buso, N.; Gur, T.; Madhusoodanan, N.; Basutkar, P.; Tivey, A.R.N.; Potter, S.C.; Finn, R.D.; et al. The EMBL-EBI search and sequence analysis tools APIs in 2019. Nucleic Acids Res. 2019, 47, W636–W641. [Google Scholar] [CrossRef] [Green Version]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef] [PubMed]
- Katoh, K.; Standley, D.M. MAFFT Multiple Sequence Alignment Software Version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lane, D. 16S/23S rRNA Sequencing. In Nucleic Acid Techniques in Bacterial Systematics; Stackebrandt, E., Goodfellow, M., Eds.; John Wiley and Sons: New York, NY, USA, 1991; pp. 115–175. [Google Scholar]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A Rapid Bootstrap Algorithm for the RAxML Web Servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef] [PubMed]
- Ciancio, O.; Garfi, V.; Iovino, F.; Menguzzato, G. Il pino insigne nell’arboricoltura da legno: Analisi di interventi realizzati nell’Italia meridionale. Ital. For. Mont. 2006, 61, 217–231. Available online: http://ojs.aisf.it/index.php/ifm/article/view/36 (accessed on 7 July 2021). [CrossRef] [Green Version]
- Tsopelas, P.; Barnes, I.; Soulioti, N.; Wingfield, M.J. Dothistroma septosporum identified in Greece on Pinus brutia and Pinus nigra plantations. Plant Dis. 2013, 97, 1247. [Google Scholar] [CrossRef]
- Doğmuş-Lehtijärvi, H.T.; Lehtijärvi, A.; Oskay, F.; Aday Kaya, A.G.; Örtel, E.; Datumani, A. Dothistroma needle blight in Turkey. In Book of Abstract, IUFRO WP 7.02.02 Foliage Shoot and Stems Diseases: Biosecurity in Natural Forests and Plantations, Genomics and Biotechnology for Biosecurity and Forestry; Cerno Hora and Brno,: Czech Republic, 20–25 May 2013; Mendel University: Brno, Czech Republic, 2013; pp. 68–69. [Google Scholar]
- Tunalı, Z.; Doğmuș-Lehtİjärvİ, H.T.; Oskay, F. Detection of Fungal Needle Disease Agents of Turkish Pine (Pinus brutia Ten.) forests in Burdur Province Using Molecular Techniques. Süleyman Demirel Üniversitesi Fen Bilimleri Enstitüsü Dergisi; 2018; Volume 22, pp. 628–636, (In Turkish with English Abstract); Available online: https://www.cabdirect.org/cabdirect/abstract/20203105032 (accessed on 7 July 2021).
- Oskay, F.; Tunalı, Z.; Lehtijärvi, A.T.; Doğmuş-Lehtijärvi, H.T.; Woodward, S.; Mullett, M. Distribution and genetic diversity of Dothistroma septosporum in Pinus brutia forests of south-western Turkey. Plant Pathol. 2020, 69, 1551–1564. [Google Scholar] [CrossRef]
- Lazarević, J.; Davidenko, K.; Millberg, H. Incidence of Dothistroma septosporum in different pine forests in Montenegro. Mycol. Montenegrina 2014, 17, 119–131. [Google Scholar]
- Lazarević, J.; Davidenko, K.; Millberg, H. Dothistroma needle blight on high altitude pine forests in Montenegr. Balt. For. 2017, 23, 294–302. [Google Scholar]
- Adamson, K.; Mullett, M.S.; Solheim, H.; Barnes, I.; Müller, M.M.; Hantula, J.; Vuorinen, M.; Kačergius, A.; Markovskaja, S.; Musolin, D.L.; et al. Looking for relationships between the populations of Dothistroma septosporum in northern Europe and Asia. Fungal Genet. Biol. 2018, 110, 15–25. [Google Scholar] [CrossRef]
- Dale, A.L.; Lewis, K.J.; Murray, B.W. Sexual reproduction and gene flow in the pine pathogen Dothistroma septosporum in British Columbia. Phytopathology 2011, 101, 68–76. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mullett, M.S.; Brown, A.V.; Fraser, S.; Baden, R.; Tubby, K.V. Insights into the pathways of spread and potential origins of Dothistroma septosporum in Britain. Fungal Ecol. 2017, 26, 85–98. [Google Scholar] [CrossRef]
- Capron, A.; Feau, N.; Heinzelmann, R.; Barnes, I.; Benowicz, A.; Bradshaw, R.E.; Dale, A.; Lewis, K.J.; Owen, T.J.; Reich, R.; et al. Signatures of post-glacial genetic isolation and human-driven migration in the Dothistroma needle blight pathogen in Western Canada. Phytopathology 2021, 111, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Mullett, M.S.; Drenkhan, R.; Adamson, K.; Boroń, P.; Lenart-Boroń, A.; Barnes, I.; Tomšovský, M.; Jánošíková, Z.; Adamčíková, K.; Ondrušková, E.; et al. Worldwide genetic structure elucidates the Eurasian origin and invasion pathways of Dothistroma septosporum, causal agent of Dothistroma needle blight. J. Fungi 2021, 7, 111. [Google Scholar] [CrossRef] [PubMed]
- Woods, A.J.; Martín-García, J.; Bulman, L.; Vasconcelos, M.W.; Boberg, J.; La Porta, N.; Peredo, H.; Vergara, G.; Ahumada, R.; Brown, A.; et al. Dothistroma needle blight, weather and possible climatic triggers for the disease’s recent emergence. For. Pathol. 2016, 46, 443–452. [Google Scholar] [CrossRef] [Green Version]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aglietti, C.; Benigno, A.; Scali, E.; Capretti, P.; Ghelardini, L.; Moricca, S. Molecular-Based Reappraisal of a Historical Record of Dothistroma Needle Blight in the Centre of the Mediterranean Region. Forests 2021, 12, 983. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080983
Aglietti C, Benigno A, Scali E, Capretti P, Ghelardini L, Moricca S. Molecular-Based Reappraisal of a Historical Record of Dothistroma Needle Blight in the Centre of the Mediterranean Region. Forests. 2021; 12(8):983. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080983
Chicago/Turabian StyleAglietti, Chiara, Alessandra Benigno, Edoardo Scali, Paolo Capretti, Luisa Ghelardini, and Salvatore Moricca. 2021. "Molecular-Based Reappraisal of a Historical Record of Dothistroma Needle Blight in the Centre of the Mediterranean Region" Forests 12, no. 8: 983. https://0-doi-org.brum.beds.ac.uk/10.3390/f12080983