
Analysis of Olive Grove Destruction by Xylella fastidiosa Bacterium on the Land Surface Temperature in Salento Detected Using Satellite Images

 ,
,  , ,
, ,  ,
,  ,
,  and
and 
Abstract
:1. Introduction
2. Materials and Methods
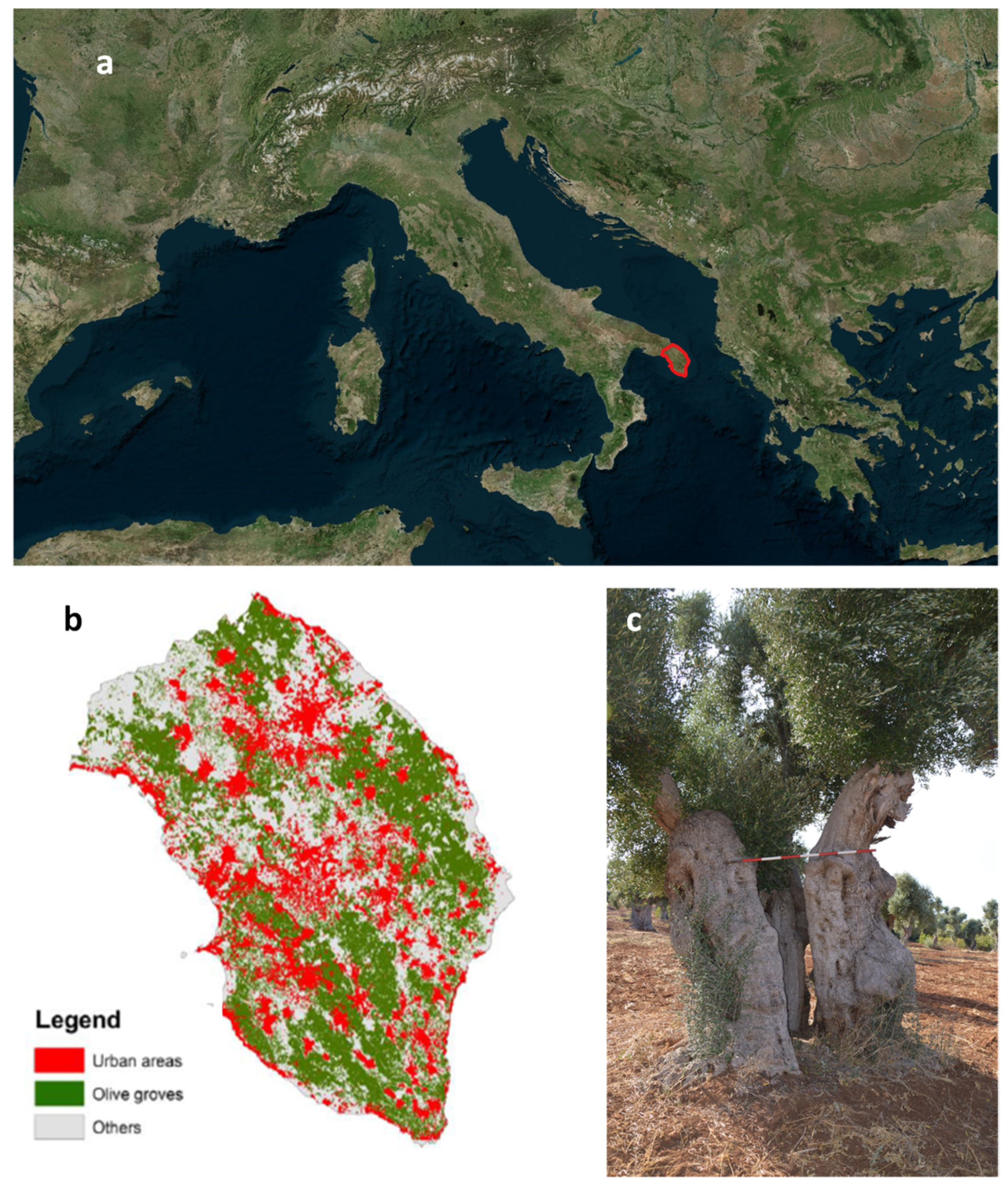
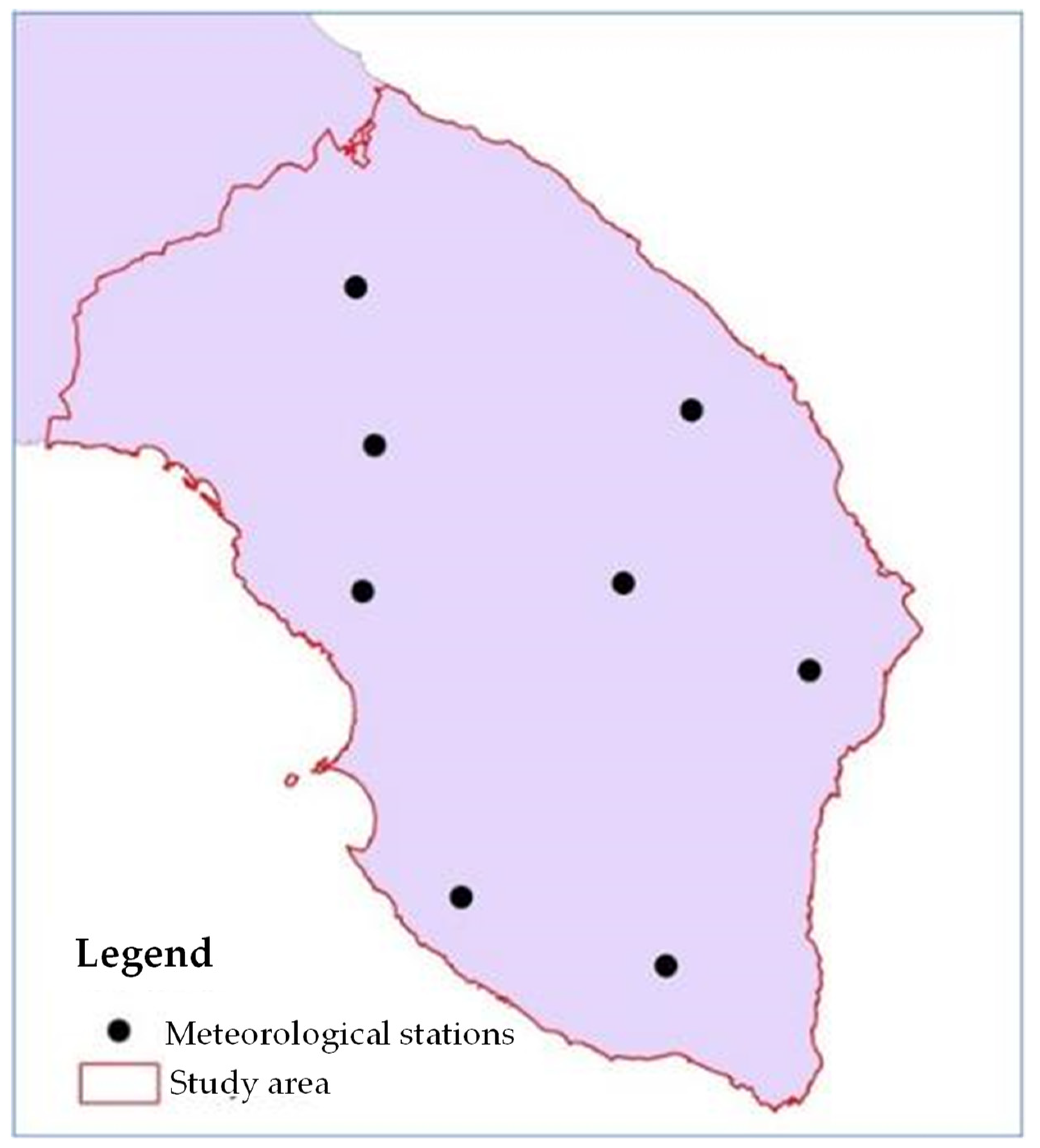
2.1. Study Area
2.2. Satellite Data
2.3. Analysis of LST Landsat 8 Data for Land-Cover Types
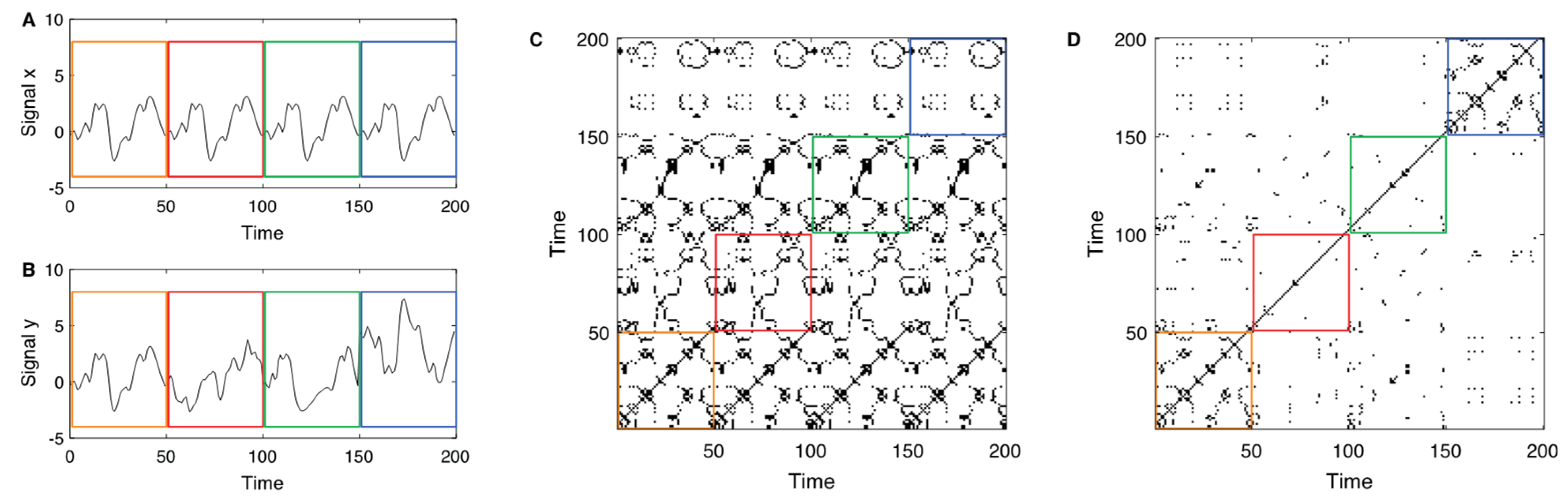
2.4. Recurrence Analysis of LST Time-Series Using MODIS Data
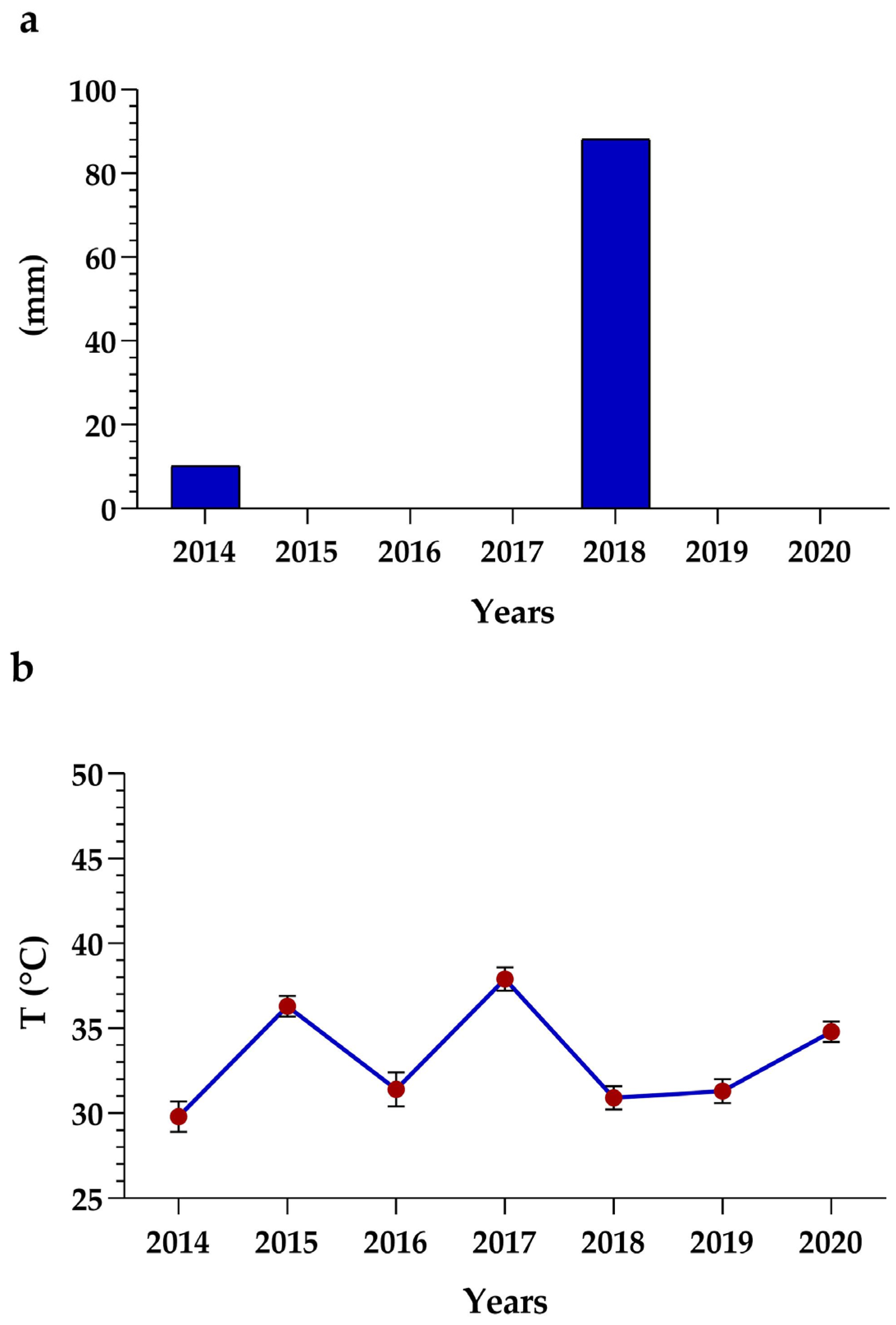
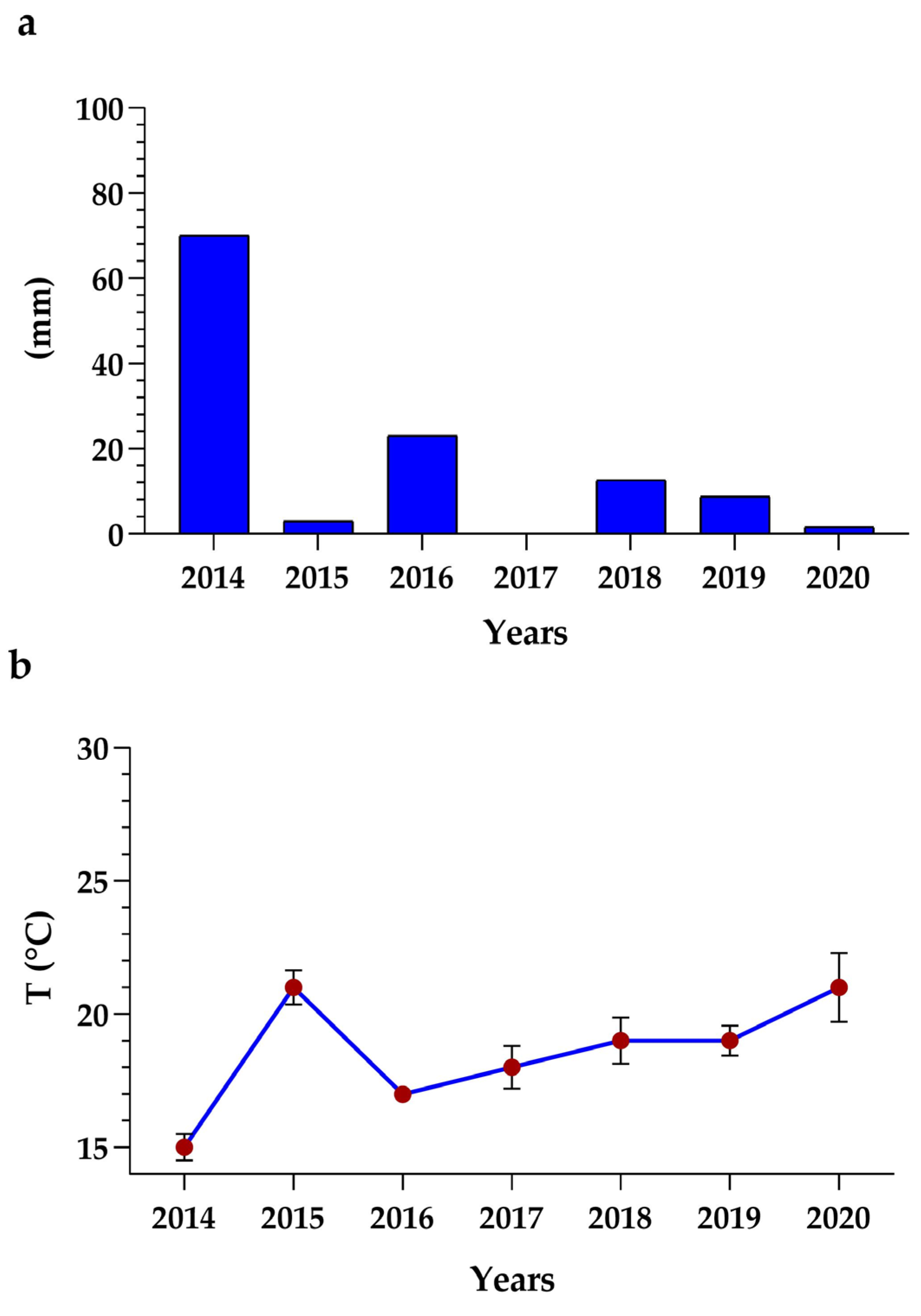
2.5. Analysis of Climate Data
3. Results
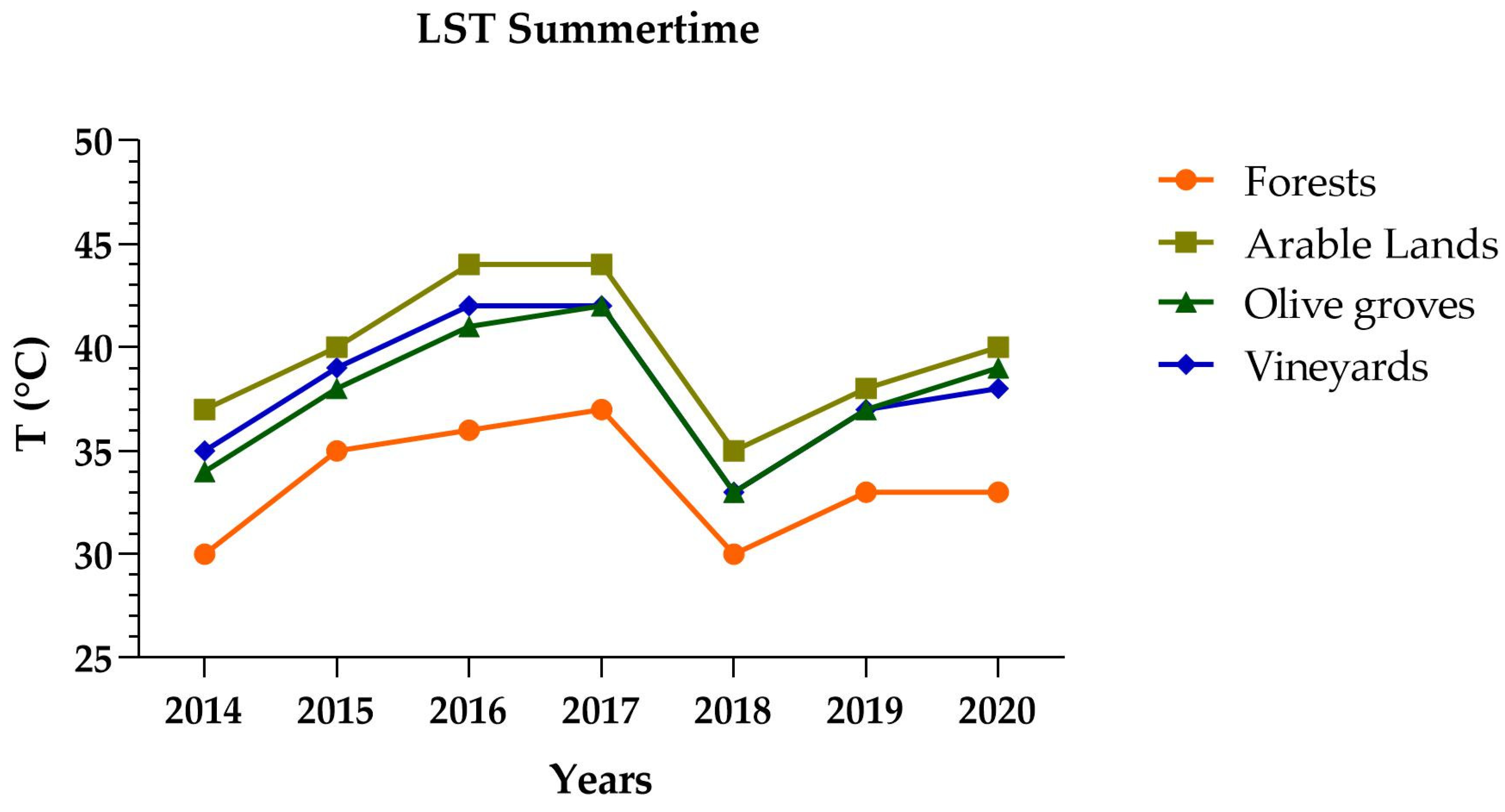
3.1. LST Landsat 8 Analysis
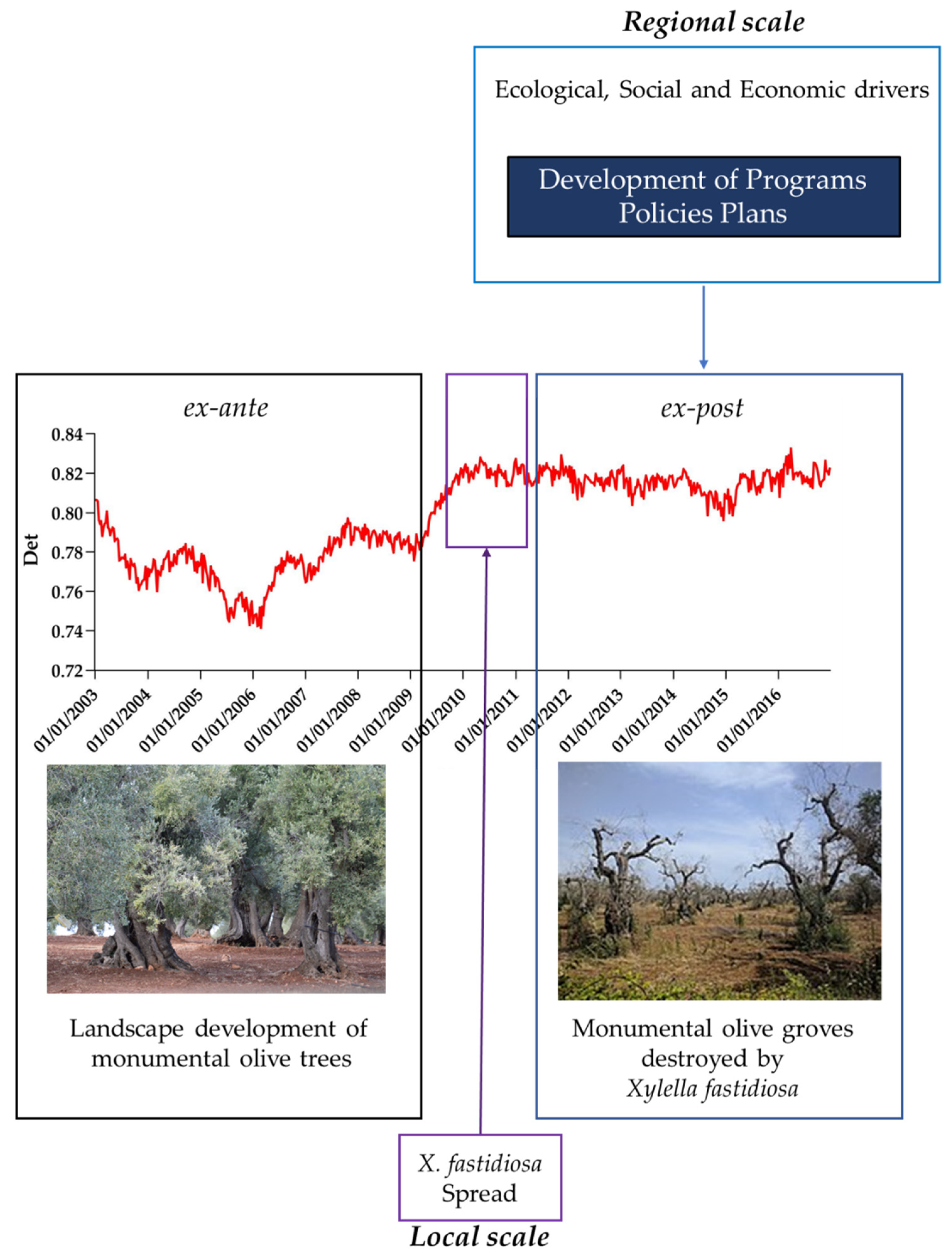
3.2. Recurrence Quantification Analysis of LST Time Series
4. Discussion
The Vision of Recurrence Analysis in Panarchy Approach
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Antrop, M. Landscape Change: Plan or Chaos? Landsc. Urban Plan. 1998, 41, 155–161. [Google Scholar] [CrossRef]
- Haines-Young, R.; Potschin, M. Valuing and assessing multifunctional landscapes: An approach based on the natural capital concept. In Multifunctional Landscapes: Theory, Values and History; Brandt, J., Vejre, H., Eds.; WIT Press: Southampton, UK, 2004; pp. 181–192. [Google Scholar]
- Lambin, E.F.; Meyfroidt, P. Land use transitions: Socio-ecological feedback versus socio-economic change. Land Use Policy 2010, 27, 108–118. [Google Scholar] [CrossRef]
- Virapongse, A.; Brooks, S.; Metcalf, E.C.; Zedalis, J.G.; Kliskey, A.; Alessa, L. A social-ecological systems approach for environmental management. J. Environ. Manag. 2016, 178, 83–91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maggiore, G.; Semeraro, T.; Aretano, R.; De Bellis, L.; Luvisi, A. GIS Analysis of Land-Use Change in Threatened Landscapes by Xylella fastidiosa. Sustainability 2019, 11, 253. [Google Scholar] [CrossRef] [Green Version]
- Turner, B.L.; Meyer, W.B. Land use and land cover in global environmental change: Considerations for study. Int. Soc. Sci. J. 1991, 130, 667–669. [Google Scholar]
- Turner, B.L.; Skole, D.; Sanderson, S.; Fischer, G.; Fresco, L.; Leemans, R. Land-Use and Land-Cover Change Science/Research Plan. In Joint Publication of the International Geosphere-Biosphere Programme (Report No. 35) and the Human Dimensions of Global Environmental Change Programme (Report No. 7); Royal Swedish Academy of Sciences: Stockholm, Sweden, 1995. [Google Scholar]
- Roger, A.; Pielke, S.R. Land Use and Climate Change. Science 2005, 310, 1625–1626. [Google Scholar]
- Righter, K.; Shearer, C.K. Magmatic fractionation of Hf and W: Constraints on the timing of core formation and differentiation in the Moon and Mars. Geochim. Cosmochim. Acta 2003, 67, 2497. [Google Scholar] [CrossRef]
- Palme, H.; Wänke, H.; Palme, H.; Wanke, H. A unified trace element model for the evolution of the lunar mantle and crust. Proc. Lunar Sci. Conf. 1975, 6, 1179–1202. [Google Scholar]
- Feddema, J.J.; Oleson, K.W.; Bonan, G.B.; Mearns, L.O.; Buja, L.E.; Meehl, G.A.; Washington, W.M. The importance of land-cover change in simulating future climates. Science 2005, 310, 1674–1678. [Google Scholar] [CrossRef] [Green Version]
- Holling, C.S.; Gunderson, L.; Ludwig, D. In Quest of a Theory of Adaptive Change. In Panarchy: Understanding Transformations in Human and Natural Systems; Gunderson, L.H., Holling, C.S., Eds.; Island Press: Washington, DC, USA, 2002; pp. 3–24. [Google Scholar]
- Gissi, E.; Burkhard, B.; Verburg, P.H. Ecosystem services: Building informed policies to orient landscape dynamics. Int. J. Biodivers. Sci. Ecosyst. Serv. Manag. 2015, 11, 185–189. [Google Scholar] [CrossRef]
- Semeraro, T.; Luvisi, A.; Lillo, A.; Aretano, R.; Buccolieri, R.; Marwan, N. Recurrence Analysis of Vegetation Indices for Highlighting the Ecosystem Response to Drought Events: An Application to the Amazon Forest. Remote Sens. 2020, 12, 907. [Google Scholar] [CrossRef] [Green Version]
- Saponari, M.; Boscia, D.; Nigro, F.; Martelli, G.P. Identification of DNA sequences related to Xylella fastidiosa in oleander, almond and olive trees exhibiting leaf scorch symptoms in Apulia (Southern Italy). J. Plant. Pathol. 2013, 95, 3. [Google Scholar]
- Martelli, G.P.; Boscia, D.; Porcelli, F.; Saponari, M. The olive quick decline syndrome in south-east Italy: A threatening phytosanitary emergency. Eur. J. Plant. Pathol. 2016, 144, 235–243. [Google Scholar] [CrossRef]
- Luvisi, A.; Aprile, A.; Sabella, E.; Vergine, M.; Nicolì, F.; Nutricati, E.; Miceli, A.; Negro, C.; De Bellis, L. Xylella fastidiosa subsp. pauca (CoDiRO strain) infection in four olive (Olea europaea L.) cultivars: Profile of phenolic compounds in leaves and progression of leaf scorch symptoms. Phytopathol. Mediterr. 2017, 56, 259–273. [Google Scholar]
- Vergine, M.; Meyer, J.B.; Cardinale, M.; Sabella, E.; Hartmann, M.; Cherubini, P.; De Bellis, L.; Luvisi, A. The Xylella fastidiosa-Resistant olive cultivar “Leccino” has stable endophytic microbiota during the Olive Quick Decline Syndrome (OQDS). Pathogens 2020, 9, 35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Almeida, R.P.P. Can Apulia’s olive trees be saved? Science 2016, 353, 346–348. [Google Scholar] [CrossRef] [Green Version]
- Semeraro, T.; Gatto, E.; Buccolieri, R.; Vergine, M.; Gao, Z.; De Bellis, L.; Luvisi, A. Changes in Olive Urban Forests Infected by Xylella fastidiosa: Impact on Microclimate and Social Health in urban areas. Int. J. Environ. Res. Public Health 2019, 16, 2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semeraro, T.; Radicchio, B.; Medagli, P.; Arzeni, S.; Turco, A.; Geneletti, D. Integration of Ecosystem Services in Strategic Environmental Assessment of a Peri-Urban Development Plan. Sustainability 2021, 13, 122. [Google Scholar] [CrossRef]
- De Groot, R.S.; Alkemade, R.; Braat, L.; Hein, L.; Willemen, L. Challenges in integrating the concept of ecosystem services and values in landscape planning, management and decision making. Ecol. Complex. 2010, 7, 260–272. [Google Scholar] [CrossRef]
- Wani, A.M.; Sahoo, G. Forest Ecosystem Services and Biodiversity. In Spatial Modeling in Forest Resources Management; Shit, P.K., Pourghasemi, H.R., Das, P., Bhunia, G.S., Eds.; Environmental Science and Engineering; Springer: Cham, Switzerland, 2021. [Google Scholar] [CrossRef]
- Lioubimtseva, E.; Cole, R.; Adams, J.M.; Kapustin, G. Impacts of climate and land-cover changes in arid lands of Central Asia. J. Arid Environ. 2005, 62, 285–308. [Google Scholar] [CrossRef]
- Samie, A.; Abbas, A.; Azeem, M.M.; Hamid, S.; Iqbal, M.A.; Hasan, S.S.; Deng, X. Examining the impacts of future land use/land cover changes on climate in Punjab province, Pakistan: Implications for environmental sustainability and economic growth. Environ. Sci. Pollut. Res. 2020, 27, 25415–25433. [Google Scholar] [CrossRef]
- National Institute of Statistics. Available online: https://www.istat.it/it/agricoltura (accessed on 25 June 2021).
- USGS-Science for a Changing World. Available online: https://www.usgs.gov/core-science-systems/nli/landsat/landsat-8?qt-science_support_page_related_con=0#qt-science_support_page_related_con (accessed on 5 April 2021).
- MODIS Images. Available online: https://lpdaac.usgs.gov/dataset_discovery/modis/modis_products_table (accessed on 12 October 2020).
- Jeevalakshmi, D.; Reddy, N.S.; Mnikiam, D. Land Surface Temperature Retrieval from LANDSAT data using Emissivity Estimation. Int. J. Appl. Eng. Res. 2017, 12, 9679–9687. [Google Scholar]
- Sekertkin, A.; Bonafin, S. Land Surface Temperature Retrieval from Landsat 5, 7, and 8 over Rural Areas: Assessment of Different Retrieval Algorithms and Emissivity Models and Toolbox Implementation. Remote Sens. 2020, 12, 294. [Google Scholar] [CrossRef] [Green Version]
- Boles, S.; Xiao, X.; Liu, J.; Zhang, Q.; Munkhutya, S.; Chen, S.; Ojima, D. Land cover characterization of Temperate East Asia: Using multi-temporal image data of vegetation sensor. Remote Sens. Environ. 2004, 90, 477–489. [Google Scholar] [CrossRef]
- SIT Puglia. Available online: http://www.sit.puglia.it/ (accessed on 6 February 2020).
- Protezione Civile of the Puglia Region. Available online: http://93.57.89.4:8081/temporeale/meteo/stazioni?codstaz=492 (accessed on 25 June 2021).
- Barsi, J.A.; Schott, J.R.; Hook, S.J.; Raqueno, N.G.; Markham, B.L.; Radocinski, R.G. Landsat-8 Thermal Infrared Sensor (TIRS) Vicarious Radiometric Calibration. Remote Sens. 2014, 6, 11607–11626. [Google Scholar] [CrossRef] [Green Version]
- Zhibin, R.; Haifeng, Z.; Xingyuan, H.; Dan, Z.; Xingyang, Y. Estimation of the Relationship between Urban Vegetation Configuration and Land Surface Temperature with Remote Sensing. J. Indian Soc. Remote Sens. 2015, 43, 89–100. [Google Scholar] [CrossRef]
- Sobrino, J.A.; Raissouni, N. Toward Remote Sensing Methods For Land Cover Dynamic Monitoring: Application to Morocco. Int. J. Remote Sens. 2010, 21, 353–366. [Google Scholar] [CrossRef]
- Stathopoulou, M.; Cartalis, C. Daytime urban heat islands from Landsat ETM+ and Corine land cover data: An application to major cities in Greece. Sol. Energy 2007, 81, 358–368. [Google Scholar] [CrossRef]
- Semeraro, T.; Pomes, A.; Del Giudice, C.; Negro, D.; Aretano, R. Planning ground based utility scale solar energy as green infrastructure to enhance ecosystem services. Energy Policy 2018, 117, 218–227. [Google Scholar] [CrossRef]
- Marwan, N.; Carmen Romano, M.; Thiel, M.; Kurths, J. Recurrence plots for the analysis of complex systems. Phys. Rep. 2007, 438, 237–329. [Google Scholar] [CrossRef]
- Marwan, N.; Kurths, J.; Foerster, S. Analysing spatially extended high-dimensional dynamics by recurrence plots. Phys. Lett. A 2015, 379, 894–900. [Google Scholar] [CrossRef] [Green Version]
- Marwan, N.; Thiel, M.; Nowaczyk, N.R. Cross Recurrence Plot Based Synchronization of Time Series. Non-Linear Process. Geophys. 2002, 9, 325–331. [Google Scholar] [CrossRef]
- Zurlini, G.; Marwan, N.; Semeraro, T.; Jones, K.B.; Areatno, R.; Pasimeni, M.R.; Valente, D.; Mulder, C.; Petrosillo, I. Investigating landscape phase transitions in Mediterranean rangelands by recurrence analysis. Landsc. Ecol. 2018, 33, 1617–1631. [Google Scholar] [CrossRef]
- Semeraro, T.; Vacchiano, G.; Aretano, R.; Ascoli, D. Application of vegetation index time series to value fire effect on primary production in a Southern European rare wetland. Ecol. Eng. 2019, 134, 9–17. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Z.Q.; Li, S.C.; Gao, J.B.; Wang, Y.L. Identifying Spatial Patterns And Dynamics Of Climate Change Using Recurrence Quantification Analysis: A Case Study Of Qinghai–Tibet Plateau. Int. J. Bifurcat. Chaos 2011, 21, 1127–1139. [Google Scholar] [CrossRef]
- Ñauñay, A.A.; Benito, R.M.; Quemada, M.; Losada, J.C.; Tarquis, A.M. Recurrence Quantification Techniques of vegetation time-series indices in semiarid grasslands. EGU Gen. Assem. 2020. [Google Scholar] [CrossRef]
- Nichols, J.M.; Trickey, S.T.; Seaver, M. Damage detection using multivariate recurrence quantification analysis. Mech. Syst. Signal. Process. 2006, 20, 421–443. [Google Scholar] [CrossRef]
- Li, S.; Zhao, Z.; Wang, Y.; Wang, Y. Identifying spatial patterns of synchronization between NDVI and climatic determinants using joint recurrence plots. Environ. Earth Sci. 2011, 64, 851–859. [Google Scholar] [CrossRef]
- Semeraro, T.; Gatto, E.; Buccolieri, R.; Catanzaro, V.; De Bellis, L.; Cotrozzi, L.; Lorenzini, G.; Vergine, M.; Luvisi, A. How Ecosystem Services Can Strengthen the Regeneration Policies for Monumental Olive Groves Destroyed by Xylella fastidiosa Bacterium in a Peri-Urban Area. Sustainability 2021, 13, 8778. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| NDVI Range | Emissivity Value (ԑ) | Land-Cover Type |
|---|---|---|
| NDVI ≤0 | 0.991 | Water |
| 0 < NDVI < 0.2 | 0.966 | Soil |
| 0.2 ≤ NDVI ≤ 0.5 | Applied Equation (2) | Mixture of soil and vegetation cover |
| NDVI > 0.5 | 0.973 | Natural vegetation (forest or wetland) |
| Years | O-A | O-F | O-V | A-F | A-V | F-V | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α = 5% | α = 5% | α = 5% | α = 5% | α = 5% | α = 5% | |||||||
| Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | |
| F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | |
| 2014 | 1.385 | 406.902 | 1.806 | 279.342 | 1.140 | 99.681 | 1.304 | 394.615 | 1.579 | 166.853 | 2.059 | 290.473 |
| 2015 | 1.119 | 418.556 | 2.604 | 273.148 | 1.520 | 143.660 | 2.328 | 412.609 | 1.700 | 143.995 | 3.958 | 304.558 |
| 2016 | 1.161 | 478.017 | 2.240 | 388.510 | 1.207 | 76.147 | 1.930 | 539.160 | 1.401 | 237.241 | 2.704 | 360.905 |
| 2017 | 1.607 | 393.575 | 3.532 | 483.039 | 1.108 | 87.044 | 2.198 | 519.295 | 1.450 | 298.324 | 3.188 | 306.241 |
| 2018 | 1.156 | 298.964 | 1.189 | 231.560 | 1.813 | 18.900 | 1.374 | 374.642 | 1.568 | 231.429 | 2.155 | 226.394 |
| 2019 | 1.152 | 142.507 | 2.382 | 333.169 | 1.446 | 99.207 | 2.067 | 354.766 | 1.666 | 182.318 | 3.443 | 230.991 |
| 2020 | 1.126 | 180.511 | 2.137 | 487.116 | 1.455 | 114.136 | 1.898 | 514.249 | 1.638 | 222.178 | 3.109 | 362.063 |
| Years | O-A | O-F | O-V | A-F | A-V | F-V | ||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| α = 5% | α = 5% | α = 5% | α = 5% | α = 5% | α = 5% | |||||||
| Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | Ftab = 1.01 | Ttab = 1.65 | |
| F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | F-Test | t-Test | |
| 2014 | 1.240 | 208.260 | 1.169 | 51.722 | 1.004 | 214.159 | 1.061 | 132.766 | 1.236 | 57.069 | 1.165 | 167.513 |
| 2015 | 1.659 | 5.193 | 1.131 | 282.100 | 1.962 | 207.320 | 1.876 | 216.525 | 1.183 | 162.384 | 2.219 | 294.208 |
| 2016 | 2.534 | 114.628 | 1.333 | 53.919 | 1.433 | 261.573 | 3.378 | 76.085 | 1.769 | 108.880 | 1.910 | 185.564 |
| 2017 | 1.066 | 99.611 | 1.070 | 98.618 | 1.026 | 150.788 | 1.141 | 143.301 | 1.040 | 80.391 | 1.097 | 175.149 |
| 2018 | 1.909 | 13.458 | 1.206 | 247.566 | 2.406 | 184.654 | 1.629 | 172.361 | 1.225 | 141.003 | 1.995 | 236.795 |
| 2019 | 1.041 | 148.201 | 1.089 | 124.289 | 1.185 | 44.067 | 1.046 | 183.474 | 1.233 | 55.436 | 1.290 | 141.247 |
| 2020 | 1.703 | 54.835 | 2.463 | 278.256 | 1.636 | 177.178 | 1.446 | 233.004 | 1.041 | 104.748 | 1.506 | 268.300 |
| F-Recurrence | Mean | Std. Dev. | F_b-a | Ftab | Alpha |
|---|---|---|---|---|---|
| LST-before | 0.207 | 0.199 | 1.08 | 1.41 | p > 0.05 |
| LST-after | 0.208 | 0.207 |
| Years | LST Difference between Farmland and Olive Groves (°C) |
|---|---|
| 2014 | 2.8 |
| 2015 | 2.2 |
| 2016 | 3.0 |
| 2017 | 2.0 |
| 2018 | 1.8 |
| 2019 | 0.8 |
| 2020 | 0.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Semeraro, T.; Buccolieri, R.; Vergine, M.; De Bellis, L.; Luvisi, A.; Emmanuel, R.; Marwan, N. Analysis of Olive Grove Destruction by Xylella fastidiosa Bacterium on the Land Surface Temperature in Salento Detected Using Satellite Images. Forests 2021, 12, 1266. https://0-doi-org.brum.beds.ac.uk/10.3390/f12091266
Semeraro T, Buccolieri R, Vergine M, De Bellis L, Luvisi A, Emmanuel R, Marwan N. Analysis of Olive Grove Destruction by Xylella fastidiosa Bacterium on the Land Surface Temperature in Salento Detected Using Satellite Images. Forests. 2021; 12(9):1266. https://0-doi-org.brum.beds.ac.uk/10.3390/f12091266
Chicago/Turabian StyleSemeraro, Teodoro, Riccardo Buccolieri, Marzia Vergine, Luigi De Bellis, Andrea Luvisi, Rohinton Emmanuel, and Norbert Marwan. 2021. "Analysis of Olive Grove Destruction by Xylella fastidiosa Bacterium on the Land Surface Temperature in Salento Detected Using Satellite Images" Forests 12, no. 9: 1266. https://0-doi-org.brum.beds.ac.uk/10.3390/f12091266