
Comprehensive Identification and Profiling of miRNAs Involved in Terpenoid Synthesis of Gleditsia sinensis Lam.


Abstract
:1. Introduction
2. Materials and Methods
2.1. Material Collection and Determination of Saponin Contents
2.2. RNA Extraction
2.3. Library Preparation and Sequencing
2.4. Sequence Filtration and sRNA Clustering
2.5. Identification of Conserved and Novel miRNAs
2.6. Differential Expression Analysis of miRNA and Functional Annotation of Target Genes
2.7. Target Prediction and Functional Annotation
3. Results
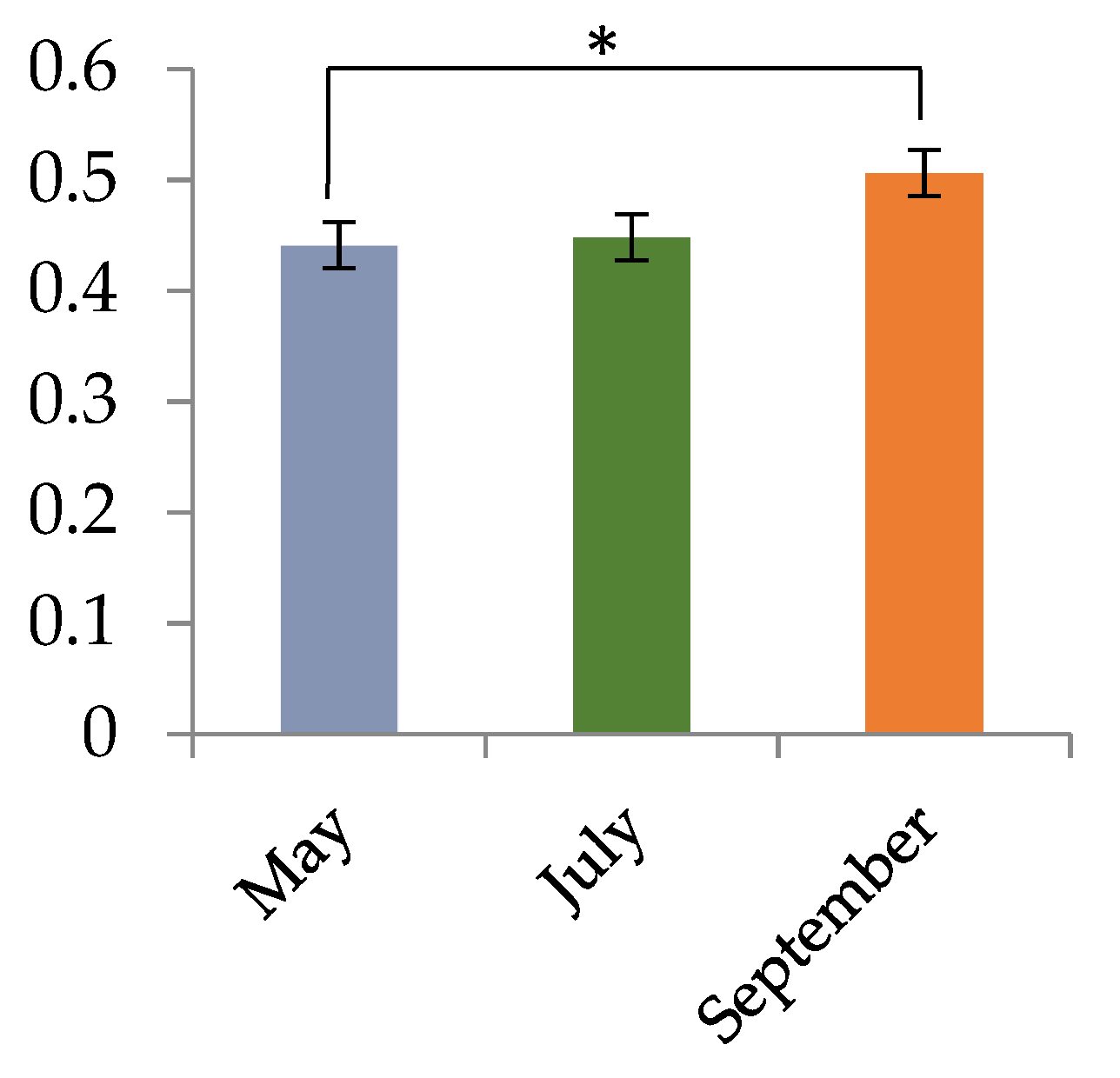
3.1. Accumulation of Saponins in G. sinensis Pods with Different Developmental Stages
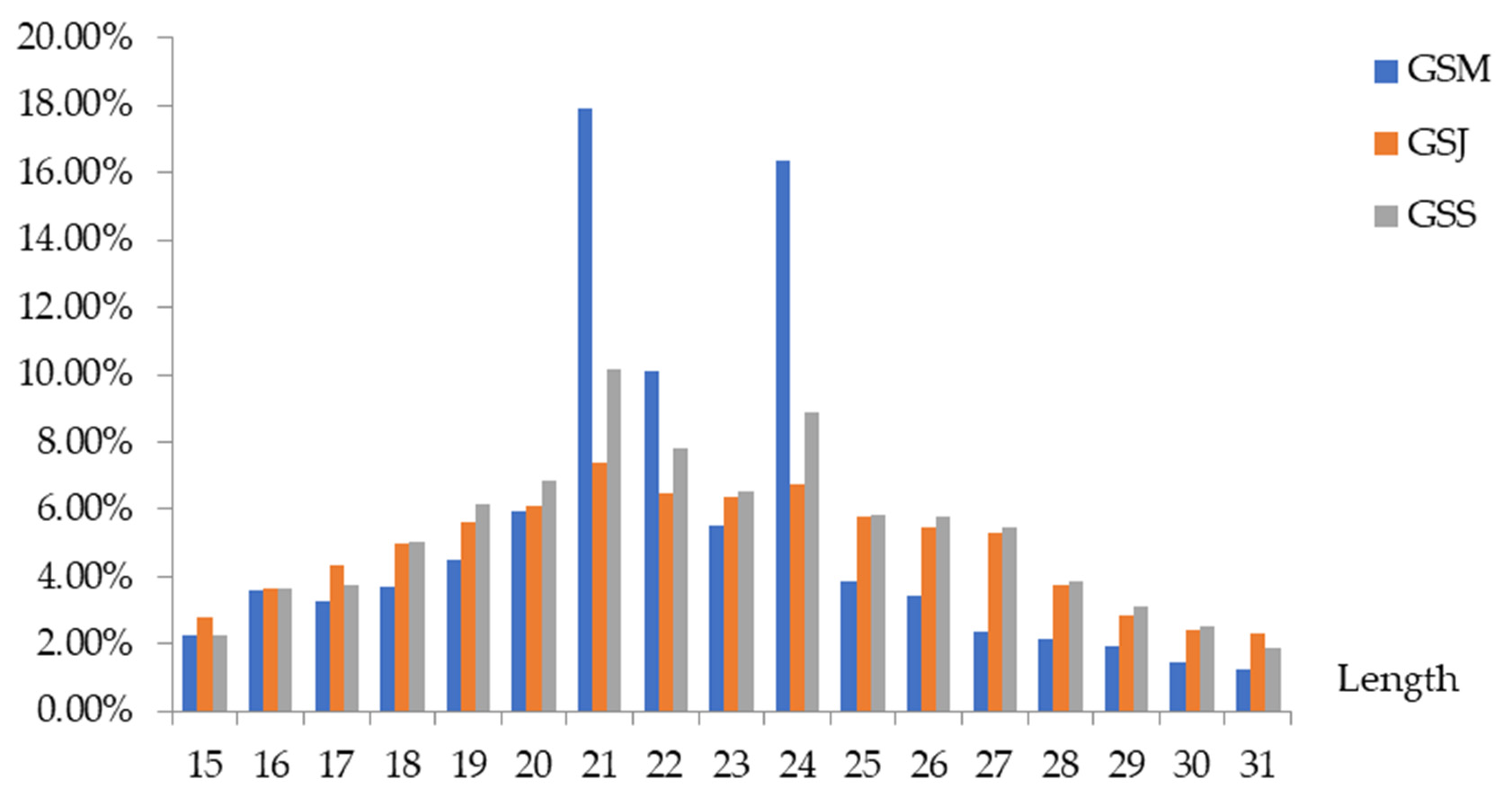
3.2. Small RNA Sequencing of G. sinensis Pods at Three Different Developmental Stages
3.3. Identification of Conserved miRNAs in G. sinensis Pods
3.4. Prediction of Novel miRNA Candidates in G. sinensis Pods
3.5. Expression Analysis of miRNAs Identified in G. sinensis Pods
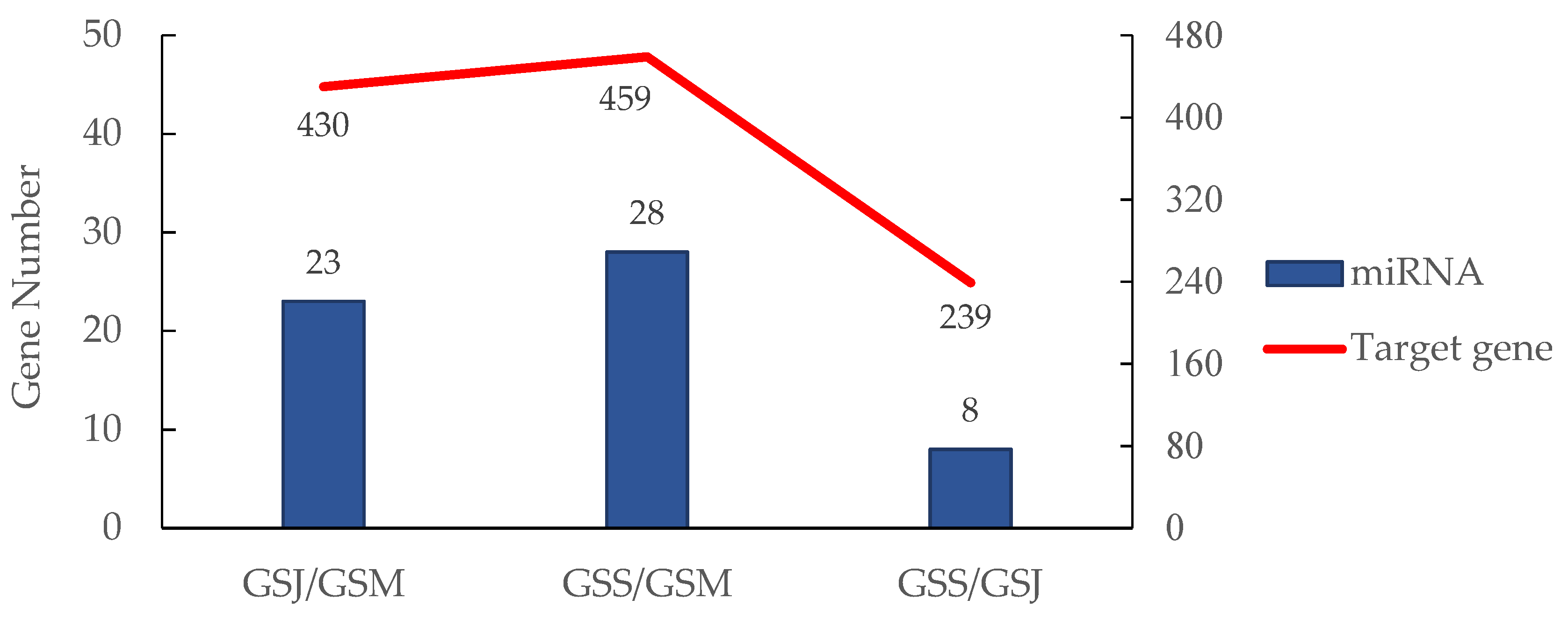
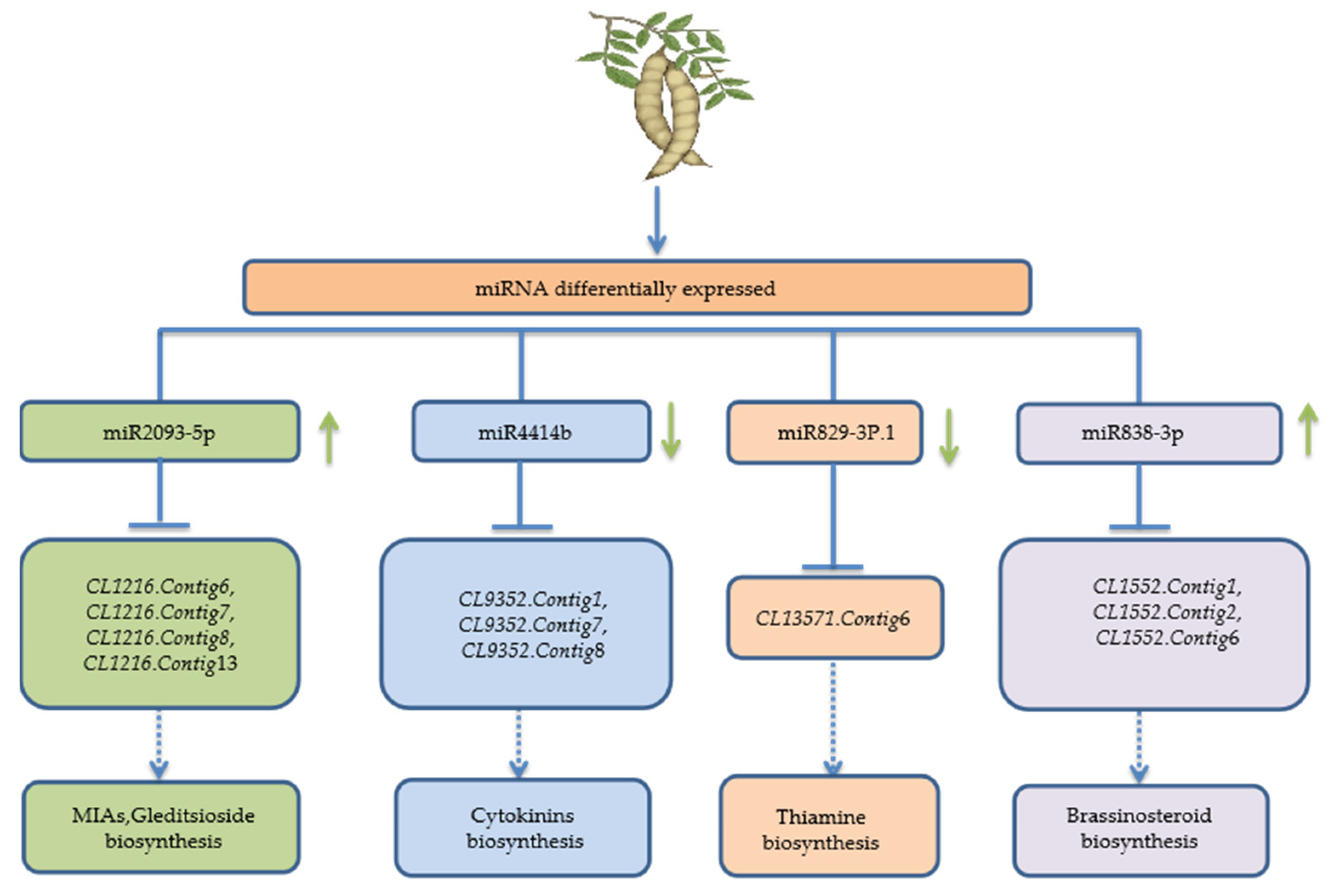
3.6. Target Prediction of miRNAs with Differential Expression
3.7. GO Analysis of Targets of Differentially Expressed miRNAs
3.8. KEGG Analysis of Targets of Differentially Expressed miRNAs
4. Discussion
4.1. Legume-Specific miRNAs in Gleditsia sinensis
4.2. miRNAs Involved in the Development of G. sinensis Pods
4.3. miRNAs Involved in Terpenoid Synthesis of Gleditsia sinensis Pods
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, J.P.; Tian, X.H.; Yang, Y.X.; Liu, Q.X.; Wang, Q.; Chen, L.P.; Li, H.L.; Zhang, W.D. Gleditsia species: An ethnomedical, phytochemical and pharmacological review. J. Ethnopharmacol. 2016, 178, 155–171. [Google Scholar]
- Liang, J.Y.; An, X.N.; Jiang, J.X.; Zhu, L.W.; Zhang, W.M. Study on the chemical constituents of the pod of G. sinensis. Chin. Wild Plant Resour. 2003, 22, 44–46. [Google Scholar]
- Lian, X.Y.; Zhang, Z. Quantitive analysis of gleditsia saponins in the fruits of G. sinensis Lam. by high performance liquid chromatography. J. Pharm. Biomed. Anal. 2013, 75, 41–46. [Google Scholar]
- Jin, S.K.; Yang, H.S.; Choi, J.S. Effect of G. sinensis Lam. Extract on physico-chemical properties of emulsion-type pork sausages. Korean J. Food Sci. Anim. Resour. 2017, 37, 274–287. [Google Scholar]
- Kim, K.H.; Han, C.W.; Yoon, S.H.; Kim, Y.S.; Kim, J.I.; Joo, M.; Choi, J.Y. The fruit hull of G. sinensis enhances the anti-tumor effect of cis-diammine dichloridoplatinum II (Cisplatin). Evid.-Based Complement. Altern. Med. 2016, 2016, 7480971. [Google Scholar]
- Kim, Y.; Koh, J.H.; Ahn, Y.J.; Oh, S.; Kim, S.H. The synergic anti-inflammatory impact of G. sinensis Lam. and Lactobacillus brevis KY21 on intestinal epithelial cells in a DSS-induced colitis model. Korean J. Food Sci. Anim. Resour. 2015, 35, 604–610. [Google Scholar] [PubMed]
- Kuwahara, Y.; Nakajima, D.; Shinpo, S.; Nakamura, M.; Kawano, N.; Kawahara, N.; Yamazaki, M.; Saito, K.; Suzuki, H.; Hirakawa, H. Identification of potential genes involved in triterpenoid saponins biosynthesis in Gleditsia sinensis by transcriptome and metabolome analyses. J. Nat. Med. 2019, 73, 369–380. [Google Scholar] [PubMed] [Green Version]
- Zhao, S.; Wang, L.; Liu, L.; Liang, Y.; Sun, Y.; Wu, J. Both the mevalonate and the non-mevalonate pathways are involved in ginsenoside biosynthesis. Plant Cell Rep. 2014, 33, 393–400. [Google Scholar] [PubMed]
- Thimmappa, R.; Geisler, K.; Louveau, T.; O’Maille, P.; Osbourn, A. Triterpene biosynthesis in plants. Annu. Rev. Plant Biol. 2014, 65, 225–257. [Google Scholar] [PubMed]
- Seki, H.; Tamura, K.; Muranaka, T. P450s and UGTs: Key players in the structural diversity of triterpenoid saponins. Plant Cell Physiol. 2015, 56, 1463–1471. [Google Scholar]
- Lange, B.M.; Ghassemian, M. Genome organization in Arabidopsis thaliana: A survey for genes involved in isoprenoid and chlorophyll metabolism. Plant Mol. Biol. 2003, 51, 925–948. [Google Scholar] [PubMed]
- Sakakibara, H. Cytokinins: Activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 2006, 57, 431–449. [Google Scholar] [PubMed] [Green Version]
- Qiu, L.; Qi, Y.; Wang, M.; Jia, X. Relationship between secondary metabolite autotoxic to plant and continuous cropping obstacles. Soils 2010, 42, 1–7. [Google Scholar]
- Fan, R.; Li, Y.; Li, C.; Zhang, Y. Differential microRNA analysis of glandular trichomes and young leaves in Xanthium strumarium L. reveals their putative roles in regulating terpenoid biosynthesis. PLoS ONE 2015, 10, e0139002. [Google Scholar]
- Gupta, O.P.; Karkute, S.G.; Banerjee, S.; Meena, N.L.; Dahuja, A. Contemporary understanding of miRNA-based regulation of secondary metabolites biosynthesis in plants. Front. Plant Sci. 2017, 29, 374. [Google Scholar]
- Soto-Suarez, M.; Baldrich, P.; Weigel, D.; Rubio-Somoza, I.; San, S.B. The Arabidopsis miR396 mediates pathogen-associated molecular pattern-triggered immune responses against fungal pathogens. Sci. Rep. 2017, 7, 44898. [Google Scholar]
- Smoczynska, A.; Szweykowska-Kulinska, Z. MicroRNA-mediated regulation of flower development in grasses. Acta Biochim. Pol. 2016, 63, 687–692. [Google Scholar]
- Wang, Y.; Zou, W.; Xiao, Y.; Cheng, L.; Liu, Y.; Gao, S.; Shi, Z.; Jiang, Y.; Qi, M.; Xu, T.; et al. MicroRNA1917 targets CTR4 splice variants to regulate ethylene responses in tomato. J. Exp. Bot. 2018, 69, 1011–1025. [Google Scholar] [PubMed] [Green Version]
- Hou, Y.; Zhai, L.; Li, X.; Xue, Y.; Wang, J.; Yang, P.; Cao, C.; Li, H.; Cui, Y.; Bian, S. Comparative analysis of fruit ripening-related miRNAs and their targets in blueberry using small RNA and degradome sequencing. Int. J. Mol. Sci. 2017, 18, 2767. [Google Scholar]
- Wang, Y.; Wang, Y.; Song, Z.; Zhang, H. Repression of MYBL2 by both microRNA858a and HY5 leads to the activation of anthocyanin biosynthetic pathway in Arabidopsis. Mol. Plant 2016, 9, 1395–1405. [Google Scholar] [PubMed] [Green Version]
- Kajal, M.; Singh, K. Small RNA profiling for identification of miRNAs involved in regulation of saponins biosynthesis in Chlorophytum borivilianum. BMC Plant Biol. 2017, 17, 265. [Google Scholar]
- Chang, S.; Puryear, J.; Cairney, J. A simple and efficient method for isolating RNA from pine trees. Plant Mol. Biol. Rep. 1993, 11, 113–116. [Google Scholar]
- Hou, J.; Sun, F.S.; Wu, Q.M.; Yang, Y.; He, W.; Wang, Y.W. An efficient method for total RNA extraction of poplar bark infected with pathogen and the application. Plant Physiol. J. 2014, 50, 223–228. [Google Scholar]
- Zhai, R.; Feng, Y.; Wang, H.; Zhan, X.; Shen, X.; Wu, W.; Zhang, Y.; Chen, D.; Dai, G.; Yang, Z.; et al. Transcriptome analysis of rice root heterosis by RNA-Seq. BMC Genom. 2013, 14, 19. [Google Scholar]
- Meyers, B.C.; Axtell, M.J.; Bartel, B.; Bartel, D.P.; Baulcombe, D.; Bowman, J.L.; Cao, X.; Carrington, J.C.; Chen, X.; Green, P.J.; et al. Criteria for annotation of plant microRNAs. Plant Cell 2008, 20, 3186–3190. [Google Scholar]
- Allen, E.; Xie, Z.; Gustafson, A.M.; Carrington, J.C. microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 2005, 121, 207–221. [Google Scholar] [PubMed] [Green Version]
- Dai, X.; Zhao, P.X. psRNATarget: A plant small RNA target analysis server. Nucleic Acids Res. 2011, 39, 155–159. [Google Scholar]
- Kanehisa, M.; Araki, M.; Goto, S.; Hattori, M.; Hirakawa, M.; Itoh, M.; Katayama, T.; Kawashima, S.; Okuda, S.; Tokimatsu, T.; et al. KEGG for linking genomes to life and the environment. Nucleic Acids Res. 2008, 36, 480–484. [Google Scholar]
- Ren, Y.Y.; Chen, L.; Zhang, Y.Y.; Kang, X.Y.; Zhang, Z.Y.; Wang, Y.W. Identification and characterization of salt-responsive microRNAs in Populus tomentosa by high-throughput sequencing. Biochimie 2015, 15, 93–105. [Google Scholar]
- Wang, X.; Zheng, Y.Q.; Su, S.C.; Ao, Y. Discovery and profiling of microRNAs at the critical period of sex differentiation in Xanthoceras sorbifolium Bunge. Forests 2019, 10, 1141. [Google Scholar]
- Jia, X.; Shen, J.; Liu, H.; Li, F.; Ding, N.; Gao, C.; Pattanaik, S.; Patra, B.; Li, R.; Yuan, L. Small tandem target mimic-mediated blockage of microRNA858 induces anthocyanin accumulation in tomato. Planta 2015, 242, 283–293. [Google Scholar]
- Gou, J.Y.; Felippes, F.F.; Liu, C.J.; Weigel, D.; Wang, J.W. Negative regulation of anthocyanin biosynthesis in Arabidopsis by a miR156-targeted SPL transcription factor. Plant Cell 2011, 23, 1512–1522. [Google Scholar] [PubMed] [Green Version]
- Li, H.; Lin, Q.; Yan, M.; Wang, M.; Wang, P.; Zhao, H.; Wang, Y.; Ni, D.; Guo, F. Relationship between Secondary Metabolism and miRNA for Important Flavor Compounds in Different Tissues of Tea Plant (Camellia sinensis) As Revealed by Genome-Wide miRNA Analysis. J. Agric. Food Chem. 2021, 69, 2001–2012. [Google Scholar]
- Zhang, X.; Rui, H.; Zhang, F.; Hu, Z.; Xia, Y.; Shen, Z. Overexpression of a functional Vicia sativa PCS1 homolog increases cadmium tolerance and phytochelatins synthesis in Arabidopsis. Front. Plant Sci. 2018, 9, 107. [Google Scholar]
- Li, Y.; Wan, L.; Bi, S.; Wan, X.; Li, Z.; Cao, J.; Tong, Z.; Xu, H.; He, F.; Li, X. Identification of drought-responsive microRNAs from roots and leaves of alfalfa by high-throughput sequencing. Genes 2017, 8, 119. [Google Scholar]
- Gururani, M.A.; Venkatesh, J.; Upadhyaya, C.P.; Nookaraju, A.; Pandey, S.K.; Park, S.W. Plant disease resistance genes: Current status and future directions. Physiol. Mol. Plant Pathol. 2012, 78, 51–65. [Google Scholar]
- Cakir, O.; Candar-Cakir, B.; Zhang, B. Small RNA and degradome sequencing reveals important microRNA function in Astragalus chrysochlorus response to selenium stimuli. Plant Biotechnol. J. 2016, 14, 543–556. [Google Scholar] [PubMed]
- Bao, D.; Ganbaatar, O.; Cui, X.; Yu, R.; Bao, W.; Falk, B.W.; Wuriyanghan, H. Down-regulation of genes coding for core RNAi components and disease resistance proteins via corresponding microRNAs might be correlated with successful Soybean mosaic virus infection in soybean. Mol. Plant Pathol. 2018, 19, 948–960. [Google Scholar]
- Ning, P.; Zhou, Y.; Gao, L.; Sun, Y.; Zhou, W.; Liu, F.; Yao, Z.; Xie, L.; Wang, J.; Gong, C. Unraveling the microRNA of Caragana korshinskii along a precipitation gradient on the Loess Plateau, China, using high-throughput sequencing. PLoS ONE 2017, 12, e172017. [Google Scholar]
- Zhang, S.; Wang, Y.; Li, K.; Zou, Y.; Chen, L.; Li, X. Identification of cold-responsive miRNAs and their target genes in nitrogen-fixing nodules of soybean. Int. J. Mol. Sci. 2014, 15, 13596–13614. [Google Scholar]
- Cui, X.; Yan, Q.; Gan, S.; Xue, D.; Dou, D.; Guo, N.; Xing, H. Overexpression of gma-miR1510a/b suppresses the expression of a NB-LRR domain gene and reduces resistance to Phytophthora sojae. Gene 2017, 621, 32–39. [Google Scholar]
- Yan, Z.; Hossain, S.; Valdés-López, O.; Hoang, N.T.; Zhai, J.; Wang, J.; Libault, M.; Brechenmacher, L.; Findley, S.; Joshi, T.; et al. Identification and functional characterization of soybean root hair microRNAs expressed in response to Bradyrhizobium japonicum infection. Plant Biotechnol. J. 2016, 14, 332–341. [Google Scholar]
- Xie, F.; Wang, Q.; Sun, R.; Zhang, B. Deep sequencing reveals important roles of microRNAs in response to drought and salinity stress in cotton. J. Exp. Bot. 2015, 66, 789–804. [Google Scholar] [PubMed] [Green Version]
- Wang, Y.; Zhang, C.; Hao, Q.; Sha, A.; Zhou, R.; Zhou, X.; Yuan, L. Elucidation of miRNAs-mediated responses to low nitrogen stress by deep sequencing of two soybean genotypes. PLoS ONE 2013, 8, e67423. [Google Scholar]
- Lin, Y.; Lai, Z. Comparative analysis reveals dynamic changes in miRNAs and their targets and expression during somatic embryogenesis in longan (Dimocarpus longan Lour.). PLoS ONE 2013, 8, e60337. [Google Scholar]
- Lu, Y.B.; Qi, Y.P.; Yang, L.T.; Guo, P.; Li, Y.; Chen, L.S. Boron-deficiency-responsive microRNAs and their targets in Citrus sinensis leaves. BMC Plant Biol. 2015, 15, 271. [Google Scholar]
- Su, Y.; Zhang, Y.; Huang, N.; Liu, F.; Su, W.; Xu, L.; Ahmad, W.; Wu, Q.; Guo, J.; Que, Y. Small RNA sequencing reveals a role for sugarcane miRNAs and their targets in response to Sporisorium scitamineum infection. BMC Genom. 2017, 18, 325. [Google Scholar]
- Cao, D.; Wang, J.; Ju, Z.; Liu, Q.; Li, S.; Tian, H.; Fu, D.; Zhu, H.; Luo, Y.; Zhu, B. Regulations on growth and development in tomato cotyledon, flower and fruit via destruction of miR396 with short tandem target mimic. Plant Sci. 2016, 247, 1–12. [Google Scholar]
- Liu, D.; Song, Y.; Chen, Z.; Yu, D. Ectopic expression of miR396 suppresses GRF target gene expression and alters leaf growth in Arabidopsis. Physiol. Plant. 2009, 136, 223–236. [Google Scholar] [PubMed]
- Baucher, M.; Moussawi, J.; Vandeputte, O.M.; Monteyne, D.; Mol, A.; Pérez-Morga, D.; El Jaziri, M. A role for the miR396/GRF network in specification of organ type during flower development, as supported by ectopic expression of Populus trichocarpa miR396c in transgenic tobacco. Plant Biol. 2013, 15, 892–898. [Google Scholar]
- Bazin, J.; Khan, G.A.; Combier, J.P.; Bustos-Sanmamed, P.; Debernardi, J.M.; Rodriguez, R.; Sorin, C.; Palatnik, J.; Hartmann, C.; Crespi, M.; et al. miR396 affects mycorrhization and root meristem activity in the legume Medicago truncatula. Plant J. 2013, 74, 920–934. [Google Scholar] [PubMed]
- Silva, G.F.F.E.; Silva, E.M.; da Silva Azevedo, M.; Guivin, M.A.C.; Ramiro, D.A.; Figueiredo, C.R.; Carrer, H.; Peres, L.E.P.; Nogueira, F.T.S. microRNA156-targeted SPL/SBP box transcription factors regulate tomato ovary and fruit development. Plant J. 2014, 78, 604–618. [Google Scholar]
- García-Mauriño, S.M.; Rivero-Rodríguez, F.; Velázquez-Cruz, A.; Hernández-Vellisca, M.; Díaz-Quintana, A.; De la Rosa, M.A.; Díaz-Moreno, I. RNA binding protein regulation and cross-talk in the control of AU-rich mRNA fate. Front. Mol. Biosci. 2017, 4, 71. [Google Scholar]
- Plass, M.; Rasmussen, S.H.; Krogh, A. Highly accessible AU-rich regions in 3’ untranslated regions are hotspots for binding of regulatory factors. PLoS Comput. Biol. 2017, 13, e1005460. [Google Scholar]
- Curinha, A.; Oliveira, B.S.; Pereira-Castro, I.; Cruz, A.; Moreira, A. Implications of polyadenylation in health and disease. Nucleus 2014, 5, 508–519. [Google Scholar]
- Li, W.; Zhang, Y.; Zhang, C.; Pei, X.; Wang, Z.; Jia, S. Presence of poly(A) and poly(A)-rich tails in a positive-strand RNA virus known to lack 3 poly(A) tails. Virology 2014, 454–455, 1–10. [Google Scholar]
- Jing, Q.; Huang, S.; Guth, S.; Zarubin, T.; Motoyama, A.; Chen, J.; Di Padova, F.; Lin, S.C.; Gram, H.; Han, J. Involvement of microRNA in AU-rich element-mediated mRNA instability. Cell 2005, 120, 623–634. [Google Scholar] [PubMed] [Green Version]
- Yang, L.; Xu, M.; Koo, Y.; He, J.; Poethig, R.S. Sugar promotes vegetative phase change in Arabidopsis thaliana by repressing the expression of MIR156A and MIR156C. eLife 2013, 2, e260. [Google Scholar]
- Lee, J.H.; Lee, J.; Kim, H.; Chae, W.B.; Kim, S.J.; Lim, Y.P.; Oh, M.H. Brassinosteroids regulate glucosinolate biosynthesis in Arabidopsis thaliana. Physiol. Plant. 2018, 163, 450–458. [Google Scholar] [PubMed]
- Liu, X.; Yang, Q.; Wang, Y.; Wang, L.; Fu, Y.; Wang, X. Brassinosteroids regulate pavement cell growth by mediating BIN2-induced microtubule stabilization. J. Exp. Bot. 2018, 69, 1037–1049. [Google Scholar] [PubMed]
- Xia, K.; Ou, X.; Tang, H.; Wang, R.; Wu, P.; Jia, Y.; Wei, X.; Xu, X.; Kang, S.H.; Kim, S.K.; et al. Rice microRNA osa-miR1848 targets the obtusifoliol 14alpha-demethylase gene OsCYP51G3 and mediates the biosynthesis of phytosterols and brassinosteroids during development and in response to stress. New Phytol. 2015, 208, 790–802. [Google Scholar]
- Kim, B.H.; Kwon, Y.; Lee, B.H.; Nam, K.H. Overexpression of miR172 suppresses the brassinosteroid signaling defects of bak1 in Arabidopsis. Biochem. Biophys. Res. Commun. 2014, 447, 479–484. [Google Scholar]
- Feng, Y.; Kong, B.; Zhang, J.; Chen, X.; Yuan, J.; Tang, X.; Ma, C. Proteomic Analysis of vernalization responsive proteins in winter wheat Jing841. Protein Pept. Lett. 2018, 25, 260–274. [Google Scholar] [PubMed]
- Kamarudin, A.N.; Lai, K.S.; Lamasudin, D.U.; Idris, A.S.; Balia, Y.Z. Enhancement of thiamine biosynthesis in oil palm seedlings by colonization of endophytic fungus hendersonia toruloidea. Front. Plant Sci. 2017, 8, 1799. [Google Scholar] [PubMed] [Green Version]
- Copley, T.R.; Aliferis, K.A.; Kliebenstein, D.J.; Jabaji, S.H. An integrated RNAseq-(1)H NMR metabolomics approach to understand soybean primary metabolism regulation in response to Rhizoctonia foliar blight disease. BMC Plant Biol. 2017, 17, 84. [Google Scholar]
- Wu, F.; Shu, J.; Jin, W. Identification and validation of miRNAs associated with the resistance of maize (Zea mays L.) to Exserohilum turcicum. PLoS ONE 2014, 9, e87251. [Google Scholar]
- Dubouzet, J.G.; Matsuda, F.; Ishihara, A.; Miyagawa, H.; Wakasa, K. Production of indole alkaloids by metabolic engineering of the tryptophan pathway in rice. Plant Biotechnol. J. 2013, 11, 1103–1111. [Google Scholar] [PubMed]







{kind=link}
{kind=link}
{kind=link}
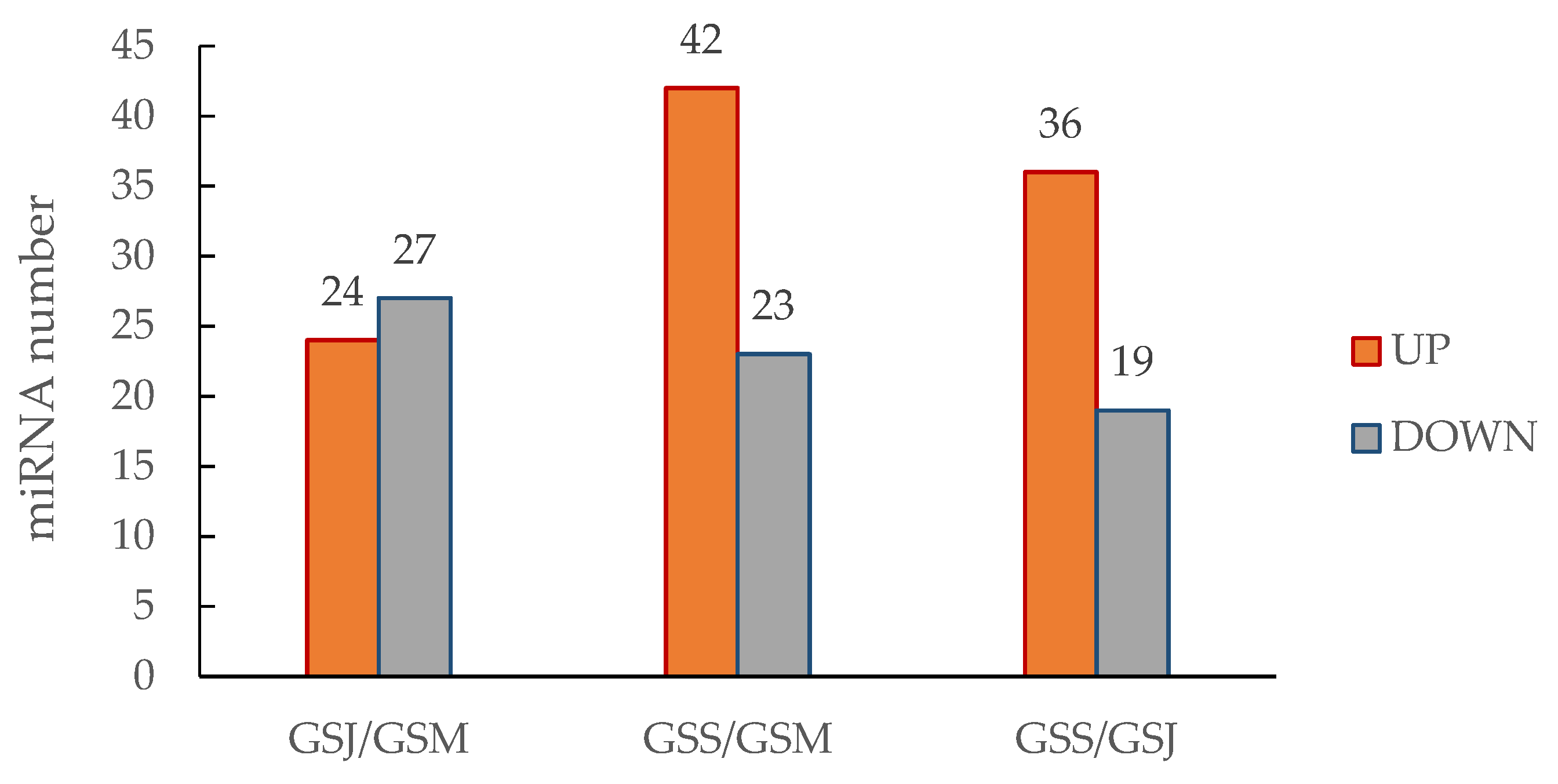{kind=link}
{kind=link}
{kind=link}
| Sample | Class | miRNA | rRNA | Repeats | snRNA | snoRNA | tRNA | Unann | |
|---|---|---|---|---|---|---|---|---|---|
| GSM | Unique | 48,994 | 451,446 | 23 | 9514 | 9217 | 18,152 | 2,380,624 | 2,917,967 |
| Total | 1,036,235 | 4,235,848 | 27 | 58,957 | 57,321 | 209,420 | 5,551,115 | 11,148,921 | |
| GSJ | Unique | 50,507 | 636,727 | 1 | 6479 | 4269 | 5706 | 658,720 | 1,362,408 |
| Total | 1,078,946 | 10,808,395 | 1 | 144,310 | 19,019 | 36,019 | 1,788,863 | 13,875,552 | |
| GSS | Unique | 49,288 | 597,554 | 14 | 5336 | 5117 | 26,131 | 1,045,473 | 1,728,910 |
| Total | 976,674 | 8,687,031 | 19 | 95,666 | 35,275 | 251,696 | 3,243,009 | 13,289,369 |
| miRNA | Read Count | Sequence (5′-3′) | ||
|---|---|---|---|---|
| GSM | GSJ | GSS | ||
| miR7984a | 82,492.5 | 268,297.5 | 208,928.5 | UCCGACUUUGUGAAAUGACUU |
| miR7696a-3p | 27,181.5 | 155,590.5 | 168,177.5 | UUCAAAUGAGAACUUUGAAG |
| miR9767 | 0 | 96,130.5 | 131,227.5 | GAUGGAAAGGACUUUGAAAAGA |
| miR2093-5p | 0 | 89,975 | 80,110.5 | GUGCUGUUACUUGGAAGAAA |
| miR5813 | 24,358.5 | 82,385.5 | 41,766 | ACAGCAGGACGGUGGUCAUGGA |
| miR2916 | 13,876.5 | 79,125 | 73,311.5 | UUGGGGGCUCGAAGACGAUCAGA |
| miR1520n | 0 | 76,947.5 | 0 | UCAACUCAGAACUGGUACGGACA |
| miR395x | 0 | 51,046.5 | 0 | GUGAAGUGUUCGGAUCGCU |
| miR7990b | 0 | 49,339 | 0 | GAAUAUUCAAAUGAGAACUUUG |
| miR5386 | 10,617 | 43,959.5 | 2505 | CGUCGGCUGUCGGCGGACUG |
| miR4342 | 0 | 38,655 | 0 | AAUGACUUGAGAGGUGUAGGAUAGGU |
| miR7532a | 12,331 | 36,495.5 | 11,995.5 | GAACAGCCUCUGGUCGAUGGA |
| miR2199 | 12,674.5 | 32,087.5 | 29,888.5 | UGAUAACUCGACGGAUCGC |
| miR5568f-3p | 9716.5 | 29,972 | 22,125.5 | GUCUGGUAAUUGGAAUGAG |
| miR1026a | 15,261 | 26,264.5 | 37,665.5 | UGUGAAAUGACUUGAGAGGUA |
| miR3444a-5p | 0 | 21,517.5 | 318 | GUUGGGAGCUCGAAGACGAUCAGA |
| miR7545 | 4872 | 20,466.5 | 12,665 | UUGAAGAAAUUAGAGUGCU |
| miR827-5p | 0 | 16,313 | 13,833.5 | UUUGUUUGAUGGUACCUACUC |
| miR8141 | 0 | 15,003.5 | 0 | UCGUCUAGUAGCUGGUU |
| miR396a-5p | 0 | 14,387 | 41,980.5 | UUCCACAGCUUUCUUGAACUG |
| miR1110 | 0 | 10,075 | 4623 | GCAGGGCGGUGGUCAUGGA |
| miR159a | 324,743 | 9399.5 | 32,021.5 | UUUGGAUUGAAGGGAGCUCUA |
| miR4994-3p | 0 | 7722.5 | 12,323.5 | UAAUUCUAGAGCUAAUACA |
| miR5671a | 2145.5 | 5020.5 | 10,828 | CAUGGUGGUGACGGGUGAC |
| miR166b | 97,140 | 2943 | 3178 | UCGGACCAGGCUUCAUUCCCC |
| miR159b-3p | 74,369 | 2309 | 14,130.5 | UUUGGAUUGAAGGGAGCUCUU |
| miR482b | 59,366.5 | 2286.5 | 6964.5 | UCUUACCUAUUCCUCCCAUGCC |
| miR1448 | 28,824 | 1770 | 3446.5 | UCUUUCCAACGCCUCCCAUACU |
| miR7767-5p | 18,941.5 | 1719.5 | 4330.5 | CCAAGAUGAGUGCUCUCCC |
| miR482a-3p | 15,428 | 916 | 1588 | UUCCCAAUGCCGCCCAUUCCGA |
| miR2118 | 13,416.5 | 878.5 | 1304 | UUGCCGAUUCCACCCAUUCCUA |
| miR472 | 27,541.5 | 827 | 8649.5 | UUUCCUACCCCUCCCAUCCC |
| miR2118a-3p | 12,621.5 | 727.5 | 1446.5 | UUGCCGAUUCCACCCAUUCCU |
| miR162a-3p | 12,305.5 | 427.5 | 4508.5 | UCGAUAAACCUCUGCAUCCAG |
| miR166m | 18,156 | 343.5 | 331.5 | UCGGACCAGGCUUCAUUCCCU |
| miR159f | 22,346.5 | 75 | 1056 | AACUGCCGACUCAUUCGUAC |
| miR396b-5p | 37,325.5 | 0 | 0 | UUCCACAGCUUUCUUGAACUU |
| miR1510a-5p | 18,193 | 0 | 0 | UUUCUUACCUAUUCCUCCCAUG |
| miR1077-5p | 16,190.5 | 0 | 0 | UUGAAGUGUUCGGAUCGCGGC |
| miR858-5p | 13,839.5 | 0 | 5 | UUCAUUGUCUGUUCGGCCGUA |
| miR2095-3p | 0 | 0 | 82,480.5 | CUUGGAUUUAUGAAAGUU |
| miR396b | 0 | 0 | 50,319 | UUCCACAGCUUUCUUGAACU |
| miR482a-5p | 0 | 0 | 12,190.5 | GGAAUGGGCUGUUUGGGAAGA |
| miR5037c | 0 | 0 | 17,883.5 | AGUGAGAACUUUGAAGGCCG |
| miR6194 | 0 | 0 | 69,261.5 | UAGUAGGGAUUGACAGACUGAG |
| miR8682 | 0 | 0 | 47,369 | AUAUCUCGGCUCUCGCAG |
| miR8752 | 0 | 0 | 20,994 | UGAUGGGGAUAGGUCAUUGCA |
| miR9736 | 0 | 0 | 49,416.5 | UGAAAGACAAACAACUGCG |
| miRNA | Species | miRNA | Species |
|---|---|---|---|
| miR9762 | Glycine max | miR4995 | Glycine max |
| miR9748 | Glycine max | miR4994-3p | Glycine max |
| miR9736 | Glycine max | miR4416a | Glycine max |
| miR862b | Glycine max | miR4415a-5p | Glycine max |
| miR7545 | Lotus japonicus | miR4415a-3p | Glycine max |
| miR7532a | Lotus japonicus | miR4412-3p | Glycine max |
| miR5780d | Glycine max | miR4387d | Glycine max |
| miR5770a | Glycine max | miR4380b | Glycine max |
| miR5741a | Medicago truncatula | miR2670a | Medicago truncatula |
| miR5678 | Glycine max | miR2658 | Medicago truncatula |
| miR5677 | Glycine max | miR2624 | Medicago truncatula |
| miR5672 | Glycine max | miR2619a | Medicago truncatula |
| miR5671a | Glycine max | miR2592bm-5p | Medicago truncatula |
| miR5559-5p | Medicago truncatula | miR2199 | Medicago truncatula |
| miR5282 | Medicago truncatula | miR1535a | Glycine max |
| miR5258 | Medicago truncatula | miR1520n | Glycine max |
| miR5209 | Medicago truncatula | miR1507a | Glycine max |
| miR5208a | Medicago truncatula | Vigna unguiculata | |
| miR5037c | Glycine max | Glycine soja | |
| Medicago truncatula | Lotus japonicus |
| miRNA | Sequence (5′-3′) | Precursor Length (nt) | MFE | Read Count | ||
|---|---|---|---|---|---|---|
| GSM | GSJ | GSS | ||||
| gsi-smR1 | CCUUCUCUUCCAUUCUUCUAG | 226 | −59.7 | 72 | 0 | 133.5 |
| gsi-smR5 | UUGGACUCUCUUCUUCUCAUG | 148 | −85.3 | 41.5 | 2.5 | 206 |
| gsi-smR6 | UCUUACCCACACCACCUAGCCC | 300 | −97.2 | 1139 | 60 | 182.5 |
| gsi-smR7 | UGCAGAACAAGUCCCAGCUUU | 211 | −68.6 | 0 | 20 | 2.5 |
| gsi-smR9 | AAGAACUCUUAUACCAAUUCG | 103 | −51.1 | 0 | 0 | 11 |
| gsi-smR10 | UGGACUCUCUUCUUCUCAUG | 143 | −82 | 0 | 0 | 63 |
| gsi-smR25 | AAGUUGAGAACAUUGAUGGC | 120 | −49.4 | 0 | 2.5 | 3 |
| gsi-smR34 | UGGUGAUCACGGGAUGAAGCU | 226 | −79.1 | 571.5 | 8.5 | 0 |
| gsi-smR35 | UGGUGAUCACGGGAUGAAGCU | 349 | −118.4 | 0 | 11.5 | 0 |
| gsi-smR54 | UUUUCGUCUUCGAGUUUCUUA | 254 | −108.6 | 1134 | 18 | 392.5 |
| gsi-smR55 | UUUUCGUCUUCGAGUUUCUUA | 235 | −99.4 | 1886 | 0 | 28.5 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Wang, J.; Wang, C.; Chen, H.; Liu, Y.; Wang, Y.; Gao, W. Comprehensive Identification and Profiling of miRNAs Involved in Terpenoid Synthesis of Gleditsia sinensis Lam. Forests 2022, 13, 108. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010108
Yang Y, Wang J, Wang C, Chen H, Liu Y, Wang Y, Gao W. Comprehensive Identification and Profiling of miRNAs Involved in Terpenoid Synthesis of Gleditsia sinensis Lam. Forests. 2022; 13(1):108. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010108
Chicago/Turabian StyleYang, Yuzhang, Jing Wang, Chun Wang, Hui Chen, Yanping Liu, Yanwei Wang, and Wei Gao. 2022. "Comprehensive Identification and Profiling of miRNAs Involved in Terpenoid Synthesis of Gleditsia sinensis Lam." Forests 13, no. 1: 108. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010108