Morphological Variation in Absorptive Roots in Downy Birch (Betula pubescens) and Norway Spruce (Picea abies) Forests Growing on Drained Peat Soils


 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Characteristics of the Stands
2.2. Collection of Root Samples and Preparation
2.3. Absorptive Root Morphology
2.4. Data Analyses
3. Results
3.1. Stand and Soil Characteristics
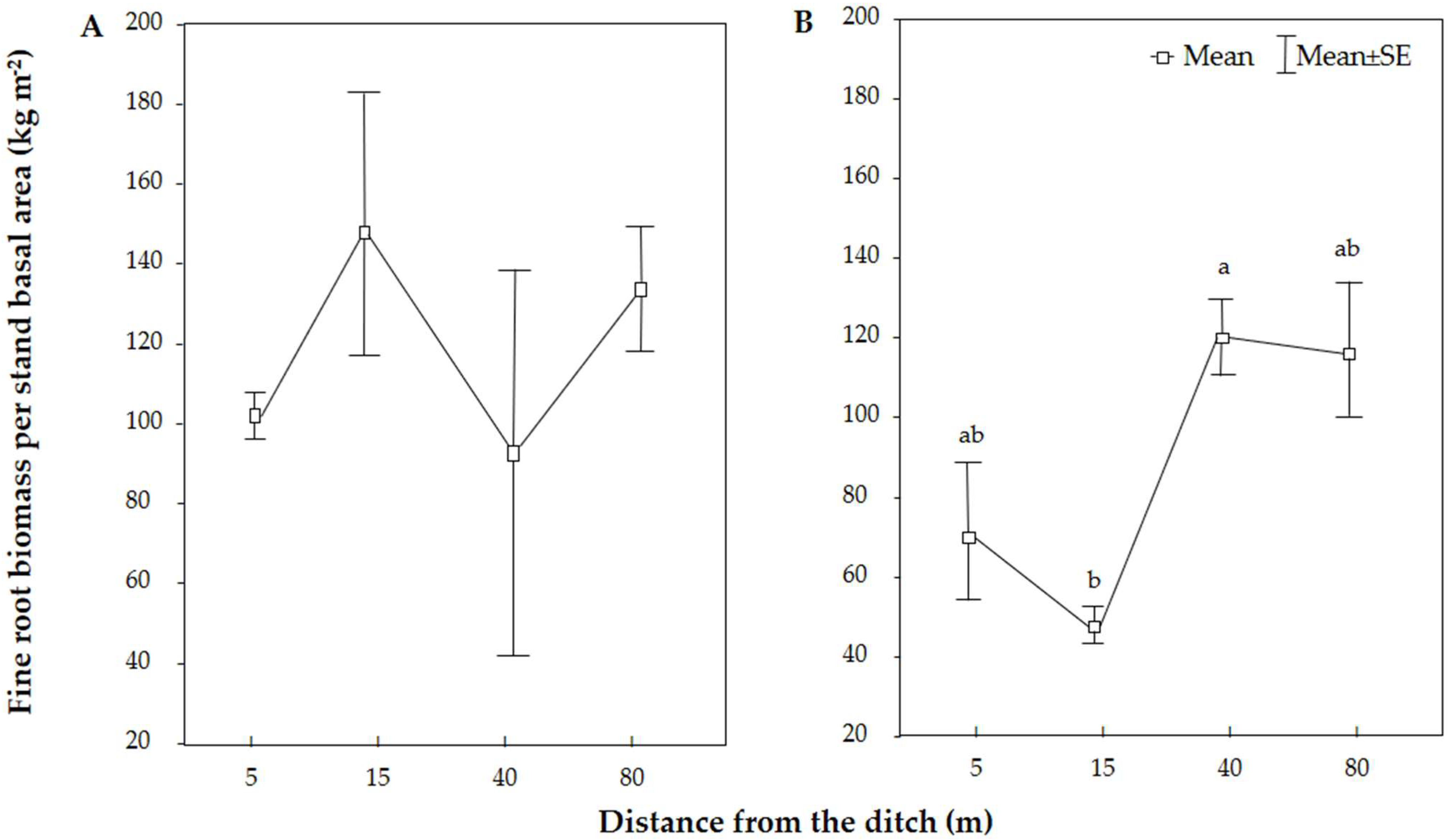
3.2. Variation in Absorptive Root Morphology across Tree Species and Distance from the Ditch
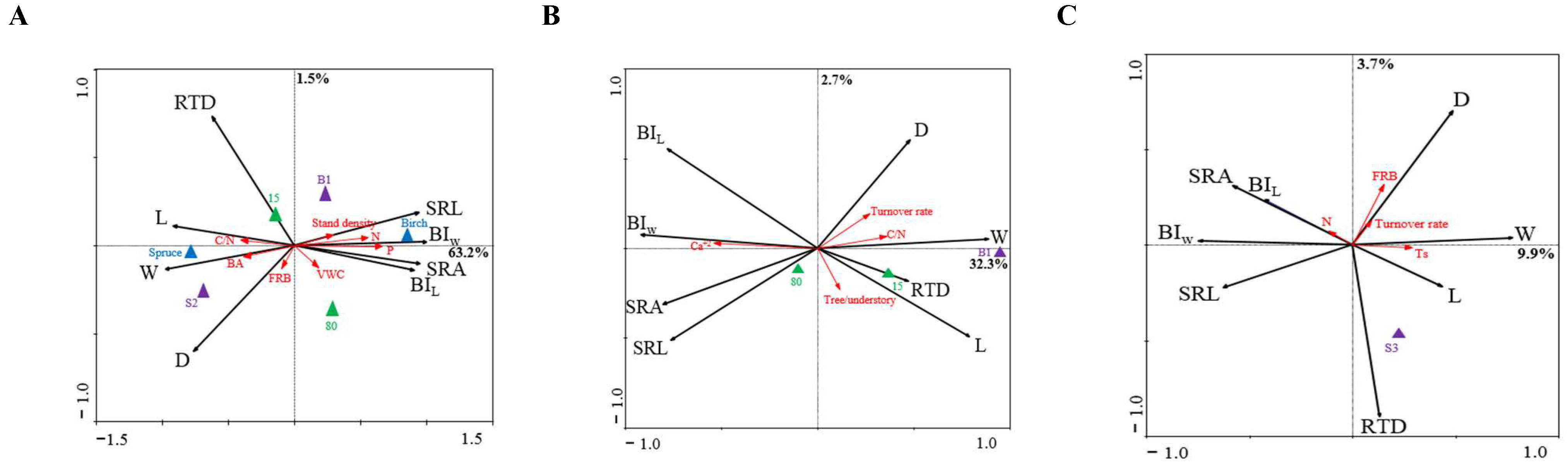
3.3. Relationships between Absorptive Root Morphology and the Characteristics of Soil and Stands
4. Discussion
4.1. Absorptive Root Morphological Variation in the Tree Species at Drained Peat Soils
4.2. Absorptive Root Morphological Variation along with the Distance from the Ditch
4.3. Absorptive Root Morphological Variation with the Stand and Soil Properties
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Sikström, U.; Hökkä, H. Interactions between soil water conditions and forest stands in boreal forests with implications for ditch network maintenance. Silva Fenn. 2016, 50, 1416. [Google Scholar] [CrossRef] [Green Version]
- Becker, H.; Aosaar, J.; Varik, M.; Morozov, G.; Aun, K.; Mander, Ü.; Soosaar, K.; Uri, V. Annual net nitrogen mineralization and litter flux in well-drained downy birch, Norway spruce and Scots pine forest ecosystems. Silva Fenn. 2018, 52, 10013. [Google Scholar] [CrossRef]
- Raudsaar, M.; Kaia-Liisa, S.; Valgepea, M. Yearbook Forest; Estonian Environment Agency: Tartu, Estonia, 2016. [Google Scholar]
- Gustavsen, H.G.; Heinonen, R.; Paavilainen, E.; Reinikainen, A. Growth and yield models for forest stands on drained peatland sites in southern Finland. For. Ecol. Manag. 1998, 107, 1–17. [Google Scholar] [CrossRef]
- Macdonald, S.E.; Yin, F. Factors influencing size inequality in peatland black spruce and tamarack: Evidence from post-drainage release growth. J. Ecol. 1999, 87, 404–412. [Google Scholar] [CrossRef]
- Miina, J. Spatial growth model for Scots pine on drained peatland. Silva Fenn. 1994, 28, 15–27. [Google Scholar] [CrossRef] [Green Version]
- Hökkä, H.; Laurén, A.; Stenberg, L.; Launiainen, S.; Leppä, K.; Nieminen, M. Defining guidelines for ditch depth in drained Scots pine dominated peatland forests. Silva Fenn. 2021, 55, 10494. [Google Scholar] [CrossRef]
- Roy, V.; Plamondon, A.P.; Bernier, P.Y. Draining forested wetland cutovers to improve seedling root zone conditions. Scand. J. For. Res. 2000, 15, 58–67. [Google Scholar] [CrossRef]
- Truu, M.; Nõlvak, H.; Ostonen, I.; Oopkaup, K.; Maddison, M.; Ligi, T.; Espenberg, M.; Uri, V.; Mander, Ü.; Truu, J. Soil Bacterial and Archaeal Communities and Their Potential to Perform N-Cycling Processes in Soils of Boreal Forests Growing on Well-Drained Peat. Front. Microbiol. 2020, 11, 591358. [Google Scholar] [CrossRef]
- Laiho, R.; Vasander, H.; Penttilä, T.; Laine, J. Dynamics of plant-mediated organic matter and nutrient cycling following water-level drawdown in boreal peatlands. Glob. Biogeochem. Cycles 2003, 17, 111–119. [Google Scholar] [CrossRef]
- Bhuiyan, R.; Minkkinen, K.; Helmisaari, H.-S.; Ojanen, P.; Penttilä, T.; Laiho, R. Estimating fine-root production by tree species and understorey functional groups in two contrasting peatland forests. Plant Soil 2017, 412, 299–316. [Google Scholar] [CrossRef] [Green Version]
- Iversen, C.M.; Childs, J.; Norby, R.J.; Ontl, T.A.; Kolka, R.K.; Brice, D.J.; McFarlane, K.J.; Hanson, P.J. Fine-root growth in a forested bog is seasonally dynamic, but shallowly distributed in nutrient-poor peat. Plant Soil 2018, 424, 123–143. [Google Scholar] [CrossRef]
- Pregitzer, K.S.; DeForest, J.L.; Burton, A.J.; Allen, M.F.; Ruess, R.W.; Hendrick, R.L. Fine root architecture of nine North American trees. Ecol. Monogr. 2002, 72, 293–309. [Google Scholar] [CrossRef]
- McCormack, M.L.; Dickie, I.A.; Eissenstat, D.M.; Fahey, T.J.; Fernandez, C.W.; Guo, D.; Helmisaari, H.S.; Hobbie, E.A.; Iversen, C.M.; Jackson, R.B. Redefining fine roots improves understanding of below-ground contributions to terrestrial biosphere processes. New Phytol. 2015, 207, 505–518. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Lasn, R. The role of soil conditions in fine root ecomorphology in Norway spruce (Picea abies (L.) Karst.). Plant Soil 1999, 208, 283–292. [Google Scholar] [CrossRef]
- Guo, D.; Xia, M.; Wei, X.; Chang, W.; Liu, Y.; Wang, Z. Anatomical traits associated with absorption and mycorrhizal colonization are linked to root branch order in twenty-three Chinese temperate tree species. New Phytol. 2008, 180, 673–683. [Google Scholar] [CrossRef]
- Zwetsloot, M.J.; Goebel, M.; Paya, A.; Grams, T.E.; Bauerle, T.L. Specific spatio-temporal dynamics of absorptive fine roots in response to neighbor species identity in a mixed beech–spruce forest. Tree Physiol. 2019, 39, 1867–1879. [Google Scholar] [CrossRef]
- Ostonen, I.; Lõhmus, K.; Alama, S.; Truu, J.; Kaar, E.; Vares, A.; Uri, V.; Kurvits, V. Morphological adaptations of fine roots in Scots pine (Pinus sylvestris L.), silver birch (Betula pendula Roth.) and black alder (Alnus glutinosa (L.) Gaertn.) stands in recultivated areas of oil shale mining and semicoke hills. Oil Shale 2006, 23, 187–202. [Google Scholar]
- Robles-Aguilar, A.A.; Pang, J.; Postma, J.A.; Schrey, S.D.; Lambers, H.; Jablonowski, N.D. The effect of pH on morphological and physiological root traits of Lupinus angustifolius treated with struvite as a recycled phosphorus source. Plant Soil 2019, 434, 65–78. [Google Scholar] [CrossRef] [Green Version]
- Doi, R.; Tanikawa, T.; Wada, R.; Hirano, Y. Morphological traits of Chamaecyparis obtusa fine roots are sensitive to soil acid buffering capacity. Plant Soil 2020, 452, 73–85. [Google Scholar] [CrossRef]
- Defrenne, C.E.; McCormack, M.L.; Roach, W.J.; Addo-Danso, S.D.; Simard, S.W. Intraspecific Fine-root trait-environment relationships across interior Douglas-fir forests of Western Canada. Plants 2019, 8, 199. [Google Scholar] [CrossRef] [Green Version]
- Olmo, M.; Lopez-Iglesias, B.; Villar, R. Drought changes the structure and elemental composition of very fine roots in seedlings of ten woody tree species. Implications for a drier climate. Plant Soil 2014, 384, 113–129. [Google Scholar] [CrossRef]
- Lozano, Y.M.; Aguilar-Trigueros, C.A.; Flaig, I.C.; Rillig, M.C. Root trait responses to drought are more heterogeneous than leaf trait responses. Funct. Ecol. 2020, 34, 2224–2235. [Google Scholar] [CrossRef]
- Wang, P.; Diao, F.; Yin, L.; Huo, C. Absorptive roots trait plasticity explains the variation of root foraging strategies in Cunninghamia lanceolata. Environ. Exp. Bot. 2016, 129, 127–135. [Google Scholar] [CrossRef]
- Leuschner, C.; Hertel, D.; Schmid, I.; Koch, O.; Muhs, A.; Hölscher, D. Stand fine root biomass and fine root morphology in old-growth beech forests as a function of precipitation and soil fertility. Plant Soil 2004, 258, 43–56. [Google Scholar] [CrossRef]
- Fitter, A.H. Functional Significance of Root Morphology and Root System Architecture. Spec. Publ. Ser. Br. Ecol. Soc. 1985, 4, 87–106. [Google Scholar]
- Lõhmus, K.; Oja, T.; Lasn, R. Specific root area: A soil characteristic. Plant Soil 1989, 119, 245–249. [Google Scholar] [CrossRef]
- Weemstra, M.; Mommer, L.; Visser, E.J.; van Ruijven, J.; Kuyper, T.W.; Mohren, G.M.; Sterck, F.J. Towards a multidimensional root trait framework: A tree root review. New Phytol. 2016, 211, 1159–1169. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ostonen, I.; Truu, M.; Helmisaari, H.S.; Lukac, M.; Borken, W.; Vanguelova, E.; Godbold, D.L.; Lõhmus, K.; Zang, U.; Tedersoo, L. Adaptive root foraging strategies along a boreal–temperate forest gradient. New Phytol. 2017, 215, 977–991. [Google Scholar] [CrossRef] [Green Version]
- Luke McCormack, M.; Adams, T.S.; Smithwick, E.A.; Eissenstat, D.M. Predicting fine root lifespan from plant functional traits in temperate trees. New Phytol. 2012, 195, 823–831. [Google Scholar] [CrossRef] [PubMed]
- Kottke, I. Mycorrhizae-rhizosphere determinants of plant communities. In Plant Roots: The Hidden Half, 3rd ed.; Marcel Dekker: New York, NY, USA, 2002; pp. 919–932. [Google Scholar]
- Isaac, M.E.; Martin, A.R.; de Melo Virginio Filho, E.; Rapidel, B.; Roupsard, O.; Van den Meersche, K. Intraspecific trait variation and coordination: Root and leaf economics spectra in coffee across environmental gradients. Front. Plant Sci. 2017, 8, 1196. [Google Scholar] [CrossRef] [Green Version]
- Liese, R.; Alings, K.; Meier, I.C. Root branching is a leading root trait of the plant economics spectrum in temperate trees. Front. Plant Sci. 2017, 8, 315. [Google Scholar] [CrossRef] [Green Version]
- Beck, P.; Caudullo, G.; de Rigo, D.; Tinner, W. Betula Pendula, Betula Pubescens and Other Birches in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 70–73. [Google Scholar]
- Seppä, H.; Alenius, T.; Bradshaw, R.H.; Giesecke, T.; Heikkilä, M.; Muukkonen, P. Invasion of Norway spruce (Picea abies) and the rise of the boreal ecosystem in Fennoscandia. J. Ecol. 2009, 97, 629–640. [Google Scholar] [CrossRef]
- Lõhmus, E. Estonian Forest Site Types; Valgus Publishers: Tallinn, Estonia, 1984. (In Estonian) [Google Scholar]
- Schad, P. The International Soil Classification System WRB. 2014 Novel Methods for Monitoring and Managing Land and Water Resources in Siberia, 3rd ed.; Springer International Publishing: New York, NY, USA, 2016; pp. 563–571. [Google Scholar]
- Clément, R.; Pärn, J.; Maddison, M.; Henine, H.; Chaumont, C.; Tournebize, J.; Uri, V.; Espenberg, M.; Günther, T.; Mander, Ü. Frequency-domain electromagnetic induction for upscaling greenhouse gas fluxes in two hemiboreal drained peatland forests. Appl. Geophys. 2020, 173, 103944. [Google Scholar] [CrossRef]
- Federation, Water Environmental, and APH Association. Standard Methods for the Examination of Water and Wastewater; American Public Health Association (APHA): Washington, DC, USA, 2005. [Google Scholar]
- Ostonen, I.; Lõhmus, K.; Helmisaari, H.-S.; Truu, J.; Meel, S. Fine root morphological adaptations in Scots pine, Norway spruce and silver birch along a latitudinal gradient in boreal forests. Tree Physiol. 2007, 27, 1627–1634. [Google Scholar] [CrossRef] [Green Version]
- Liu, B.; Li, H.; Zhu, B.; Koide, R.T.; Eissenstat, D.M.; Guo, D. Complementarity in nutrient foraging strategies of absorptive fine roots and arbuscular mycorrhizal fungi across 14 coexisting subtropical tree species. New Phytol. 2015, 208, 125–136. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.; Zhou, B.; Ge, X.; Cao, Y.; Brunner, I.; Shi, J.; Li, M.-H. Species-Specific Responses of Root Morphology of Three Co existing Tree Species to Nutrient Patches Reflect Their Root Foraging Strategies. Front. Plant Sci. 2021, 11, 2322. [Google Scholar] [CrossRef] [PubMed]
- Bauhus, J.; Messier, C. Soil exploitation strategies of fine roots in different tree species of the southern boreal forest of eastern Canada. Can. J. For. Res. 1999, 29, 260–273. [Google Scholar] [CrossRef]
- Werger, L.; Bergmann, J.; Weber, E.; Heinze, J. Wind intensity affects fine root morphological traits with consequences for plant-soil feedback effects. AoB Plants 2020, 12, plaa050. [Google Scholar] [CrossRef]
- Zhou, M.; Wang, J.; Bai, W.; Zhang, Y.; Zhang, W.H. The response of root traits to precipitation change of herbaceous species in temperate steppes. Funct. Ecol. 2019, 33, 2030–2041. [Google Scholar] [CrossRef]
- McCormack, M.L.; Guo, D. Impacts of environmental factors on fine root lifespan. Front. Plant Sci. 2014, 5, 205. [Google Scholar] [CrossRef] [Green Version]
- Young, K.L.; Woo, M.-k.; Edlund, S.A. Influence of local topography, soils, and vegetation on microclimate and hydrology at a high Arctic site, Ellesmere Island, Canada. Arct. Antarct. Alp. Res. 1997, 29, 270–284. [Google Scholar] [CrossRef]
- Waddington, J.; Morris, P.; Kettridge, N.; Granath, G.; Thompson, D.; Moore, P. Hydrological feedbacks in northern peatlands. Ecohydrology 2015, 8, 113–127. [Google Scholar] [CrossRef]
- Leppä, K.; Korkiakoski, M.; Nieminen, M.; Laiho, R.; Hotanen, J.-P.; Kieloaho, A.-J.; Korpela, L.; Laurila, T.; Lohila, A.; Minkkinen, K. Vegetation controls of water and energy balance of a drained peatland forest: Responses to alternative harvesting practices. Agric. For. Meteorol. 2020, 295, 108198. [Google Scholar] [CrossRef]
- Bardgett, R.D. The Biology of Soil: A Community and Ecosystem Approach; Oxford University Press: Oxford, UK, 2005; p. 242. [Google Scholar]
- Fang, Y.; Wang, F.; Chen, J.; Lin, C.; Lin, X. Microbial responses in a cold waterlogged paddy field to different distances from a drainage ditch. Chil. J. Agric. Res. 2017, 77, 87–94. [Google Scholar] [CrossRef] [Green Version]
- Qiu, S.; Wang, M.; Wang, F.; Chen, J.; Li, X.; Li, Q.; Lin, C.; Lin, X. Effects of open drainage ditch design on bacterial and fungal communities of cold waterlogged paddy soils. Braz. J. Microbiol. 2013, 44, 983–991. [Google Scholar] [CrossRef] [Green Version]
- Freschet, G.T.; Valverde-Barrantes, O.J.; Tucker, C.M.; Craine, J.M.; McCormack, M.L.; Violle, C.; Fort, F.; Blackwood, C.B.; Urban-Mead, K.R.; Iversen, C.M. Climate, soil and plant functional types as drivers of global fine-root trait variation. J. Ecol. 2017, 105, 1182–1196. [Google Scholar] [CrossRef] [Green Version]
- Marañón, T.; Navarro-Fernández, C.M.; Gil Martínez, M.; Domínguez, M.T.; Madejón, P.; Villar Montero, R. Variation in morphological and chemical traits of Mediterranean tree roots: Linkage with leaf traits and soil conditions. Plant Soil 2020, 449, 389–403. [Google Scholar] [CrossRef]
- Parts, K.; Tedersoo, L.; Schindlbacher, A.; Sigurdsson, B.D.; Leblans, N.I.; Oddsdóttir, E.S.; Borken, W.; Ostonen, I. Acclimation of fine root systems to soil warming: Comparison of an experimental setup and a natural soil temperature gradient. Ecosystems 2019, 22, 457–472. [Google Scholar] [CrossRef] [Green Version]
- Onwuka, B.; Mang, B. Effects of soil temperature on some soil properties and plant growth. Adv. Plants Agric. Res. 2018, 8, 34. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Replicate Stands | Geographical Coordinates | Fine Root Biomass (g m−2) | Peat Depth (cm) | pH-H2O | Soil Temperature-10 cm (°C) | Soil Water Content-10 cm (m3 m−3) |
|---|---|---|---|---|---|---|
| B1 | N 58°18′24.8 E 27°15′23.1 | 222 ± 33 | 43 ± 2 | 4.38 ± 0.05 | 11.76 ± 0.04 | 0.21 ± 0.02 |
| B2 | N 58°17′21.4 E 27°19′3.2 | 206 ± 50 | 87 ± 1 | 5.68 ± 0.05 | 12.00 ± 0.21 | 0.23 ± 0.03 |
| B3 | N 58°18′37.0 E 27°21′11.8 | 251 ± 25 | 77 ± 3 | 4.85 ± 0.03 | 11.86 ± 0.08 | 0.30 ± 0.06 |
| Means for birch | 226 ± 20 | 69 ± 6 | 4.97 ± 0.16 | 11.87 ± 0.07 | 0.25 ± 0.03 | |
| S1 | N 58°18′6.3 E 27°16′54.0 | 242 ± 40 | 49 ± 2 | 5.08 ± 0.13 | 11.68 ± 0.18 | 0.32 ± 0.03 |
| S2 | N 58°17′49.3 E 27°14′53.4 | 289 ± 71 | 36 ± 5 | 4.93 ± 0.30 | 10.76 ± 0.10 | 0.24 ± 0.03 |
| S3 | N 58°15′14.5 E 27°17′44.2 | 140 ± 42 | 65 ± 6 | 4.04 ± 0.05 | 11.23 ± 0.07 | 0.19 ± 0.02 |
| Means for spruce | 217 ± 33 | 51 ± 4 | 4.63 ± 0.16 | 11.23 ± 0.12 | 0.25 ± 0.02 | |
| Stand Characteristics | Species | Replicate Stands | Stand Values (Per Stand) | Distances from the Ditch (m) | Stand Values (Per Sampling Plot) |
|---|---|---|---|---|---|
| Stand density (trees ha−1) | Birch | B1 | 2000 ± 294 | 5 | 1400 ± 0 |
| B2 | 2100 ± 268 | 15 | 1633 ± 260 | ||
| B3 | 1325 ± 229 | 40 | 1867 ± 546 | ||
| 80 | 2333 ± 233 | ||||
| Spruce | S1 | 1075 ± 138 b | 5 | 1200 ± 178 | |
| S2 | 1900 ± 183 a | 15 | 1600 ± 289 | ||
| S3 | 1340 ± 169 ab | 40 | 1433 ± 376 | ||
| 80 | 1550 ± 150 | ||||
| BA (m2 ha−1) | Birch | B1 | 25 ± 4 | 5 | 24 ± 1 |
| B2 | 20 ± 3 | 15 | 23 ± 5 | ||
| B3 | 18 ± 3 | 40 | 21 ± 6 | ||
| 80 | 16 ± 1 | ||||
| Spruce | S1 | 32 ± 6 | 5 | 30 ± 7 | |
| S2 | 28 ± 4 | 15 | 29 ± 3 | ||
| S3 | 22 ± 1 | 40 | 20 ± 2 | ||
| 80 | 25 ± 4 | ||||
| DBH (cm) | Birch | B1 | 15 ± 2 | 5 | 15 ± 0 a |
| B2 | 14 ± 1 | 15 | 16 ± 2 a | ||
| B3 | 14 ± 1 | 40 | 16 ± 1 a | ||
| 80 | 11 ± 0 b | ||||
| Spruce | S1 | 20 ± 1 | 5 | 19 ± 1 | |
| S2 | 17 ± 1 | 15 | 17 ± 2 | ||
| S3 | 16 ± 2 | 40 | 16 ± 2 | ||
| 80 | 17 ± 0 |
| Distances from the Ditch (m) | ||||||
|---|---|---|---|---|---|---|
| Root Traits | Stands | 5 | 15 | 40 | 80 | Means of Four Sampling Plots |
| D (mm) | Birch | 0.227 ± 0.013 | 0.231 ± 0.012 | 0.221 ± 0.003 | 0.229 ± 0.011 | 0.227 ± 0.002 B |
| Spruce | 0.278 ± 0.007 ab | 0.265 ± 0.004 c | 0.272 ± 0.005 abc | 0.281 ± 0.009 ab | 0.274 ± 0.003 A | |
| L (mm) | Birch | 1.12 ± 0.14 abc | 1.27 ± 0.09 a | 1.01 ± 0.19 c | 1.16 ± 0.13 abc | 1.14 ± 0.05 B |
| Spruce | 1.90 ± 0.06 | 1.82 ± 0.07 | 1.86 ± 0.07 | 1.78 ± 0.02 | 1.84 ± 0.02 A | |
| W (mg) | Birch | 0.0124 ± 0.0035 abc | 0.0141 ± 0.0026 a | 0.0105 ± 0.0025 c | 0.0123 ± 0.0027 abc | 0.0124 ± 0.0007 B |
| Spruce | 0.0339 ± 0.0021 a | 0.0301 ± 0.0012 c | 0.0324 ± 0.0020 abc | 0.0305 ± 0.0016 abc | 0.0317 ± 0.0009 A | |
| BIL (cm−1) | Birch | 9.58 ± 1.10 bc | 8.45 ± 0.67 bc | 11.26 ± 1.90 a | 9.40 ± 1.15 bc | 9.67 ± 0.58 A |
| Spruce | 5.44 ± 0.12 | 5.60 ± 0.23 | 5.53 ± 0.23 | 5.73 ± 0.03 | 5.58 ± 0.06 B | |
| BIw (mg−1) | Birch | 99.0 ± 22.9 abc | 82.2 ± 15.4 c | 121.4 ± 24.6 ab | 106.3 ± 26.8 ab | 102.2 ± 8.1 A |
| Spruce | 31.1 ± 1.6 c | 34.3 ± 1.4 a | 32.5 ± 2.3 abc | 34.6 ± 1.6 abc | 33.1 ± 0.8 B | |
| SRA (m2 kg−1) | Birch | 69 ± 6 abc | 68 ± 4 c | 71 ± 4 abc | 75 ± 6 a | 71 ± 2 A |
| Spruce | 50 ± 1 bc | 51 ± 1 abc | 50 ± 1 bc | 53 ± 2 a | 51 ± 1 B | |
| SRL (m g−1) | Birch | 100 ± 14 abc | 95 ± 11 c | 105 ± 4 abc | 108 ± 14 a | 103 ± 3 A |
| Spruce | 57 ± 2 c | 62 ± 1 a | 59 ± 2 abc | 60 ± 3 abc | 59 ± 1 B | |
| RTD (kg m−3) | Birch | 262 ± 11 ab | 262 ± 4 ab | 264 ± 17 ab | 242 ± 10 c | 258 ± 5 B |
| Spruce | 295 ± 6 ab | 300 ± 7 ab | 300 ± 4 ab | 276 ± 1 c | 293 ± 6 A | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rezapour, A.; Truu, M.; Maddison, M.; Rohula-Okunev, G.; Tullus, A.; Uri, V.; Mander, Ü.; Ostonen, I. Morphological Variation in Absorptive Roots in Downy Birch (Betula pubescens) and Norway Spruce (Picea abies) Forests Growing on Drained Peat Soils. Forests 2022, 13, 112. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010112
Rezapour A, Truu M, Maddison M, Rohula-Okunev G, Tullus A, Uri V, Mander Ü, Ostonen I. Morphological Variation in Absorptive Roots in Downy Birch (Betula pubescens) and Norway Spruce (Picea abies) Forests Growing on Drained Peat Soils. Forests. 2022; 13(1):112. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010112
Chicago/Turabian StyleRezapour, Azadeh, Marika Truu, Martin Maddison, Gristin Rohula-Okunev, Arvo Tullus, Veiko Uri, Ülo Mander, and Ivika Ostonen. 2022. "Morphological Variation in Absorptive Roots in Downy Birch (Betula pubescens) and Norway Spruce (Picea abies) Forests Growing on Drained Peat Soils" Forests 13, no. 1: 112. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010112