
Heterotrophic Bacteria Play an Important Role in Endemism of Cephalostachyum pingbianense (Hsueh & Y.M. Yang ex Yi et al.) D.Z. Li & H.Q. Yang, 2007, a Full-Year Shooting Woody Bamboo


Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Selection and Soil Sampling
2.2. Soil Chemical Characterization and Enzyme Activity Measurements
2.3. DNA Extraction, PCR Amplification, and MiSeq Sequencing
2.4. Bioinformatics and Statistical Analysis
3. Results
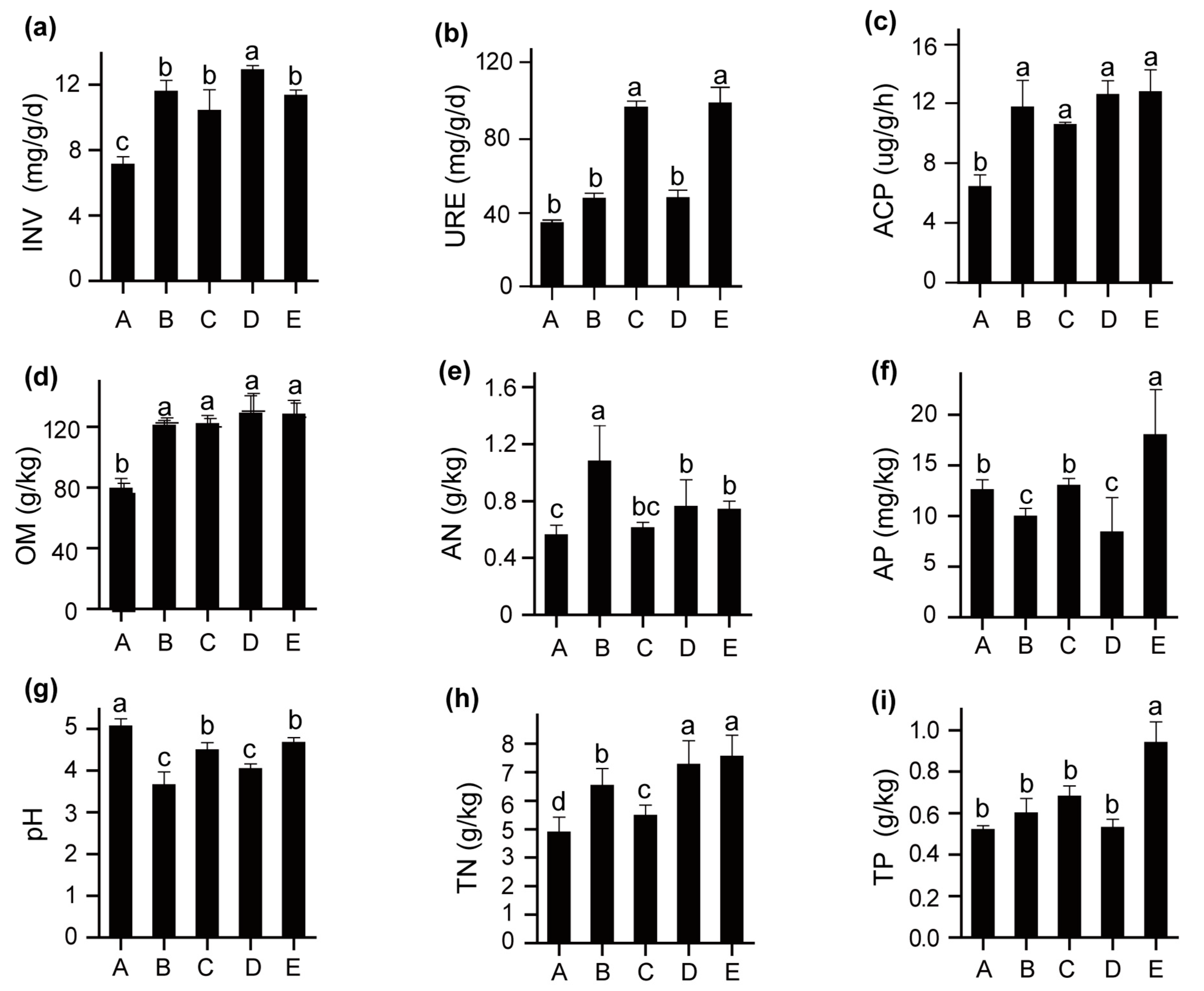
3.1. Soil Chemical Properties and Soil Enzymatic Activities
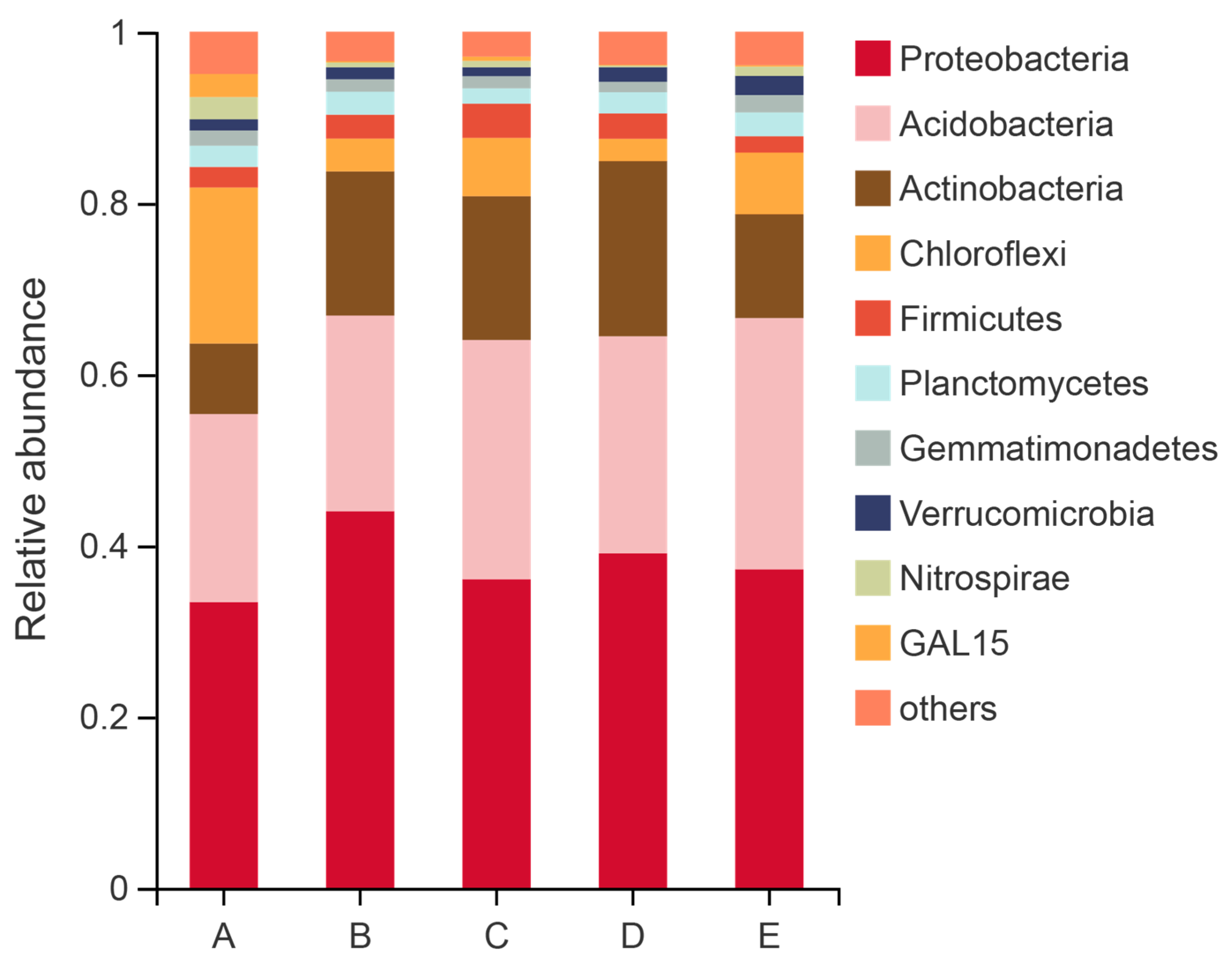
3.2. Changes in Soil Bacterial Community Composition and Diversity
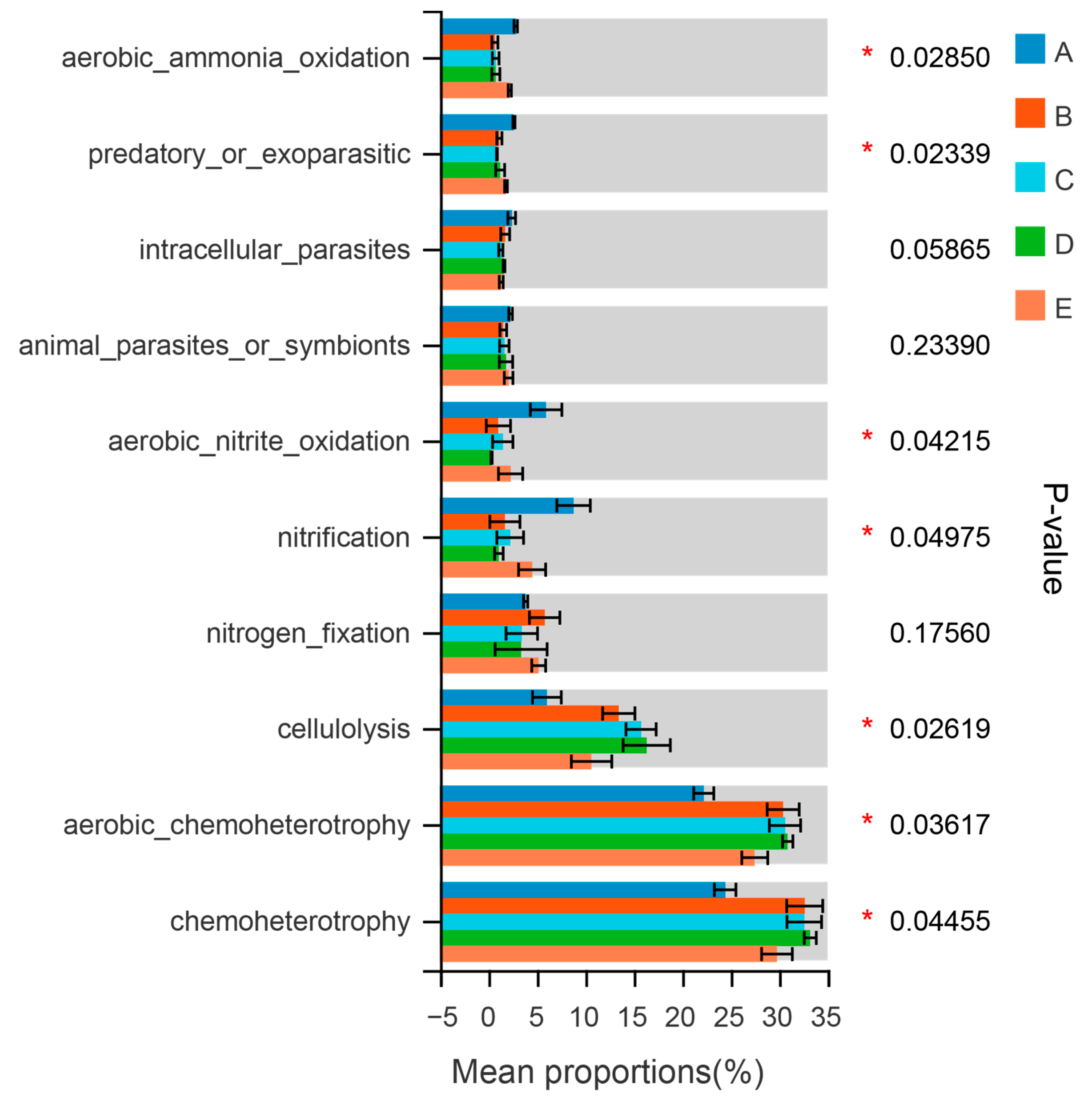
3.3. Ecological Function of Soil Bacterial Community
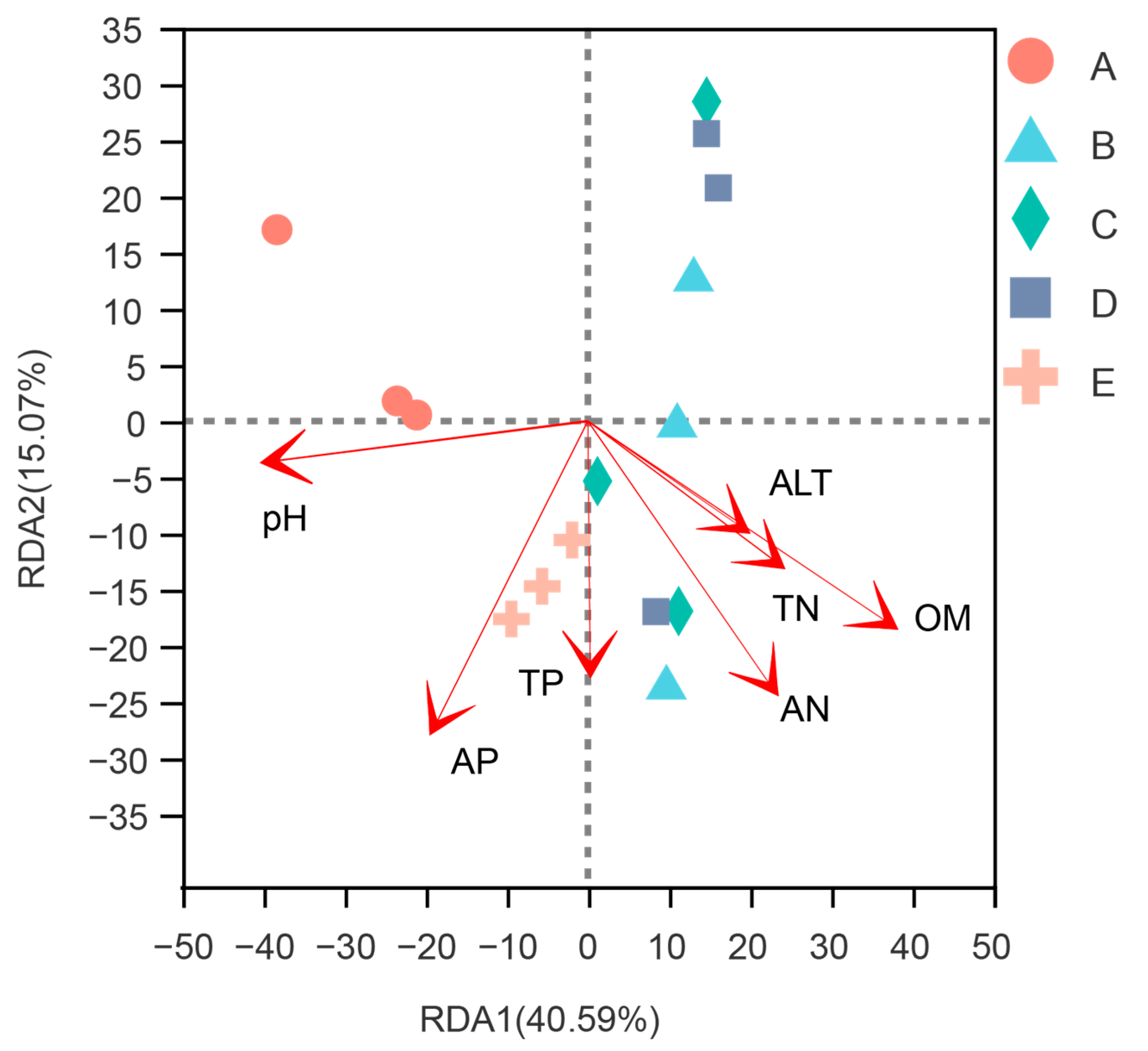
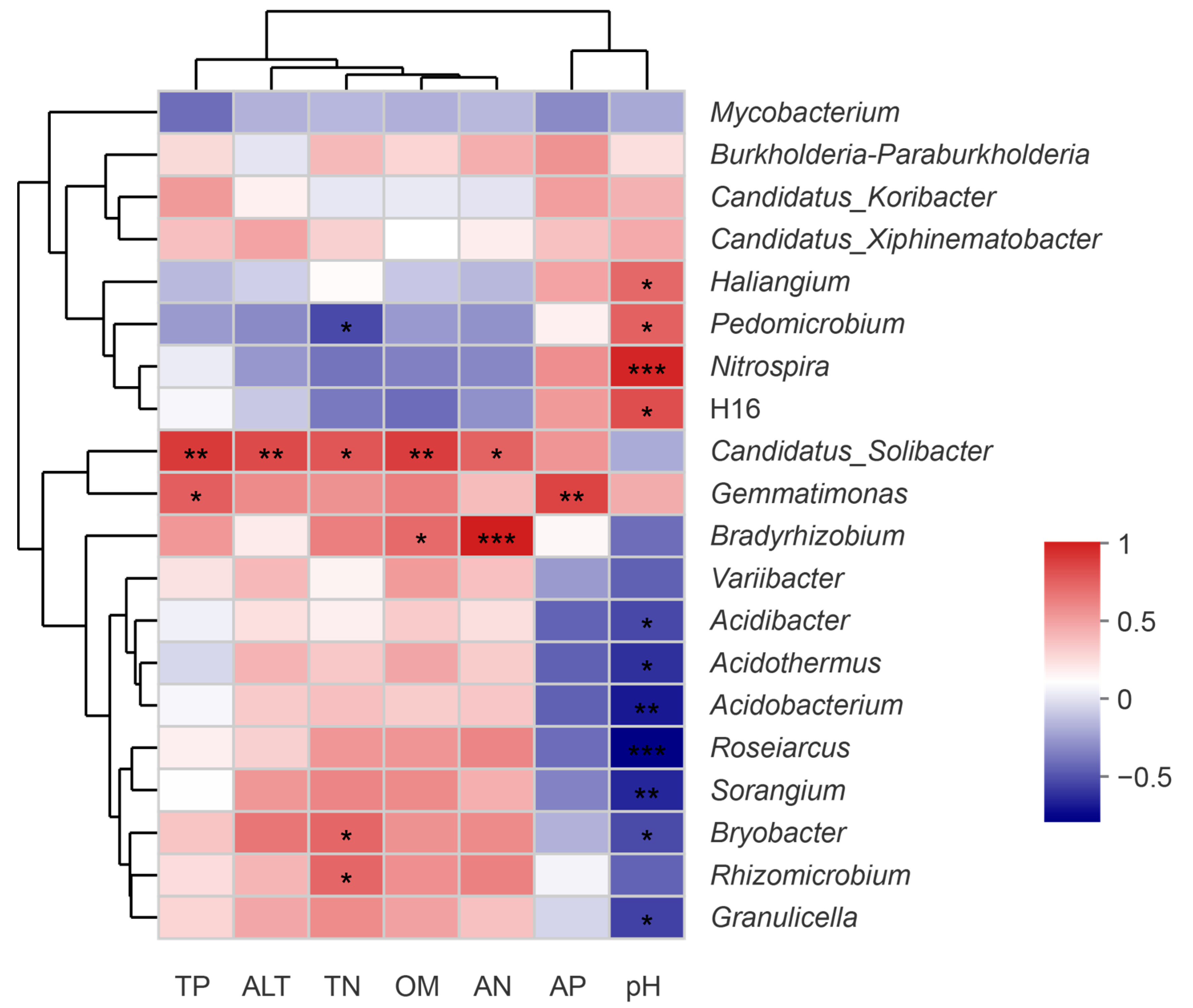
3.4. Relationships between Soil Chemical Properties, Altitude, and Soil Bacterial Community
4. Discussion
4.1. Soil Chemical Properties and Soil Enzymatic Activities
4.2. Changes of Soil Bacterial Community
4.3. Effects of Soil Chemical Properties and Altitude on Soil Bacterial Community
4.4. Endemism of C. pingbianense and Implication on Forest Management
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yang, H.Q.; Li, D.Z. Revision on Cephalostachyum Munro (Poaceae: Bambusoideae) in China. Plant Divers. Resour. 2015, 37, 546–550. [Google Scholar]
- Zheng, X.Q.; Cui, Y.Z.; Chen, L.N.; Yang, H.Q. Study on bamboo shooting and shoot growth of Cephalostachyum pingbianense. For. Res. 2018, 31, 131–136. [Google Scholar] [CrossRef]
- Yang, Y.M.; Xue, J.R. The preliminary study on natural bamboo forest in Yunnan Dawei mountain area. J. Southwest For. Univ. 1990, 1, 21–30. [Google Scholar]
- Mayr, E. Change of Genetic Environment and Evolution; Allen and Unwin: London, UK, 1954; pp. 157–180. [Google Scholar]
- Angert, A.L.; Schemske, D.W. The evolution of species’ distributions: Reciprocal transplants across the elevation ranges of Mimulus cardinalis and M. lewisii. Evolution 2005, 59, 1671–1684. [Google Scholar] [CrossRef] [PubMed]
- Ettinger, A.K.; Ford, K.R.; HilleRisLambers, J. Climate determines upper, but not lower, altitudinal range limits of Pacific Northwest conifers. Ecology 2011, 92, 1323–1331. [Google Scholar] [CrossRef] [Green Version]
- Grinnell, J. The Niche-Relationships of the California Thrasher. Auk 1917, 34, 427–433. [Google Scholar] [CrossRef]
- Benning, J.W.; Moeller, D.A. Maladaptation beyond a geographic range limit driven by antagonistic and mutualistic biotic interactions across an abiotic gradient. Evology 2019, 73, 2044–2059. [Google Scholar] [CrossRef]
- Theobald, E.J.; Gabrielyan, H.; HilleRisLambers, J. Lilies at the limit: Variation in plant-pollinator interactions across an elevational range. Am. J. Bot. 2016, 103, 189–197. [Google Scholar] [CrossRef] [Green Version]
- Benning, J.W.; Eckhart, V.M.; Geber, M.A.; Moeller, D.A. Biotic Interactions Contribute to the Geographic Range Limit of an Annual Plant: Herbivory and Phenology Mediate Fitness beyond a Range Margin. Am. Nat. 2019, 193, 786–797. [Google Scholar] [CrossRef] [Green Version]
- Lankau, R.A.; Keymer, D.P. Ectomycorrhizal fungal richness declines towards the host species’ range edge. Mol. Ecol. 2016, 25, 3224–3241. [Google Scholar] [CrossRef]
- Wallace, J.; Laforest-Lapointe, I.; Kembel, S.W. Variation in the leaf and root microbiome of sugar maple (Acer saccharum) at an elevational range limit. PeerJ 2018, 6, e5293. [Google Scholar] [CrossRef] [Green Version]
- Schütz, V.; Frindte, K.; Cui, J.; Zhang, P.; Hacquard, S.; Schulze-Lefert, P.; Knief, C.; Schulz, M.; Dörmann, P. Differential Impact of Plant Secondary Metabolites on the Soil Microbiota. Front. Microbiol. 2021, 12, 666010. [Google Scholar] [CrossRef]
- Huang, A.C.; Jiang, T.; Liu, Y.-X.; Bai, Y.-C.; Reed, J.; Qu, B.; Goossens, A.; Nützmann, H.-W.; Bai, Y.; Osbourn, A. A specialized metabolic network selectively modulates Arabidopsis root microbiota. Science 2019, 364, eaau6389. [Google Scholar] [CrossRef]
- Sánchez-Galindo, L.M.; Sandmann, D.; Marian, F.; Krashevska, V.; Maraun, M.; Scheu, S. Leaf litter identity rather than diversity shapes microbial functions and microarthropod abundance in tropical montane rainforests. Ecol. Evol. 2021, 11, 2360–2374. [Google Scholar] [CrossRef]
- Becklin, K.M.; Pallo, M.L.; Galen, C. Willows indirectly reduce arbuscular mycorrhizal fungal colonization in understorey communities. J. Ecol. 2012, 100, 343–351. [Google Scholar] [CrossRef]
- Trivedi, P.; Leach, J.E.; Tringe, S.G.; Sa, T.; Singh, B.K. Plant–microbiome interactions: From community assembly to plant health. Nat. Rev. Genet. 2020, 18, 1–15. [Google Scholar] [CrossRef]
- de Wit, R.; Bouvier, T. ‘Everything is everywhere, but, the environment selects’; what did Baas Becking and Beijerinck really say? Environ. Microbiol. 2006, 8, 755–758. [Google Scholar] [CrossRef]
- Burns, J.H.; Anacker, B.L.; Strauss, S.; Burke, D.J. Soil microbial community variation correlates most strongly with plant species identity, followed by soil chemistry, spatial location and plant genus. AoB Plants 2015, 7, plv030. [Google Scholar] [CrossRef] [Green Version]
- Lu, R.K. Analysis Methods of Soil Agrochemistry; China Agricultural Science and Technology Press: Beijing, China, 2000; pp. 147–195. [Google Scholar]
- Guan, S.Y. Soil Enzymes and Their Research Methods; Agriculture Press: Beijing, China, 1986. [Google Scholar]
- Xu, N.; Tan, G.; Wang, H.; Gai, X. Effect of biochar additions to soil on nitrogen leaching, microbial biomass and bacterial community structure. Eur. J. Soil Biol. 2016, 74, 1–8. [Google Scholar] [CrossRef]
- Kaushal, R.; Singh, I.; Thapliyal, S.D.; Gupta, A.K.; Mandal, D.; Tomar, J.M.S.; Kumar, A.; Alam, N.M.; Kadam, D.; Singh, D.V.; et al. Rooting behaviour and soil properties in different bamboo species of Western Himalayan Foothills, India. Sci. Rep. 2020, 10, 1–17. [Google Scholar] [CrossRef]
- Zhao, T.X.; Mao, X.W.; Cheng, M.; Chen, J.H.; Qin, H.; Li, Y.C.; Liang, C.F.; Xu, Q.F. Effects of Phyllostachys edulis cultivation on soil bacterial and fungal community structure and diversity. Chin. J. Appl. Ecol. 2017, 28, 3740–3750. [Google Scholar]
- Zhang, X.; Gao, G.; Wu, Z.; Wen, X.; Zhong, H.; Zhong, Z.; Yang, C.; Bian, F.; Gai, X. Responses of soil nutrients and microbial communities to intercropping medicinal plants in moso bamboo plantations in subtropical China. Environ. Sci. Pollut. Res. 2019, 27, 2301–2310. [Google Scholar] [CrossRef]
- Ahemad, M.; Zaidi, A.; Khan, M.S.; Oves, M. Biological Importance of Phosphorus and Phosphate Solubilizing Microbes; Nova Science Publishers: New York, NY, USA, 2009; pp. 1–14. [Google Scholar]
- Bojórquez-Quintal, E.; Escalante-Magaña, C.; Echevarría-Machado, I.; Martínez-Estévez, M. Aluminum, a Friend or Foe of Higher Plants in Acid Soils. Front. Plant Sci. 2017, 8, 1767. [Google Scholar] [CrossRef]
- Onweremadu, E. Predicting Phosphorus Sorption Characteristics in Highly Weathered Soils of South-Eastern Nigeria. Res. J. Environ. Sci. 2007, 1, 47–55. [Google Scholar] [CrossRef]
- Peng, Y.; Jiang, Y.; Liu, D.L.; Ruan, J.J. Effects of organic matter and microbial inoculants on agronomic characters of tartary buckwheat continuous cropping and soil enzyme activities. Mol. Plant Breed. 2021, 1–16. Available online: https://kns.cnki.net/kcms/detail/46.1068.S.20210415.1516.008.html (accessed on 6 November 2021).
- Chunyan, X.; Tao, W.; Chenbo, J.; Yang, G.; Jianyu, S. Effects of different desert plants on the soil chemical properties and enzyme activities in Helanshan eastern region. Ecol. Environ. Sci. 2020, 29, 2346–2354. [Google Scholar] [CrossRef]
- Haiyan, Z.; Fuli, X.; Weiling, W.; Weidong, W.; Qincheng, C.; Yafang, Z.; Yajuan, M. Soil nutrients and enzyme activities in Larix principis-rupprechtii plantations in the Qinling Mountains, China. Acta Ecol. Sin. 2015, 35, 1086–1094. [Google Scholar] [CrossRef] [Green Version]
- Talia, P.; Sede, S.M.; Campos, E.; Rorig, M.; Principi, D.; Tosto, D.; Hopp, H.E.; Grasso, D.; Cataldi, A. Biodiversity characterization of cellulolytic bacteria present on native Chaco soil by comparison of ribosomal RNA genes. Res. Microbiol. 2012, 163, 221–232. [Google Scholar] [CrossRef]
- Morvan, S.; Meglouli, H.; Sahraoui, A.L.; Hijri, M. Into the wild blueberry (Vaccinium angustifolium) rhizosphere microbiota. Environ. Microbiol. 2020, 22, 3803–3822. [Google Scholar] [CrossRef]
- Nie, H.; Qin, T.; Yan, D.; Lv, X.; Wang, J.; Huang, Y.; Lv, Z.; Liu, S.; Liu, F. How do tree species characteristics affect the bacterial community structure of subtropical natural mixed forests? Sci. Total Environ. 2021, 764, 144633. [Google Scholar] [CrossRef]
- Zi, H.; Jiang, Y.; Cheng, X.; Li, W.; Huang, X. Change of rhizospheric bacterial community of the ancient wild tea along elevational gradients in Ailao mountain, China. Sci. Rep. 2020, 10, 1–11. [Google Scholar] [CrossRef]
- Liu, W.; Wang, F.; Sun, Y.; Yang, L.; Chen, H.; Liu, W.; Zhu, B.; Hui, C.; Wang, S. Influence of dragon bamboo with different planting patterns on microbial community and physicochemical property of soil on sunny and shady slopes. J. Microbiol. 2020, 58, 906–914. [Google Scholar] [CrossRef]
- Fierer, N.; Bradford, M.; Jackson, R.B. Toward an ecological classification of soil bacteria. Ecology 2007, 88, 1354–1364. [Google Scholar] [CrossRef]
- Daims, H.; Lebedeva, E.V.; Pjevac, P.; Han, P.; Herbold, C.; Albertsen, M.; Jehmlich, N.; Palatinszky, M.; Vierheilig, J.; Bulaev, A.; et al. Complete nitrification by Nitrospira bacteria. Nature 2015, 528, 504–509. [Google Scholar] [CrossRef]
- Kits, K.D.; Sedlacek, C.; Lebedeva, E.V.; Han, P.; Bulaev, A.; Pjevac, P.; Daebeler, A.; Romano, S.; Albertsen, M.; Stein, L.Y.; et al. Kinetic analysis of a complete nitrifier reveals an oligotrophic lifestyle. Nature 2017, 549, 269–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Q.N. The Effects for Phyllostachys pubscens Expansion on Nitrogen and Phosphorus Distribution Pattern and Process of Evergreen Broadleaved Forest. Ph.D. Thesis, Tsinghua University, Beijing, China, 2017. [Google Scholar]
- Junjie, X.; Zhonghua, C.; Jin, Z. Research progress of carbohydrate-active enzymes and the degradation mechanisms by marine bacteria. Acta Microbiol. Sin. 2021; accepted. [Google Scholar] [CrossRef]
- Li, S.X. Soil and Plant Nitrogen in Dryland Areas of China; Science Press: Beijing, China, 2008. [Google Scholar]
- Song, Q.N.; Yang, Q.P.; Liu, J.; Yu, D.K.; Fang, K.; Xu, P.; He, Y.J. Effects of Phyllostachys edulis expansion onsoil nitrogen mineralization and its availability in evergreen broadleaf forest. Chin. J. Appl. Ecol. 2013, 24, 338–344. [Google Scholar] [CrossRef]
- Rheault, K.; Lachance, D.; Morency, M.-J.; Thiffault, É.; Guittonny, M.; Isabel, N.; Martineau, C.; Séguin, A. Plant Genotype Influences Physicochemical Properties of Substrate as Well as Bacterial and Fungal Assemblages in the Rhizosphere of Balsam Poplar. Front. Microbiol. 2020, 11, 575625. [Google Scholar] [CrossRef]
- Kitamura, K.; Uchiyama, K.; Ueno, S.; Ishizuka, W.; Tsuyama, I.; Goto, S. Geographical Gradients of Genetic Diversity and Differentiation among the Southernmost Marginal Populations of Abies sachalinensis Revealed by EST-SSR Polymorphism. Forests 2020, 11, 233. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Sork, V.L.; Meng, H.; Jiang, M. Genetic evidence for central-marginal hypothesis in a Cenozoic relict tree species across its distribution in China. J. Biogeogr. 2016, 43, 2173–2185. [Google Scholar] [CrossRef]
- Bahram, M.; Hildebrand, F.; Forslund, S.K.; Anderson, J.L.; Soudzilovskaia, N.A.; van Bodegom, P.; Bengtsson-Palme, J.; Anslan, S.; Coelho, L.P.; Harend, H.; et al. Structure and function of the global topsoil microbiome. Nature 2018, 560, 233–237. [Google Scholar] [CrossRef]
- Yao, F.; Yang, S.; Wang, Z.; Wang, X.; Ye, J.; Wang, X.; De Bruyn, J.; Feng, X.; Jiang, Y.; Li, H. Microbial Taxa Distribution Is Associated with Ecological Trophic Cascades along an Elevation Gradient. Front. Microbiol. 2017, 8, 2071. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.; Wang, C.; Luo, Y. Meta-analysis of the impacts of global change factors on soil microbial diversity and functionality. Nat. Commun. 2020, 11, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Huntemann, M.; Lu, M.; Nolan, M.; Lapidus, A.; Lucas, S.; Hammon, N.; Deshpande, S.; Cheng, J.-F.; Tapia, R.; Han, C.; et al. Complete genome sequence of the thermophilic sulfur-reducer Hippea maritima type strain (MH 2 T). Stand. Genom. Sci. 2011, 4, 303–311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, Q.; Liu, Z.; Huang, Y.; Li, F.; Nengzi, L.; Zhang, J. Influence of temperature on CODMn and Mn2+ removal and microbial community structure in pilot-scale biofilter. Bioresour. Technol. 2020, 316, 123968. [Google Scholar] [CrossRef]
- Xun, W.; Liu, Y.; Li, W.; Ren, Y.; Xiong, W.; Xu, Z.; Zhang, N.; Miao, Y.; Shen, Q.; Zhang, R. Specialized metabolic functions of keystone taxa sustain soil microbiome stability. Microbiome 2021, 9, 1–15. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Types | Shannon | Chao1 | Richness | Phylogenetic Diversity |
|---|---|---|---|---|
| A | 6.47 ± 0.07 a | 3325 ± 211 a | 2431 ± 174 a | 210 ± 10 a |
| B | 5.90 ± 0.08 c | 2762 ± 29 b | 1830 ± 44 b | 154 ± 6 b |
| C | 5.83 ± 0.09 c | 2757 ± 168 b | 1736 ± 138 b | 148 ± 9 b |
| D | 5.90 ± 0.04 c | 2764 ± 52 b | 1860 ± 31 b | 155 ± 2 b |
| E | 6.15 ± 0.05 b | 2780 ± 97 b | 1963 ± 58 b | 163 ± 4 b |
| Bacteria with Significant Differences | A (%) | B (%) | C (%) | D (%) | E (%) |
|---|---|---|---|---|---|
| Actinobacteria | 8.24 ± 1.74 b | 16.84 ± 0.94 ab | 16.80 ± 2.79 ab | 20.46 ± 1.49 a | 12.13 ± 3.76 b |
| Chloroflexi | 18.22 ± 4.14 a | 3.83 ± 3.23 b | 6.82 ± 2.58 b | 2.61 ± 0.21 b | 7.20 ± 2.87 b |
| Gemmatimonadetes | 1.76 ± 0.48 a | 1.45 ± 0.46 a | 1.42 ± 0.65 a | 1.23 ± 0.31 b | 2.04 ± 0.26 a |
| Nitrospirae | 2.61 ± 0.52 a | 0.56 ± 0.63 b | 0.75 ± 0.48 b | 0.21 ± 0.04 b | 1.63 ± 0.42 b |
| Variibacter | 4.33 ± 1.45 b | 8.60 ± 1.12 a | 8.34 ± 1.25 a | 8.37 ± 0.12 a | 6.66 ± 0.45 ab |
| Acidothermus | 2.27 ± 0.83 b | 6.94 ± 0.75 ab | 7.59 ± 1.91 ab | 9.59 ± 2.02 a | 4.19 ± 1.23 b |
| Acidibacter | 1.88 ± 0.28 b | 2.89 ± 0.62 ab | 2.53 ± 0.43 ab | 3.03 ± 0.74 a | 2..02 ± 0.39 b |
| Roseiarcus | 0.69 ± 0.15 b | 3.21 ± 1.53 a | 1.76 ± 0.38 ab | 2.28 ± 0.23 ab | 1.13 ± 0.39 b |
| Nitrospira | 2.61 ± 0.52 a | 0.56 ± 0.63 b | 0.75 ± 0.48 b | 0.21 ± 0.04 b | 1.09 ± 0.56 b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, T.; Li, L.; Li, B.; Dou, P.; Yang, H. Heterotrophic Bacteria Play an Important Role in Endemism of Cephalostachyum pingbianense (Hsueh & Y.M. Yang ex Yi et al.) D.Z. Li & H.Q. Yang, 2007, a Full-Year Shooting Woody Bamboo. Forests 2022, 13, 121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010121
Xia T, Li L, Li B, Dou P, Yang H. Heterotrophic Bacteria Play an Important Role in Endemism of Cephalostachyum pingbianense (Hsueh & Y.M. Yang ex Yi et al.) D.Z. Li & H.Q. Yang, 2007, a Full-Year Shooting Woody Bamboo. Forests. 2022; 13(1):121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010121
Chicago/Turabian StyleXia, Tize, Lushuang Li, Bin Li, Peitong Dou, and Hanqi Yang. 2022. "Heterotrophic Bacteria Play an Important Role in Endemism of Cephalostachyum pingbianense (Hsueh & Y.M. Yang ex Yi et al.) D.Z. Li & H.Q. Yang, 2007, a Full-Year Shooting Woody Bamboo" Forests 13, no. 1: 121. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010121