
The Effects of Plant and Soil Characteristics on Partitioning Different Rainfalls to Soil in a Subtropical Chinese Fir Forest Ecosystem

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Site Description

2.2. Rainfall Event Selection and Sample Collection
2.3. Sample Preparation and Stable Isotope Analysis
2.4. Contribution of Rainfall to Soil Water
2.5. Measurement of the Vegetation and Soil Parameters
2.6. Data Analysis
3. Results
3.1. Temporal Variation in the Soil Water Content after Different Rainfall Events
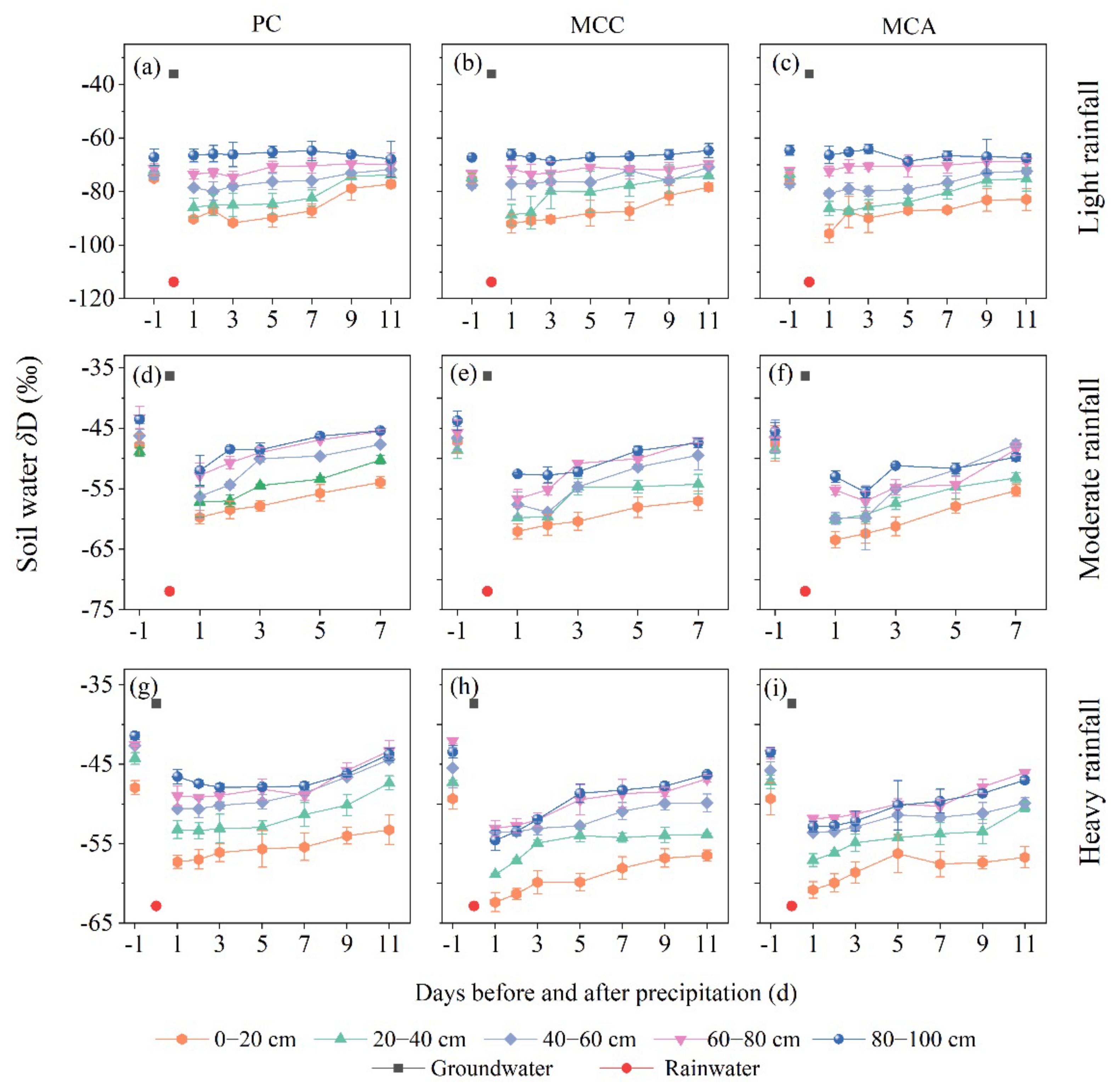
3.2. Temporal Variation in the Soil Water δD after Different Rainfall Events
3.3. Variation of CRSW among the Three Plantations
3.4. The Correlation of Vegetation Biomass, Litter Characteristics, Root Biomass and Soil Properties with the CRSW

3.5. The Factors Affecting the CRSW under Different Rainfall Events
4. Discussion
4.1. The Response of the SWC and Soil Water δD to Rainfall Events
4.2. The Variation in the CRSW in Different Plantations and Its Direct Regulators
4.3. Key Indirect Regulators of the CRSW
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Legates, D.R.; Mahmood, R.; Levia, D.F.; Deliberty, T.L.; Quiring, S.M.; Houser, C.; Nelson, F.E. Soil moisture: A central and unifying theme in physical geography. Prog. Phys. Geog. 2011, 35, 65–86. [Google Scholar] [CrossRef]
- Moreno-Gutierrez, C.; Dawson, T.E.; Nicolas, E.; Querejeta, J.I. Isotopes reveal contrasting water use strategies among coexisting plant species in a Mediterranean ecosystem. New Phytol. 2012, 196, 489–496. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Lei, H.; Yang, D.; Huang, M.; Liu, D.; Yuan, X. Impact of vegetation dynamics on hydrological processes in a semi-arid basin by using a land surface-hydrology coupled model. J. Hydrol. 2017, 551, 116–131. [Google Scholar] [CrossRef]
- Wang, C.; Fu, B.; Zhang, L.; Xu, Z. Soil moisture-plant interactions: An ecohydrological review. J. Soil Sediment. 2019, 19, 1–9. [Google Scholar] [CrossRef]
- Jiao, L.; An, W.; Li, Z.; Gao, G.; Wang, C. Regional variation in soil water and vegetation characteristics in the Chinese Loess Plateau. Ecol. Indic. 2020, 115, 106399. [Google Scholar] [CrossRef]
- Chen, G.; Tan, Q.; Xu, M.; Liu, G. Mixed-species plantations can alleviate water stress on the Loess Plateau. For. Ecol. Manag. 2020, 458, 117767. [Google Scholar]
- Fischer, C.; Roscher, C.; Jensen, B.; Eisenhauer, N.; Baade, J.; Attinger, S.; Scheu, S.; Weisser, W.W.; Schumacher, J.; Hildebrandt, A. How do earthworms, soil texture and plant composition affect infiltration along an experimental plant diversity gradient in grassland? PLoS ONE 2014, 9, e98987. [Google Scholar] [CrossRef] [PubMed]
- Taylor, C.M.; Belusic, D.; Guichard, F.; Arker, D.; Vischel, T.; Bock, O.; Harris, P.P.; Janicot, S.; Klein, C.; Panthou, G. Frequency of extreme Sahelian storms tripled since 1982 in satellite observations. Nature 2017, 544, 475–478. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Myhre, G.; Samset, B.; Hodnebrog, Ø.; Andrews, T.; Boucher, O.; Faluvegi, G.; Fläschner, D.; Forster, P.M.; Kasoar, M.; Kharin, V.; et al. Sensible heat has significantly affected the global hydrological cycle over the historical period. Nat. Commun. 2018, 9, 1922. [Google Scholar] [CrossRef] [PubMed]
- Metzger, J.C.; Wutzler, T.; Valle, N.D.; Filipzik, J.; Hildebrandt, A. Vegetation impacts soil water content patterns by shaping canopy water fluxes and soil properties. Hydrol. Process. 2017, 31, 3783–3795. [Google Scholar] [CrossRef]
- Solomou, A.D.; Skoufogianni, E.; Danalatos, N.G. Exploitation of soil properties for controlling herbaceous plant communities in an organic cultivation of lippia citriodora in the mediterranean landscape. Bulg. J. Agric. Sci. 2020, 26, 7983. [Google Scholar]
- Li, X.; Xiao, Q.; Niu, J.; Dymond, S.; Doorn, N.; Yu, X.; Xie, B.; Lv, X.; Zhang, K.; Li, J. Process-based rainfall interception by small trees in Northern China: The effect of rainfall traits and crown structure characteristics. Agric. For. Meteorol. 2016, 218, 65–73. [Google Scholar] [CrossRef]
- Song, X.; Lyu, S.; Wen, X. Limitation of soil moisture on the response of transpiration to vapor pressure deficit in a subtropical coniferous plantation subjected to seasonal drought. J. Hydrol. 2020, 591, 125301. [Google Scholar] [CrossRef]
- Zhao, C.; Jia, X.; Zhu, Y.; Shao, M. Long-term temporal variations of soil water content under different vegetation types in the Loess Plateau, China. Catena 2017, 158, 55–62. [Google Scholar] [CrossRef]
- Saija, H.; Timo, D.; Leena, F.; Jarkko, H.; Juho, M.; Jari, M.; Seppo, N.; Pentti, N.; Ari, N.; Tuula, P.; et al. What is the potential for replacing monocultures with mixed-species stands to enhance ecosystem services in boreal forests in Fennoscandia? For. Ecol. Manag. 2021, 479, 118558. [Google Scholar]
- Bouwman, M.; Forrester, D.I.; Ouden, J.D.; Nabuurs, G.J.; Mohren, G. Species interactions under climate change in mixed stands of Scots pine and pedunculate oak. For. Ecol. Manag. 2020, 481, 118615. [Google Scholar] [CrossRef]
- Pretzsch, H.; Schütze, G.; Uhl, E. Resistance of European tree species to drought stress in mixed versus pure forests: Evidence of stress release by inter-specific facilitation. Plant Biology 2013, 15, 483–495. [Google Scholar] [CrossRef] [PubMed]
- Metz, J.; Annighofer, P.; Schall, P.; Zimmermann, J.; Kahl, T.; Schulze, E.D.; Ammer, C. Site-adapted admixed tree species reduce drought susceptibility of mature European beech. Glob. Chang. Biol. 2016, 22, 903–920. [Google Scholar] [CrossRef]
- Sui, M.; Zhang, B.; Xu, Q.; Gao, D.; Zhang, Y.; Wang, S. Effects of plantation types and patterns on rainfall partition in soil in a mid-subtropical region of China. Plant Soil 2021, 466, 1–15. [Google Scholar] [CrossRef]
- Primicia, I.; Camarero, J.J.; Imbert, J.B.; Castillo, F.J. Effects of thinning and canopy type on growth dynamics of Pinus sylvestris: Inter-annual variations and intra-annual interactions with microclimate. Eur. J. For. Res. 2013, 132, 121–135. [Google Scholar] [CrossRef]
- Niether, W.; Schneidewind, U.; Armengot, L.; Adamtey, N.; Schneider, M.; Gerold, G. Spatial-temporal soil moisture dynamics under different cocoa production systems. Catena 2017, 158, 340–349. [Google Scholar] [CrossRef]
- Sun, L.; Yang, L.; Chen, L.; Li, S.; Zhao, F.; Sun, S. Tracing the soil water response to autumn rainfall in different land uses at multi-day timescale in a subtropical zone. Catena 2019, 180, 355–364. [Google Scholar] [CrossRef]
- Li, B.; Biswas, A.; Wang, Y.; Li, Z. Identifying the dominant effects of climate and land use change on soil water balance in deep loessial vadose zone. Agr. Water Manag. 2020, 245, 106637. [Google Scholar] [CrossRef]
- Dai, L.; Yuan, Y.; Guo, X.; Du, Y.; Cao, G. Soil water retention in alpine meadows under different degradation stages on the northeastern Qinghai-Tibet Plateau. J. Hydrol. 2020, 590, 125397. [Google Scholar] [CrossRef]
- Wei, X.; Liang, W. Regulation of stand density alters forest structure and soil moisture during afforestation with Robinia pseudoacacia L. Pinus tabulaeformis Carr. On the Loess Plateau. For. Ecol. Manag. 2021, 491, 119196. [Google Scholar]
- Korres, W.; Reichenau, T.G.; Fiener, P.; Koyama, C.N.; Bogena, H.R.; Cornelissen, T.; Baatz, R.; Herbst, V.; Diekkrüger, B.; Vereecken, H.; et al. Spatio-temporal soil moisture patterns—A meta-analysis using plot to catchment scale data. J. Hydrol. 2014, 520, 326–341. [Google Scholar] [CrossRef] [Green Version]
- Yue, K.; Frenne, P.D.; Fornara, D.A.; Meerbeek, K.V.; Li, W.; Peng, X.; Ni, X.; Peng, Y.; Wu, F.; Yang, Y.; et al. Global patterns and drivers of rainfall partitioning by trees and shrubs. Glob. Chang. Biol. 2021, 27, 3350–3357. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Xu, Q.; Gao, D.; Jiang, C.; Ma, Y. Higher soil capacity of intercepting heavy rainfall in mixed stands than in pure stands in riparian forests. Sci. Total Environ. 2019, 658, 1514–1522. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Guo, L.; Huang, Z.; Wu, G.; Lopez-Vicente, M. Root morphological characteristics and soil water infiltration capacity in semi-arid artificial grassland soils. Agric. Water Manag. 2020, 235, 106153. [Google Scholar] [CrossRef]
- Thompson, S.E.; Harman, C.J.; Heine, P.; Katul, G.G. Vegetation-infiltration relationships across climatic and soil type gradients. J. Geophys. Res. Biogeo. 2015, 115, G02023. [Google Scholar] [CrossRef] [Green Version]
- Dubbert, M.; Werner, C. Water fluxes mediated by vegetation: Emerging insights at the soil and atmosphere interfaces. New Phytol. 2018, 221, 1754–1763. [Google Scholar] [CrossRef]
- Zhou, G.; Houlton, B.Z.; Wang, W.; Huang, W.; Xiao, Y.; Zhang, Q.; Liu, S.; Cao, M.; Wang, X.; Wang, S.; et al. Substantial reorganization of China’s tropical and subtropical forests: Based on the permanent plots. Glob. Chang. Biol. 2014, 20, 240–250. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Peng, C.; Li, Y.; Liu, S.; Zhang, Q.; Tang, X.; Liu, J.; Yan, J.; Zhang, D.; Chu, G. A climate change-induced threat to the ecological resilience of a subtropical monsoon evergreen broad-leaved forest in Southern China. Glob. Chang. Biol. 2013, 19, 1197–1210. [Google Scholar] [CrossRef]
- Gaisberger, H.; Fremout, T.; Kettle, C.J.; Barbara, V.; Kemalasari, D.; Kanchanarak, T.; Thomas, E.; Serra-Diaz, J.M.; Svenning, J.C.; Slik, F.; et al. Tropical and subtropical Asia’s valued tree species under threat. Conserv. Biol. 2021, accepted. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhang, Z.; Zhu, B.; Cheng, X.; Yang, L.; Gao, M.; Kong, R. Maxent modeling based on CMIP6 models to project potential suitable zones for Cunninghamia lanceolata in China. Forests 2021, 12, 752. [Google Scholar] [CrossRef]
- SFA. China Forest Resources Report (2009–2013); State Forest Administration: Beijing, China, 2014; 75p. [Google Scholar]
- Guan, F.; Tang, X.; Fan, S.; Zhao, J.; Peng, C. Changes in soil carbon and nitrogen stocks followed the conversion from secondary forest to Chinese fir and Moso bamboo plantations. Catena 2015, 133, 455–460. [Google Scholar] [CrossRef]
- Penna, D.; Hopp, L.; Scandellari, F.; Allen, S.T.; Benettin, P.; Beyer, M.; Geris, J.; Klaus, J.; Marshall, J.D.; Schwendenmann, L.; et al. Ideas and perspectives: Tracing terrestrial ecosystem water fluxes using hydrogen and oxygen stable isotopes—Challenges and opportunities from an interdisciplinary perspective. Biogeosciences 2018, 15, 6399–6415. [Google Scholar] [CrossRef] [Green Version]
- Sprenger, M.; Leistert, H.; Gimbel, K.; Weiler, M. Illuminating hydrological processes at the soil-vegetation-atmosphere interface with water stable isotopes. Rev. Geophy. 2016, 54, 674–704. [Google Scholar] [CrossRef] [Green Version]
- Gazis, C.; Feng, X. A stable isotope study of soil water: Evidence for mixing and preferential flow paths. Geoderma 2004, 119, 97–111. [Google Scholar] [CrossRef]
- Wan, H.; Liu, W. An isotope study (δ18O and δD) of water movements on the Loess Plateau of China in arid and semiarid climates. Ecol. Eng. 2016, 93, 226–233. [Google Scholar] [CrossRef]
- Xu, Q.; Liu, S.; Wan, X.; Jiang, C.; Song, X.; Wang, J. Effects of rainfall on soil moisture and water movement in a subalpine dark coniferous forest in southwestern China. Hydrol. Process. 2012, 26, 3800–3809. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, S.; Yu, H. Comparisons of litterfall, litter decomposition and nutrient return in a monoculture Cunninghamia lanceolata and a mixed stand in southern China. For. Ecol. Manag. 2008, 255, 1210–1218. [Google Scholar] [CrossRef]
- Zhao, P.; Tang, X.; Zhao, P.; Zhang, W.; Tang, J. Mixing of event and pre-event water in a shallow Entisol in sloping farmland based on isotopic and hydrometric measurements, SW China. Hydrol. Process. 2016, 30, 3478–3493. [Google Scholar] [CrossRef]
- Lin, G.; Phillips, S.L.; Ehleringer, J.R. Monosoonal precipitation responses of shrubs in a cold desert community on the Colorado Plateau. Oecologia 1996, 106, 8–17. [Google Scholar] [CrossRef] [PubMed]
- West, A.G.; Patrickson, S.J.; Ehleringer, J.R. Water extraction times for plant and soil materials used in stable isotope analysis. Rapid Commun. Mass Sp. 2006, 20, 1317–1321. [Google Scholar] [CrossRef]
- White, J.; Cook, E.R.; Lawrence, J.R.; Broecker, W.S. The D/H ratios of sap in trees: Implications for water sources and tree ring D/H ratios. Geochim. Cosmochim. Acta 1985, 49, 237–246. [Google Scholar] [CrossRef]
- Bai, Y.; Zhou, Y.; Du, J.; Zhang, X.; Di, N. Effects of a broadleaf-oriented transformation of coniferous plantations on the hydrological characteristics of litter layers in subtropical China. Glob. Ecol. Conserv. 2020, 25, e01400. [Google Scholar] [CrossRef]
- Özcan, M.; GÖkbulak, F.; Hizal, A. Exclosure effects on recovery of selected soil properties in a mixed broadleaf forest recreation site. Land Degrad. Dev. 2013, 24, 266–276. [Google Scholar] [CrossRef]
- Dunkerley, D. Percolation through leaf litter: What happens during rainfall events of varying intensity? J. Hydrol. 2015, 525, 737–746. [Google Scholar] [CrossRef]
- Meier, I.C.; Knutzen, F.; Eder, L.M.; Müller-Haubold, H.; Leuschner, C. The deep root system of Fagus sylvatica on sandy soil: Structure and variation across a precipitation gradient. Ecosystems 2017, 21, 280–296. [Google Scholar] [CrossRef]
- Wu, H.; Li, X.Y.; Jiang, Z.; Chen, H.; Zhang, C.; Xiao, X. Contrasting water use pattern of introduced and native plants in an alpine desert ecosystem, Northeast Qinghai–Tibet Plateau, China. Sci. Total Environ. 2016, 542, 182–191. [Google Scholar] [CrossRef]
- Crockford, R.H.; Richardson, D.P. Partitioning of rainfall into throughfall, stemflow and interception: Effect of forest type, ground cover and climate. Hydrol. Process. 2015, 14, 2903–2920. [Google Scholar] [CrossRef]
- Daly, E.; Porporato, A. A review of soil moisture dynamics: From rainfall infiltration to ecosystem response. Environ. Eng. Sci. 2005, 22, 9–24. [Google Scholar] [CrossRef]
- Geroy, I.J.; Gribb, M.M.; Marshall, H.P.; Chandler, D.G.; Benner, S.G.; Mcnamara, J.P. Aspect influences on soil water retention and storage. Hydrol. Process. 2011, 25, 3836–3842. [Google Scholar] [CrossRef]
- Sun, D.; Yang, H.; Guan, D.; Yang, M.; Wu, J.; Yuan, F.; Jin, C.; Wang, A.; Zhang, Y. The effects of land use change on soil infiltration capacity in China: A meta-analysis. Sci. Total Environ. 2018, 626, 1394–1401. [Google Scholar] [CrossRef]
- Yinglan, A.; Wang, G.; Liu, T.; Shrestha, S.; Xue, B.; Tan, Z. Vertical variations of soil water and its controlling factors based on the structural equation model in a semi-arid grassland. Sci. Total Environ. 2019, 691, 1016–1026. [Google Scholar]
- Gao, X.; Li, H.; Zhao, X.; Ma, W.; Wu, P. Identifying a suitable revegetation technique for soil restoration on water-limited and degraded land: Considering both deep soil moisture deficit and soil organic carbon sequestration. Geoderma 2018, 319, 61–69. [Google Scholar] [CrossRef]
- Leung, A.; Garg, A.; Coo, J.; Ng, C.; Hau, C. Effects of the roots of Cynodon dactylon and Schefflera heptaphylla on water infiltration rate and soil hydraulic conductivity. Hydrol. Process. 2015, 29, 3342–3354. [Google Scholar] [CrossRef] [Green Version]
- Marin, C.T.; Bouten, W.; Sevink, J. Gross rainfall and its partitioning into throughfall, stemflow and evaporation of intercepted water in four forest ecosystems in western Amazonia. J. Hydrol. 2000, 237, 40–57. [Google Scholar] [CrossRef]
- Liu, Z.; Ma, D.; Hu, W.; Li, X. Land use dependent variation of soil water infiltration characteristics and their scale-specific controls. Soil Till. Res. 2018, 178, 139–149. [Google Scholar] [CrossRef]
- Zhao, S.W.; Zhao, Y.G.; Wu, J.S. Quantitative analysis of soil pores under natural vegetation successions on the Loess Plateau. Sci. China Earth Sci. 2010, 53, 617–625. [Google Scholar] [CrossRef]
- Alaoui, A. Modelling susceptibility of grassland soil to macropore flow. J. Hydrol. 2015, 525, 536–546. [Google Scholar] [CrossRef]
- Zhi, W.; Hua, Z.; Jrsc, D.; Zoa, B. Plant functional diversity mediates indirect effects of land-use intensity on soil water conservation in the dry season of tropical areas. For. Ecol. Manag. 2021, 480, 118646. [Google Scholar]
- Sun, J.; Yu, X.; Wang, H.; Jia, G.; Chen, J. Effects of forest structure on hydrological processes in China. J. Hydrol. 2018, 561, 187–199. [Google Scholar] [CrossRef]
- Huang, Z.; Tian, F.; Wu, G.; Liu, Y.; Dang, Z. Legume grasslands promote precipitation infiltration better than Gramineous grasslands in the arid regions. Land Deg. Dev. 2016, 28, 309–316. [Google Scholar] [CrossRef]
- Hudek, C.; Stanchi, S.; D’Amico, M.E.; Freppaz, M. Quantifying the contribution of the root system of alpine vegetation in the soil aggregate stability of moraine. J. Soil Water Conserv. 2017, 5, 36–42. [Google Scholar] [CrossRef]
- Bormann, H.; Klaassen, K. Seasonal and land use dependent variability of soil hydraulic and soil hydrological properties of two Northern German soils. Geoderma 2008, 145, 295–302. [Google Scholar] [CrossRef]
- Clemmensen, K.E.; Bahr, A.; Ovaskainen, O.; Dahlberg, A.; Ekblad, A.; Wallander, H.; Stenlid, J.; Finlay, R.D.; Wardle, D.A.; Lindahl, B.D. Roots and associated fungi drive long-term carbon sequestration in boreal forest. Science 2013, 339, 1615–16188. [Google Scholar] [CrossRef]
- Perie and Ouimet. Organic carbon, organic matter and bulk density relationships in boreal forest soils. Can. J. Soil 2008, 88, 315–325. [Google Scholar] [CrossRef]
- Yan, T.; Wang, Z.; Liao, C.; Xu, W.; Wan, L. Effects of the morphological characteristics of plants on rainfall interception and kinetic energy. J. Hydrol. 2020, 592, 106689. [Google Scholar] [CrossRef]
- Wang, B.; Verheyen, K.; Baeten, L.; Smedt, P.D. Herb litter mediates tree litter decomposition and soil fauna composition. Soil Biol. Biochem. 2020, 152, 10863. [Google Scholar] [CrossRef]
- Mishra, S.; Singh, K.; Sahu, N.; Singh, S.N.; Manika, N.; Chaudhary, L.B.; Jain, M.K.; Kumar, V.; Behera, S.K. Understanding the relationship between soil properties and litter chemistry in three forest communities in tropical forest ecosystem. Environ. Monit. Assess. 2019, 191, 797. [Google Scholar] [CrossRef] [PubMed]
- Jin, T.T.; Fu, B.J.; Liu, G.H.; Wang, Z. Hydrologic feasibility of artificial forestation in the semi-arid Loess Plateau of China. Hydrol. Earth Syst. Sci. 2011, 15, 2519–2530. [Google Scholar] [CrossRef] [Green Version]
- Robichaud, P.R. Fire effects on infiltration rates after prescribed fire in Northern Rocky Mountain forests, USA. J. Hydrol. 2000, 231, 220–229. [Google Scholar] [CrossRef]
- Pavão, L.L.; Sanches, L.; Júnior, O.P.; Spolador, J. The influence of litter on soil hydro-physical characteristics in an area of Acuri palm in the Brazilian Pantanal. Ecohydrol. Hydrobiol. 2019, 19, 642–650. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vegetation and Litter Characteristics | Variable | PC | MCC | MCA |
|---|---|---|---|---|
| Above-ground biomass | Tree biomass (kg·m−2) | 10.34 ± 1.69 b | 20.14 ± 0.93 a | 22.46 ± 1.00 a |
| Shrub biomass (g·m−2) | 2.68 ± 0.02 b | 12.54 ± 2.12 a | 10.93 ± 0.84 a | |
| Herb biomass (g·m−2) | 269.31 ± 6.43 b | 399.35 ± 46.22 a | 319.40 ± 8.54 b | |
| Litter characteristics | Litter mass (t·hm−2) | 7.94 ± 0.22 b | 8.83 ± 0.17 a | 8.88 ± 0.53 a |
| EHC (t·hm−2) | 12.62 ± 1.44 b | 17.94 ± 0.97 a | 17.62 ± 2.26 a | |
| MWAR (g·kg−1·h−1) | 81.44 ± 7.31 b | 100.27 ± 4.52 a | 97.47 ± 10.65 a |
| Variable | Layers (cm) | PC | MCC | MCA |
|---|---|---|---|---|
| Bulk density (g·m−3) | 0–20 | 1.31 ± 0.02 a | 1.25 ± 0.02 b | 1.24 ± 0.03 b |
| 20–40 | 1.36 ± 0.01 a | 1.33 ± 0.02 b | 1.33 ± 0.01 b | |
| 40–60 | 1.42 ± 0.03 a | 1.34 ± 0.02 b | 1.34 ± 0.04 b | |
| 60–80 | 1.46 ± 0.02 a | 1.41 ± 0.05 b | 1.40 ± 0.05 a | |
| 80–100 | 1.46 ± 0.01 a | 1.42 ± 0.03 ab | 1.42 ± 0.06 b | |
| Field capacity (%) | 0–20 | 35.01 ± 0.99 b | 41.48 ± 1.19 a | 40.69 ± 3.61 a |
| 20–40 | 28.75 ± 1.34 c | 38.23 ± 0.56 a | 35.10 ± 1.37 b | |
| 40–60 | 27.88 ± 1.91 b | 42.33 ± 3.65 a | 31.99 ± 1.57 a | |
| 60–80 | 34.49 ± 4.10 a | 35.98 ± 4.61 a | 31.58 ± 1.44 a | |
| 80–100 | 25.27 ± 3.50 b | 32.18 ± 0.48 a | 36.82 ± 3.18 a | |
| Total porosity (%) | 0–20 | 45.79 ± 0.60 b | 50.90 ± 2.45 a | 49.58 ± 1.49 a |
| 20–40 | 39.07 ± 1.82 b | 48.03 ± 3.08 a | 45.34 ± 2.96 a | |
| 40–60 | 39.68 ± 2.50 a | 46.65 ± 5.81 a | 43.31 ± 1.56 a | |
| 60–80 | 43.70 ± 1.00 a | 46.65 ± 3.41 a | 44.62 ± 0.27 a | |
| 80–100 | 40.43 ± 2.44 b | 46.82 ± 1.02 a | 45.24 ± 1.67 a | |
| Root biomass (g·m−2) | 0–20 | 128.60 ± 54.43 b | 136.17 ± 13.62 b | 325.72 ± 58.58 a |
| 20–40 | 103.37 ± 17.94 a | 157.39 ± 25.42 a | 152.31 ± 44.20 a | |
| 40–60 | 119.09 ± 27.26 a | 96.08 ± 51.56 a | 96.19 ± 4.79 a | |
| 60–80 | 111.08 ± 37.92 a | 90.07 ± 48.65 a | 133.11 ± 67.48 a | |
| 80–100 | 29.74 ± 19.45 b | 30.93 ± 3.76 b | 110.78 ± 9.81 a | |
| Total | 491.87 ± 53.87 b | 510.63 ± 128.25 b | 818.11 ± 124.80 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Zhang, B.; Xu, Q.; Gao, D.; Xu, W.; Ren, R.; Jiang, J.; Wang, S. The Effects of Plant and Soil Characteristics on Partitioning Different Rainfalls to Soil in a Subtropical Chinese Fir Forest Ecosystem. Forests 2022, 13, 123. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010123
Zhang Y, Zhang B, Xu Q, Gao D, Xu W, Ren R, Jiang J, Wang S. The Effects of Plant and Soil Characteristics on Partitioning Different Rainfalls to Soil in a Subtropical Chinese Fir Forest Ecosystem. Forests. 2022; 13(1):123. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010123
Chicago/Turabian StyleZhang, Ying, Beibei Zhang, Qing Xu, Deqiang Gao, Wenbin Xu, Ranran Ren, Jing Jiang, and Silong Wang. 2022. "The Effects of Plant and Soil Characteristics on Partitioning Different Rainfalls to Soil in a Subtropical Chinese Fir Forest Ecosystem" Forests 13, no. 1: 123. https://0-doi-org.brum.beds.ac.uk/10.3390/f13010123