1. Introduction
With the expansion of the pure forest planting area and the increase in the number of rotations used, the continuous decline of soil fertility and the productivity of plantations has attracted national attention [
1,
2,
3,
4]. Ecosystem function depends largely on the functional diversity and activity of the underground microbial system [
5]. Soil microbial functional diversity is extremely sensitive to changes in the soil microenvironment [
6] and so can reflect changes in soil quality [
7,
8]. It is often used together with soil microbial biomass as an important indicator for evaluating soil fertility and health [
9,
10]. Deforestation and land management influence the function of soil microorganisms [
11,
12,
13], with some effects lasting for decades [
14], and have nonnegligible impacts on forest health and the long-term maintenance of productivity [
15]. Though the optimization of forest cutting and regeneration methodologies is necessary to improve the functional diversity of soil microorganisms, the effects of harvest residual treatment on the functional diversity of soil microorganisms remain unclear.
Traditional slash-and-burn cultivation is a common method of forest cutting and regeneration in plantations, but often has negative effects on the physicochemical and biological characteristics of soil [
2,
16]. Studies have shown that 93% of the total biomass is lost as gases and fly ash after burning, as well as nitrogen (N), phosphorus (P), and potassium (K), including losses of nutrients from dead plants [
17]. In addition, runoff occurs easily and causes substantial soil erosion due to the loss of the surface protective layer [
18]. In general, slash-and-burn cultivation is generally believed to change the structure of the soil food web, destroy soil organic matter [
19], reduce soil total nutrient contents [
17], and alter the soil microenvironment [
20,
21], thus disrupting the structure and affecting the functional diversity of soil microorganisms [
16,
22,
23].
Compared with traditional slash-and-burn cultivation, the functional diversity of soil microorganisms is higher when there is little or no interference [
22,
24]. Moreover, the negative impact of wood and biofuel-oriented forestry practices on forest ecosystems can also be alleviated by mulching residues after harvesting [
25,
26,
27]. However, some studies have indicated that the composition of animals and microorganisms needed for decomposing coniferous forest residues in woodland is single, resulting in the easy accumulation of residues, which leads to a low rate of nutrient cycling and thus a decline in soil fertility [
28,
29]. It is even possible to remove harvest residues while maintaining productivity [
30]. Although there is no direct evidence that whole-tree harvesting reduces woodland productivity, mulching residues in woodland can reduce at least some of the direct nutrient losses of ecosystems [
31]. One study showed that the rate of removal of N, P, K, calcium (Ca), and magnesium (Mg) with whole-tree harvesting increased by 216%, 304%, 152%, 254%, and 151%, respectively, compared to stem-only harvesting, and the removals and displacements of nutrients were unacceptably large [
32]. Therefore, the appropriate removal and effective utilization of harvest residues is likely to affect the functional diversity of soil microorganisms and ultimately affect the forest productivity, especially in plantations with short-term rotations.
Mulching with crushing straw is widely used in agriculture, as it facilitates nutrient release, lessens nutrient loss, and reduces human disturbance to the ecosystem by avoiding the use of fire [
33,
34,
35,
36]. However, there have been few investigations on forestry management in the hilly areas of South China, and whether crushing residues can accelerate the nutrient transfer process, improve soil quality, enhance soil microbial functional diversity, and maintain a long-term high yield in the forest remains unclear.
Chinese fir (
Cunninghamia lanceolata (Lamb.) Hook.) is a representative fast-growing tree species of plantations in the subtropical area of China [
37], with an accumulative planting area of over 12 million hectares [
38]. It has been used for large-scale afforestation for at least 1000 years and plays an important role in the construction of the timber forest base in southern China [
39]. With the increase in the planting area of pure Chinese fir forest, the number of rotations, soil fertility, and plant productivity have significantly reduced, which has affected the construction of timber forest sites in this region [
40]. Soil microbial change is the key to aboveground and underground ecological function [
5]. In order to ensure the sustainability of high-productivity Chinese fir plantations, we hypothesized, based on the important role of harvest residues in nutrient cycling and soil fertility conservation in plantations, that the management of harvest residues could enhance the functional diversity of soil microorganisms. Our aim is to study the effects of three harvest residual treatments in Chinese fir plantations (RB, residual burning; MT, crushing and mulching treatment; and NR, no residuals) on the functional diversity of soil microorganisms and their changes over time, as well as to explore the response mechanism of soil microbial functional diversity to the three types of residual management in two years (2018–2020). This will provide scientific evidence and a technological basis for solving the problem of soil fertility decline in Chinese fir plantations.
4. Discussion
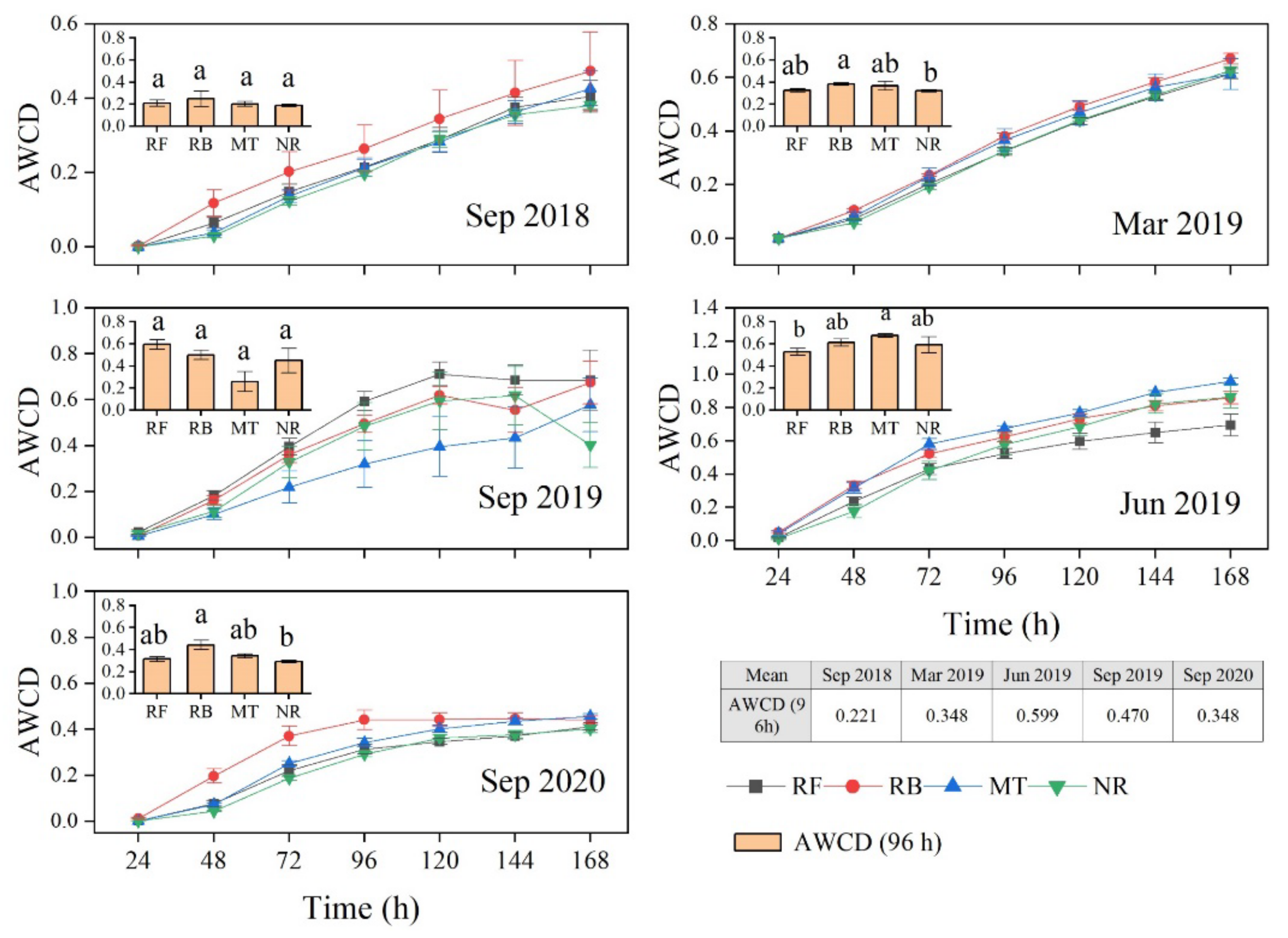
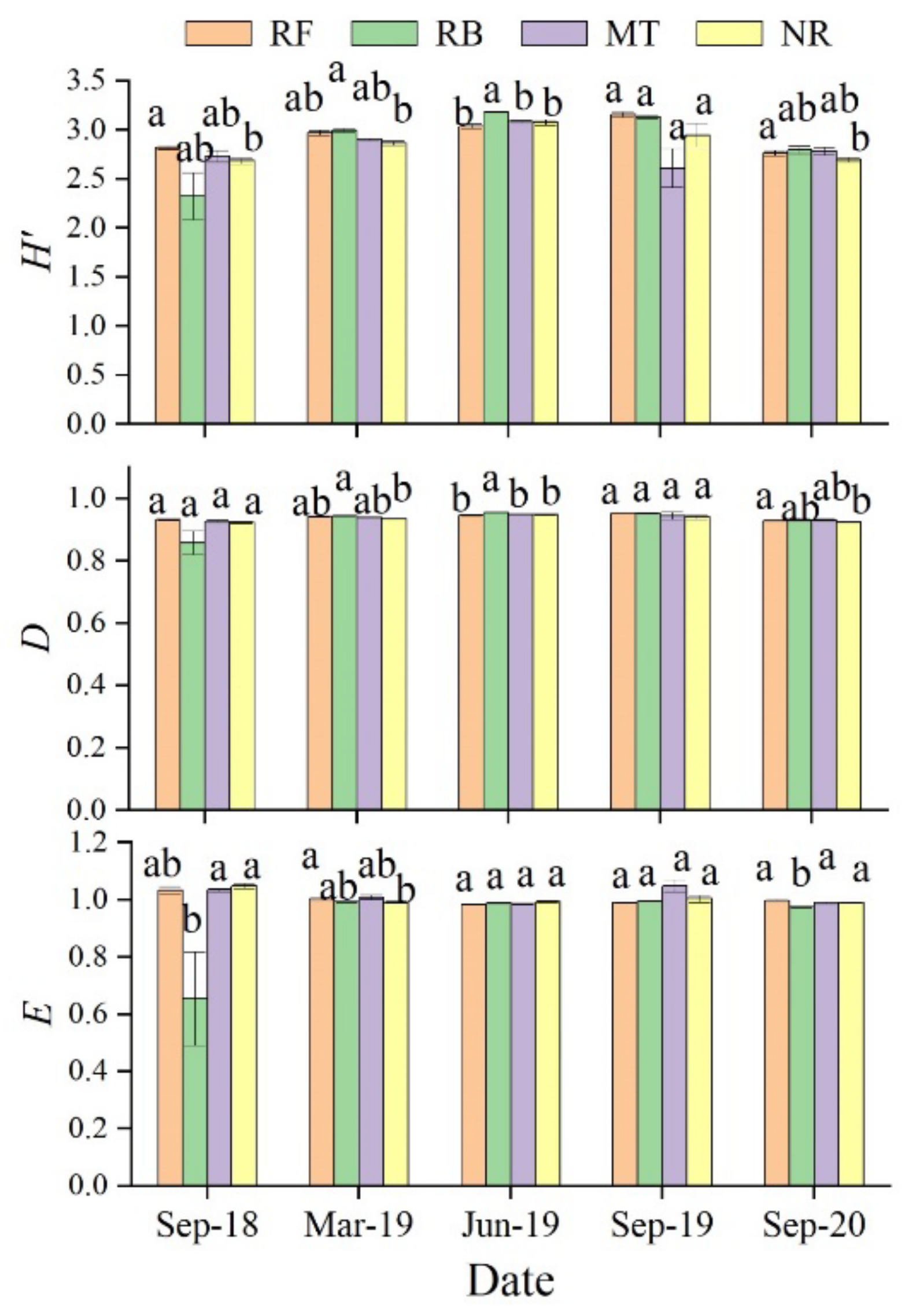
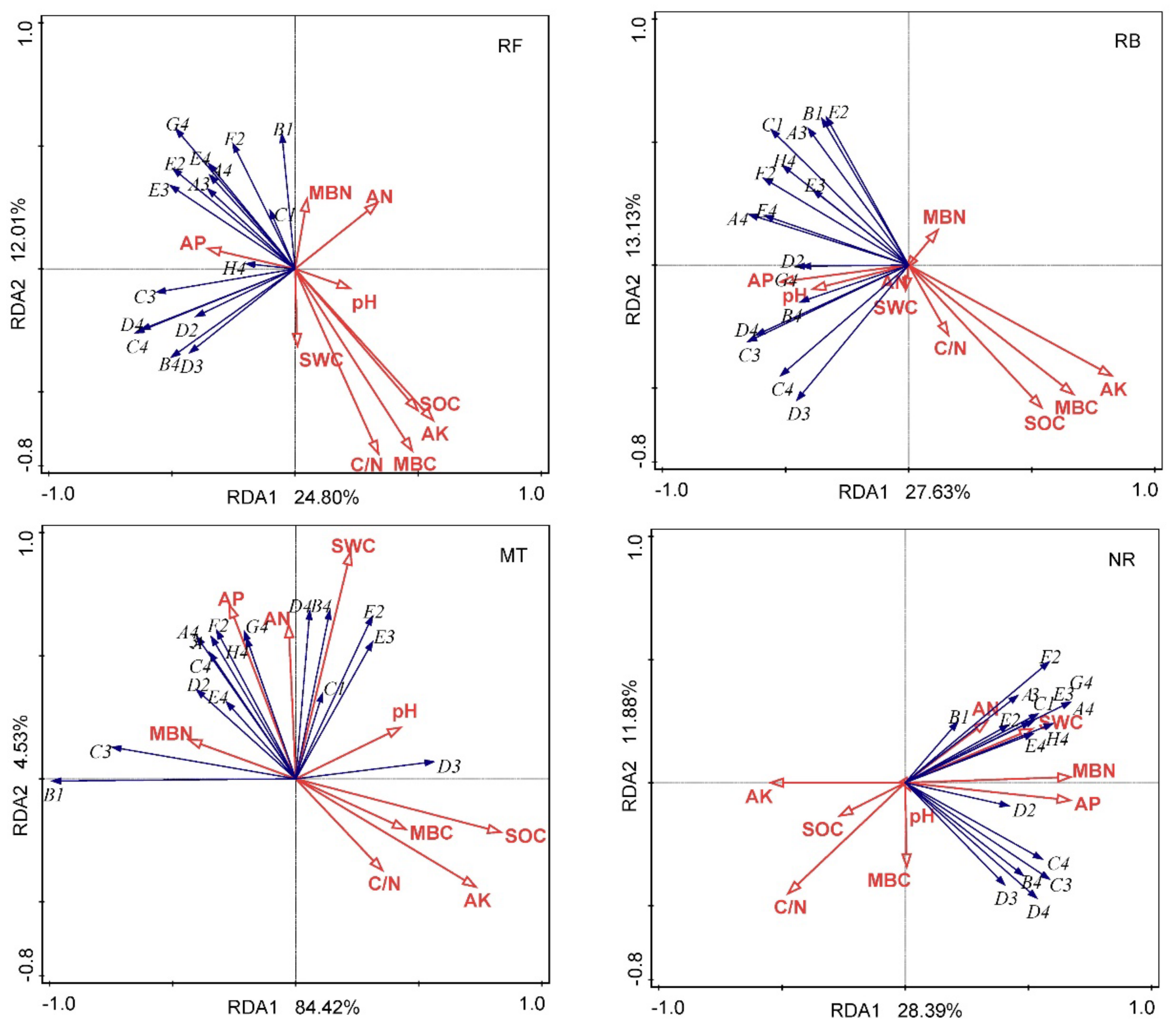
Based on two years of observation, we studied the characteristics of the variation of the functional diversity of soil microorganisms under different cutting and regeneration methodologies in a Chinese fir plantation and analyzed the driving factors in order to understand the mode of response of the soil microorganisms to plantation management. Our study showed that the modes of response of soil microbial functional diversity in a forest ecosystem to various cutting and regeneration methodologies varied, and that they were also affected by soil physical and chemical properties, as well as the sampling period.
Forest management can affect the metabolic activity of the soil microbial community by changing the soil physical and chemical properties [
50,
51,
52]. In our study, the AWCD values (
Figure 3 and
Figure 4) and diversity index (
Figure 5) were combined to show that the soil microbes in the burned area had a higher metabolic activity, including dominance and richness, than those in the unburned area, but that the community evenness was poor. Rosaria et al. (2005) [
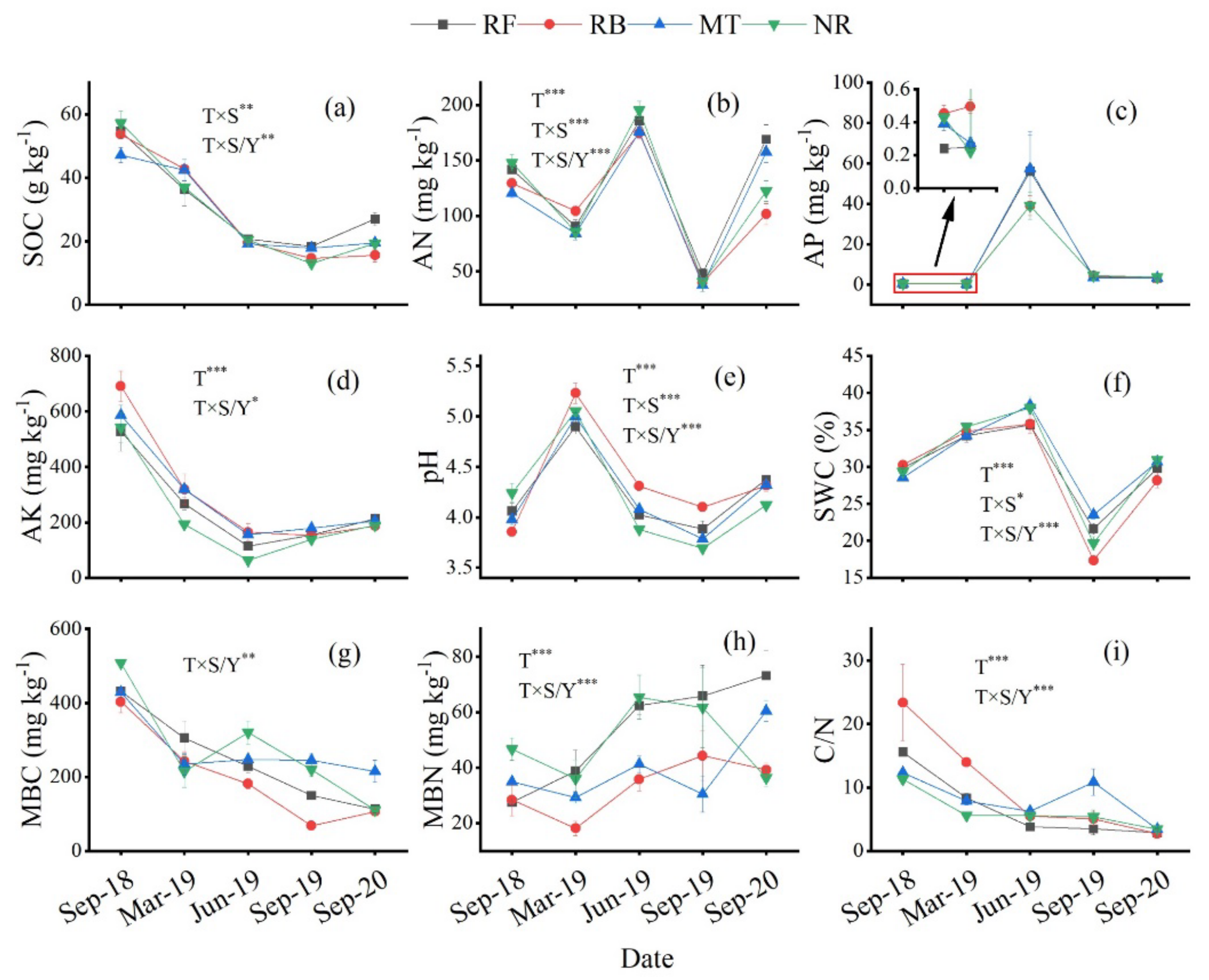
53] reported that in the first week after a fire disturbance, the carbon source substrate used by soil microorganisms changed slightly and transiently, but the microbial community quickly restored its function and diversity. They also showed that, compared to unburning, fire disturbance increases the functional diversity of soil microorganisms. The nutrients in organic matter are returned to the soil in a relatively short time after residual burning [
54,
55]. We found that, especially at the early stage, the content of available nutrients such as AP and AN was higher (
Figure 2), was positively correlated with the metabolic activity of the soil microbial carbon source, and could effectively improve the metabolic activity of the soil microbial community. However, the comparison of soil samples collected 5 years after burning and unburned soil samples demonstrated that burning significantly reduced the microbial functional diversity [
56], suggesting that severe forest disturbances, such as significant burning, should be avoided in order to reduce their effect on the soil microbial community. This may because soil nutrients are only increased for a short period of time after slash-and-burn cultivation and become substantially reduced after many years of using this technique, especially in the rainy, hilly regions of the south of China [
57]. A large number of nutrients are directly lost in the form of gas or dust in the instant of slash burning, and the ash left after burning is easily lost by erosion caused by rain in rainy seasons [
17]. Unexpectedly, no significant decrease in the metabolic capacity of soil microorganisms in the burning area was observed during our 2-year study, and some still showed a significant increase after two years of treatment (
Figure 3), although the soil nutrients in the soil given this treatment had begun to decrease (
Figure 2), which may have been caused by the time lag between microbial response and substrate reduction or other factors not observed in this study. However, the community evenness in soil given the burning treatment was always poor (
Figure 5). Slash burning can kill and weaken certain plants, animals, and microorganisms, creating an ecological niche [
58,
59,
60]. An adequate supply of nutrients would help microorganisms to perform all their functions, encourage some microorganisms to compete successfully and come to occupy a dominant position, and thus improve the richness and dominance of the soil microbial community [
21,
61,
62]; however, it would also destroy the microbial community structure and lead to the deterioration of community evenness.
The soil microbial community structure and diversity have been found to be related to the variation in soil nutrients caused by organic mulch [
63,
64]. Studies have shown that the soil microbial functional diversity in Chinese fir plantations was significantly affected by silvicultural treatments, which was related to the microclimate and soil substrate input [
65,
66]. Chinese fir could supply a huge amount of litterfall, which was found to be significantly correlated with the input of soil C, N, etc. [
67,
68]. Therefore, litter may be an important factor supporting soil microorganisms [
69]. Once harvest residues reach the ground, they can also be treated as litterfall to an extent.
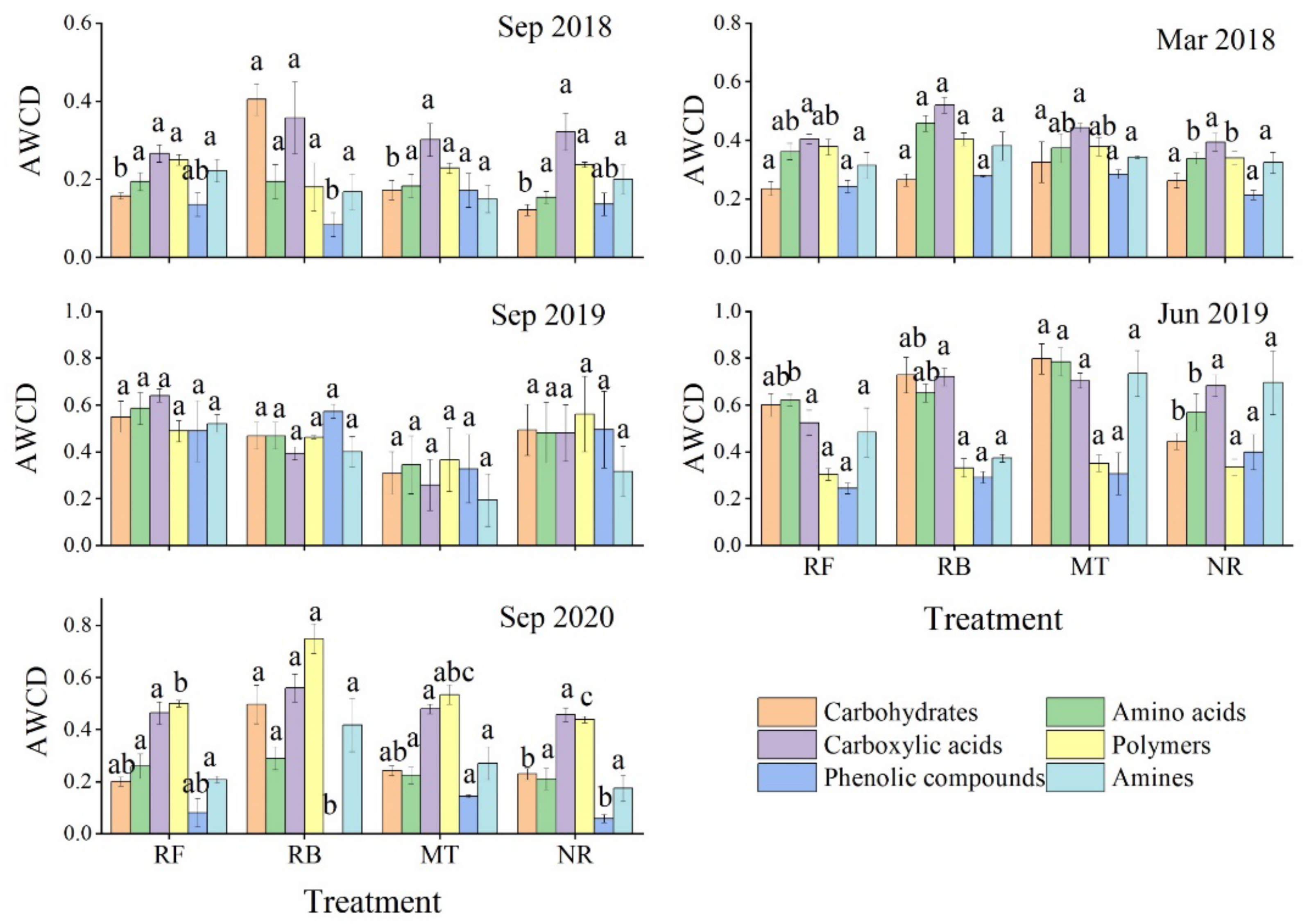
In our study, the soil given the crushing and mulching treatment had a higher metabolic capacity for microbial carbon sources in summer, especially for amino acids and carbohydrates, which are two major carbon sources used by microorganisms (
Figure 3 and
Figure 4). Huang et al. (2008) argued that plant residues such as mulch significantly increased the diversity of soil microbial functional communities in Australian subtropical plantations compared with communities given no mulch [
70]. When topsoil is mulching, the evaporation rate will be reduced, higher water contents and smaller soil temperature changes will be maintained for a longer period of time [
71,
72,
73], soil loss will be effectively controlled [
74,
75], and more suitable habitats can be provided for microbial metabolism activities. This can also increase the substrate required for microbial metabolic activities through the decomposition and leaching of organic matter [
76,
77], and even help to control weed growth [
78,
79] and reduce the loss of nutrients [
80]. The addition of organic matter can also increase the microbial biomass, change the composition of the microbial community, and lead it to develop a higher abundance and activity, thus enhancing the functional diversity of microorganisms [
81]. Compared with untreated soil, mulching crushed residues can cause the non-synchronous decomposition of organic matter, providing different habitats and nutrients for microorganisms and thus changed the metabolic mechanism of microorganisms. As the host of microorganisms, fresh organic matter, after being subjected to crushing, not only greatly increased the area available for microbial lodging but also increased the contact area of organic matter with air, water, and sunlight and accelerated the decomposition rate of organic matter, especially in the hot and rainy summer. This provided a more favorable habitat and greater amount of substrate for microorganisms [
77] and in turn improved the carbon source metabolic activity of soil microorganisms.
A plot with no residues will have a reduced soil quality [
4], which may be the main reason for the low metabolic activity of microorganisms seen in this treatment. Large amounts of organic matter were removed from the ecosystem in this treatment, leading to the direct loss of large amounts of nutrients [
4,
82]. Although the SOC was higher in the initial stage of treatment, this was only due to the rapid decomposition of the surface humus layer due to the exposure of the surface and direct sunlight [
83]. At a later stage, having no residues to reduce the nutrients, SWC, and pH affected the functional diversity of soil microorganisms [
82,
83,
84,
85].
Overall, the use of burning and crushing treatments showed certain advantages for the functional diversity of soil microorganisms. However, the microbial biomass seen in the burning treatment was lower throughout the whole observation process, which may have had a profound impact on the sustainability of the soil ecosystem and had no sustained effect on promoting the supply of soil nutrients [
21]. Moreover, it has been proven that burning can only increase the functional diversity of microorganisms for a short period [
56]. Therefore, we prefer the use of the crushing treatment, which may be applied for the future management of Chinese fir plantations. Of course, longer-term data are needed to verify the effects of this process.


 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





