1. Introduction
The European spruce bark beetle,
Ips typographus (L.) (Coleoptera: Curculionidae: Scolytinae), is the most important biotic disturbance agent of Norway spruce,
Picea abies (L.) Karst. (Pinales: Pinaceae). Its preferred breeding substrates are wind-thrown and drought-stressed trees with impaired defenses [
1,
2]. Over the last several decades, the high abundance of predisposed spruce stands, as well as an increasingly warmer and drier climate, have promoted the transition from endemic to epidemic population densities of
I. typographus in many regions of Eurasia [
3,
4,
5,
6]. Under outbreak conditions,
I. typographus can colonize vigorous trees due to mass attacks mediated by aggregation pheromones [
7]. At low attack rates, however, successful brood establishment depends on the landing of male pioneer beetles on suitable, weakened hosts, where tree defenses can easily be overcome. Thus, the availability and recognition of highly susceptible hosts in the initial attack phase strongly influence the extent of population build-up. Bark beetles may randomly encounter trees during long-range dispersal, but at short distances, they likely make a deliberate choice guided by visual, olfactory, gustatory, and thigmotactic (haptic) cues [
8]. To efficiently detect suitable breeding sites,
I. typographus antennae have highly specific olfactory sensilla for identifying odor signals, composed of volatile organic compounds emitted from host and non-host trees, such as terpenoids or green-leaf alcohols [
9,
10]. Moreover, the European spruce bark beetle’s olfactory system is sensitive to oxygenated host volatiles related to tree stress, which can serve as repellent or attractive cues for host selection [
11].
The odorant receptors of
I. typographus do not only respond to tree odors and pheromone components, but also to specific oxygenated monoterpenes and fusel alcohols produced by its fungal symbionts [
12,
13]. Ophiostomatoid (blue-stain fungal) communities of
I. typographus involve different species varying in virulence towards Norway spruce, with some of them helping beetles to overcome and exhaust host defenses and promote tree death [
14,
15,
16]. Unable to penetrate intact tree bark, these fungi depend on being vectored on the body’s surface and in the gut of dispersing beetles, subsequently being introduced into breeding galleries in the phloem tissue [
17]. Among the most frequently found and potentially most virulent fungal associates,
Endoconidiophora polonica (Siemaszko) Z.W. de Beer, T.A. Duong & M.J. Wingf. and
Grosmannia penicillata (Grosmann) Goid. cause the tree to locally establish hypersensitive wound reaction zones around infection sites, enclosing areas of necrotic phloem, where terpenes and phenolics are accumulated [
18,
19]. The experimental inoculation of blue-stain fungi is an established method to test for tree resistance towards subcortical fungal and insect attacks [
20,
21]. Larger wound reaction zones indicate a slower containment of fungal growth due to the impaired defense reactions of stressed trees [
22]. Moreover, in field experiments with young Norway spruce, increased terpene induction in response to fungal inoculation strongly inhibited subsequent spruce bark beetle colonization [
23]. Yet, experimental data that make connections between drought stress, susceptibility to ophiostomatoid fungi, and the attractiveness of Norway spruce for
I. typographus males searching for hosts are still lacking.
Studies on the preference of bark beetles for trees of various stress and defense states under field conditions are hampered by seasonal weather conditions, outbreak dynamics, and the problem that susceptible trees can be detected only after natural infestation [
24]. Experimental approaches to control tree attacks, such as the use of small plastic tubes to expose beetles to the bark, have the disadvantage that the beetles’ choices are confined to host acceptance or no host acceptance [
25]. In field bioassays with attack boxes, which allow beetles to move freely, explore a larger bark area, or exit the system, drought-stressed trees were more successfully attacked [
1]. While empirical evidence for the increased attractiveness of drought-stressed trees as hosts for pioneer beetles is still scarce, choice experiments using Y-tube olfactometers proved that mass aggregation in
I. typographus is elicited by aggregation pheromones released from unmated males [
26]. In similarly designed bioassays, the repellent effects of trans-4-thujanol were observed, showing the increased susceptibility of older spruce trees to beetle attacks due to a natural decrease in concentrations of this oxygenated monoterpene [
27]. Choice experiments merely based on olfaction may, however, only partially reflect the natural behavior of insect herbivores [
28]. A relatively simple method to also include gustatory and thigmotactic cues are Petri dish arena choice experiments. For instance, Kandasamy et al. [
29] used such arenas to show that European spruce bark beetles preferentially chose and tunneled in spruce bark agar colonized by
E. polonica,
G. penicillata, and other ophiostomatoid fungal species. Beetles even preferred agar sections containing fungal associates of the North American spruce beetle,
Dendroctonus rufipennis (Kirby), over the pure spruce agar medium [
30], which suggests a high ecological relevance of these symbionts for bark beetle brood establishment.
Here, we performed Petri dish arena choice tests to study the preferences of male I. typographus for bark cores sampled from trees of various levels of drought stress and susceptibility to an ophiostomatoid fungus. The examined trees were part of a rainfall exclusion field study including two treatments, rain-out shelters (roofed trees) and non-roofed trees. Drought stress of trees was assessed by repeated measurements of twig water potentials in the field and their defense states by the extent of hypersensitive wound reactions in response to inoculations with G. penicillata. We addressed (1) whether male I. typographus preferred the bark cores of either drought-stressed (roofed) or well-water-supplied trees (non-roofed) over those of randomly chosen non-manipulated control trees at the study site; (2) if the time beetles remained in Petri dish sections was related to their final choice; (3) whether the extent of the drought stress of study trees increased attractiveness of sampled bark cores to I. typographus; and (4) if tree defense status, as indicated by the size of hypersensitive wound reaction zones in response to inoculation with G. penicillata, influenced a beetle’s final choice of a certain bark sample. Our results contribute important data for understanding I. typographus host colonization behavior and the beetles’ potential to successfully attack and breed in Norway spruce.
2. Materials and Methods
To understand the host selection behavior of male
I. typographus towards Norway spruce trees differing in drought-stress status and resistance, we conducted Petri dish arena choice experiments. Trees for our experimental trials were selected from the Rosalia Roof Study II, a rainfall exclusion study conducted in the Austrian Rosalia Mountains, forming the eastern foothills of the Alps [
1,
22,
31]. The main project goals of the Rosalia study were to examine the influence of site water supply on Norway spruce biochemistry, physiology and chemical defense, the subsequent effects on tree attractiveness and susceptibility to attacks by the European spruce bark beetle,
I. typographus, and susceptibility to its associated ophiostomatoid fungi. The experimental plots were located in an 80-year-old Norway spruce stand interspersed with Scots pine,
Pinus sylvestris (L.), and European beech,
Fagus sylvatica (L.). Climatic and other site-related characteristics of the study area are described in Netherer et al. [
1].
2.1. Field Experiments and Sampling of Bark Cores
The drought manipulation treatment site was installed between October and December 2018 and involved ten roofed (R) trees excluded from direct precipitation by 8 × 8 m sized rain-out shelters and ten non-roofed (nR) trees (
Figure 1). These 20 study trees were monitored for various tree physiological and defense-related parameters in 2019 and 2020 (results not shown) and sampled for bark cores used in the Petri dish arena choice experiments. In addition, two non-manipulated and non-monitored control trees (C1, C2) in close vicinity to the study trees were sampled for the experiments. These control trees were of a similar age and diameter, faced the same site and water supply conditions, and showed similar vigorous crown conditions.
To evaluate the water supply of R and nR trees, the pre-dawn twig water potential, which is considered a reliable tree physiological indicator of water stress in rainless periods [
32,
33], was measured repeatedly over the study season of 2020 (20 May, 1 July, and 5 August), following the pressure chamber method described in Netherer et al. [
1].
To assess the defense response (hypersensitive wound reaction) in the bark of R and nR trees towards blue-stain fungal infection, the study trees were inoculated with the ophiostomatoid fungal species
G. penicillata. This ascomycete is a frequent and dominant symbiont of
I. typographus and considered to accelerate the exhaustion of tree defense and tree death during bark beetle attacks [
8,
18,
34]. The isolate of
G. penicillata (strain LF/Ia/9/1, stored in the fungal culture collection of IFFF-BOKU) was obtained in July 2020 from desiccated sapwood of an
I. typographus-attacked Norway spruce trap tree nearby the study site and grown for 13 days on ash leaf malt extract agar (AMEA, [
35]) in plastic Petri dishes (diameter 5.2 cm). On 6 August 2020, the ten R and ten nR trees were wound-inoculated with the fungus, following the methods described in detail by Netherer et al. [
22]. Each tree received six inoculations of
G. penicillata, which were evenly distributed around the trunk. Three fungal inoculations were arranged around the trunk at lower stem heights of 160 cm (R) or 210 cm (nR) and three at upper stem heights of 210 cm (R) or 260 cm (nR). The control trees C1 and C2 remained untreated. Areas in mm
2 of hypersensitive wound reaction zones (lesion sizes) were recorded six weeks after inoculation, on 16 September 2020. Short/small lesions indicated increased tree defense ability due to a fast containment of fungal growth in the affected bark sections [
36]. The measurements of lesion size per tree were averaged and mean lesion size per tree was used as a parameter to characterize the defense states of the R and nR trees in the data analysis (see
Section 2.3).
Between 20 and 40 bark cores per tree with a diameter of 1 cm were sampled with a cork borer from seven R (R1, R2, R3, R4, R5, R6, R9) and eight nR (nR1, nR3, nR4, nR5, nR7, nR8, nR9, nR10) trees and from the two additional control trees (C1, C2) (
Figure 1) at breast height on 16 September 2020. Samples included outer bark and vigorous phloem unaffected by fungal infection or other damage. All cores were shock-frozen with liquid nitrogen in the field and preserved at −80 °C until experiments were performed. Shortly before Petri dish arena trials (starting in October 2020) the stored bark cores were transferred to −20 °C to facilitate an easier handling of samples.
2.2. Petri Dish Arena Choice Experiments
The laboratory trials were mainly run from October 2020 to February 2021. Additional Petri dish arena choice experiments to study possible differences in the attractiveness of C1 and C2 and between particular R and nR bark cores, were conducted in August 2021 (
Table 1). Test insects were obtained from our permanent bark beetle rearing facility at the institute.
Ips typographus individuals that had recently completed their maturation feeding and emerged from logs were collected from rearing cages. The beetles were sexed as described by Schlyter and Cederholm [
37], and only male beetles were used in the experiments.
All choice experiments were performed in plastic Petri dishes (14 cm diameter), covered with white filter paper to facilitate walking of beetles. Petri dishes were divided into equally sized T (test, mainly for bark samples from R or nR trees) and C (control, mainly for bark samples from C1 or C2 trees) sections, each equipped with a white PET bottle cap with four small (5 mm diameter) entrance holes (
Figure 2). Before the start of a test run, frozen bark cores were put in the caps (for details on tested pairs, see
Table 1) and thawed for ten minutes at room temperature. Afterwards, one
I. typographus male was placed in the center of the Petri dish, which was subsequently closed with a transparent lid. Bark beetle activity/behavior was monitored within a maximum observation period of 60 min.
Test beetles could move freely in the Petri dishes and enter the caps with bark cores through entrance holes and from above. Time was recorded when a beetle changed between T and C sections and when it made a distinct, final choice for the T or C bark core. The choice was considered as definite (“T” or “C” choice), and the test run was stopped, once the beetle entered a cap and remained with the sample for at least two minutes (
Video S1). In case a beetle was not within a cap after a maximum of 60 min, the outcome of the trial was categorized as “no choice”. Beetles showing no movement for at least ten minutes anytime during the test run were categorized as “inactive”, replaced with a new male, and the trial was started all over again. Fresh bark samples were used for each test run to provide each beetle with an odor blend of freshly thawed bark cores. Individual bark beetles were tested only once.
Choice tests performed until February 2021 between “T and C” (i.e., “R/nR and C1/C2”) or “T and T” (i.e., “R/nR and R/nR”) involved 10 or 20 test runs (i.e., replicates) per experiment (i.e., compared tree pairs) (
Table 1). Additional experiments performed in August 2021 (“C1 and C2” and “R9 and nR9”) involved 29 and 23 test runs, respectively. To evaluate the ability of bark beetles from the permanent rearing to perceive olfactory stimuli and to track odor sources in the Petri dish arena, ten “zero tests” were conducted by placing a C2 bark core into cap C, while cap T stayed empty.
2.3. Data Analysis
Behavioral observations were analyzed by calculating the total numbers and proportions (%) of “T”, “C” and “no” choices across all test runs conducted within a particular choice experiment (same pair of test and control trees). Test runs that had been terminated due to the inactivity of beetles were not considered in the final analysis. Furthermore, the absolute and relative time, i.e., the percental share of time a beetle spent in each section (mean % time ± standard deviation) for sections T and C across all test runs (i.e., replicates) per experiment was calculated. Similarly, the mean absolute time ± standard deviation (in sec) until beetles made an ultimate choice for a bark sample (no matter whether T or C) for each experiment was evaluated. Average times (± standard deviation) until a choice for a bark core was made were also calculated for all test runs ending with a T and a C choice, separately. Independent samples t-tests were performed to evaluate differences in mean absolute and relative time spent in T and C sections, and the average time until a final choice was made between the experiments ending with a T and those with a C choice. The normal distribution of data was controlled by a Shapiro–Wilk test and homogeneity of variances by a Levene’s test. We used Fisher’s exact test to examine whether the number of T and C choices significantly deviated from an expected equal distribution. Relationships between the beetles’ final choices for T or C (% T and % C choices), relative time spent in sections T and C, drought-stress states of study trees (pre-dawn twig water potentials recorded in the field in May, July, and August 2020), and defense states of trees (mean lesion size following inoculation with G. penicillata) were examined by Pearson correlation r (p ≤ 0.05 significant correlation; p ≤ 0.001 highly significant correlation). Figures were prepared and analyses were performed in Excel © Microsoft 365 and IBM SPSS Statistics 22.
3. Results
The behavior of
I. typographus males, which could decide between bark cores of differentially stressed Norway spruce (R, nR) and two untreated control trees (C1, C2), was studied in a total of 292 independent test runs of Petri dish arena choice experiments (
Table 1). Overall, seven trials (i.e., 2%) had to be terminated without a final choice for test or control samples due to inactivity of beetles. In more than 90% of the tests, beetles made a final choice, which means they entered a cap either containing the T or the C sample and stayed there for at least two minutes (
Table 1,
Figure 3). Bark cores of the study trees R1, R6, and R9 were only tested against C1, the cores of R2, R5, nR4, nR5, and nR10 were only tested against C2, while those of R3, R4, nR1, nR3, nR7, nR8, and nR9 were tested against both C1 and C2 samples (
Figure 1,
Table 1).
In “zero tests” (i.e., experiments with one empty cap to evaluate whether beetles responded to volatiles in this setup), all beetles made a clear choice for the bark sample (
Table 1,
Table S1,
Figure 3f). Zero tests and additional “T and T” and “C and C” comparisons accounted for 92 test runs. The two untreated control trees were similarly attractive for the test beetles when they could choose between them, i.e., in a “C and C” experimental set-up (43% C1, 50% C2, 7% no choice) (
Figure 3e).
3.1. Beetles Preferred Bark Samples of Roofed Trees over C1, but Both Controls over Non-Roofed Trees
Male
I. typographus showed a slight preference for bark samples of roofed study trees (specifically R1, R3, R6, and R9) over control samples (mainly C1), and clearly chose C1 and C2 cores over non-roofed tree samples (except for nR1 and nR8) (
Table 1;
Figure 3a–d).
The nR trees were affected in the same way by the measurements and bioassays performed in the Rosalia study as R trees but supposedly were similarly well-supplied with water as the C trees. While 52% of test runs ended with the choice of R and 34% for C1 (Fisher’s exact test:
p = 0.033; Χ
2 = 5.12; df = 1), only 27% chose nR, but 65% chose C1 samples (
p = 0.049; Χ
2 = 4.41; df = 1). Compared to C2 cores, test beetles showed a similar preference for R and nR samples (41% and 39%, respectively), but only nR choices deviated from the expected 50% (
p = 0.018; Χ
2 = 6.025; df = 1). The proportion of beetles making no choice for any bark core was low in all experiments (6–14%). Additional “T and T” tests to directly compare study trees partly confirmed the preferences of beetles for specific R and nR samples observed in the “T and C” tests. Bark cores of R3 (preferred over C1 and C2) were also preferred over R2 and nR10 (less often chosen than C2), but R2 was preferred over nR8 (preferred over C2 but not C1). In further choice experiments conducted in August 2021, beetles preferred nR9 (less often chosen than C1 and C2) over R9 (preferred over C1) (
Table 1).
3.2. The Time That Beetles Spent in T and C Petri Dish Sections Was Related to Their Final Choices
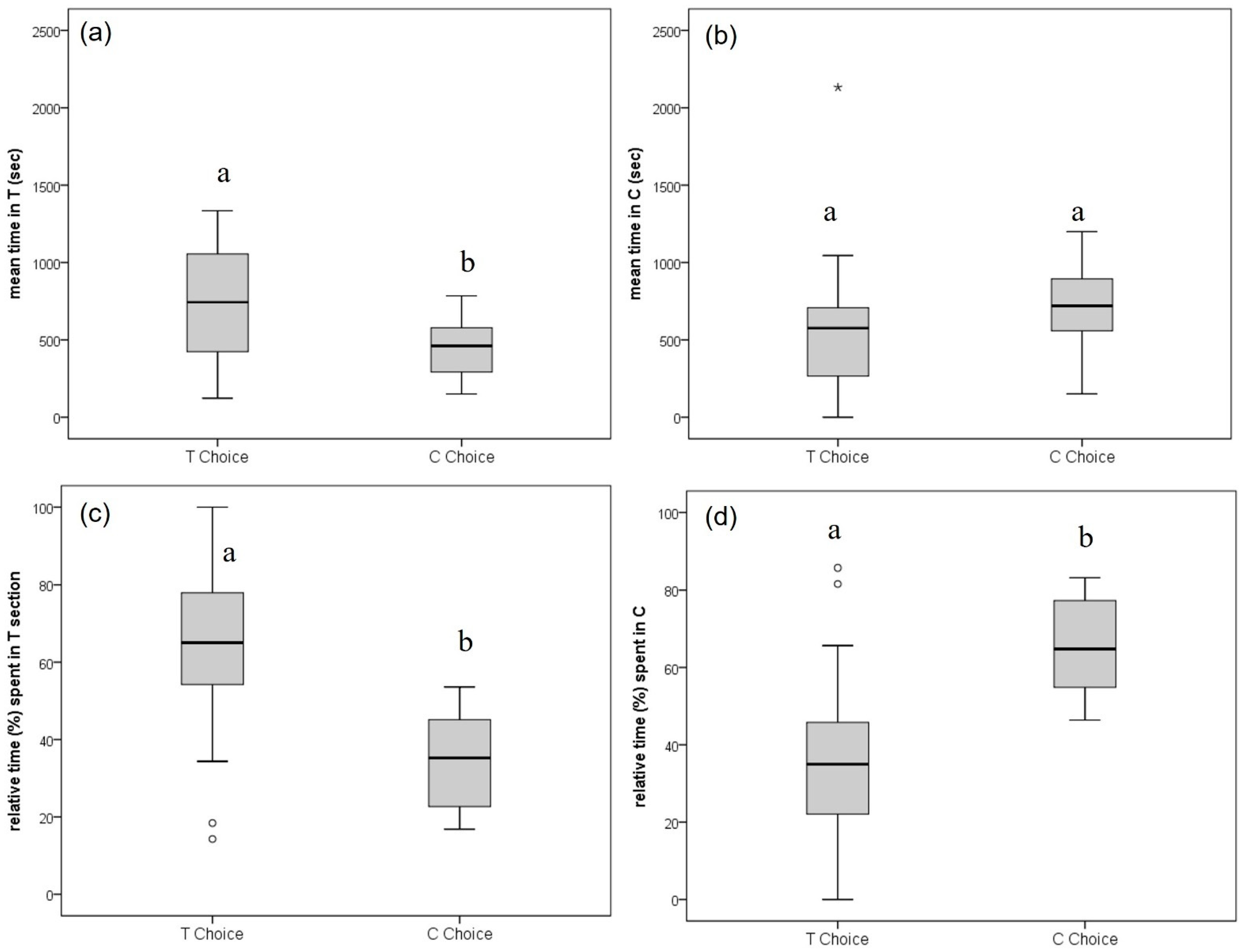
After placing test beetles in the center, they actively explored the arena and moved between T and C sections, both on the floor and the lid of Petri dishes. We commonly observed that beetles changed sections multiple times, attempted to fly, or entered a cap and stayed only briefly (less than two minutes) with a particular bark core before they made a definite choice, or until the test run ended after 60 min. The time taken until the beetles finally decided on a bark sample ranged from a few seconds to a major part of the maximum test time (
Table S1) but did not differ between an ultimate T (1205 ± 631 s) and C choice (1102 ± 370 s) (T = 0.66; df = 42;
p = 0.52). However, the mean time spent in area T was significantly higher when beetles finally decided for T (750 ± 350 s compared to 448 ± 179 s in area T when beetles decided for C; T = 3.60; df = 42;
p ≤ 0.001) (
Figure 4a). While the differences were not significant for mean times spent in area C (
p = 0.147;
Figure 4b), they were highly significant regarding the relative time spent in the T and C sections depending on the final choice (mean % time in T: 63 ± 21 for T and 34 ± 11 for C choices; mean % time in C: 37 ± 21 for T and 66 ± 11 for C choices; T = 5.52; df = 42;
p ≤ 0.001) (
Figure 4c,d).
The mean time test beetles stayed in the section of tree samples corresponded with their choices for or against a particular tree. This means that the proportions of the final T and C choices observed across all test runs of an experiment and relative duration of stay (mean % time) in Petri dish sections T and C, respectively, showed clear positive linear relationships (
Figure 5a,b). To provide some examples, beetles spent more than 50% of their time in section C when they preferred the C sample, which was the case for nR3 (70% C1, 55% C2 choice), nR9 (80% C1, 70% C2) and nR10 (80% C2). Similarly, beetles spent more time in section T when they more often chose the T sample, as was the case for R6 (50% T, 30% C1) and R9 (80% T, 10% C1).
3.3. Drought Stress of Study Trees Increased Attractiveness of Bark Cores to Bark Beetles
Pre-dawn twig water potentials of R trees were lowest in May 2020, indicating minor drought stress due to the roof treatment, and increased (i.e., values were less negative) in July and August 2020 (
Table S2), indicating an even lower drought stress during summer 2020. The proportion of final decisions of test beetles for R or nR, against C1 or C2 bark cores, as well as the relative time spent in T sections increased with more negative pre-dawn twig water potentials of the test trees. In particular, a clear positive relationship between the final choice for the study tree (T) or the control tree (C1) and twig water potentials recorded in May 2020 was observed (
Figure 6a,b). Notably, all nR trees tested in these experiments showed twig water potentials above the value of −0.65 MPa and T choices below 40%, while R trees were more stressed, with water potentials between −0.78 and −0.93 MPa, and were chosen to 40–80% (
Table 1,
Tables S2 and S3). The proportions of T choices (against C1) and twig water potentials in July 2020 (in MPa
−1) also showed significant positive correlations (r = 0.69;
n = 10;
p = 0.028), showing a similar pattern as described above. T choices showed no significant relationship with August water potentials (r = 0.39;
n = 10;
p = 0.27 for “T and C1” tests) (
Table S3).
Although being rather scattered, a distinct linear relationship between the drought-stress status of trees and % T choices and relative times spent in T, respectively, was found in all “T and C” experiments (including C1 and C2). Beetles spent 41–68% of the time in the Petri dish section containing samples of R trees (compared to 22–54% of time with samples of nR trees), which were clearly more drought-stressed (twig water potentials as low as −1.13 MPa) than non-roofed trees in May 2020 (
Figure 6c). Despite some outliers of % T choices (such as R3 tested against C2 and nR8), the decisions of beetles were significantly correlated (r = 0.45;
n = 22;
p = 0.037) with pre-dawn twig water potentials measured in July 2020 (
Figure 6d), but again, not with values measured in August (r = 0.37;
n = 22;
p = 0.09). Nonetheless, the mean water potential values over the May, July, and August measurements, characterizing the water supply of trees for a longer period from spring to summer, were significantly correlated with relative times spent by bark beetles in section T (r = 0.57;
n = 22;
p = 0.005) (
Table S3).
3.4. Beetles More Often Chose Bark Cores of Trees with Larger Necrotic Lesions after Fungal Inoculation
Roofed trees inoculated with
G. penicillata showed a trend towards larger hypersensitive wound reaction zones six weeks after fungal infection (2845.1 ± 1407.6 mm
2), as compared to non-roofed trees (1903.5 ± 458.0 mm
2) (T = 1.795; df = 13;
p = 0.096). Areas of wound reaction were significantly correlated with the pre-dawn twig water potentials recorded in July (Pearson correlation r = 0.56;
n = 15;
p = 0.028) and in August (r = 0.72;
n = 15;
p = 0.002), indicating a decrease in tree defense ability with drought stress. However, lesion size was not related to twig water potentials in May (r = 0.19;
n = 15;
p = 0.50), when these measurements were lowest (
Table S2). Proportions of T choices were not correlated with lesion sizes when including the outliers R9 and nR8 (
Figure 7a). The results obtained from Petri dish arena choice experiments with R9 and nR8 were also slightly ambiguous (
Table 1). The bark cores of nR8 were highly preferred over C2 (70% T) but not over C1 (30% T). R9 samples were preferred over C1 (80% T) but not over nR9, which was less often chosen than C1/C2 (20/30% T). When these outliers were omitted, lesion size and the proportion of T choices were highly significantly correlated for remaining “T and C” observations (r = 0.65;
n = 19;
p = 0.003) (
Figure 7a). More specifically, the increasingly frequent selection of study trees with larger lesion size (and increased drought stress) was highly significant across all “T and C2” observations (r = 0.82;
n = 11;
p = 0.002) and this trend was also apparent for “T and C1” test results (r = 0.66;
n = 9;
p = 0.055) (
Table S3). Mean relative time in section T did not depend on lesion size (r = 0.229;
n = 22;
p = 0.306) across all 22 experiments (
Figure 7b;
Table S3).
4. Discussion
Rising temperatures and extreme weather events, such as drought periods, are major drivers of the recent, unprecedented mass outbreaks of
I. typographus in European Norway spruce forests [
38]. Warmer and drier climate leads to accelerated bark beetle development, an increased number of generations per year as well as stressed forest stands, promoting exponential beetle population growth and attack of standing live trees. High
I. typographus population densities facilitate mass attacks, even of moderately stressed hosts, mediated by the release of aggregation pheromones [
39]. In addition, volatiles emitted from trees are assumed to play a role in initial attacks by pioneer beetles [
8,
40]. To obtain a better understanding of the relevance of the drought stress of trees for short-range primary attraction, we performed Petri dish arena choice tests using bark cores from study trees of a rainfall exclusion field experiment (Rosalia Roof Study II). Our laboratory bioassays provided empirical evidence that
I. typographus males are attracted to bark and phloem of Norway spruce and specifically choose drought-stressed trees with weakened defenses.
At the forest stand level, swarming bark beetles commonly attack trees in close vicinity to their emergence sites [
41,
42]. Before accepting a new host for colonization and brood establishment, beetles likely land on several trees to examine substrate quality and defense state [
43]. According to the hypothesis of random landing, beetles do not determine tree suitability based on long-range host volatiles [
44]. In our Petri dish arena study, freshly emerged male beetles showed a clear short-range host search behavior, most likely mediated by host volatiles, as they exclusively preferred bark samples over empty bottle caps in the “zero tests”. We consider it unlikely that beetles were mainly reacting to wound-induced volatiles emitted from bark cores, as samples were immediately shock-frozen in the field to stop degradation processes in the tissues.
The Petri dish setup allowed beetles to enter and leave the bottle caps through entrance holes and from above, and thereby, explore the smell, flavor, and texture of the bark samples. We observed diverse behaviors of bark beetles in the Petri dish arenas, involving immediate decisions within a couple of seconds, switching back and forth between bark cores before choices, or the refusal of both samples. Yet, in more than 90% of test runs, the beetles made a final choice for one of the two offered bark cores. Thus, Petri dish arenas have the advantage that insects can move and explore the test environment extensively, as opposed to Y-shaped olfactometers, which restrict options for entering one of the tubes in the reaction to pure odor information, without the possibility of coming back to the starting point [
28]. In our Petri dish arena choice experiments, bark beetles generally remained longer in those Petri dish sections containing the sample that was finally selected. We conclude that the observed behavioral patterns of the beetles were clearly influenced by olfactory, gustatory, and tactile traits of the bark tissues, and that arena tests are a simple and reliable method to study the host choice behavior of
I. typographus. The developed test system may have further application for studying the behavior of other “aggressive” bark beetles, particularly those with episodic eruptive population dynamics, such as other
Ips and
Dendroctonus species.
In most Petri dish arena choice experiments, roofed (R) or non-roofed (nR) study trees were compared with one of the two non-manipulated control (C) trees, which may represent any vigorous Norway spruce in a forest composed of differentially water-supplied and resistant trees. In May 2020, the roofed trees were mildly drought-stressed, with minimum twig water potentials between −0.93 and −1.0 MPa. Due to a humid and rainy season, mean twig water potentials increased to −0.54/−0.59 MPa (R) and −0.35/−0.41 MPa (nR) in July/August, respectively, indicating generally low drought stress levels [
32]. Pre-dawn twig water potentials recorded in May, July, and August 2020 reflected the seasonal water status history of the study trees, which were sampled for bark cores in September 2020. Despite the supposedly equal water supply of nR and C trees, test beetles generally preferred the control samples in the Petri dish arena choice experiments. It cannot be excluded that manipulation by the sampling of bark and twigs and repeatedly conducting field bioassays [
1] over two vegetation periods influenced the quality of the study trees. In particular, R and nR trees were affected by infection with
G. penicillata in early August 2020, but necrotic lesions remained local in response to these artificial low-density inoculations [
45]. Thus, wound reaction zones covered only a small part of the stems of mature trees examined in our study. Therefore, we assume that the areas of hypersensitive wound reaction mainly indicated stress and defense states but did not additionally weaken the study trees.
Drought stress status of the study trees in May and July was positively correlated with the relative time spent by beetles near the bark cores from these trees and the proportions of final choices of study tree bark cores, as observed for the subset of “T and C1” experiments. Such a strong relationship was not found for C2 samples, although beetles did not show a clear preference for any control tree in “C1 and C2” tests (
Figure 3e). Overall, bark samples of R trees were preferred in 40–80% of cases (% T choice) over C1 or C2, and 40–70% of the time was spent in Petri dish sections containing bark cores from the more drought-stressed study trees. Preferences for and relative time spent near nR bark cores were much lower (
Figure 6c,d). The twig water potentials recorded in May and July 2020 turned out to be most relevant for final beetle choices as well as the mean values calculated for all three measurement dates, although stress levels were comparably low for R and nR trees in August 2020. The clear preferences of beetles for trees with a seasonal drought stress history observed in the bioassays confirm the previous identification of water deficits accumulated in spring and summer as significant predictors of European spruce bark beetle infestations in forest stands dominated by Norway spruce [
5].
Drought-stressed trees were not only more attractive to bark beetles but also less resistant to biotic attack. Roofed trees showed larger hypersensitive wound reaction zones in response to
G. penicillata inoculations than nR trees, indicating increased damage severity caused by the fungus and/or decelerated inhibition of fungal growth [
46]. Similar observations were made in a Swedish study, where higher precipitation sums accumulated two months prior to the inoculation with
Grosmannia europhioides (E.F. Wright & Cain) Zipfel, Z.W. de Beer & M.J. Wingf. led to smaller lesions in 20-year-old spruce trees [
47]. It is important to note that tissues were sampled from vital bark sections (unaffected by fungal infection) below the inoculation points, so that beetles could not be influenced in their decisions by potential odor signals of
G. penicillata. Adult
I. typographus are in fact strongly attracted to the more virulent species of their fungal assemblage, such as
G. penicillata and
E. polonica, while they respond indifferently to or are even repelled by the non-pathogenic and loosely associated
Ophiostoma piceae (Münch) H. Sydow & Sydow [
29]. Furthermore,
Grosmannia species contribute to
I. typographus mass aggregation by producing an important pheromone component [
48] and to a faster depletion of host tree resources by inducing strong defense responses (i.e., by causing larger necrotic lesions and terpene production) [
19,
49].
The size of the hypersensitive wound reaction caused by G. penicillata on study trees was not related to the duration of stay of bark beetles in Petri dish sections with bark cores from corresponding trees but were clearly positively correlated with proportions of final choice. However, a significant relationship between defense states and beetle choices was found solely when omitting the outliers R9 and nR8, which were highly attractive to beetles despite small lesion areas. The drought-stressed R9 tree (twig water potential of −0.89 MPa in May) was preferred over C1 in eight out of ten test runs, but was less attractive to beetles than the non-stressed nR9 (in R9/nR9 tests). Ambiguous test results were also found for nR8, which was highly preferred over C2, but not over C1. Yet, tree R2 with the lowest twig water potential measured in May (−1.13 MPa) was clearly more attractive to test beetles than the sufficiently water supplied nR8 (in R2/nR8 tests). Unclear beetle preferences may reflect to some extent the random components of bark beetle decisions in the bioassays or result from unknown characteristics of study and control trees.


 ,
, 







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}