
Linking Changes in Fatty Acid Composition to Postharvest Needle Abscission Resistance in Balsam Fir Trees

 and
and
Abstract
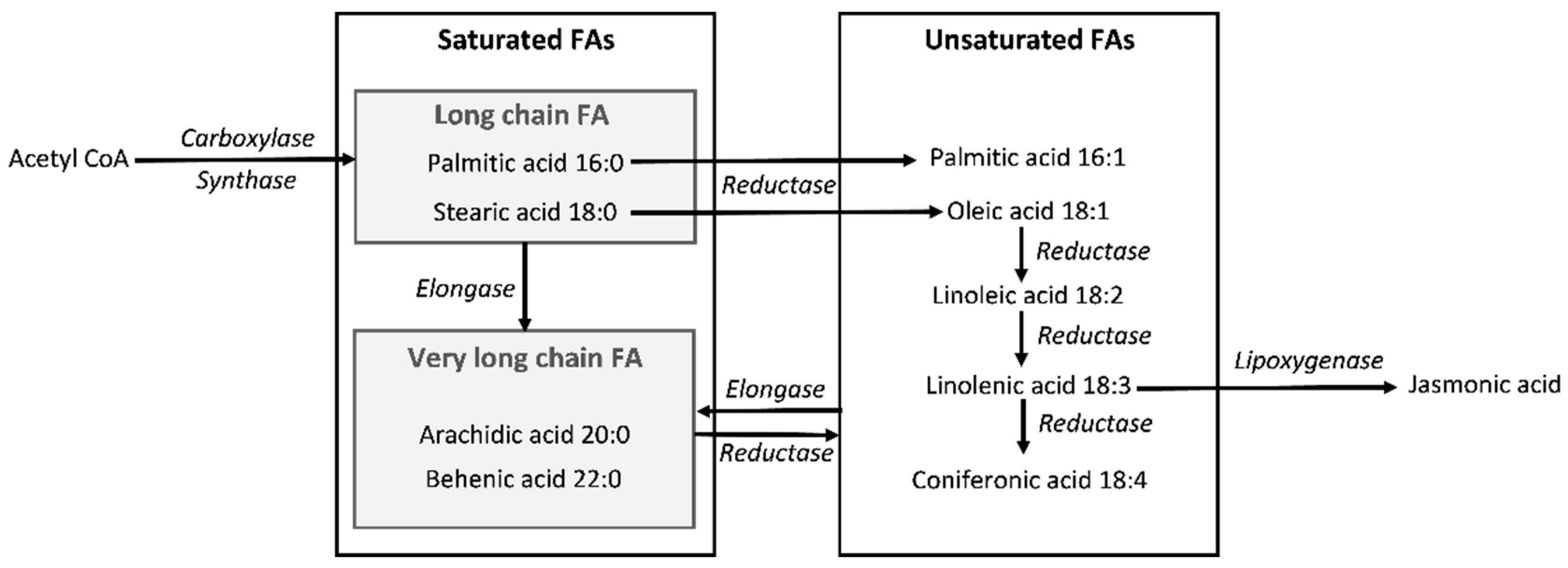
:1. Introduction
2. Materials and Methods
2.1. Sampling and Experimental Set-Up
2.2. Needle Abscission
2.3. FA Extraction
2.4. FA Analysis
2.5. Statistical Analysis
3. Results
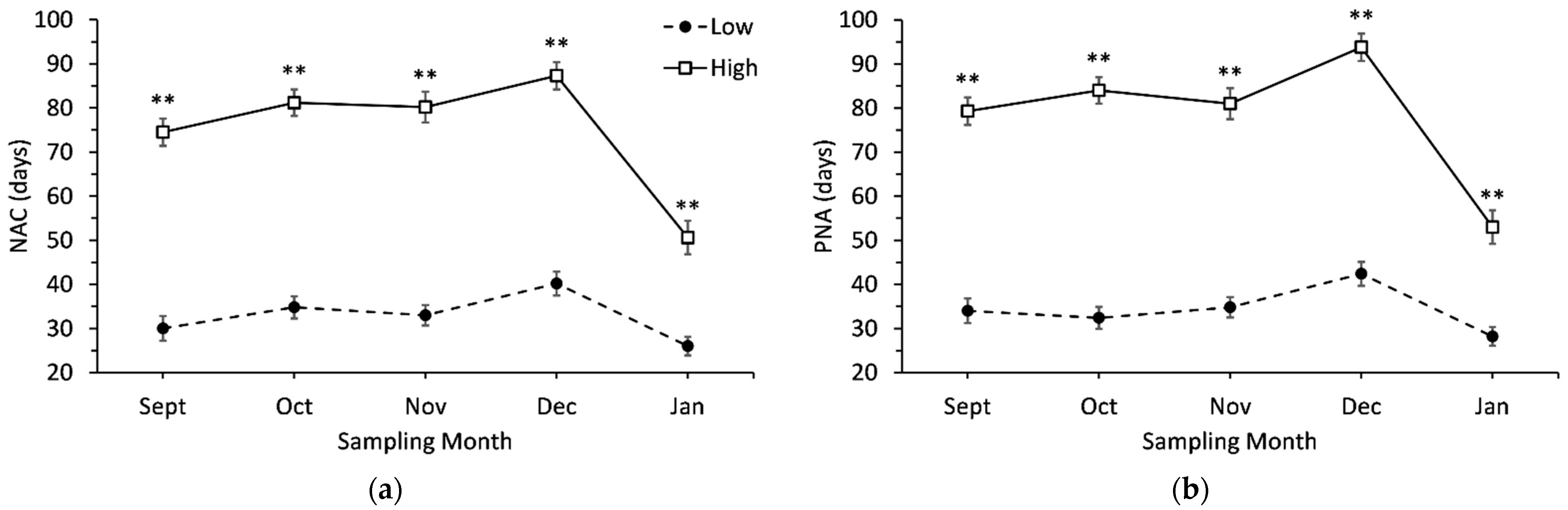
3.1. Needle Abscission Resistance
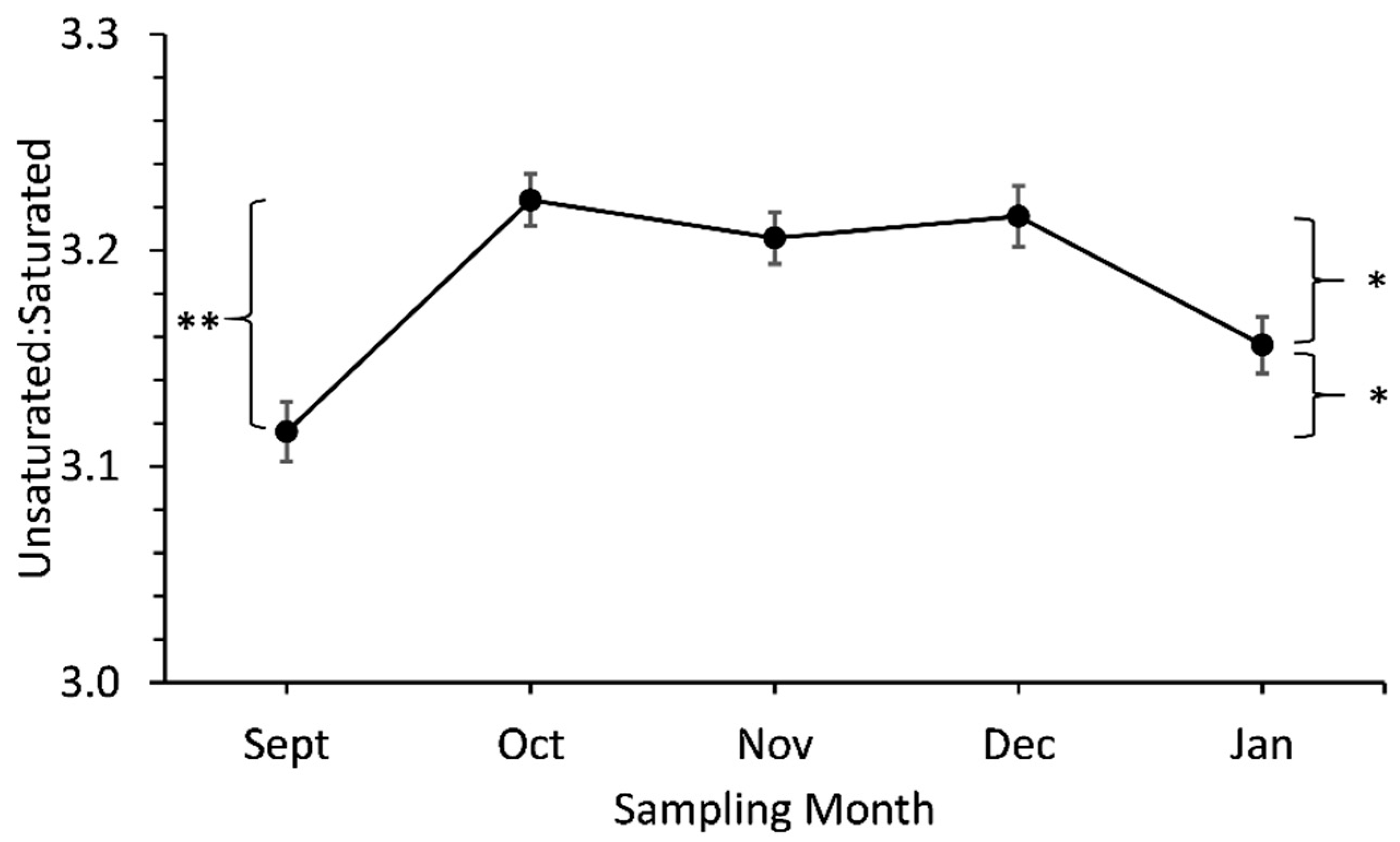
3.2. Unsaturated: Saturated FAs
3.3. Baseline FA Profiles
3.4. Genotype and Sampling Date Main Effects
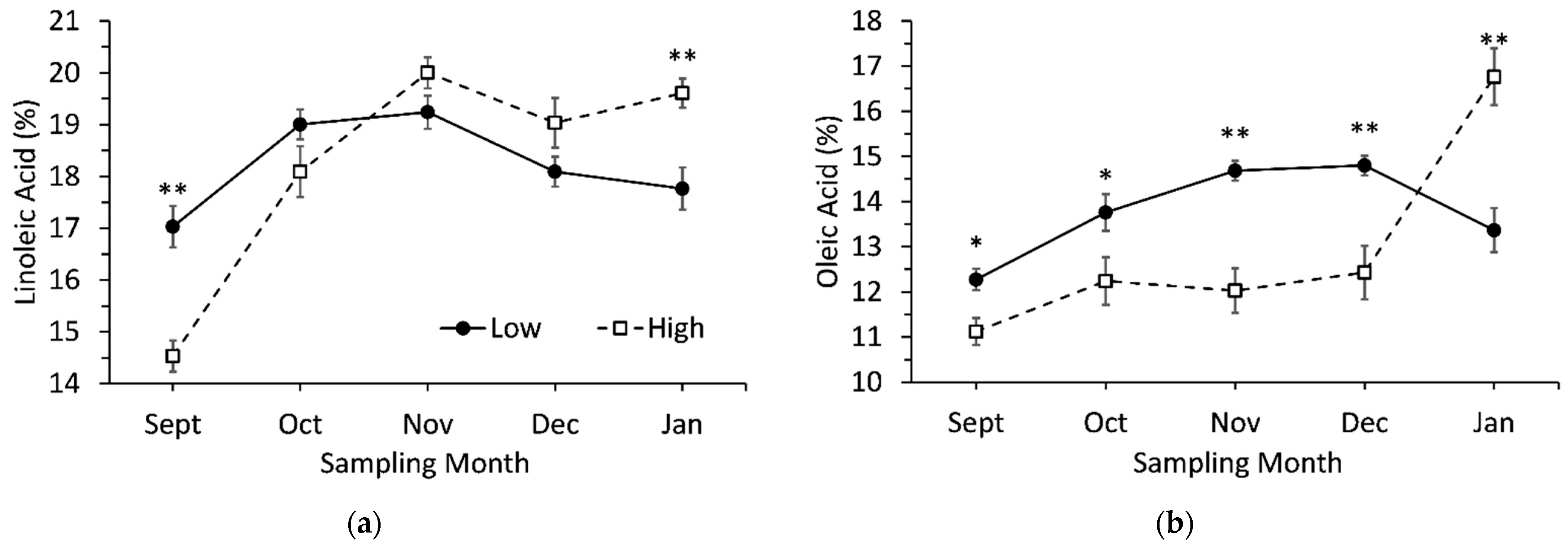
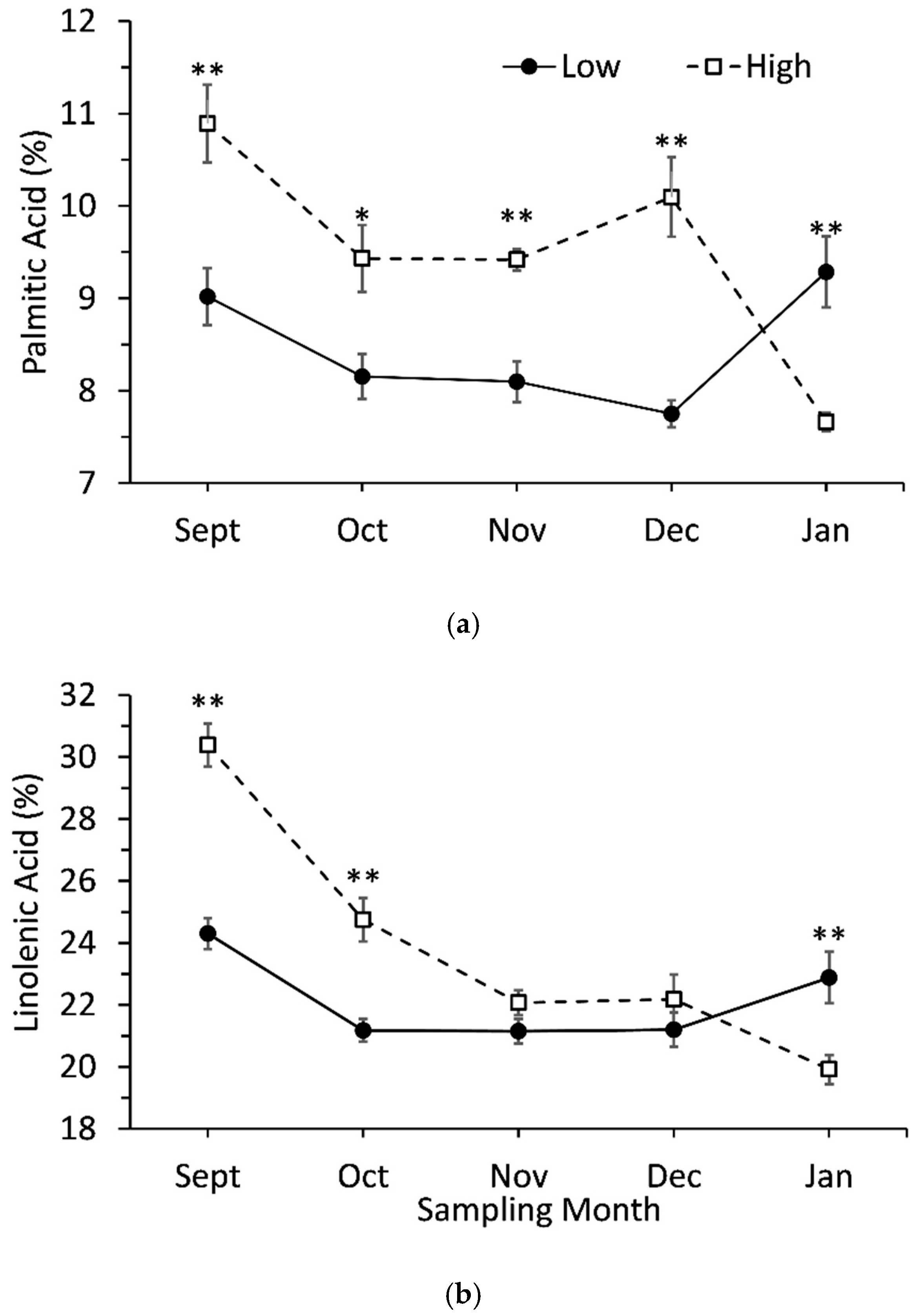
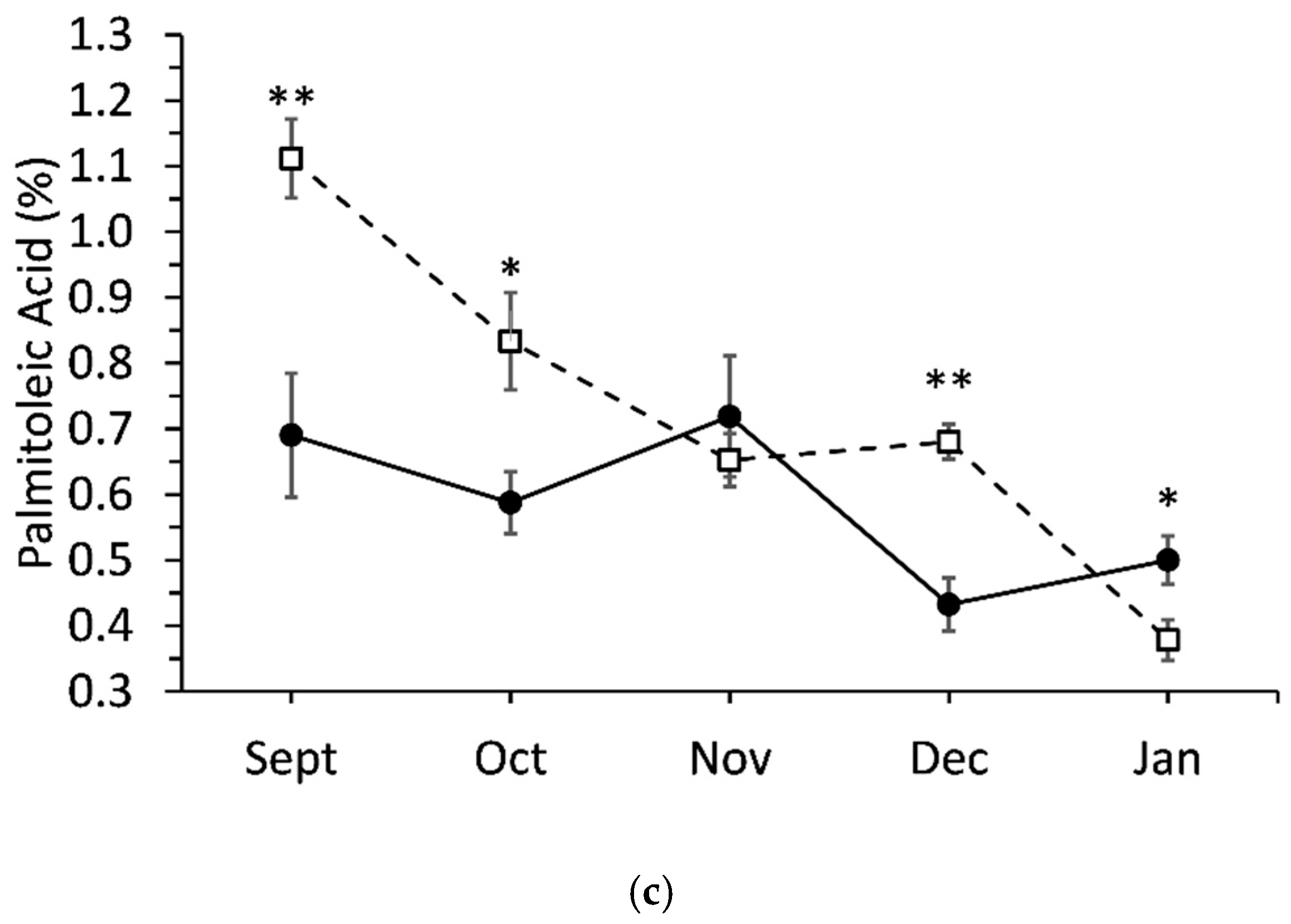
3.5. Interaction Effects between Genotypes and Sampling Date
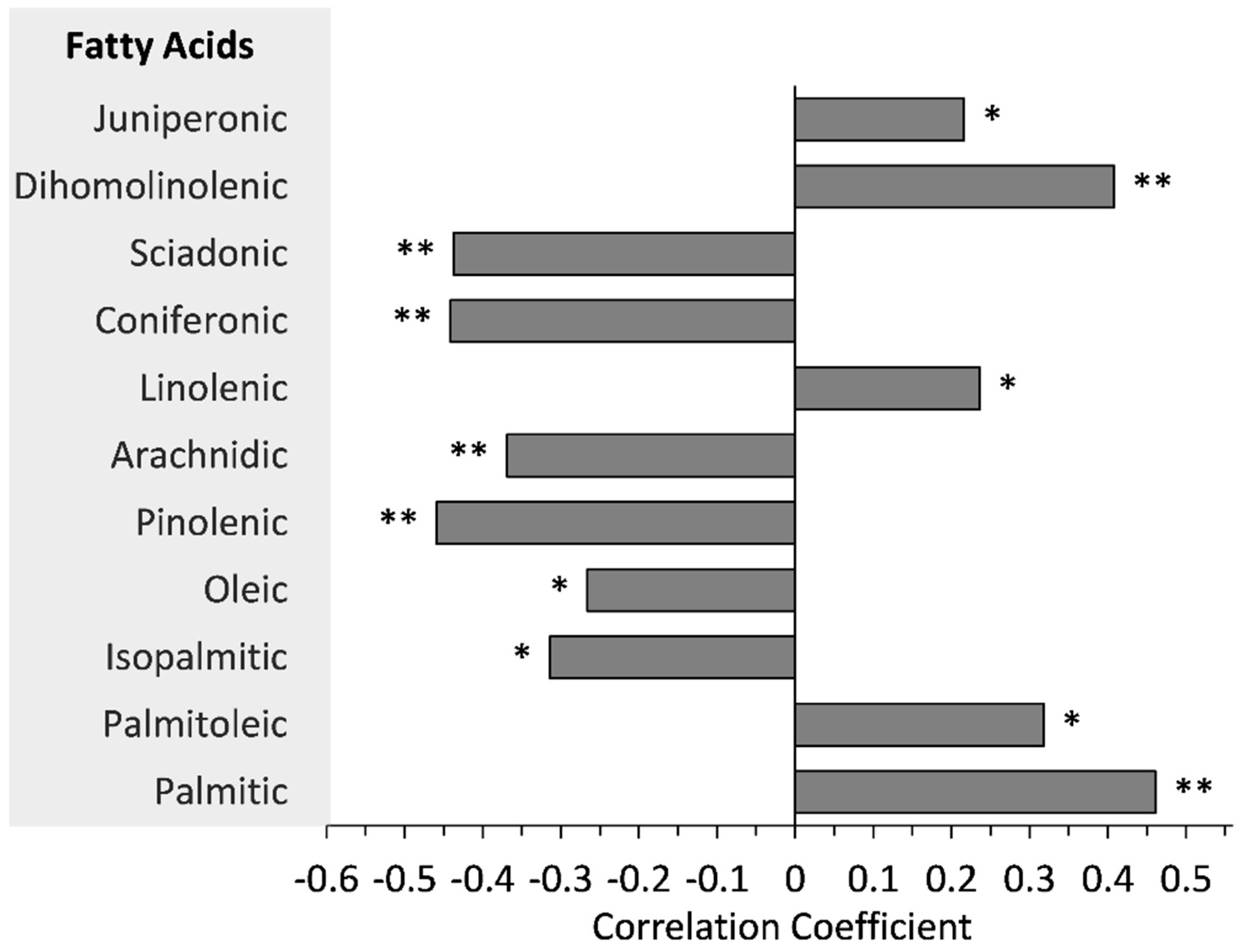
3.6. Relationships between FAs and NAR
4. Discussion
4.1. Genotypic Changes
4.2. Changes in FAs and Sampling Date
4.3. Interactive Effects Due to Cold Acclimation
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thiagarajan, A.; MacDonald, M.T.; Lada, R. Environmental and hormonal physiology of postharvest needle abscission in Christmas trees. Crit. Rev. Plant Sci. 2016, 35, 1–17. [Google Scholar] [CrossRef]
- Gusta, L.; Trischuk, R.; Weiser, C. Plant cold acclimation: The role of abscisic acid. J. Plant Growth Regul. 2005, 24, 308–318. [Google Scholar] [CrossRef]
- Odlum, K.D.; Blake, T.J. A comparison of analytical approaches for assessing freezing damage in black spruce using electrolyte leakage methods. Can. J. Bot. 1996, 74, 952–958. [Google Scholar] [CrossRef]
- MacDonald, M.T.; Lada, R.R.; Veitch, R.S. Seasonal changes in balsam fir needle abscission patterns and links to environmental factors. Scand. J. For. Res. 2016, 32, 438–445. [Google Scholar] [CrossRef]
- MacDonald, M.T.; Lada, R.R.; Veitch, R.S.; Thiagarajan, A.; Adams, A.D. Postharvest needle abscission resistance of balsam fir (Abies balsamea) is modified by harvest date. Can. J. For. Res. 2014, 44, 1394–1401. [Google Scholar] [CrossRef]
- Lada, R.R.; MacDonald, M.T. Understanding the physiology of postharvest needle abscission in balsam for. Front. Plant Sci. 2015, 6, 1069. [Google Scholar] [CrossRef]
- Jan, N.; Hussain, M.; Andrabi, K.I. Cold resistance in plants: A mystery unresolved. Electron. J. Biotechnol. 2009, 12, 14–15. [Google Scholar] [CrossRef]
- Ding, Y.; Shi, Y.; Yang, S. Advances and challenges in uncovering cold tolerance regulatory mechanisms in plants. New Phytol. 2019, 222, 1690–1704. [Google Scholar] [CrossRef] [Green Version]
- Quehenberger, O.; Armando, A.M.; Dennis, E.A. High sensitivity quantitative lipidomics analysis of fatty acids in biological samples by gas chromatography-mass spectrometry. Biochim. Biophys. Acta 2011, 1811, 648–656. [Google Scholar] [CrossRef] [Green Version]
- Harayama, T.; Riezman, H. Understanding the diversity of membrane lipid composition. Nat. Rev. Mol. Cell Biol. 2018, 19, 281–296. [Google Scholar] [CrossRef]
- He, M.; Qin, C.X.; Wang, X.; Ding, N.Z. Plant unsaturated fatty acids: Biosynthesis and regulation. Front. Plant Sci. 2020, 23, 390. [Google Scholar] [CrossRef] [PubMed]
- MacDonald, G.E.; Lada, R.R.; Caldwell, C.D.; Undigwe, C.; MacDonald, M.T. Lipid and fatty acid changes linked to post-harvest needle abscission in balsam fir, Abies balsamea. Trees 2020, 34, 297–305. [Google Scholar] [CrossRef]
- Partelli, F.L.; Batista-Santos, P.; Scotti-Campos, P.; Pais, I.P.; Quartin, V.L.; Vieira, H.D.; Ramalho, J.C. Characterization of the main lipid components of chloroplast membranes and cold induced changes in Coffea spp. Environ. Exp. Bot. 2011, 74, 194–204. [Google Scholar] [CrossRef]
- Sommerville, C.; Browse, J. Dissecting desaturation: Plants prove advantageous. Trends Cell Biol. 1996, 6, 148–153. [Google Scholar] [CrossRef]
- MacDonald, G.E.; Lada, R.R.; Caldwell, C.D.; Undigwe, C.; MacDonald, M.T. Seasonal changes in polar lipids linked to needle loss in four genotypes of balsam fir. Scand. J. For. 2022, in press.
- MacDonald, M.T.; Lada, R.R.; Veitch, R.S. Linking certain physical characteristics with postharvest needle abscission resistance in balsam fir. J. Appl. Hortic. 2014, 16, 37–39. [Google Scholar] [CrossRef] [Green Version]
- Mitcham-Butler, E.J.; Hinesley, L.E.; Pharr, D.M. Soluble carbohydrate concentration of Fraser fir foliage and its relationship to postharvest needle retention. J. Am. Soc. Hortic. Sci. 1987, 112, 672–676. [Google Scholar]
- Bligh, E.G.; Dwyer, W.J. A rapid method for total lipid extraction and purification. Can. J. Biochem. Physiol. 1959, 37, 911–917. [Google Scholar] [CrossRef]
- Christie, W.W. A simple procedure for rapid transmethylation of glycerolipids and cholesteryl esters. J. Lipid Res. 1982, 23, 1072–1075. [Google Scholar] [CrossRef]
- Miquel, M.; Cassagne, C.; Browse, J. A new class of Arabidopsis mutants with reduced hexadecatrienoic acid fatty acid levels. Plant Physiol. 1998, 117, 923–930. [Google Scholar] [CrossRef] [Green Version]
- Wolff, R.L.; Christie, W.W.; Coakley, D. The unusual occurrence of 14-methylhexadecanoic acid in Pinaceae seed oils among plants. Lipids 1997, 32, 971–973. [Google Scholar] [CrossRef] [PubMed]
- Semenova, N.V.; Makarenko, S.P.; Shmakov, V.N.; Konstabtinov, Y.M.; Dudareva, L.V. Fatty acid composition of total lipids from needles and cultured calluses of conifers Pinus sylvestris L., Picea pungens Engelm., Pinus koraiensis Siebold & Zucc., and Larix sibirica Ledeb. Biochemistry 2017, 11, 287–295. [Google Scholar] [CrossRef]
- Sato, M.; Yunoki, K.; Ohkubo, T.; Seki, K.; Tanaka, Y.; Ohnishi, M. Content and fatty acid composition of sulfoquinovosyldiacyglycerol in conifer leaves grown in Hokkaido, Japan. Biosci. Biotechnol. Biochem. 2010, 74, 1283–1285. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Wu, M.; Zhang, B.; Shrestha, P.; Petrie, J.; Green, A.G.; Singh, S.P. Genetic enhancement of palmitic acid accumulation in cotton seed oil through RNAi down-regulation of ghKAS2 encoding β-ketoacyl-ACP synthase II (KASII). Plant Biotechnol. J. 2016, 15, 132–143. [Google Scholar] [CrossRef] [Green Version]
- Yeats, T.H.; Rose, J.K.C. The formation and function of plant cuticles. Plant Physiol. 2013, 163, 5–29. [Google Scholar] [CrossRef] [Green Version]
- Mata-Perez, C.; Sanchez-Calvo, B.; Begara-Morales, J.C.; Luque, F.; Jimenez-Ruiz, J.; Padilla, M.N.; Fierro-Risco, J.; Valderrama, R.; Fernandez-Ocana, A.; Corpas, F.J.; et al. Transcriptome profiling of linolenic acid-responsive genes in ROS signaling from RNA-seq data in Arabidopsis. Front. Plant Sci. 2015, 6, 122. [Google Scholar] [CrossRef] [Green Version]
- Wasternack, C. Action of jasmonates in plant stress responses and development-applied aspects. Biotechnol. Adv. 2014, 32, 31–39. [Google Scholar] [CrossRef]
- Wasternack, C. Jasmonates: An update on biosynthesis, signal transduction, and action in plant stress response, growth and development. Ann. Bot. 2007, 100, 681–697. [Google Scholar] [CrossRef] [Green Version]
- MacDonald, M.T.; Lada, R.R. Changes in endogenous hormone levels explain season variation in balsam fir needle abscission patterns. J. Plant Growth Regul. 2017, 36, 723–733. [Google Scholar] [CrossRef]
- Huo, K.; Shui, L.; Mai, Y.; Zhou, N.; Liu, Y.; Zhang, C.; Niu, J. Effects of exogenous abscisic acid on oil content, fatty acid composition, biodiesel properties and lipid components in developing Siberian apricot (Prunus sibirica) seeds. Plant Physiol. Biochem. 2020, 154, 260–267. [Google Scholar] [CrossRef]
- Ju, Y.L.; Liu, M.; Zhao, H.; Meng, J.F.; Fang, Y.L. Effect of exogenous abscisic acid and methyl jasmonate on anthocyanin composition, fatty acids, and volatile compounds of cabernet sauvignon (Vitis vinifera L.) grape berries. Molecules 2016, 21, 1354. [Google Scholar] [CrossRef] [Green Version]
- Shi, P.; Hua, W.; Htwe, Y.M.; Zhang, D.; Li, J.; Wang, Y. Abscisic acid improves linoleic acid accumulation possibly by promoting expression of EgFAD2 and other fatty acid biosynthesis genes in oil palm mesocarp. Front. Plant Sci. 2021, 12, 748130. [Google Scholar] [CrossRef] [PubMed]
- Makarenko, S.P.; Konstaninov, Y.U.; Shmakov, V.M.; Konenkina, T.A. Fatty acid composition of fatty acids in the calluses of two pine species Pinus sibirica and Pinus sylvestris. Russ. J. Plant Physiol. 2010, 57, 739–743. [Google Scholar] [CrossRef]
- Wolff, R.L.; Christie, W.W.; Pedrono, F.; Marpeau, A.M.; Tsevegsuren, N.; Aitzetmuller, K.; Gunstone, F.D. Delta5-olefinic acids in the seed lipids from four Ephedra species and their distribution between alpha and beta positions of triacylglycerols. Characteristics common to coniferophytes and cycadophytes. Lipids 1999, 34, 855–864. [Google Scholar] [CrossRef] [PubMed]
- Zadernowski, R.; Naczk, M.; Czaplicki, S. Chemical composition of Pinus sibririca nut oils. Eur. J. Lipid Sci. 2009, 111, 698–704. [Google Scholar] [CrossRef]
- Interpretative Guide for Fatty Acids. Available online: https://www.gdx.net/core/interpretive-guides/Fatty-Acids-IG.pdf (accessed on 30 March 2022).
- Yaeno, T.; Matsuda, O.; Iba, K. Role of chloroplast trienoic fatty acids in plant disease defense responses. Plant J. 2004, 40, 931–941. [Google Scholar] [CrossRef]
- Chen, F.; Zhang, W.; Yu, K.; Sun, L.; Gao, J.; Zhou, X.; Peng, Q.; Fu, S.; Hu, M.; Long, W.; et al. Unconditional and conditional QTL analyses of seed fatty acid composition in Brassica napus L. BMC Plant Biol. 2018, 18, 49. [Google Scholar] [CrossRef] [Green Version]
- Janila, P.; Pandey, M.K.; Shasidhar, Y.; Variath, M.T.; Sriswathi, M.; Khera, P.; Manohar, S.S.; Nagesh, P.; Vishwakarma, M.K.; Mishra, G.P.; et al. Molecular breeding for introgression of fatty acid desaturase mutant alleles (ahFAD2A and ahFAD2B) enhances oil quality in high and low oil containing genotypes. Plant Sci. 2016, 242, 203–213. [Google Scholar] [CrossRef] [Green Version]
- Gelvin, S.B. Agrobacterium-mediated plant transformation: The biology behind the “gene-jockeying” tool. Microbiol. Mol. Biol. 2003, 67, 16–37. [Google Scholar] [CrossRef] [Green Version]
- Ruiz-Lopez, N.; Usher, S.; Sayanova, O.V.; Napier, J.A.; Haslam, R.P. Modifying the lipid content and composition of plant seeds: Engineering the production of LC-PUFA. Appl. Microbiol. Biotechnol. 2015, 99, 143–154. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Fatty Acid | Low NAR | High NAR | Sig. |
|---|---|---|---|
| TOTAL SATURATED | 23.7 ± 0.5 | 23.7 ± 0.4 | |
| Myristic acid (14:0) | 1.4 ± 0.1 | 1.5 ± 0.1 | |
| Palmitic acid (16:0) | 9.1 ± 0.3 | 10.9 ± 0.4 | ** |
| Isopalmitic acid (14-methyl-16:0) | 5.3 ± 0.3 | 4.8 ± 0.3 | |
| Isomargaric acid (14-methyl-17:0) | 0.4 ± 0.1 | 0.3 ± 0.1 | |
| Stearic acid (18:0) | 2.9 ± 0.2 | 2.5 ± 0.3 | |
| Arachidic acid (20:0) | 1.0 ± 0.1 | 0.4 ± 0.1 | ** |
| Behenic acid (22:0) | 1.5 ± 0.2 | 1.3 ± 0.3 | |
| Tricosylic acid (23:0) | 1.0 ± 0.1 | 0.8 ± 0.1 | |
| Lignoceric acid (24:0) | 1.1 ± 0.1 | 1.2 ± 0.1 | |
| TOTAL UNSATURATED | 76.3 ± 0.5 | 76.3 ± 0.4 | |
| Palmitoleic acid (9-cis-16:1) | 0.8 ± 0.1 | 1.1 ± 0.1 | * |
| Oleic acid (9-cis-18:1) | 12.1 ± 0.2 | 11.2 ± 0.3 | * |
| Taxoleic acid (5,9-cis-18:2) | 3.0 ± 0.2 | 3.2 ± 0.2 | |
| Linoleic acid (9,12-cis-18:2) | 17.0 ± 0.4 | 14.5 ± 0.3 | ** |
| Pinolenic acid (5,9,12-cis-18:3) | 7.0 ± 0.3 | 5.1 ± 0.3 | ** |
| Linolenic acid (9,12,15-cis-18:3) | 24.3 ± 0.5 | 30.4 ± 0.7 | ** |
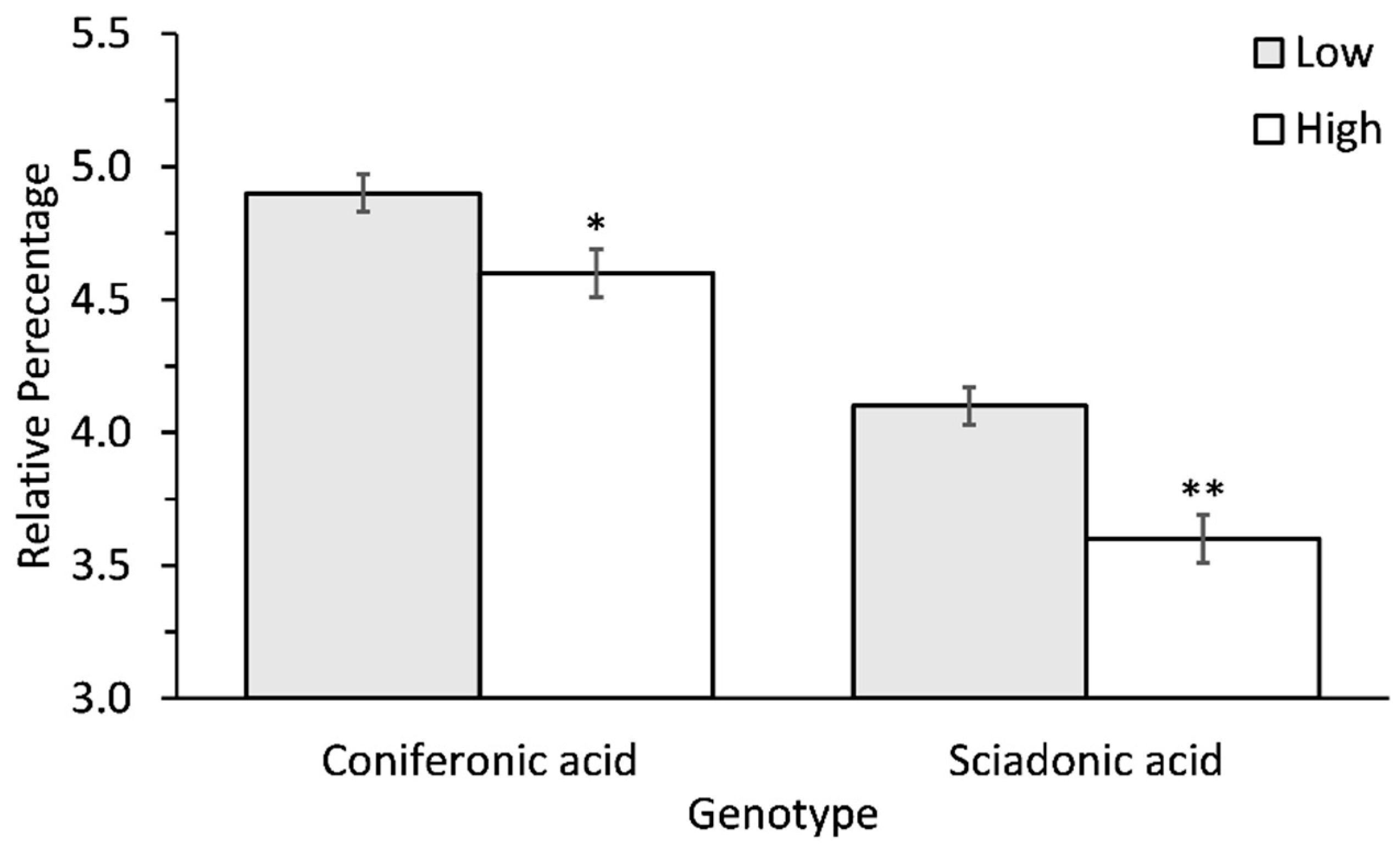
| Coniferonic acid (5,9,12,15-cis-18:4) | 4.7 ± 0.1 | 4.4 ± 0.1 | * |
| Icosadienoic acid (11,14-cis-20:2) | 0.6 ± 0.1 | 0.3 ± 0.1 | * |
| Sciadonic acid (5,11,14-cis-20:3) | 4.5 ± 0.2 | 3.7 ± 0.1 | ** |
| Dihomolinolenic acid (11,14,17-cis-20:3) | 0.4 ± 0.1 | 0.4 ± 0.1 | |
| Juniperonic acid (5,11,14,17-cis-20:4) | 1.9 ± 0.1 | 2.0 ± 0.2 |
| Fatty Acid | Sept | Oct | Nov | Dec | Jan | Sig. |
|---|---|---|---|---|---|---|
| Stearic acid (18:0) | 2.7 ± 0.1 | 2.6 ± 0.1 | 3.0 ± 0.1 | 3.2 ± 0.1 | 3.0 ± 0.1 | ** |
| Icosadienoic acid (11,14-cis-20:2) | 0.4 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | 0.6 ± 0.1 | * |
| Sciadonic acid (5,11,14-cis-20:3) | 4.3 ± 0.1 | 4.3 ± 0.2 | 3.8 ± 0.1 | 3.8 ± 0.1 | 3.9 ± 0.1 | * |
| Behenic acid (22:0) | 1.4 ± 0.1 | 1.6 ± 0.1 | 1.9 ± 0.1 | 1.9 ± 0.1 | 2.0 ± 0.1 | ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MacDonald, G.E.; Lada, R.R.; Caldwell, C.D.; Udenigwe, C.C.; MacDonald, M.T. Linking Changes in Fatty Acid Composition to Postharvest Needle Abscission Resistance in Balsam Fir Trees. Forests 2022, 13, 800. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050800
MacDonald GE, Lada RR, Caldwell CD, Udenigwe CC, MacDonald MT. Linking Changes in Fatty Acid Composition to Postharvest Needle Abscission Resistance in Balsam Fir Trees. Forests. 2022; 13(5):800. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050800
Chicago/Turabian StyleMacDonald, Gaye E., Rajasekaran R. Lada, Claude D. Caldwell, Chibuike C. Udenigwe, and Mason T. MacDonald. 2022. "Linking Changes in Fatty Acid Composition to Postharvest Needle Abscission Resistance in Balsam Fir Trees" Forests 13, no. 5: 800. https://0-doi-org.brum.beds.ac.uk/10.3390/f13050800