
Foliar Water Uptake Capacity in Six Mangrove Species
by
 , and
, and
Katrien H. D. Schaepdryver
1,* ,
,
Willem Goossens
1,
Abdulla Naseef
2,
Sreejith Kalpuzha Ashtamoorthy
2 and
Kathy Steppe
1 1
Laboratory of Plant Ecology, Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, Coupure Links 653, 9000 Ghent, Belgium
2
Department of Forest Ecology, Kerala Forest Research Institute, Thrissur 680653, India
*
Author to whom correspondence should be addressed.
Forests 2022, 13(6), 951; https://0-doi-org.brum.beds.ac.uk/10.3390/f13060951
Submission received: 3 May 2022
/
Revised: 9 June 2022
/
Accepted: 15 June 2022
/
Published: 17 June 2022
(This article belongs to the Special Issue Advances in Mangrove Ecology)
Abstract
:Foliar water uptake (FWU) is a mechanism that enables plants to acquire water from the atmosphere through their leaves. As mangroves live in a saline sediment water environment, the mechanism of FWU might be of vital importance to acquire freshwater and grow. The goal of this study was to assess the FWU capacity of six different mangrove species belonging to four genera using a series of submersion experiments in which the leaf mass increase was measured and expressed per unit leaf area. The foliar water uptake capacity differed between species with the highest and lowest average water uptake in Avicennia marina (Forssk.) Vierh. (1.52 ± 0.48 mg H2O cm−2) and Bruguiera gymnorhiza (L.) Lam. (0.13 ± 0.06 mg H2O cm−2), respectively. Salt-excreting species showed a higher FWU capacity than non-excreting species. Moreover, A. marina, a salt-excreting species, showed a distinct leaf anatomical trait, i.e., trichomes, which were not observed in the other species and might be involved in the water absorption process. The storage of leaves in moist Ziplock bags prior to measurement caused leaf water uptake to already occur during transport to the field station, which proportionately increased the leaf water potential (A. marina: −0.31 ± 0.13 MPa and B. gymnorhiza: −2.70 ± 0.27 MPa). This increase should be considered when performing best practice leaf water potential measurements but did not affect the quantification of FWU capacity because of the water potential gradient between a leaf and the surrounding water during submersion. Our results highlight the differences that exist in FWU capacity between species residing in the same area and growing under the same environmental conditions. This comparative study therefore enhances our understanding of mangrove species’ functioning.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Mangroves occupy about 13,700,000 ha of coastline worldwide, providing multiple ecosystem services, such as food supply and protection against storms [1,2]. However, climate change will lead to a global increase in drought frequency and duration as a consequence of altering precipitation patterns and more extreme weather events [3]. This seasonal variation in rainfall patterns will most likely have a profound effect on species’ distribution and the productivity of mangrove forests [4,5]. Periods with abnormally low rainfall can therefore alter the average salinity and complicate the ability of mangroves to take up water [6]. Accompanied by higher evaporation and transpiration rates due to future temperature extremes, the saline conditions will further intensify, leading to a decrease in the net primary production, seedling survival and conversion of the upper tidal zone to hypersaline mudbanks [7,8].
In assessment studies about the resilience of mangrove species (and plant species in general) under changing climatic conditions, the phenomenon of foliar water uptake (FWU) is often overlooked. In contrast to the classical cohesion–tension theory of Dixon and Joly [9], in which transpiration is defined as the driving force for the ascent of sap in trees [10,11], with the resulting water movement from roots through the stem and towards the canopy in the so-called soil–plant–atmosphere continuum (SPAC) [12], FWU describes the movement of water in the opposite direction [12,13]. This implies that plants are capable of passively absorbing a substantial amount of water directly through their leaves during leaf wetting events (e.g., rainfall, fog or dew) [14,15]; after which, it is transported from the leaves to the stem, roots [16,17] and possibly even the soil, resulting in a sap flow reversal [18]. FWU has been demonstrated in at least 200 species spanning various biomes, which shows that it is a widespread phenomenon [13,19]. Researchers have elaborated on this topic in mangroves to address the potential importance of this mechanism in a future with more unpredictable climate conditions. A previous study of Steppe et al. [20] showed that the mangrove species Avicennia marina (Forssk.) Vierh. is capable of freshwater uptake through the leaves. In fact, it provided evidence that these seemingly small water inputs resulted in turgor-driven growth spurts and can therefore decouple, at least to some extent, the water status from the sediment water uptake [21]. The large-scale 2015 dieback of mangroves in the Gulf of Carpentaria, Australia, demonstrated that prolonged drought events are detrimental to mangroves [22] with, in this particular case, FWU apparently not being able to mitigate tree drought stress.
Four entering pathways for FWU are commonly accepted: absorption by (i) stomata, (ii) trichomes or (iii) hydathodes and (iv) diffusion through the cuticle [13,19,23,24]. Recently, Bryant et al. [25] identified cork warts as a novel entering pathway in the mangrove species Sonneratia alba Sm. and Sonneratia caseolaris (L.) Engl. Liquid water transport through the stomata was previously thought to be impossible due to water surface tension and stomatal morphology, combined with the hydrophobicity of the cuticle [26]. Currently, this opinion is no longer shared in the scientific community, as Burkhardt et al. [27] showed that stomata can be ‘activated’ (hydraulic activation of stomata, HAS). HAS can occur by fungal hyphae entering the stomata [15], by stomatal mucilage [27] or by hygroscopic particles (e.g., salts and trace gases) on the leaf surface [19,27]. Coopman et al. [28] showed that deliquescence of secreted salt on the leaf surface of A. marina enables the absorption of atmospheric moisture through the leaves, even in unsaturated atmospheres. According to Guzmán-Delgado et al. [24], leaves with open stomata hydrated two times faster than leaves with closed stomata, indicating the importance of stomata for FWU. Schreel et al. [23] discovered that trichomes can also play a major role in the FWU of beech (Fagus sylvatica L.). However, trichomes are highly variable in their morphology (e.g., hair or scale-like) and function (e.g., glandular or non-glandular), with every type having a different FWU capacity [19]. Martin and von Willert [29] showed the ability of Crassula spp. to take up water through their hydathodes during nocturnal misting. Schreel and Steppe [13] questioned this sole conclusion, because the nocturnal opening of stomata in this CAM (Crassulacean Acid Metabolism) plant may also play a role in the observed FWU. The diffusion of water through the cuticle depends on the cuticular wettability and permeability [30]. While the cuticular thickness does not appear to affect the permeability [31], the composition of cuticular wax does [32]. Kamtsikakis et al. [33] demonstrated that the water permeation properties of plant cuticles can be asymmetric, with the water-responsive, plasticizable cutin-rich outer cuticular side controlling the water transport directionality. This enables the leaf to retain water in dry conditions and to take up moisture through the cuticle during fog and rain by swelling of the cuticle from the outer side [33].
The unique ecosystem of mangrove forests survives in a challenging environment of fluctuating salinity and waterlogged sediments. Mangroves have some remarkable adaptations to survive in this physiologically dry environment. In the literature, mangrove species have been classified into three putative groups of mechanisms of coping with salt, the so-called ‘salt management strategies’: (i) salt excretors, (ii) salt excluders and (iii) salt accumulators [34,35,36]. The exclusive classification of mangrove species into one of these three salt management strategies can be misleading if not interpreted wisely [37]. In fact, all mangrove species exclude salt at root level, but some exclude up to 99% [34,38]. Similarly, all mangrove species accumulate salt in their vacuoles, but some will be better at it. In contrast, only some species possess salt glands that allow them to excrete salt at the leaf level, which makes classification into salt excretors versus non-excretors more useful.
The main objective of the present study is to assess the FWU capacity in six mangrove species with a different physiological background (i.e., salt management strategies) and different leaf functional traits (e.g., presence of trichomes). This variability in physiological and anatomical features likely reflects a continuum in FWU capacity across species in the mangrove group. Our specific working hypothesis was that there will be significant differences in FWU capacity between the six species. This knowledge is crucial to understanding the impact and importance of freshwater uptake via leaves in mangrove species, which are keystones in an ecosystem most vulnerable to a rapidly changing climate.
2. Materials and Methods
2.1. Site Description
The experiments were conducted at Ashtamudi Lake, Kollam District, Kerala, southwestern coast of India (8°56′ N, 76°33′ E). This water body, with an area of about 32 km2, is an estuarine wetland connected to the Arabian Sea. Leaf samples were collected from a group of five sedimentary islands located along the west side of the lake, 2.5 km from the Arabian Sea (Figure S1). These islands are located close to each other and have a limited surface area, with the largest island being 6738 m2. All sampled trees were located within a 500-m radius, resulting in similar salinity conditions. The salinity largely differs, depending on the location in the 14-km-long lake and on the time of the year, and ranges between 14 and 25‰ [39]. The saline lake is fed with freshwater by the Kallada River, which originates from the Western Ghats at an altitude of 1100 m. Since 2002, the Ashtamudi wetland is included in the list of wetlands of international importance, as defined by the Ramsar Convention for conservation and the sustainable utilization of wetlands [39]. The average annual rainfall at Ashtamudi Lake is 2298 mm [40], of which one-third falls during the Monsoon months (June–September) [41]. The total annual rainfall in 2020 was 2405 mm, recorded with a manual rain gauge onsite. The mean annual temperature is 27.8 °C [40].
One species (S. caseolaris) was collected from Ponnarimangalam (10°00′ N, 76°16′ E) on the banks of Vembanadu Lake, near Vallarpadam, Ernakulam District, Kerala, as this species could not be found at Ashtamudi Lake. This site is about 120 km away from the main sampling site. The salinity in Vembanadu Lake ranges between 9 and 30‰, depending on the distance from the estuary [42]. The average annual rainfall and mean annual temperature at Vembanadu Lake are 2977 mm and 27.6 °C, respectively [40].
2.2. Experimental Set-Up
To assess the foliar water uptake (FWU) capacity in six mangrove species belonging to four different genera, an adapted version of the FWU capacity submersion experiment of Limm et al. [14] was performed (see below). The selected mangrove species were: Avicennia marina (Forssk.) Vierh., Aegiceras corniculatum (L.) Blanco, Bruguiera cylindrica (L.) Blume, Bruguiera gymnorhiza (L.) Lam., Sonneratia alba Sm. and Sonneratia caseolaris (L.) Engl. Considering the different genera from which samples were taken, one or more salt management strategies can be assigned based on previous research. Aegiceras spp. combine the excreting (through salt glands) and excluding (by ultrafiltration) salt management strategies [43]. Avicennia spp. adds to this the accumulating strategy by the storage of toxic levels of sodium ions in their vacuoles, maintaining the cytoplasmic concentration below 50 mm [44]. Bruguiera spp. and Sonneratia spp. act as both accumulators [45,46] and excluders, with the latter filtering almost 100% of salt at root level [47,48].
For each species, six individual trees were selected. From these individuals, five nodes with a minimum of two leaves were collected by cutting below and/or above the node with pruning shears (Figure 1). The nodes were randomly selected from different branches in such a way that the leaves were sun-exposed, fully expanded and not damaged by pathogens or herbivore attacks. All samples were collected between 31 July and 2 August 2019 between 2 and 4 pm. On the sampling days, the temperature was approximately 30 °C, and no precipitation occurred.
Leaf samples of each individual were put in a plastic Ziplock bag together with moist paper tissue. Before sealing the bag, air was exhaled into the bag to create a humid environment. The bags were kept in a thermostatic, lightproof box during transportation and until measurements were performed. Logistically, measurements could not be performed directly in the field. Therefore, leaves spent two to four hours in the bags, with the exception of leaves of the more remote S. caseolaris (7 h).
2.3. Foliar Water Uptake Capacity
At the field station, FWU capacity was determined according to the adapted submersion protocol of Limm et al. [14]. The only aspect in which the protocol differed from that of Limm et al. [14] is that residual water on the leaf surface after towel-drying was not considered, because this difference was minimal and not larger than the error of the balance on which the leaf samples were weighed. From each node, one leaf was picked. Simultaneously, an adjacent leaf was picked to measure the water potential (Ψleaf; MPa) (see below). Petioles of the picked leaves were sealed by non-acid vinyl glue, leaves were weighed (Mfresh; mg), taped to petri dishes, submerged in distilled water while avoiding contact between water and petioles and kept in a dark environment. After three hours, the leaves were patted dry and reweighed (Msub; mg), and the leaf area (A; cm2) was measured. The water uptake per unit of area (FWU capacity; mg H2O cm−2) was calculated:
The oven dry mass (Mdry; mg) was determined after drying the leaves in a dry oven for 72 h at 65 °C. The difference in leaf water content (ΔLWC; %) was calculated:
2.4. Water Potential
The leaf water potential (Ψleaf; MPa) was determined with a pressure chamber (Model 610, PMS Instrument Company, Albany, OR, USA). For every leaf used to determine the FWU capacity, the water potential of the adjacent leaf was measured.
2.5. Leaf Anatomy
Small segments of leaf tissue along the midrib at a position approximately halfway between base and apex were excised and processed for embedding in paraffin following Yeung et al. [50]. Transverse sections of 10 micron were made with a rotation microtome (Automatic MT3, NK Systems Limited, Nishi-Shinbashi, Tokyo) and collected on microscopic slides coated with silane. Safranin O (1% w/v in aqueous solution) and fast green (1% w/v in clove oil) double staining were carried out according to Johansen [51]. After staining, the sections were washed with clove oil and mounted in glycerin. Micrographs were taken using a Leica DFC295 camera mounted on a Leica DM1000 LED microscope.
2.6. Statistical Analysis
Statistical analyses were conducted using R software (v.3.6.1) [52] with the RStudio interface [53]. Linear models were adjusted using the ‘stats’ package [52] to assess differences in FWU capacity, ΔLWC and Ψleaf (variables) between species (factor). For all models, the five values per tree were averaged. When a significant effect was found, Tukey’s comparisons were tested using the ‘multcomp’ package [54]. The assumption of data normality was tested with the Shapiro–Wilk Normality test (‘stats’ package [52]), and the data were log-transformed if necessary. The latter was the case for the variable FWU capacity and ΔLWC. The assumption of data homoscedasticity was tested with the Levene test (‘car’ package [55]). To assess differences in FWU capacity and Ψleaf between excreting and non-excreting species, a Welch’s t-test (‘stats’ package [52]) was conducted, because the data were heteroscedastic. Again, the variable FWU capacity was log-transformed to fulfill the normality assumption. Observations exceeding 2.5 standard deviations above and below the mean were considered outliers and excluded from analyses. In the following sections, the mean values ± standard deviation are reported. When data had to be log-transformed for statistical analysis, the back-transformed mean values are reported.
3. Results
3.1. Species Level
The foliar water uptake capacity of the six mangrove species, belonging to four genera, ranged from 0.08 mg H2O cm−2 to 2.24 mg H2O cm−2 (Figure 2a). The difference in leaf water content (ΔLWC) varied between 0.05% and 1.71%. As a linear relationship (marginal R2 = 0.74, conditional R2 = 0.99) existed between the FWU capacity and ΔLWC, similar patterns between species for both variables were found (Figure 2a and Figure S2). Measurements of the water potential (Ψleaf) of the leaves ranged from −3.03 MPa to −0.14 MPa (Figure 2b).
The results show that all the leaves took up water during the submersion experiment. B. gymnorhiza showed with 0.13 ± 0.06 mg H2O cm−2 the lowest FWU capacity (p < 0.05) (Figure 2a). A. marina had the highest average FWU capacity with 1.52 ± 0.48 mg H2O cm−2, but this was not significantly different from the species A. corniculatum and S. caseolaris (p > 0.05) (Figure 2a). Bruguiera spp. had the most negative average Ψleaf (i.e., −2.70 ± 0.26 MPa), followed by Sonneratia spp. (i.e., −1.62 ± 0.44 MPa) and with A. marina and A. corniculatum showing the least negative average values (i.e., −0.31 ± 0.13 MPa and −0.35 ± 0.12 MPa, respectively) (Figure 2b).
It is known that phylogenetically more closely related species (e.g., same genus) tend to show similarities in functional traits due to coexistence in ecologically similar niches [56,57]. The micrographs (see below, Figure 3) indeed show a similar leaf architecture between species belonging to the same genus. However, there is a significant difference in FWU capacity between B. cylindrica and B. gymnorhiza (Figure 2a), both belonging to the Bruguiera genus. The same is true for Ψleaf of the two Sonneratia species (Figure 2b). For this reason, the discussion will mainly focus on the species level.
3.2. Leaf Anatomy
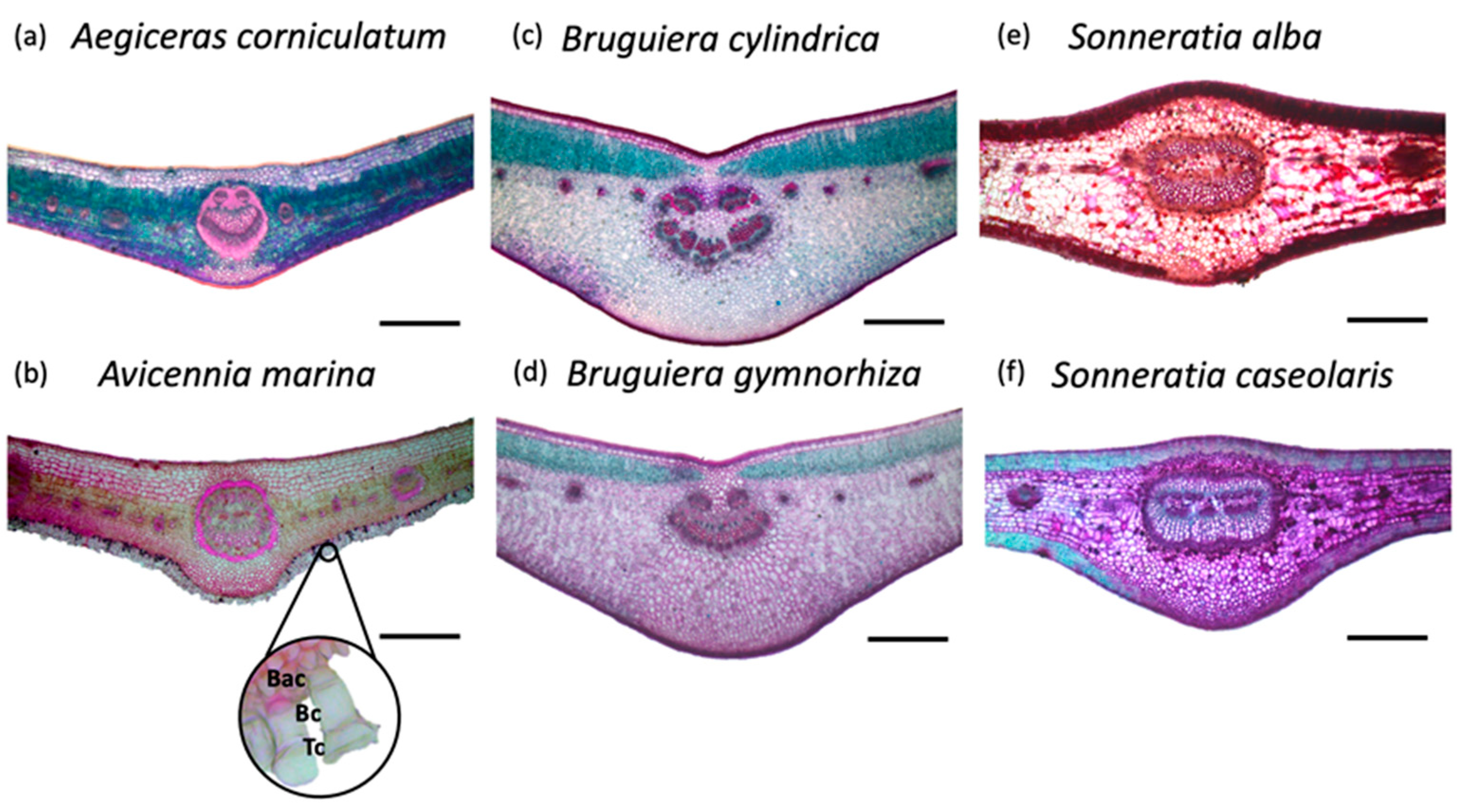
A survey of the micrographs (Figure 3) showed that the leaf architecture is similar between species belonging to the same genus. Species of the Aegiceras, Avicennia and Bruguiera genera have dorsiventral leaves in contrast to the Sonneratia genus, which has an isolateral leaf structure. A clear anatomical distinction of A. marina compared to the other species is the abundance of abaxial, awl-shaped multicellular trichomes, consisting of a top cell, one or two body cells and ending in a basal cell (Figure 3b). Although Bryant et al. [25] identified cork warts in Sonneratia sp. and Evans et al. [58] in B. gymnorhiza, none were observed on the transverse micrographs (Figure 3d–f).
3.3. Salt Excretors versus Non-Excretors
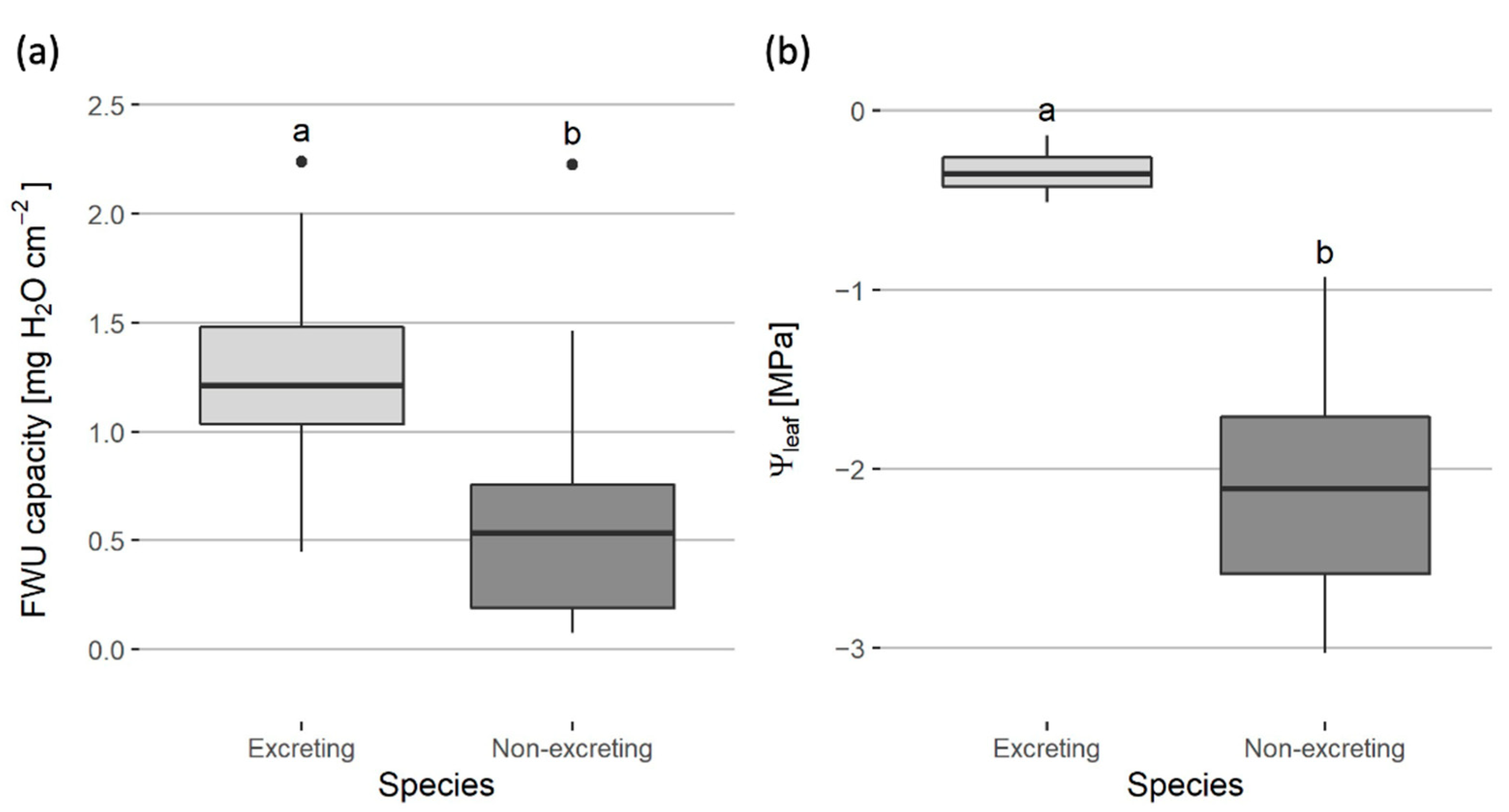
As exclusive classification of the mangrove species into the three putative salt management strategies is debated, the results were instead analyzed by dividing species based on the presence or absence of salt glands, referred to as salt ‘excretors’ and ‘non-excretors’, respectively (Figure 4). The excretors consist of the species A. marina and A. corniculatum, the non-excretors of the species B. cylindrica, B. gymnorhiza, S. alba and S. caseolaris. The FWU capacity of the excretors (1.20 ± 0.50 mg H2O cm−2) is significantly higher than that of the non-excretors (0.41 ± 0.52 mg H2O cm−2) (p < 10−4).
4. Discussion
The FWU capacities measured in this study can be used to compare the relative importance of FWU between the different studied mangrove species. It is worth noting that the obtained FWU capacities by using cut leaves may be an underestimation of FWU in intact trees, as water absorbed by attached leaves will be redistributed toward the stem and possibly the roots and can, therefore, in reality, be orders of magnitude larger [16,17].
For water to flow passively into a leaf, Ψleaf must be more negative than the water immediately surrounding the leaf. During a leaf wetting event, the water potential of the wet surface layer approaches zero [19,20], making it possible for water to enter the leaf by one of the entering pathways. Before the start of the FWU capacity submersion experiment, the Ψleaf of all species was negative (Figure 2b), creating the required gradient for FWU when the leaves were submerged in distilled water (free available water—zero water potential). A. marina and A. corniculatum showed a higher FWU capacity than Bruguiera spp., while they had the least negative water potential (Figure 2a,b). This co-occurrence of the highest FWU capacity and least negative water potential can be explained by the storage of the leaves in moist Ziplock bags prior to measurement, causing leaf water uptake to occur already during transport (see discussion below). These results also demonstrate that it is sufficient for a water potential gradient to exist between a submerged leaf and the surrounding water and that its magnitude is of less importance to determine the FWU capacity as long as certain anatomical characteristics enable water to enter the leaf.
Hayes et al. [59] also performed the submersion experiment of Limm et al. [14] on three neotropical mangrove species, with two salt excretors (Avicennia germinans (L.) L. and Laguncularia racemosa (L.) C. F. Gaertn. and one non-excretor (Rhizophora mangle L.). They found that L. racemosa had the highest FWU capacity and R. mangle the lowest, with 10.99 ± 0.98 mg H2O cm−2 and 6.96 ± 1.16 mg H2O cm−2, respectively. Although these values are approximately 10 times larger than our values, it confirms that FWU capacity varies across mangrove species and, more importantly, that non-excretors have a lower FWU capacity than excretors (Figure 4).
Interestingly, of all species, A. marina not only had the highest FWU capacity but was also the only species showing an abundant number of trichomes (Figure 3b). This distinctive leaf functional trait, accompanied by a high FWU capacity, raises the question of whether it has a role in FWU for this particular mangrove species. Previous studies indicate that these trichomes act as extracellular storage units for water and are the first to be applicable during a persistent drought in A. marina [60]. Besides buffering water losses from underlying tissue due to transpiration, trichomes are involved in the uptake of water and replenishment of leaf water [61]. Up to 10% of a leaf’s total relative water content can be attributed to the storage of water in trichomes and consequently play an important role in the water budget of A. marina [60]. Access to an alternative water source, e.g., dew or intercepting rainfall, was even required to obtain a fully hydrated leaf water status [62]. This process of foliar absorption when the atmospheric humidity is high is aided by the re-absorption of water through aquaporins when salt glands excrete ions [63]. These strategies using water from other sources, rather than sediment water alone [20], explain the vast area where A. marina can thrive, even under high sediment water salinities. Future research should examine the chemical properties of trichomes in A. marina and how these are affected by environmental factors, e.g., atmospheric temperature and sediment salinity. Caution should be taken when attributing the highest FWU capacity of A. marina solely due to the presence of leaf trichomes, as other water entry points may also be involved in the uptake of water. Recently, a new point of water entry has been elucidated, i.e., cork warts in both S. alba and S. caseolaris [25]. Although this leaf trait was also found in B. gymnorhiza by Evans et al. [58], no comparable mode of action for this mechanism has been allocated to this species yet, nor were cork warts observed in our study (Figure 3). A possible reason for the absence of cork warts is that leaves are organs showing high plasticity due to sensitivity to variations in environmental parameters [64]. A case study of Khan et al. [65] highlighted this for Heritiera fomes Buch.-Ham., a mangrove species abundantly found in the Sundarbans, Bangladesh. On three sampling sites with different salinities (the oligohaline, mesohaline and polyhaline zones), this mangrove species showed variations in both leaf anatomical and morphological features. As the Sundarbans’ salinity conditions are largely determined by the volume of freshwater coming from upstream, both the daily and seasonal (dry or wet season) salinity fluctuations determine the environment in which these trees grow [66]. Given that the salinity levels play a major role in determining the leaf plasticity in mangroves [67,68], the leaf anatomical traits from our study (i.e., the absence of cork warts) can differ from those reported in the study of Evans et al. [58] and Bryant et al. [25], which were both carried out in Queensland, Australia.
Regarding leaf water potential measurements, our results are different from previous studies, which led us to speculate on the reasons why. Jiang et al. [69] reported a Ψleaf of −2.68 ± 0.23 MPa and −2.16 ± 0.07 MPa for, respectively, A. corniculatum and A. marina, while our study showed values of −0.35 ± 0.12 MPa and −0.31 ± 0.13 MPa, respectively. In contrast, for B. gymnorhiza, our value was more negative than the one reported by Jiang et al. [69]: −2.70 ± 0.27 MPa versus −2.13 ± 0.09 MPa, respectively. The difference in Ψleaf values between our study and the study of Jiang et al. [69] could possibly be explained by a difference in the sediment salinity between the two study sites of India and China. Zhu et al. [70] indeed showed that, by increasing the sediment salinity from 0 mmol NaCl l−1 H2O to 600 mmol NaCl l−1 H2O, the Ψleaf of A. marina saplings decreased from −1.69 ± 0.11 MPa to −3.38 ± 0.19 MPa. However, if the difference in Ψleaf values between Jiang et al. [69] and our study was due to a difference in salinity, it is not logical that only the values for B. gymnorhiza corresponded, as Jiang et al. [69] collected all the discussed species at the same location, resulting in a similar sediment salinity for all the species.
A more plausible explanation for the observed differences in Ψleaf between Jiang et al.’s study and our work is that they [69] measured the water potential in the field, almost directly after picking the leaf from the tree. They left the leaves in moist bags for only a short period. In our study, the water potential could only be measured in the field station upon return, after a period of two to four hours, with the exception of a maximum of seven hours. During this time span, the leaves were kept in a dark environment in Ziplock bags containing exhaled air and moistened paper tissue to prevent tissue dehydration during transport. In other words, the leaves were kept in a humid environment for several hours, which created an environment that enabled FWU. Additionally, the leaves in the bag were still connected to the node, making water uptake through tissues other than the leaf surface (e.g., petiole) possible. As the Ψleaf values measured in the field station prior to submergence were negative for all species (Figure 2b), the required water potential gradient developed between the leaf and surrounding water, enabling the uptake of water via the leaf and, hence, accurate quantification of FWU capacity. Storage in the moist bags did therefore not invalidate our comparison of the FWU capacity between species. The measured Ψleaf values may, however, not be considered as representative of the Ψleaf values of leaves attached to a tree in the field because of the possible increase in leaf water potential due to FWU inside the bags. Our results demonstrated that this increase is proportional to the species’ FWU capacity (Figure 2 and Figure 4). To verify whether leaves can indeed take up water when stored in moist bags, leaves of A. marina were collected from the botanical garden of Ghent University, Ghent, Belgium (51°02′ N, 3°43′ E), and measured in the same way as described above. Both the leaves that were bagged for one hour and those bagged for four hours showed FWU during the submersion experiment after the bagged period, with a tendency in the four-hour bagged leaves to take up slightly more water than the one-hour bagged leaves (p = 0.212, Figure S3a). The water potential of leaves that were bagged for four hours (−0.71 ± 0.21 MPa) was less negative than one-hour bagged leaves (−1.27 ± 0.26 MPa, p < 10−6; Figure S3b), indicating that they rehydrated during their time in the bags, which corroborates our field observations (Figure 2b).
Our Ψleaf results show that certain species alter their Ψleaf when stored in bags containing moist paper tissue and that this effect is associated with the species’ FWU capacity. With its highest FWU capacity, A. marina took up the most water in the moist bag, which brought its water potential closest to zero just before the start of the submersion experiment (Figure 2b). B. gymnorhiza, which had the smallest measured FWU capacity, took up less water in the moist bag, changing its water potential only slightly, which explained its good correspondence with the values measured by Jiang et al. [69].
The recent review paper of Rodriguez-Dominguez et al. [71] provided best practices for pressure chamber Ψleaf measurements and recommended the storage of leaves in a plastic Ziplock bag (containing exhaled air) immediately after excision. This initial bag is put into a second bag containing moist paper tissue to prevent leaf dehydration. They do note that care should be taken to prevent the absorption of moisture or rehydration, although the studies mentioned in their review did not observe any changes in Ψleaf after the bagged period. In contrast, our study clearly shows that some species do significantly alter their Ψleaf by FWU during storage. We therefore strongly encourage taking the possible effect of FWU into account when the purpose of the study is to precisely and accurately measure the Ψleaf.
As quantification of the FWU capacity was not hampered by bagging the leaves (see the discussion above), we can state that the foliar uptake capacity of freshwater differed between the studied mangroves. Considering the mangrove’s saline sediment environment, FWU might be even more important compared to other forest systems. This research contributes to advances in our knowledge about a species-dependent affinity for FWU in mangrove ecosystems. As potential changes in the non-saline water availability will affect the productivity and persistence of mangroves [72], a higher capacity to take up freshwater through the canopy could give certain species the possibility of colonizing a broader range of salt concentrations. A key factor that determines the future dispersion of mangroves is their resilience to the rise of the seawater level along the coastline habitat, together with the rate of sediment accretion [73]. Especially low-lying islands and river deltas are prone to this eustatic change in the sea level, forcing mangrove species land inward to keep up with the pace of the sea rise or to go locally into extinction [74,75]. The migration of mangrove species farther inland is enhanced in regions where rainfall decreases and saline intrusion takes place [76]. The complex structure of mangroves and associated interaction with their surroundings derives from residing in the continent–ocean demarcation [77]. This makes the prediction of flourishment, decline or extinction of mangrove ecosystems in the nearby future difficult, because climate effects widely vary at the local scale. Nevertheless, a thorough assessment of coastal mangrove distribution on a global scale for 30 species has been modeled for the year 2080 [78]. Two of our studied species, i.e., A. marina and S. alba, were indicated as two out of twelve globally most common species and were included in the distribution model. Both species are projected to gain coastal area [78]. Although this approximation of the distribution of mangroves using occurrence records and environmental data is valuable, additional data on the biotic processes (e.g., interspecific competition and propagule dispersal) and landscape development (e.g., coastal area, forestry and forest management) could further improve the model’s accuracy [79,80]. FWU capacity can be one of these important model additions, as it co-determines the fate of mangrove survival under altering precipitation patterns. Binks et al. [81] further showed that FWU, besides contributing to the plant’s water budget, also resulted in photosynthesis afforded by this extra freshwater input. Our findings could therefore be useful in predictions about the gross primary production yield of mangrove forests, regarding differences in FWU between mangrove species.
5. Conclusions
We demonstrated that there is a significant difference in FWU capacity between mangrove species thriving in the same area and under the same environmental conditions. Where A. marina has the highest FWU capacity, B. gymnorhiza has the lowest. Salt-excreting species showed a higher FWU capacity than non-excreting species. The highest FWU capacity of A. marina coincided with the presence of trichomes on the abaxial leaf side, which were not present in the five other mangrove species. Future research should therefore include leaf traits to discover the water entering pathways and should focus on the species-specific importance of FWU to better address potential natural vegetation shifts in mangrove forests under climate change. Furthermore, we conclude that leaf storage in moist bags for a couple of hours prior to the submersion experiment must be considered as a leaf treatment itself, during which FWU may occur. While bagging the leaves did not hamper the quantification of FWU capacity (i.e., as long as a water potential gradient exists between the submerged leaf and surrounding water), it did affect Ψleaf measurements and increased the values in concert with the FWU capacity. Future studies should therefore consider that the storage of leaves in moist bags prior to measurement could alter the Ψleaf.
Supplementary Materials
The following supporting information can be downloaded at https://0-www-mdpi-com.brum.beds.ac.uk/article/10.3390/f13060951/s1: Figure S1: Satellite map of Ashtamudi Lake. Figure S2: Differences in leaf water contents (ΔLWC; %). Figure S3: Additional submersion experiment on the leaves of Avicennia marina.
Author Contributions
Conceptualization, A.N., S.K.A. and K.S.; Resources, A.N., S.K.A. and K.S.; Investigation, K.H.D.S., W.G. and A.N.; Visualization, K.H.D.S. and W.G.; Formal analysis, K.H.D.S., W.G. and K.S.; Writing—original draft preparation, K.H.D.S., W.G. and K.S.; Writing—review and editing, K.H.D.S., W.G. and K.S.; Supervision, K.S.; Project administration, S.K.A. and K.S. and Funding acquisition, K.S. All authors have read and agreed to the published version of the manuscript.
Funding
Funding was provided by the Special Research Fund (BOF) of Ghent University, Belgium, for Research Project 01J07919 granted to K.S. and supporting the PhD work of K.H.D.S. A travel grant was provided to K.H.D.S. by the Commission Scientific Research (CWO) of Ghent University, Belgium, allowing the research stay at the Kerala Forest Research Institute, Kerala, India. The University Grants Commission supported A.N. with a Junior Research Fellowship (UGC JRF-375066) for his research in mangrove ecophysiology, and S.K.A. was supported by Kerala Forest Research Institute Plan Grant RP 775/19 to fund the fieldwork in mangroves and host K.H.D.S. in India.
Data Availability Statement
The data that support this study will be shared upon reasonable request to the corresponding author.
Acknowledgments
The authors thank Olivier Leroux for the opportunity to access the botanical garden at Ghent University and for his anatomical knowledge, Roberto L. Salomón for his help with statistics, Jeroen Schreel for his guidelines on the experimental set-up and Kausal A.K. and Peter Pathrose for the help during fieldwork. K.H.D.S. acknowledges the Kerala Forest Research Institute for hosting her as an international visiting research student.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Spalding, M.; Kainuma, M.; Collins, L. World Atlas of Mangroves; Earthscan: London, UK, 2010. [Google Scholar]
- Bunting, P.; Rosenqvist, A.; Lucas, R.M.; Rebelo, L.M.; Hilarides, L.; Thomas, N.; Hardy, A.; Itoh, T.; Shimada, M.; Finlayson, C.M. The Global Mangrove Watch—A New 2010 Global Baseline of Mangrove Extent. Remote Sens. 2018, 10, 1669. [Google Scholar] [CrossRef] [Green Version]
- IPCC. Special Report: Global Warming of 1.5 °C; IPCC: Geneva, Switzerland, 2018.
- Gilman, E.L.; Ellison, J.; Duke, N.C.; Field, C. Threats to Mangroves from Climate Change and Adaptation Options: A Review. Aquat. Bot. 2008, 89, 237–250. [Google Scholar] [CrossRef]
- Duke, N.C.; Field, C.; Mackenzie, J.R.; Meynecke, J.-O.; Wood, A.L.; Duke, N.C.; Field, C.; Mackenzie, J.R.; Meynecke, J.-O.; Wood, A.L. Rainfall and Its Possible Hysteresis Effect on the Proportional Cover of Tropical Tidal-Wetland Mangroves and Saltmarsh–Saltpans. Mar. Freshw. Res. 2019, 70, 1047–1055. [Google Scholar] [CrossRef]
- Ward, R.D.; Friess, D.A.; Day, R.H.; Mackenzie, R.A. Impacts of Climate Change on Mangrove Ecosystems: A Region by Region Overview. Ecosyst. Health Sustain. 2016, 2, e01211. [Google Scholar] [CrossRef] [Green Version]
- Field, C.D. Impact of Expected Climate Change on Mangroves. In Asia-Pacific Symposium on Mangrove Ecosystems; Springer: Dordrecht, The Netherlands, 1995; pp. 75–81. [Google Scholar] [CrossRef]
- Duke, N.C.; Ball, M.C.; Ellison, J.C. Factors Influencing Biodiversity and Distributional Gradients in Mangroves. Glob. Ecol. Biogeogr. Lett. 1998, 7, 27–47. [Google Scholar] [CrossRef] [Green Version]
- Dixon, H.H.; Joly, J. On the Ascent of Sap. Philos. Trans. R. Soc. Lond. 1895, 186, 563–576. [Google Scholar] [CrossRef]
- Nobel, P.S. Physicochemical and Environmental Plant Physiology, 4th ed.; Academic Press: Cambridge, MA, USA, 2009. [Google Scholar]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Goldsmith, G.R. Changing Directions: The Atmosphere-Plant-Soil Continuum. New Phytol. 2013, 199, 4–6. [Google Scholar] [CrossRef]
- Schreel, J.D.M.; Steppe, K. Foliar Water Uptake in Trees: Negligible or Necessary? Trends Plant Sci. 2020, 25, 590–603. [Google Scholar] [CrossRef]
- Limm, E.B.; Simonin, K.A.; Bothman, A.G.; Dawson, T.E. Foliar Water Uptake: A Common Water Acquisition Strategy for Plants of the Redwood Forest. Oecologia 2009, 161, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Burgess, S.; Dawson, T. The Contribution of Fog to the Water Relations of Sequoia sempervirens (D. Don): Foliar Uptake and Prevention of Dehydration. Plant. Cell Environ. 2004, 27, 1023–1034. [Google Scholar] [CrossRef]
- Nadezhdina, N.; David, T.; David, J.; Ferreira, M.; Dohnal, M.; Tesar, M.; Gartner, K.; Leitgeb, E.; Nadezhdin, V.; Cermák, J.; et al. Trees Never Rest: The Multiple Facets of Hydraulic Redistribution. Ecohydrology 2010, 3, 431–444. [Google Scholar] [CrossRef]
- Schreel, J.D.M.; Van de Wal, B.A.E.; Hervé-Fernandez, P.; Boeckx, P.; Steppe, K. Hydraulic Redistribution of Foliar Absorbed Water Causes Turgor-Driven Growth in Mangrove Seedlings. Plant Cell Environ. 2019, 42, 2437–2447. [Google Scholar] [CrossRef]
- Breazeale, E.L.; McGeorge, W.T.; Breazeale, J.F. Moisture Absorption from Plants from an Atmosphere of High Humidity. Plant Physiol. 1950, 25, 413–419. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berry, Z.C.; Emery, N.C.; Gotsch, S.G.; Goldsmith, G.R. Foliar Water Uptake: Processes, Pathways, and Integration into Plant Water Budgets. Plant Cell Environ. 2019, 42, 410–423. [Google Scholar] [CrossRef] [Green Version]
- Steppe, K.; Vandegehuchte, M.W.; Van de Wal, B.A.E.; Hoste, P.; Guyot, A.; Lovelock, C.E.; Lockington, D.A. Direct Uptake of Canopy Rainwater Causes Turgor-Driven Growth Spurts in the Mangrove Avicennia marina. Tree Physiol. 2018, 38, 979–991. [Google Scholar] [CrossRef] [Green Version]
- Simonin, K.A.; Santiago, L.S.; Dawson, T.E. Fog Interception by Sequoia sempervirens (D. Don) Crowns Decouples Physiology from Soil Water Deficit. Plant Cell Environ. 2009, 32, 882–892. [Google Scholar] [CrossRef]
- Duke, N.C.; Kovacs, J.M.; Griffiths, A.D.; Preece, L.; Hill, D.J.E.; Van Oosterzee, P.; Mackenzie, J.; Morning, H.S.; Burrows, D. Large-Scale Dieback of Mangroves in Australia’s Gulf of Carpentaria: A Severe Ecosystem Response, Coincidental with an Unusually Extreme Weather Event. Mar. Freshw. Res. 2017, 68, 1816–1829. [Google Scholar] [CrossRef]
- Schreel, J.D.M.; Leroux, O.; Goossens, W.; Brodersen, C.; Rubinstein, A.; Steppe, K. Identifying the Pathways for Foliar Water Uptake in Beech (Fagus sylvatica L.): A Major Role for Trichomes. Plant J. 2020, 103, 769–780. [Google Scholar] [CrossRef]
- Guzmán-Delgado, P.; Laca, E.; Zwieniecki, M.A. Unravelling Foliar Water Uptake Pathways: The Contribution of Stomata and the Cuticle. Plant Cell Environ. 2021, 44, 1728–1740. [Google Scholar] [CrossRef]
- Bryant, C.; Fuenzalida, T.I.; Zavafer, A.; Nguyen, H.T.; Brothers, N.; Harris, R.J.; Beckett, H.A.A.; Holmlund, H.I.; Binks, O.; Ball, M.C. Foliar Water Uptake via Cork Warts in Mangroves of the Sonneratia Genus. Plant Cell Environ. 2021, 44, 2925–2937. [Google Scholar] [CrossRef]
- Schönherr, J.; Bukovac, M.J. Penetration of Stomata by Liquids: Dependence on Surface Tension, Wettability, and Stomatal Morphology. Plant Physiol. 1972, 49, 813–819. [Google Scholar] [CrossRef] [PubMed]
- Burkhardt, J.; Basi, S.; Pariyar, S.; Hunsche, M. Stomatal Penetration by Aqueous Solutions—An Update Involving Leaf Surface Particles. New Phytol. 2012, 196, 774–787. [Google Scholar] [CrossRef]
- Coopman, R.E.; Nguyen, H.T.; Mencuccini, M.; Oliveira, R.S.; Sack, L.; Lovelock, C.E.; Ball, M.C. Harvesting Water from Unsaturated Atmospheres: Deliquescence of Salt Secreted onto Leaf Surfaces Drives Reverse Sap Flow in a Dominant Arid Climate Mangrove, Avicennia marina. New Phytol. 2021, 231, 1401–1414. [Google Scholar] [CrossRef] [PubMed]
- Martin, C.E.; von Willert, D.J. Leaf Epidermal Hydathodes and the Ecophysiological Consequences of Foliar Water Uptake in Species of Crassula from the Namib Desert in Southern Africa. Plant Biol. 2000, 2, 229–242. [Google Scholar] [CrossRef]
- Fernández, V.; Bahamonde, H.A.; Peguero-Pina, J.J.; Gil-Pelegrín, E.; Sancho-Knapik, D.; Gil, L.; Goldbach, H.E.; Eichert, T. Physico-Chemical Properties of Plant Cuticles and Their Functional and Ecological Significance. J. Exp. Bot. 2017, 68, 5293–5306. [Google Scholar] [CrossRef]
- Schreiber, L.; Hartmann, K.; Skrabs, M.; Zeier, J. Apoplastic Barriers in Roots: Chemical Composition of Endodermal and Hypodermal Cell Walls on JSTOR. J. Exp. Bot. 1999, 50, 1267–1280. [Google Scholar] [CrossRef] [Green Version]
- Boanares, D.; Ferreira, B.G.; Kozovits, A.R.; Sousa, H.C.; Isaias, R.M.S.; França, M.G.C. Pectin and Cellulose Cell Wall Composition Enables Different Strategies to Leaf Water Uptake in Plants from Tropical Fog Mountain. Plant Physiol. Biochem. 2018, 122, 57–64. [Google Scholar] [CrossRef]
- Kamtsikakis, A.; Baales, J.; Zeisler-Diehl, V.V.; Vanhecke, D.; Zoppe, J.O.; Schreiber, L.; Weder, C. Asymmetric Water Transport in Dense Leaf Cuticles and Cuticle-Inspired Compositionally Graded Membranes. Nat. Commun. 2021, 12, 1267. [Google Scholar] [CrossRef]
- Scholander, P.F. How Mangroves Desalinate Seawater. Physiol. Plant. 1968, 21, 251–261. [Google Scholar] [CrossRef]
- Tomlinson, P.B. The Botany of Mangroves; Cambridge University Press: Cambridge, UK, 1986. [Google Scholar]
- Parida, A.; Jha, B. Salt Tolerance Mechanisms in Mangroves: A Review. Trees–Struct. Funct. 2010, 24, 199–217. [Google Scholar] [CrossRef]
- Ball, M.C. Ecophysiology of Mangroves. Trees 1988, 2, 129–142. [Google Scholar] [CrossRef]
- Krishnamurthy, P.; Jyothi-Prakash, P.A.; Qin, L.; He, J.; Lin, Q.; Loh, C.S.; Kumar, P.P. Role of Root Hydrophobic Barriers in Salt Exclusion of a Mangrove Plant Avicennia Officinalis. Plant Cell Environ. 2014, 37, 1656–1671. [Google Scholar] [CrossRef]
- Antony, M.M.; Ignatius, J. A Hydrological Study of Ashtamudi Lake, Kerala, India with Special Reference to Its Ecological Difference. Int. J. Sci. Res. 2016, 5, 1841–1846. [Google Scholar]
- Osborn, T.J.; Jones, P.D. The CRUTEM4 Land-Surface Air Temperature Data Set: Construction, Previous Versions and Dissemination via Google Earth. Earth Syst. Sci. Data 2014, 6, 61–68. [Google Scholar] [CrossRef] [Green Version]
- India Meteorological Department. Customized Rainfall Information System (CRIS); India Meteorological Department: New Delhi, India, 2020.
- Devassy, V.P.; Gopinathan, C.K. Hydrobiological Features of the Kerala Backwaters during Premonsoon and Monsoon Months. Fish. Technol. 1970, 7, 190–194. [Google Scholar]
- Ball, M.C. Salinity Tolerance in the Mangroves Aegiceras Corniculatum and Avicennia marina. I. Water Use in Relation to Growth, Carbon Partitioning, and Salt Balance. Funct. Plant Biol. 1988, 15, 447–464. [Google Scholar] [CrossRef]
- Cram, J.W.; Torr, P.G.; Rose, D.A. Salt Allocation during Leaf Development and Leaf Fall in Mangroves. Trees 2002, 16, 112–119. [Google Scholar] [CrossRef]
- Yasumoto, E.; Adachi, K.; Kato, M.; Sano, H.; Sasamoto, H.; Baba, S.; Ashihara, H. Uptake of Inorganic Ions and Compatible Solutes in Cultured Mangrove Cells during Salt Stress; Society for In Vitro Biology: Raleigh, NC, USA, 1999. [Google Scholar]
- Kura-Hotta, M.; Mimura, M.; Tsujimura, T.; Washitani-Nemoto, S.; Mimura, T. High Salt-Treatment-Induced Na+ Extrusion and Low Salt-Treatment-Induced Na+ Accumulation in Suspension-Cultured Cells of the Mangrove Plant, Bruguiera sexangula. Plant Cell Environ. 2001, 24, 1105–1112. [Google Scholar] [CrossRef]
- Lawton, J.R.; Todd, A.; Naidoo, D.K. Preliminary Investigations into the Structure of the Roots of the Mangroves, Avicennia marina and Bruguiera Gymnorrhiza, in Relation to Ion Uptake; Wiley: New York, NY, USA, 1981. [Google Scholar]
- Reef, R.; Lovelock, C.E. Regulation of Water Balance in Mangroves. Ann. Bot. 2014, 115, 385–395. [Google Scholar] [CrossRef] [Green Version]
- Lovelock, C. Field Guide to the Mangroves of Queensland; Australian Institute of Marine Science: Townsville, Australia, 1993.
- Yeung, E.C.T.; Stasolla, C.; Sumner, M.J.; Huang, B.Q. Plant Microtechniques and Protocols; Springer International Publishing: Berlin/Heidelberg, Germany, 2015. [Google Scholar] [CrossRef]
- Johansen, D.A. Plant Microtechnique; McGraw-Hill: New York, NY, USA, 1940. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2020; Available online: http://www.R-project.org (accessed on 1 May 2022).
- RStudio. RStudio: Integrated Development Environment for R; RStudio: Boston, MA, USA, 2021; Available online: https//www.rstudio.com (accessed on 1 May 2022).
- Hothorn, T.; Bretz, F.; Westfall, P. Simultaneous Inference in General Parametric Models. Biom. J. 2008, 50, 346–363. [Google Scholar] [CrossRef] [Green Version]
- Fox, J.; Weisberg, S. An R Companion to Applied Regression, 3rd ed.; Sage Publications: Thousand Oaks, CA, USA, 2019; Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 1 May 2022).
- Burns, J.H.; Strauss, S.Y. More Closely Related Species Are More Ecologically Similar in an Experimental Test. Proc. Natl. Acad. Sci. USA 2011, 108, 5302–5307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ackerly, D. Conservatism and Diversification of Plant Functional Traits: Evolutionary Rates versus Phylogenetic Signal. Proc. Natl. Acad. Sci. USA 2009, 106 (Suppl. S2), 19699–19706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Evans, L.S.; Testo, Z.M.; Cerutti, J.A. Characterization of Internal Airflow within Tissues of Mangrove Species from Australia: Leaf Pressurization Processes. J. Torrey Bot. Soc. 2009, 136, 70–83. [Google Scholar] [CrossRef]
- Hayes, M.A.; Chapman, S.; Jesse, A.; O’Brien, E.; Langley, J.A.; Bardou, R.; Devaney, J.; Parker, J.D.; Cavanaugh, K.C. Foliar Water Uptake by Coastal Wetland Plants: A Novel Water Acquisition Mechanism in Arid and Humid Subtropical Mangroves. J. Ecol. 2020, 108, 2625–2637. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Meir, P.; Wolfe, J.; Mencuccini, M.; Ball, M.C. Plumbing the Depths: Extracellular Water Storage in Specialized Leaf Structures and Its Functional Expression in a Three-Domain Pressure–Volume Relationship. Plant Cell Environ. 2017, 40, 1021–1038. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Constable, L. Nocturnal Top-down Rehydration: Contribution of Atmospheric Moisture to Water Status of the Mangrove, Avicennia marina. Doctoral Dissertation, The Australian National University, Canberra, Australia, 2014. [Google Scholar]
- Nguyen, H.T.; Meir, P.; Sack, L.; Evans, J.R.; Oliveira, R.S.; Ball, M.C. Leaf Water Storage Increases with Salinity and Aridity in the Mangrove Avicennia marina: Integration of Leaf Structure, Osmotic Adjustment and Access to Multiple Water Sources. Plant Cell Environ. 2017, 40, 1576–1591. [Google Scholar] [CrossRef] [Green Version]
- Tan, W.K.; Lin, Q.; Lim, T.M.; Kumar, P.; Loh, C.S. Dynamic Secretion Changes in the Salt Glands of the Mangrove Tree Species Avicennia Officinalis in Response to a Changing Saline Environment. Plant Cell Environ. 2013, 36, 1410–1422. [Google Scholar] [CrossRef]
- Shi, G.; Cai, Q. Leaf plasticity in peanut (Arachis hypogaea L.) in response to heavy metal stress. Env. and Exp. Bot. 2009, 67, 112–117. [Google Scholar] [CrossRef]
- Khan, M.N.I.; Khatun, S.; Azad, M.S.; Mollick, A.S. Leaf morphological and anatomical plasticity in Sundri (Heritiera fomes Buch.-Ham.) along different canopy light and salinity zones in the Sundarbans mangrove forest, Bangladesh. Glob. Ecol. and Cons. 2020, 23, e01127. [Google Scholar] [CrossRef]
- Wahid, S.M.; Babel, M.S.; Bhuiyan, A.R. Hydrologic monitoring and analysis in the Sundarbans mangrove ecosystem, Bangladesh. J. of Hydr. 2007, 332, 381–395. [Google Scholar] [CrossRef]
- Hoppe-Speer, S.C.; Adams, J.B.; Rajkaran, A.; Bailey, D. The response of the red mangrove Rhizophora mucronata Lam. to salinity and inundation in South Africa. Aquat. Bot. 2011, 95, 71–76. [Google Scholar] [CrossRef]
- Vovides, A.G.; Vogt, J.; Kollert, A.; Berger, U.; Grueters, U.; Peters, R.; Lara-Domínguez, A.L.; López-Portillo, J. Morphological plasticity in mangrove trees: Salinity-related changes in the allometry of Avicennia germinans. Trees 2014, 28, 1413–1425. [Google Scholar] [CrossRef]
- Jiang, G.-F.; Goodale, U.M.; Liu, Y.-Y.; Hao, G.-Y.; Cao, K.-F. Salt Management Strategy Defines the Stem and Leaf Hydraulic Characteristics of Six Mangrove Tree Species. Tree Physiol. 2017, 37, 389–401. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Z.; Zhang, R.; Liu, T.; Zheng, H.-L. Solute Accumulation and Osmotic Adjustment Characteristics of the Mangrove Avicennia marina under NaCl-Induced Salinity Stress. Bot. Mar. 2011, 54, 335–341. [Google Scholar] [CrossRef]
- Rodriguez-Dominguez, C.M.; Forner, A.; Martorell, S.; Choat, B.; Lopez, R.; Peters, J.M.R.; Pfautsch, S.; Mayr, S.; Carins-Murphy, M.R.; McAdam, S.A.M.; et al. Leaf Water Potential Measurements Using the Pressure Chamber: Synthetic Testing of Assumptions towards Best Practices for Precision and Accuracy. Plant Cell Environ. 2022. [Google Scholar] [CrossRef]
- Lovelock, C.E.; Feller, I.C.; Reef, R.; Hickey, S.; Ball, M.C. Mangrove Dieback during Fluctuating Sea Levels. Sci. Rep. 2017, 7, 1680. [Google Scholar] [CrossRef]
- McIvor, A.L.; Spencer, T.; Möller, I.; Spalding, M. The Response of Mangrove Soil Surface Elevation to Sea Level Rise. Natural Coastal Protection Series: Report 3, Cambridge Coastal Research Unit Working Paper 42; The Nature Conservancy and Wetlands International,. 2013. Available online: https://www.conservationgateway.org/ConservationPractices/Marine/crr/library/Documents/mangrove-surface-elevation-and-sea-level-rise.pdf (accessed on 1 May 2022).
- Ellison, J.C. Long-term retrospection on mangrove development using sediment cores and pollen analysis: A review. Aquat. Bot. 2008, 89, 93–104. [Google Scholar] [CrossRef]
- Shearman, P.; Bryan, J.; Walsh, J.P. Trends in deltaic change over three decades in the Asia-pacific region. J. Coast Res. 2013, 29, 1169–1183. [Google Scholar] [CrossRef]
- Godoy, M.D.; Lacerda, L.D.D. Mangroves response to climate change: A review of recent findings on mangrove extension and distribution. An. Acad. Bras. Ciênc. 2015, 87, 651–667. [Google Scholar] [CrossRef] [Green Version]
- Alongi, D.M. The impact of climate change on mangrove forests. Curr. Clim. Chang. Rep. 2015, 1, 30–39. [Google Scholar] [CrossRef]
- Record, S.; Charney, N.D.; Zakaria, R.M.; Ellison, A.M. Projecting global mangrove species and community distributions under climate change. Ecosphere 2013, 4, 1–23. [Google Scholar] [CrossRef]
- Clarke, P.J.; Kerrigan, R.A. The effects of seed predators on the recruitment of mangroves. J. Ecol. 2002, 90, 728–736. [Google Scholar] [CrossRef] [Green Version]
- Ellison, A.M. Managing mangroves with benthic biodiversity in mind: Moving beyond roving banditry. J. Sea Res. 2008, 59, 2–15. [Google Scholar] [CrossRef]
- Binks, O.; Mencuccini, M.; Rowland, L.; da Costa, A.C.L.; de Carvalho, C.J.R.; Bittencourt, P.; Eller, C.; Teodoro, G.S.; Carvalho, E.J.M.; Soza, A.; et al. Foliar Water Uptake in Amazonian Trees: Evidence and Consequences. Glob. Chang. Biol. 2019, 25, 2678–2690. [Google Scholar] [CrossRef] [PubMed]
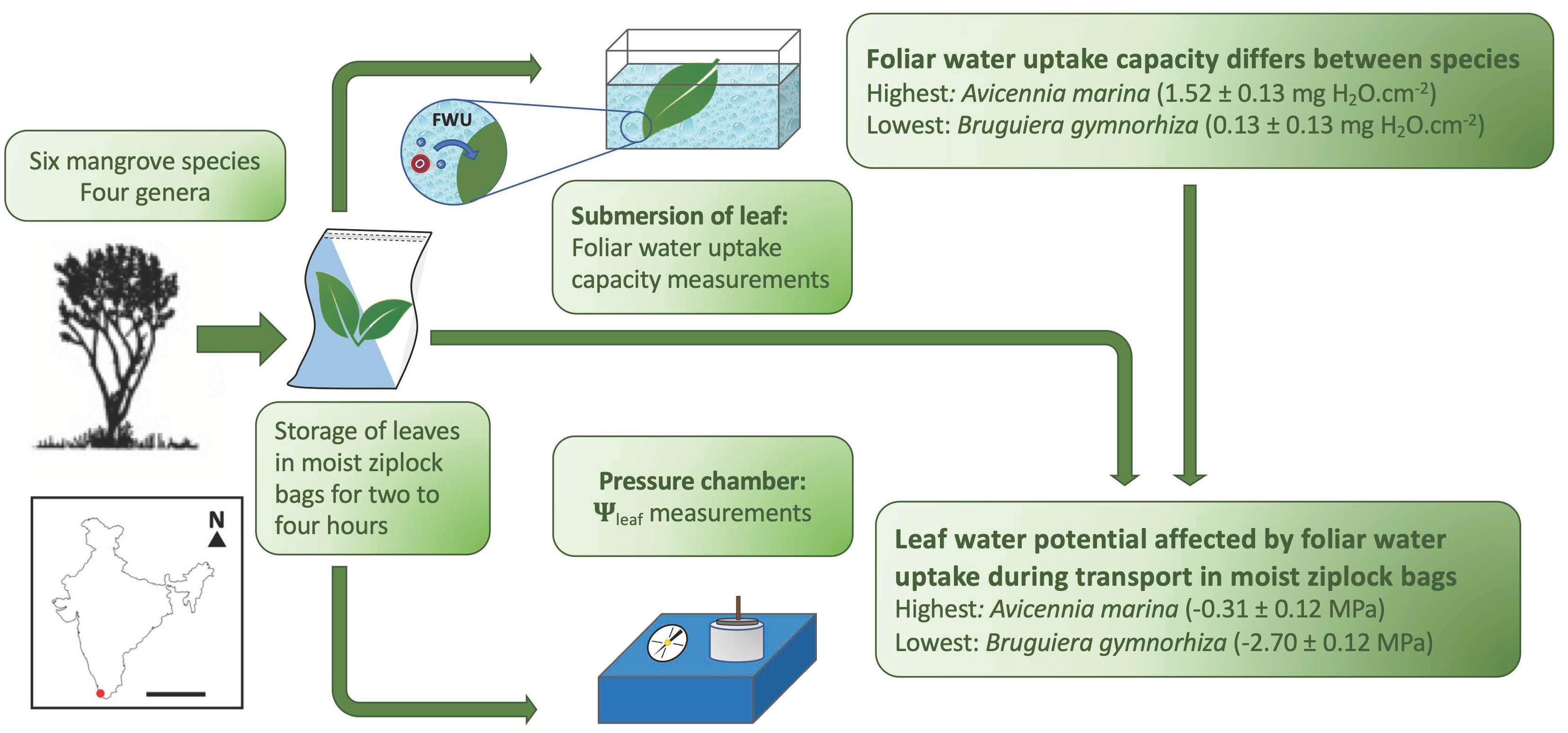
Figure 1.
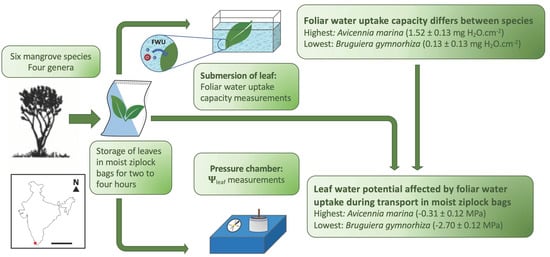
(a) Schematic overview of the leaf sampling of six mangrove species belonging to four different genera: Aegiceras corniculatum (A.c.), Avicennia marina (A.m.), Bruguiera cylindrica (B.c.), Bruguiera gymnorhiza (B.g.), Sonneratia alba (S.a.) and Sonneratia caseolaris (S.c.). From each node, one leaf was used for the foliar water uptake (FWU) capacity experiment, and one leaf was used for the water potential (Ψleaf) measurement with the pressure chamber. The illustrations of the trees are adapted from Lovelock [49] and Spalding et al. [1]. (b) The location of Ashtamudi Lake in India, where the species samples were collected. Scale bar = 1000 km.
Figure 1.
(a) Schematic overview of the leaf sampling of six mangrove species belonging to four different genera: Aegiceras corniculatum (A.c.), Avicennia marina (A.m.), Bruguiera cylindrica (B.c.), Bruguiera gymnorhiza (B.g.), Sonneratia alba (S.a.) and Sonneratia caseolaris (S.c.). From each node, one leaf was used for the foliar water uptake (FWU) capacity experiment, and one leaf was used for the water potential (Ψleaf) measurement with the pressure chamber. The illustrations of the trees are adapted from Lovelock [49] and Spalding et al. [1]. (b) The location of Ashtamudi Lake in India, where the species samples were collected. Scale bar = 1000 km.

Figure 2.
Box plots of the leaf parameters for the six different mangrove species, belonging to four genera, with the following parameters of study: (a) foliar water uptake (FWU) capacity and (b) leaf water potential (Ψleaf). Box plot midlines are medians, and box edges show first and third quartiles, with whiskers representing minimum and maximum values. Circles indicate outliers in the dataset. Letters at the top of the box plots indicate differences of significance (p < 0.05) among groups based on Tukey’s comparisons, for which the data of FWU capacity were log-transformed. Abbreviations: A.c. = Aegiceras corniculatum, A.m. = Avicennia marina, B.c. = Bruguiera cylindrica, B.g. = Bruguiera gymnorhiza, S.a. = Sonneratia alba and S.c. = Sonneratia caseolaris.
Figure 2.
Box plots of the leaf parameters for the six different mangrove species, belonging to four genera, with the following parameters of study: (a) foliar water uptake (FWU) capacity and (b) leaf water potential (Ψleaf). Box plot midlines are medians, and box edges show first and third quartiles, with whiskers representing minimum and maximum values. Circles indicate outliers in the dataset. Letters at the top of the box plots indicate differences of significance (p < 0.05) among groups based on Tukey’s comparisons, for which the data of FWU capacity were log-transformed. Abbreviations: A.c. = Aegiceras corniculatum, A.m. = Avicennia marina, B.c. = Bruguiera cylindrica, B.g. = Bruguiera gymnorhiza, S.a. = Sonneratia alba and S.c. = Sonneratia caseolaris.

Figure 3.
(a–f) Transverse microtome cross-sections through the midvein and adjacent lamina tissue of six different mangrove species’ leaves stained with safranin O–fast green. (b) Inset: awl-shaped trichomes at the abaxial leaf area. Scale bars = 500 μm. Abbreviations: Bac = Basal cell, Bc = Body cell and Tc = Top cell.
Figure 3.
(a–f) Transverse microtome cross-sections through the midvein and adjacent lamina tissue of six different mangrove species’ leaves stained with safranin O–fast green. (b) Inset: awl-shaped trichomes at the abaxial leaf area. Scale bars = 500 μm. Abbreviations: Bac = Basal cell, Bc = Body cell and Tc = Top cell.

Figure 4.
Box plots of the leaf parameters for salt-excreting and non-salt-excreting species with the following parameters of study: (a) foliar water uptake (FWU) capacity and (b) leaf water potential (Ψleaf). The excretors consist of the species Avicennia marina and Aegiceras corniculatum, the non-excretors of the species Bruguiera cylindrica, Bruguiera gymnorhiza, Sonneratia alba and Sonneratia caseolaris. Box plot midlines are medians, and box edges show first and third quartiles, with whiskers representing minimum and maximum values. Circles indicate outliers in the dataset. Letters at the top of the box plots indicate differences of significance (p < 0.05) among groups based on Welch t-tests, for which data of FWU capacity were log-transformed.
Figure 4.
Box plots of the leaf parameters for salt-excreting and non-salt-excreting species with the following parameters of study: (a) foliar water uptake (FWU) capacity and (b) leaf water potential (Ψleaf). The excretors consist of the species Avicennia marina and Aegiceras corniculatum, the non-excretors of the species Bruguiera cylindrica, Bruguiera gymnorhiza, Sonneratia alba and Sonneratia caseolaris. Box plot midlines are medians, and box edges show first and third quartiles, with whiskers representing minimum and maximum values. Circles indicate outliers in the dataset. Letters at the top of the box plots indicate differences of significance (p < 0.05) among groups based on Welch t-tests, for which data of FWU capacity were log-transformed.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Schaepdryver, K.H.D.; Goossens, W.; Naseef, A.; Kalpuzha Ashtamoorthy, S.; Steppe, K. Foliar Water Uptake Capacity in Six Mangrove Species. Forests 2022, 13, 951. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060951
AMA Style
Schaepdryver KHD, Goossens W, Naseef A, Kalpuzha Ashtamoorthy S, Steppe K. Foliar Water Uptake Capacity in Six Mangrove Species. Forests. 2022; 13(6):951. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060951
Chicago/Turabian StyleSchaepdryver, Katrien H. D., Willem Goossens, Abdulla Naseef, Sreejith Kalpuzha Ashtamoorthy, and Kathy Steppe. 2022. "Foliar Water Uptake Capacity in Six Mangrove Species" Forests 13, no. 6: 951. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060951
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.