
Identification and Functional Evaluation of Three Polyubiquitin Promoters from Hevea brasiliensis

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials
2.2. Isolation and Bioinformatic Analysis of Polyubiquitin Genes and Their Promoters in Hevea
2.3. Analysis of the Expressions of Three HbUBI Genes in Hevea Plants
2.4. Vector Construction
2.5. Hevea Protoplasts Isolation and Polyethylene Glycol (PEG)-Mediated Transformation
2.6. Agrobacterium-Mediated Transformation of Hevea Somatic Embryos
3. Results
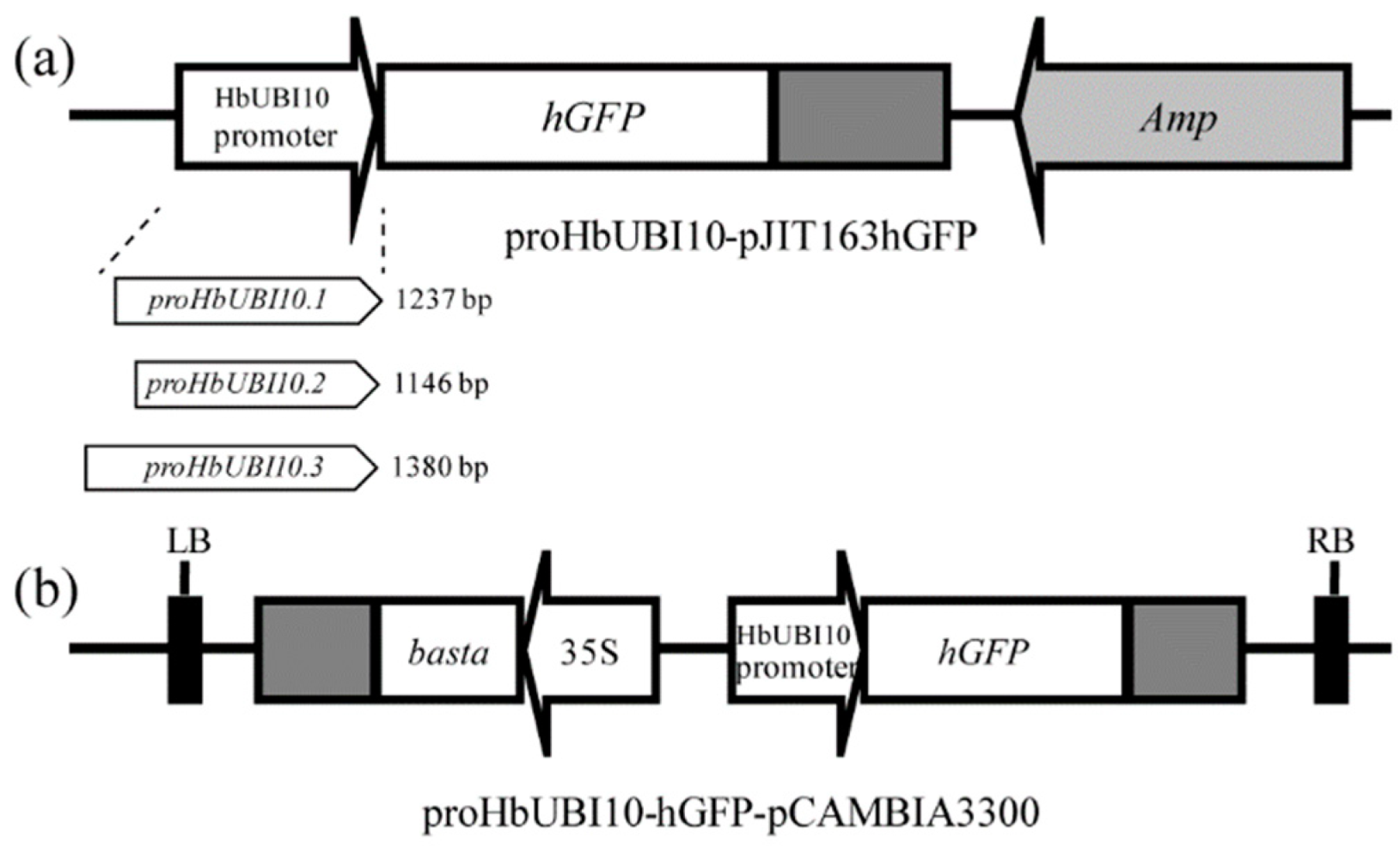
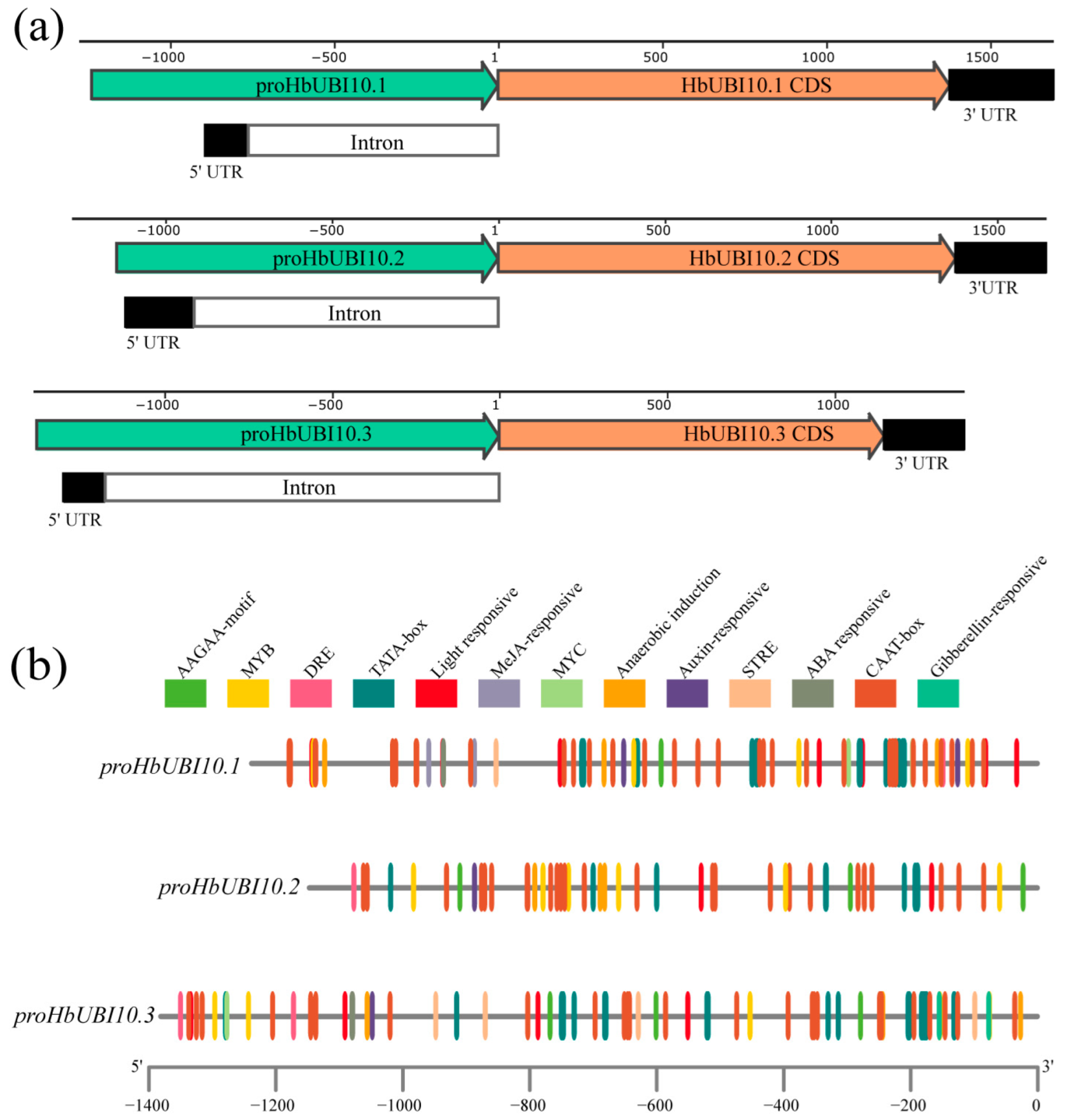
3.1. Isolation of Three Polyubiquitin Promoters in Hevea
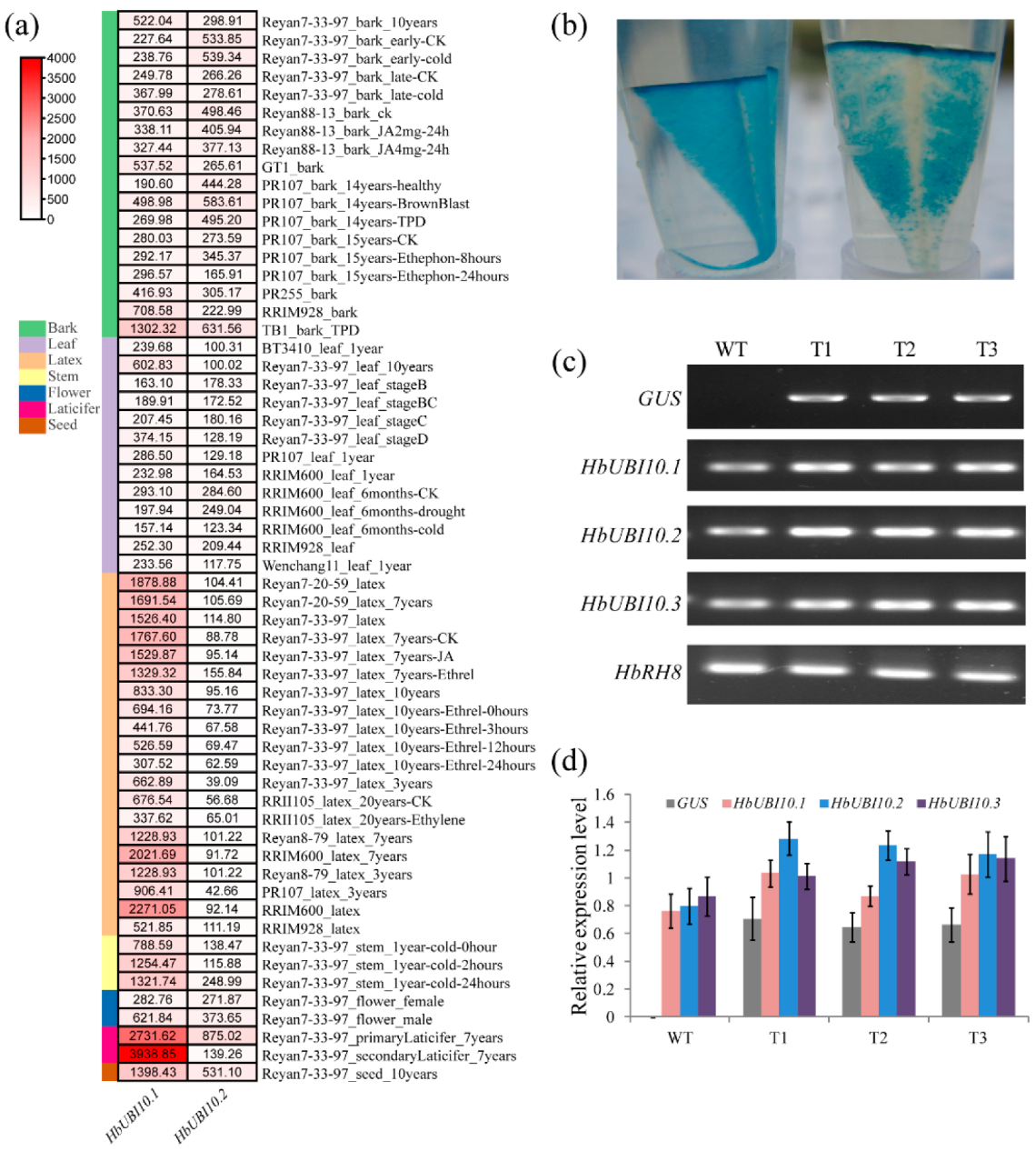
3.2. Expression Analysis of Three HbUBI Genes
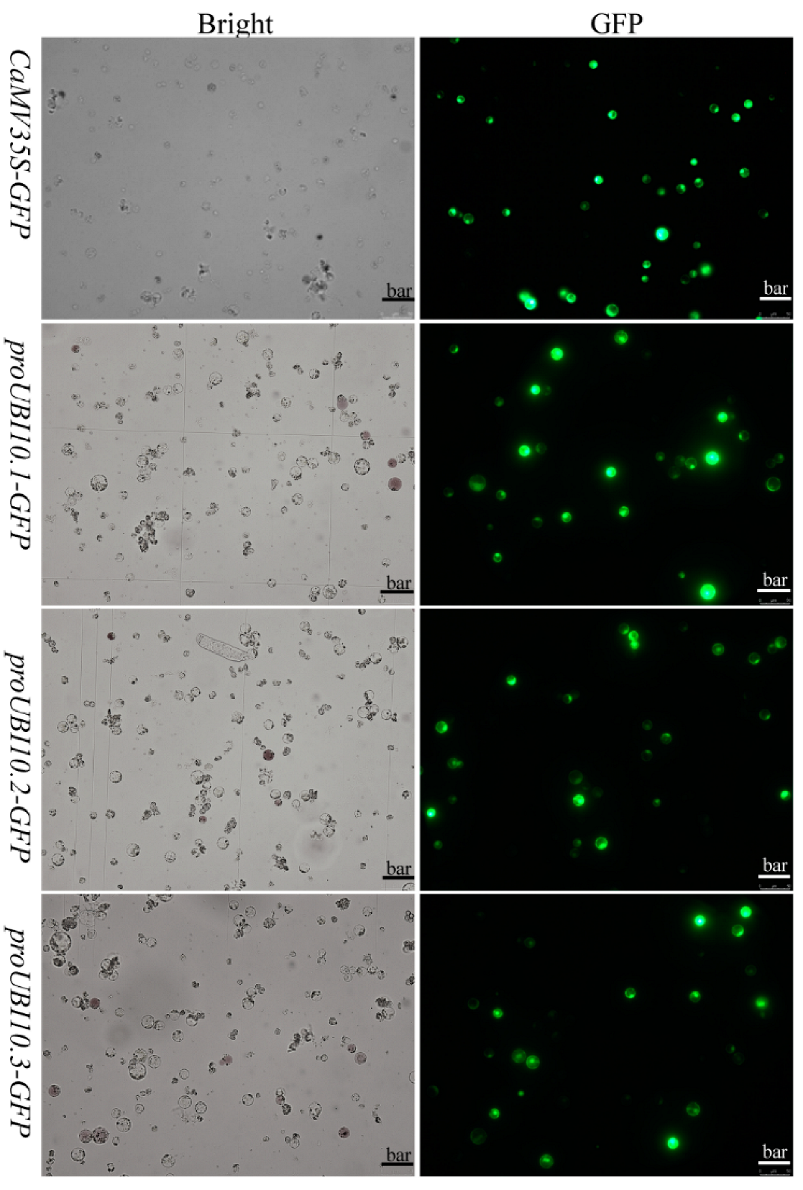
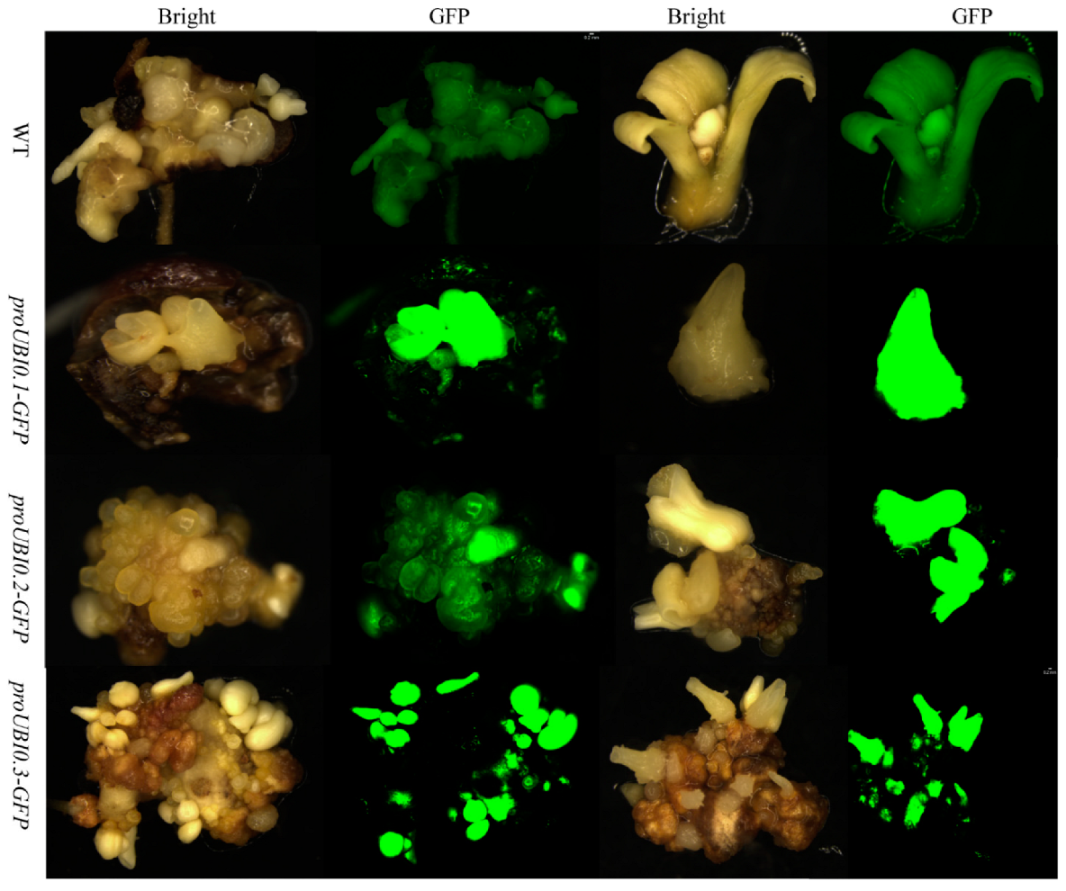
3.3. Functional Evaluation of Three HbUBI Promoters in Hevea Tissues
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thulaseedharan, A.; Jayashree, R.; Kala, R.G.; Rekha, K.; Sobha, S.; Jayasree, P.K.; Kumari, S.S.; Saleena, A.; Venkatachalam, P. Rubber Tree. In Compendium of Transgenic Crop Plants; Kole, C., Hall, T.C., Eds.; Blackwell Publishing Ltd.: Malden, MA, USA, 2009; pp. 117–140. [Google Scholar]
- Arokiaraj, P.; Florian, R.; Obermayr, E.; Samsul, B.A.; Yeang, H.Y. Expression of human serum albumin in transgenic Hevea brasiliensis. J. Rubber Res. 2002, 5, 157–166. [Google Scholar]
- Lestari, R.; Rio, M.; Martin, F.; Leclercq, J.; Woraathasin, N.; Roques, S.; Dessailly, F.; Clément-Vidal, A.; Sanier, C.; Fabre, D.; et al. Overexpression of Hevea brasiliensis ethylene response factor HbERF-IXc5 enhances growth and tolerance to abiotic stress and affects laticifer differentiation. Plant Biotechnol. J. 2018, 16, 322–336. [Google Scholar] [CrossRef] [Green Version]
- Huang, T.D.; Li, J.; Li, Y.T.; Huang, H.S.; Hua, Y.W. Somatic Embryo, an Alternative Target Tissue for Agrobacterium-Mediated Transformation in Hevea brasiliensis. J. Rubber Res. 2015, 18, 171–188. [Google Scholar]
- Martin, F.; Abati, V.; Burel, A.; Clement-Vidal, A.; Sanier, C.; Fabre, D.; Woraathasin, N.; Rio, M.; Besret, P.; Farinas, B.; et al. Overexpression of EcGSH1 induces glutathione production and alters somatic embryogenesis and plant development in Hevea brasiliensis. Ind. Crops Prod. 2018, 112, 803–814. [Google Scholar] [CrossRef]
- Fan, Y.; Xin, S.; Dai, X.; Yang, X.; Huang, H.; Hua, Y. Efficient genome editing of rubber tree (Hevea brasiliensis) protoplasts using CRISPR/Cas9 ribonucleoproteins. Ind. Crops Prod. 2020, 146, 112146. [Google Scholar] [CrossRef]
- Leclercq, J.; Lardet, L.; Martin, F.; Chapuset, T.; Oliver, G.; Montoro, P. The green fluorescent protein as an efficient selection marker for Agrobacterium tumefaciens-mediated transformation in Hevea brasiliensis (Müll. Arg). Plant Cell Rep. 2010, 29, 513–522. [Google Scholar] [CrossRef]
- Huang, T.; Xin, S.; Fang, Y.; Chen, T.; Chang, J.; Ko, N.C.K.; Huang, H.; Hua, Y. Use of a novel R2R3-MYB transcriptional activator of anthocyanin biosynthesis as visual selection marker for rubber tree (Hevea brasiliensis) transformation. Ind. Crops Prod. 2021, 174, 114225. [Google Scholar] [CrossRef]
- Ali, S.; Kim, W.-C. A Fruitful Decade Using Synthetic Promoters in the Improvement of Transgenic Plants. Front. Plant Sci. 2019, 10, 1433. [Google Scholar] [CrossRef]
- Mishiba, K.; Nishihara, M.; Nakatsuka, T.; Abe, Y.; Hirano, H.; Yokoi, T.; Kikuchi, A.; Yamamura, S. Consistent transcriptional silencing of 35S-driven transgenes in gentian. Plant J. 2005, 44, 541–556. [Google Scholar] [CrossRef]
- Kanazawa, A.; O’Dell, M.; Hellens, R.P. The Binding of Nuclear Factors to the as-1 Element in the CaMV 35S Promoter is Affected by Cytosine Methylation in vitro. Plant Biol. 2007, 9, 435–441. [Google Scholar] [CrossRef]
- Okumura, A.; Shimada, A.; Yamasaki, S.; Horino, T.; Iwata, Y.; Koizumi, N.; Nishihara, M.; Mishiba, K. CaMV-35S promoter sequence-specific DNA methylation in lettuce. Plant Cell Rep. 2016, 35, 43–51. [Google Scholar] [CrossRef] [PubMed]
- Weinhold, A.; Kallenbach, M.; Baldwin, I.T. Progressive 35S promoter methylation increases rapidly during vegetative development in transgenic Nicotiana attenuata plants. BMC Plant Biol. 2013, 13, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muskens, M.W.M.; Vissers, A.P.A.; Mol, J.N.M.; Kooter, J.M. Role of inverted DNA repeats in transcriptional and post-transcriptional gene silencing. Plant Mol. Biol. 2000, 43, 243–260. [Google Scholar] [CrossRef]
- Peremarti, A.; Twyman, R.M.; Gómez-Galera, S.; Naqvi, S.; Farré, G.; Sabalza, M.; Miralpeix, B.; Dashevskaya, S.; Yuan, D.; Ramessar, K.; et al. Promoter diversity in multigene transformation. Plant Mol. Biol. 2010, 73, 363–378. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, J.A.; Shirasu, K.; Deng, X.W. The diverse roles of ubiquitin and the 26S proteasome in the life of plants. Nat. Rev. Genet. 2003, 4, 948–958. [Google Scholar] [CrossRef]
- Han, S.-W.; Jung, B.-K.; Park, S.-H.; Ryu, K.-Y. Reversible Regulation of Polyubiquitin Gene UBC via Modified Inducible CRISPR/Cas9 System. Int. J. Mol. Sci. 2019, 20, 3168. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stone, S.L. Chapter 26—Ubiquitination of Plant Transcription Factors. In Plant Transcription Factors; Gonzalez, D.H., Ed.; Academic Press: Boston, MA, USA, 2016; pp. 395–409. [Google Scholar]
- Wang, P.-H.; Kumar, S.; Zeng, J.; McEwan, R.; Wright, T.; Gupta, M. Transcription Terminator-Mediated Enhancement in Transgene Expression in Maize: Preponderance of the AUGAAU Motif Overlapping With Poly(A) Signals. Front. Plant Sci. 2020, 11, 1498. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Martinelli, A.P.; Bouchard, R.A.; Finer, J.J. A soybean (Glycine max) polyubiquitin promoter gives strong constitutive expression in transgenic soybean. Plant Cell Rep. 2009, 28, 837–849. [Google Scholar] [CrossRef]
- Mann, D.G.J.; King, Z.R.; Liu, W.; Joyce, B.L.; Percifield, R.J.; Hawkins, J.S.; LaFayette, P.R.; Artelt, B.J.; Burris, J.N.; Mazarei, M.; et al. Switchgrass (Panicum virgatum L.) polyubiquitin gene (PvUbi1 and PvUbi2) promoters for use in plant transformation. BMC Biotechnol. 2011, 11, 74. [Google Scholar] [CrossRef] [Green Version]
- Cary, T.J.; Rylott, E.L.; Zhang, L.; Routsong, R.M.; Palazzo, A.J.; Strand, S.E.; Bruce, N.C. Field trial demonstrating phytoremediation of the military explosive RDX by XplA/XplB-expressing switchgrass. Nat. Biotechnol. 2021, 39, 1216–1219. [Google Scholar] [CrossRef]
- Wang, J.; Oard, J.H. Rice ubiquitin promoters: Deletion analysis and potential usefulness in plant transformation systems. Plant Cell Rep. 2003, 22, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Beringer, J.; Chen, W.; Garton, R.; Sardesai, N.; Wang, P.-H.; Zhou, N.; Gupta, M.; Wu, H. Comparison of the impact of viral and plant-derived promoters regulating selectable marker gene on maize transformation and transgene expression. Plant Cell Rep. 2017, 36, 519–528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolabu, T.W.; Cong, L.; Park, J.-J.; Bao, Q.; Chen, M.; Sun, J.; Xu, B.; Ge, Y.; Chai, M.; Liu, Z.; et al. Development of a Highly Efficient Multiplex Genome Editing System in Outcrossing Tetraploid Alfalfa (Medicago sativa). Front. Plant. Sci. 2020, 11, 1063. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Liu, Y.; Guo, Y.; Duan, W.; Fan, P.; Li, S.; Liang, Z. Optimizing the CRISPR/Cas9 system for genome editing in grape by using grape promoters. Hortic. Res. 2021, 8, 52. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. Identification and validation of promoters and cis-acting regulatory elements. Plant Sci. 2014, 217–218, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Joung, Y.H.; Kamo, K. Expression of a polyubiquitin promoter isolated from Gladiolus. Plant Cell Rep. 2006, 25, 1081–1088. [Google Scholar] [CrossRef]
- Hua, Y.W.; Huang, T.D.; Huang, H.S. Micropropagation of self-rooting juvenile clones by secondary somatic embryogenesis in Hevea brasiliensis. Plant Breed. 2010, 129, 202–207. [Google Scholar] [CrossRef]
- Grefen, C.; Donald, N.; Hashimoto, K.; Kudla, J.; Schumacher, K.; Blatt, M.R. A ubiquitin-10 promoter-based vector set for fluorescent protein tagging facilitates temporal stability and native protein distribution in transient and stable expression studies. Plant J. 2010, 64, 355–365. [Google Scholar] [CrossRef]
- Wolabu, T.W.; Park, J.-J.; Chen, M.; Cong, L.; Ge, Y.; Jiang, Q.; Debnath, S.; Li, G.; Wen, J.; Wang, Z. Improving the genome editing efficiency of CRISPR/Cas9 in Arabidopsis and Medicago truncatula. Planta 2020, 252, 15. [Google Scholar] [CrossRef]
- Kumar, M.; Ayzenshtat, D.; Marko, A.; Bocobza, S. Optimization of T-DNA configuration with UBIQUITIN10 promoters and tRNA–sgRNA complexes promotes highly efficient genome editing in allotetraploid tobacco. Plant Cell Rep. 2022, 41, 175–194. [Google Scholar] [CrossRef]
- Tang, C.; Yang, M.; Fang, Y.; Luo, Y.; Gao, S.; Xiao, X.; An, Z.; Zhou, B.; Zhang, B.; Tan, X.; et al. The rubber tree genome reveals new insights into rubber production and species adaptation. Nat. Plants 2016, 2, 16073. [Google Scholar] [CrossRef] [Green Version]
- Cheng, H. HeveaDB: A Hub for Rubber Tree Genetic and Genomic Resources. In The Rubber Tree Genome; Matsui, M., Chow, K.-S., Eds.; Springer International Publishing: Cham, Switzerland, 2020; pp. 137–152. [Google Scholar]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An Integrative Toolkit Developed for Interactive Analyses of Big Biological Data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Qin, Y.; Xiao, X.; Tang, C. Screening of valid reference genes for real-time RT-PCR data normalization in Hevea brasiliensis and expression validation of a sucrose transporter gene HbSUT3. Plant Sci. 2011, 181, 132–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Tang, D.; Hong, L.; Xu, W.; Huang, J.; Li, M.; Gu, M.; Xue, Y.; Cheng, Z. DEP and AFO Regulate Reproductive Habit in Rice. PLoS Genet. 2010, 6, e1000818. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horsch, R.B.; Rogers, S.G.; Fraley, R.T. Transgenic plants. Cold Spring Harb. Symp. Quant. Biol. 1985, 50, 433–437. [Google Scholar] [CrossRef]
- Hernandez-Garcia, C.M.; Bouchard, R.A.; Rushton, P.J.; Jones, M.L.; Chen, X.; Timko, M.P.; Finer, J.J. High level transgenic expression of soybean (Glycine max) GmERF and Gmubi gene promoters isolated by a novel promoter analysis pipeline. BMC Plant Biol. 2010, 10, 237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torre, C.M.D.L.; Finer, J.J. The intron and 5′ distal region of the soybean Gmubi promoter contribute to very high levels of gene expression in transiently and stably transformed tissues. Plant Cell Rep. 2015, 34, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Hernandez-Garcia, C.M.; Finer, J.J. A novel cis-acting element in the GmERF3 promoter contributes to inducible gene expression in soybean and tobacco after wounding. Plant Cell Rep. 2016, 35, 303–316. [Google Scholar] [CrossRef]
- Erpen, L.; Tavano, E.C.R.; Harakava, R.; Dutt, M.; Grosser, J.W.; Piedade, S.M.S.; Mendes, B.M.J.; Mourão Filho, F.A.A. Isolation, characterization, and evaluation of three Citrus sinensis-derived constitutive gene promoters. Plant Cell Rep. 2018, 37, 1113–1125. [Google Scholar] [CrossRef]
- Subhi, S.M.; Ghulam, K.A.P.; Omar, A.R. Isolation of an oil palm constitutive promoter derived from ubiquitin extension protein (uep2) gene. J. Oil Palm Res. 2019, 31, 28–41. [Google Scholar] [CrossRef] [Green Version]
- Yang, H.; Wang, X.; Wei, Y.; Deng, Z.; Liu, H.; Chen, J.; Dai, L.; Xia, Z.; He, G.; Li, D. Transcriptomic analyses reveal molecular mechanisms underlying growth heterosis and weakness of rubber tree seedlings. BMC Plant Biol. 2018, 18, 10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, S.; Hua, Y.; Li, J.; Dai, X.; Yang, X.; Udayabhanu, J.; Huang, H.; Huang, T. Comparative analysis of latex transcriptomes reveals the potential mechanisms underlying rubber molecular weight variations between the Hevea brasiliensis clones RRIM600 and Reyan7-33–97. BMC Plant Biol. 2021, 21, 244. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Xu, L.; Liu, X.; Xin, L.; Wu, S.; Chen, X. Development of potent promoters that drive the efficient expression of genes in apple protoplasts. Hortic. Res. 2021, 8, 211. [Google Scholar] [CrossRef] [PubMed]
- Dai, X.; Yang, X.; Wang, C.; Fan, Y.; Xin, S.; Hua, Y.; Wang, K.; Huang, H. CRISPR/Cas9-mediated genome editing in Hevea brasiliensis. Ind. Crops Prod. 2021, 164, 113418. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xin, S.; Udayabhanu, J.; Dai, X.; Hua, Y.; Fan, Y.; Huang, H.; Huang, T. Identification and Functional Evaluation of Three Polyubiquitin Promoters from Hevea brasiliensis. Forests 2022, 13, 952. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060952
Xin S, Udayabhanu J, Dai X, Hua Y, Fan Y, Huang H, Huang T. Identification and Functional Evaluation of Three Polyubiquitin Promoters from Hevea brasiliensis. Forests. 2022; 13(6):952. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060952
Chicago/Turabian StyleXin, Shichao, Jinu Udayabhanu, Xuemei Dai, Yuwei Hua, Yueting Fan, Huasun Huang, and Tiandai Huang. 2022. "Identification and Functional Evaluation of Three Polyubiquitin Promoters from Hevea brasiliensis" Forests 13, no. 6: 952. https://0-doi-org.brum.beds.ac.uk/10.3390/f13060952