Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Fungal Isolates and Plant Material
2.2. Pathogenicity Tests
2.3. Statistical Analyses
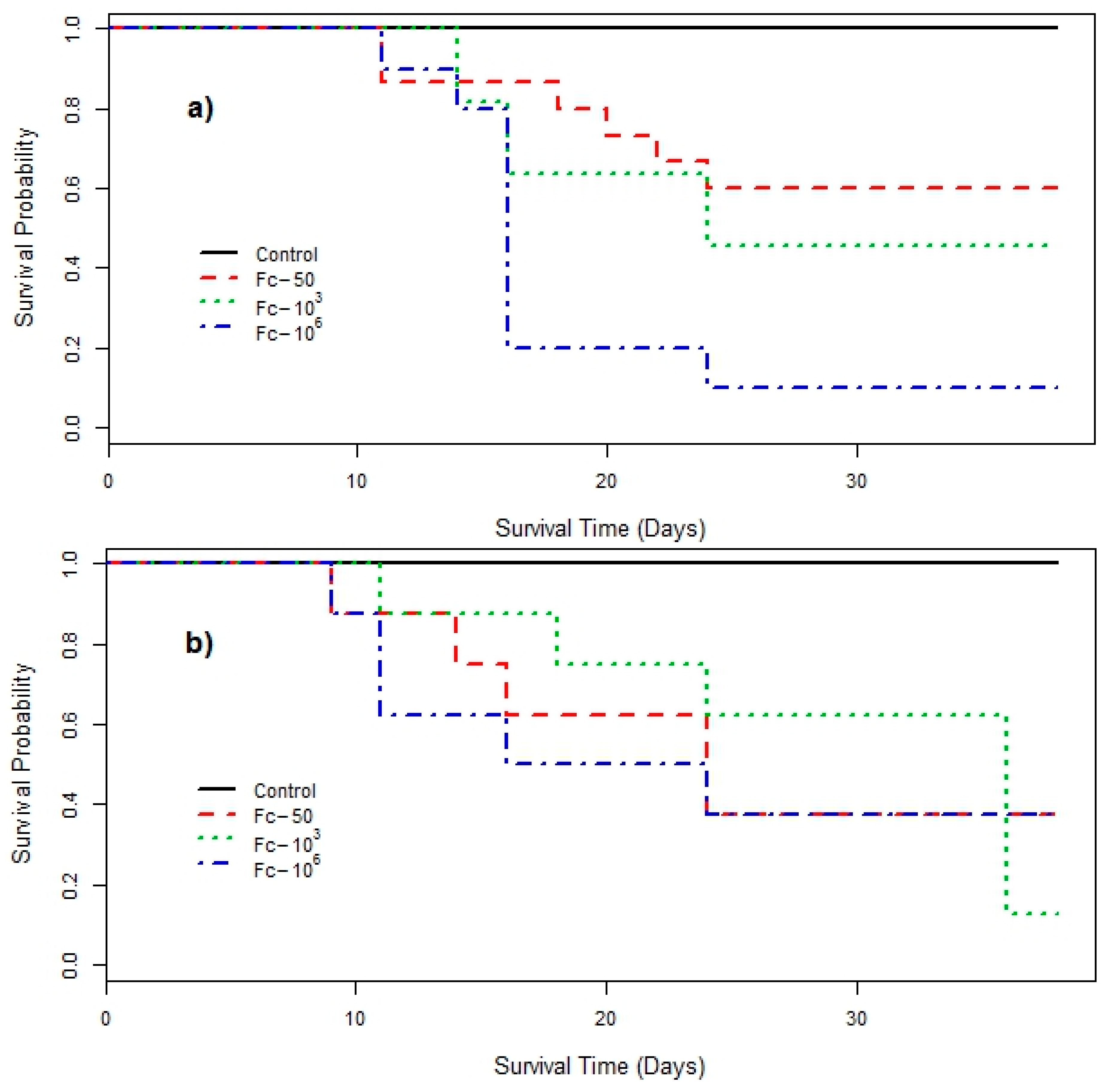
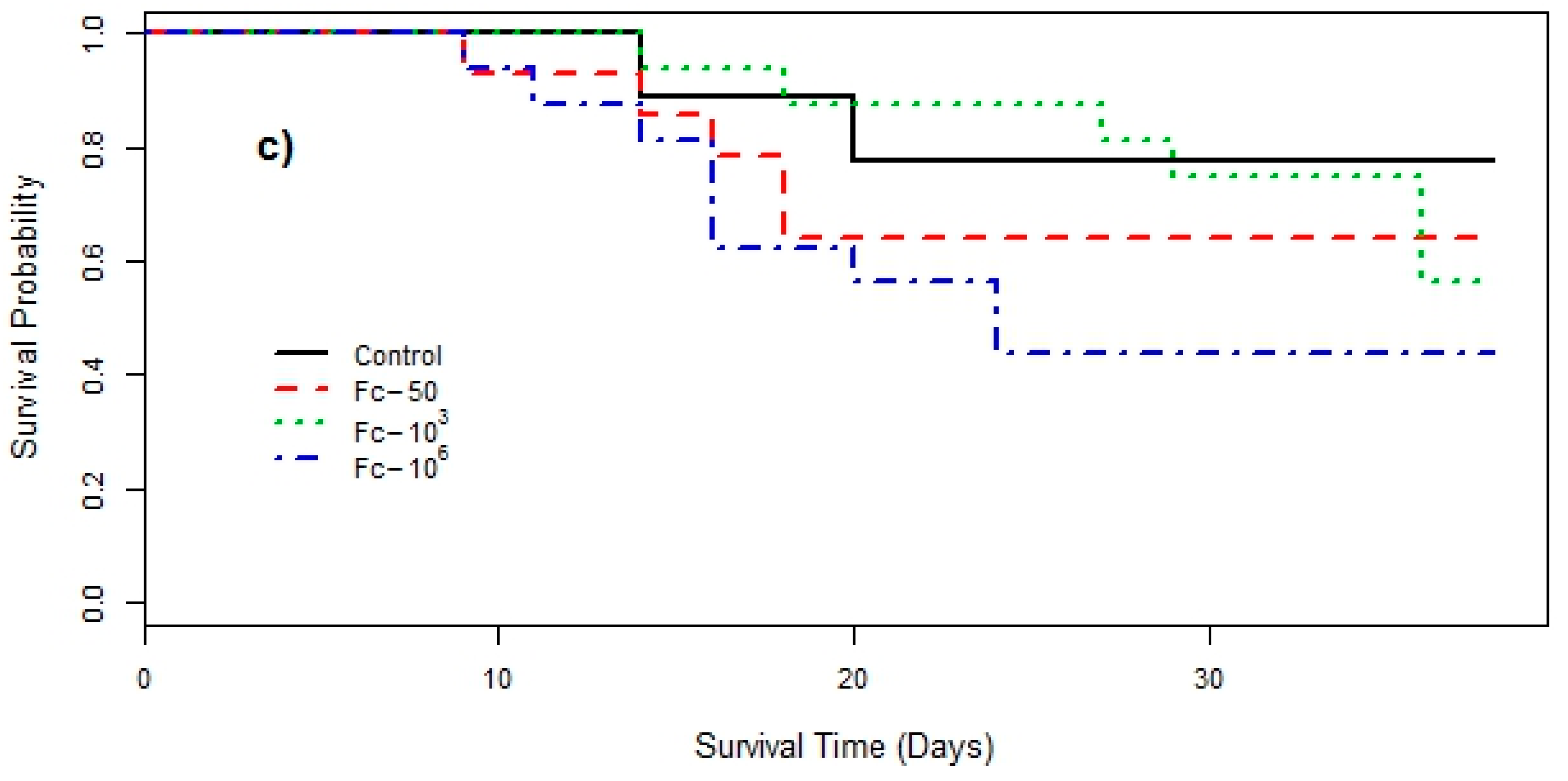
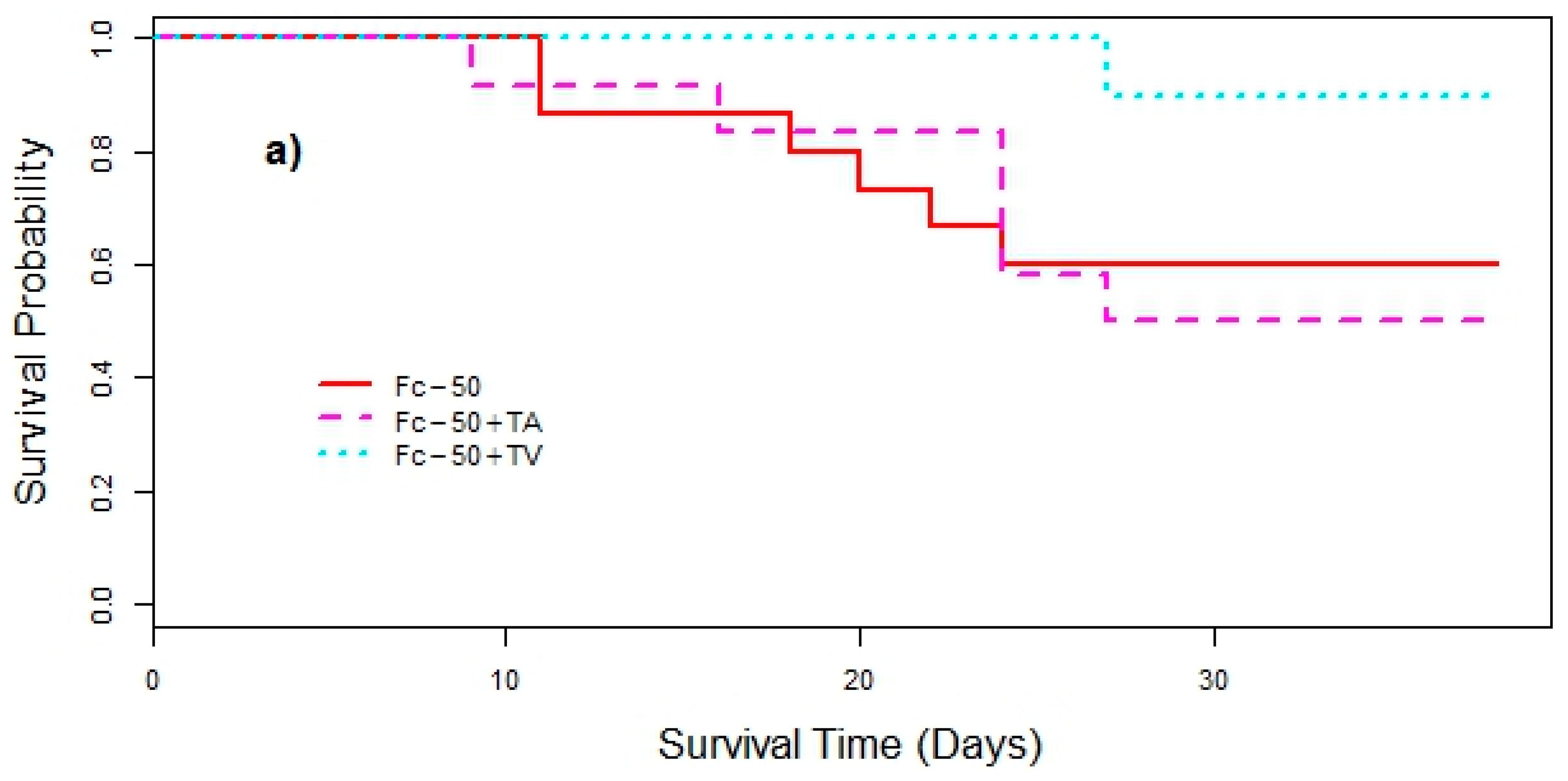
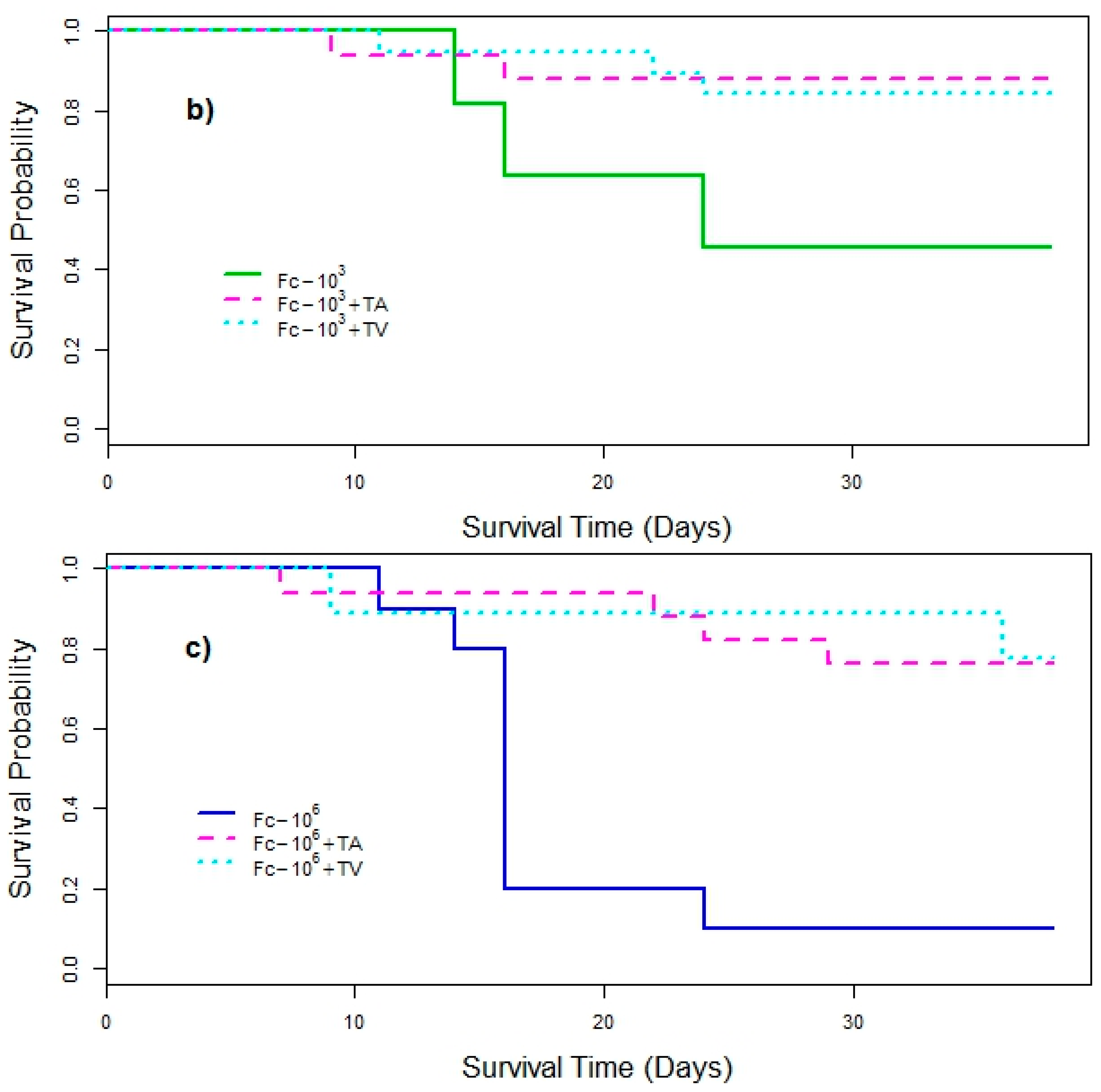
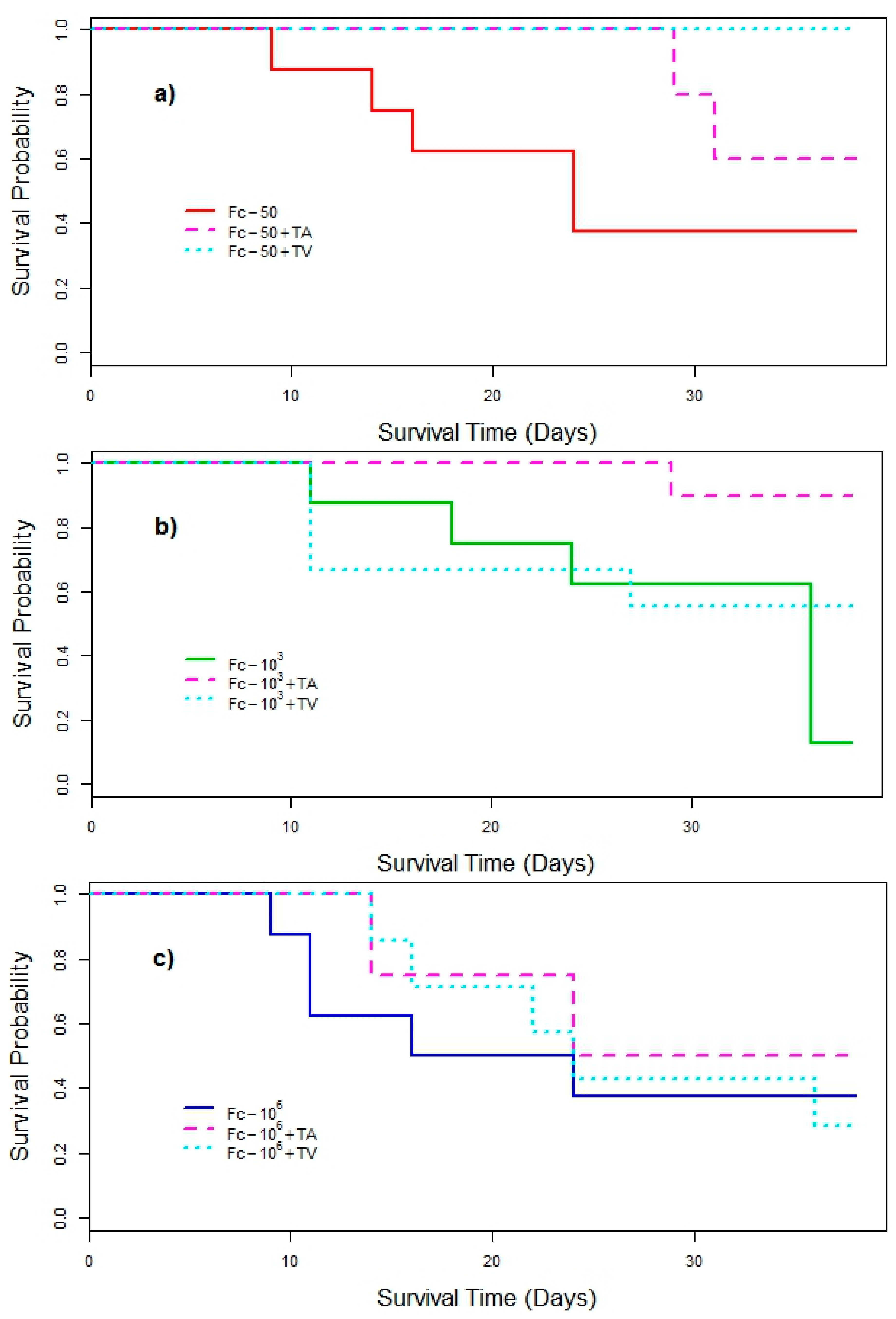
3. Results
4. Discussion
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- European Food Safety Authority (EFSA). Risk assessment of Gibberella circinata for the EU territory and identification and evaluation of risk management options. EFSA 2010, 8, 1–93. [Google Scholar] [CrossRef]
- Wingfield, M.J.; Hammerbacher, A.; Ganley, R.J.; Steenkamp, E.T.; Gordon, T.R.; Wingfield, B.D.; Coutinho, T.A. Pitch canker caused by Fusarium circinatum—A growing threat to pine plantations and forests worldwide. Australas. Plant Pathol. 2008, 37, 319–334. [Google Scholar] [CrossRef]
- Hepting, G.H.; Roth, E.R. Pitch canker, a new disease of some Southern pines. J. For. 1946, 44, 742–744. [Google Scholar]
- Hepting, G.H.; Roth, E.R. Host relations and spread of the pine pitch canker disease. Phytopathology 1953, 43, 475. [Google Scholar]
- Viljoen, A.; Wingfield, M.J.; Marasas, W.F. First report of Fusarium subglutinans F. sp. pini on seedlings in South Africa. Plant Dis. 1994, 78, 309–312. [Google Scholar] [CrossRef]
- Guerra-Santos, J.J.; Cibrián-Tovar, D. El cancro resinoso causado por Fusarium subglutinans (Wollenw y Reink) Nelson, Tousson y Marasas, Una nueva enfermedad. Revista Chapingo Serie Ciencias Forestales Y Del Ambiente 1998, 4, 279–284. (In Spanish) [Google Scholar]
- Wingfield, M.J.; Jacobs, A.; Coutinho, T.A.; Ahumada, R.; Wingfield, B.D. First report of the pitch canker fungus, Fusarium circinatum, on pines in Chile. Plant Pathol. 2002, 51, 397. [Google Scholar] [CrossRef]
- Cho, W.D.; Shin, H.D. List of Plant Diseases in Korea, 4th ed.; Korean Society of Plant Pathology: Seoul, Korea, 2004. [Google Scholar]
- Kobayashi, T.; Muramoto, M. Pitch canker of Pinus luchuensis, a new disease of Japanese forests. For. Pests 1989, 40, 169–173. [Google Scholar]
- Alonso, R.; Bettucci, L. First report of the pitch canker fungus Fusarium circinatum affecting Pinus taeda seedlings in Uruguay. Australas. Plant Dis. Notes 2009, 4, 91–92. [Google Scholar] [CrossRef]
- Steenkamp, E.T.; Rodas, C.A.; Kvas, M.; Wingfield, M.J. Fusarium circinatum and pitch canker of Pinus in Colombia. Australas. Plant Pathol. 2012, 41, 483–491. [Google Scholar] [CrossRef]
- Pfenning, L.H.; Costa, S.; de Melo, M.P.; Costa, H.; Aires, J. First report and characterization of Fusarium circinatum, the causal agent of pitch canker in Brazil. Trop. Plant Pathol. 2014, 39, 210–216. [Google Scholar] [CrossRef]
- Dwinell, D. Global Distribution of the Pitch Canker Fungus. Current and Potential Impacts of Pitch Canker in Radiata Pine. In Proceedings of the IMPACT Monterey Workshop, Monterey, CA, USA, 30 Noverber–3 December 1999. [Google Scholar]
- Landeras, E.; García, P.; Fernández, Y.; Braña, M.; Pérez-Sierra, A.; León, M.; Fernández-Alonso, O.; Méndez-Lodos, S.; Abad-Campos, P.; Berbegal, M.; et al. Outbreak of pitch canker caused by Fusarium circinatum on Pinus spp. in Northern Spain. Plant Dis. 2005, 89, 1015. [Google Scholar] [CrossRef]
- EPPO. First report of Gibberella circinata in France. EPPO Rep. Serv. 2006, 5, 9. [Google Scholar]
- Carlucci, A.; Colatruglio, L.; Frisullo, S. First report of pitch canker caused by Fusarium circinatum on Pinus halepensis and P. pinea in Apulia (Southern Italy). Plant Dis. 2007, 91, 1683. [Google Scholar] [CrossRef]
- Bragança, H.; Diogo, E.; Moniz, F.; Amaro, P. First report of pitch canker on pines caused by Fusarium circinatum. Plant Dis. 2009, 93, 1079. [Google Scholar] [CrossRef]
- CAB International. Reports—Gibberella Circinata (Pitch Canker)-Pinus-Crop Protection Compendium; CABI: Wallingford, UK, 2007. [Google Scholar]
- Bezos, D.; Martínez-Álvarez, P.; Fernández, M.; Diez, J.J. Epidemiology and management of pine pitch canker disease in Europe—A Review. Balt. For. 2017, 23, 279–293. [Google Scholar]
- Ganley, R.J.; Watt, M.S.; Manning, L.; Iturritxa, E. A global climatic risk assessment of pitch canker disease. Can. J. For. Res. 2009, 39, 2246–2256. [Google Scholar] [CrossRef]
- Watt, M.S.; Ganley, R.J.; Kriticos, D.J.; Manning, L.K. Dothistroma needle blight and pitch canker: The current and future potential distribution of two important diseases of Pinus species. Can. J. For. Res. 2011, 424, 412–424. [Google Scholar] [CrossRef]
- Houston Durrant, T.; De Rigo, D.; Caudullo, G. Pinus sylvestris in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., de Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 132–133. [Google Scholar]
- Pérez-Sierra, A.; Landeras, E.; León, M.; Berbegal, M.; García-Jiménez, J.; Armengol, J. Characterization of Fusarium circinatum from Pinus spp. in northern Spain. Mycol. Res. 2007, 111, 832–839. [Google Scholar] [CrossRef] [PubMed]
- Iturritxa, E.; Ganley, R.J.; Raposo, R.; García-Serna, I.; Mesanza, N.; Kirkpatrick, S.C.; Gordon, T.R. Resistance levels of Spanish conifers against Fusarium circinatum and Diplodia pinea. For. Pathol. 2013, 43, 488–495. [Google Scholar] [CrossRef]
- Martínez-Alvarez, P.; Pando, V.; Diez, J.J. Alternative species to replace Monterey pine plantations affected by pitch canker caused by Fusarium circinatum in northern Spain. Plant Pathol. 2014, 63, 1086–1094. [Google Scholar] [CrossRef]
- Donnelly, K.; Cavers, S.; Cottrell, J.E.; Ennos, R.A. Genetic variation for needle traits in Scots pine (Pinus sylvestris L.). Tree Genet. Genomes 2016, 12, 40. [Google Scholar] [CrossRef]
- Belletti, P.; Ferrazzini, D.; Piotti, A.; Monteleone, I.; Ducci, F. Genetic variation and divergence in Scots pine (Pinus sylvestris L.) within its natural range in Italy. Eur. J. For. Res. 2012, 131, 1127–1138. [Google Scholar] [CrossRef]
- Wójkiewicz, B.; Cavers, S.; Wachowiak, W. Current approaches and perspectives in population genetics of Scots pine (Pinus sylvestris L.). For. Sci. 2016, 62, 343–354. [Google Scholar] [CrossRef]
- Caudullo, G.; Tinner, W.; De Rigo, D. Picea abies in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., De Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 115–116. [Google Scholar]
- Lagercrantz, U.; Ryman, N. Genetic structure of Norway spruce (Picea abies): Concordance of morphological and allozymic variation. Evolution 1990, 44, 38–53. [Google Scholar] [PubMed]
- Heuertz, M.; De Paoli, E.; Ka, T.; Larsson, H.; Jurman, I.; Morgante, M.; Lascoux, M.; Gyllenstrand, N. Multilocus patterns of nucleotide diversity, linkage disequilibrium and demographic history of Norway spruce [Picea abies (L.) Karst]. Genetics 2006, 174, 2095–2105. [Google Scholar] [CrossRef] [PubMed]
- Ballian, D.; Ravazzi, C.; De Rigo, D.; Caudullo, G. Pinus mugo in Europe: Distribution, habitat, usage and threats. In European Atlas of Forest Tree Species; San-Miguel-Ayanz, J., De Rigo, D., Caudullo, G., Houston Durrant, T., Mauri, A., Eds.; Publication Office of the European Union: Luxembourg, 2016; pp. 124–125. [Google Scholar]
- Bendel, M.; Kienast, F.; Rigling, D.; Bugmann, H. Impact of root-rot pathogens on forest succession in unmanaged Pinus mugo stands in the Central Alps. Can. J. For. Res. 2006, 2674, 2666–2674. [Google Scholar] [CrossRef]
- Alves-Santos, F.M.; Diez, J.J. Biological control of Fusarium. In Control of Fusarium Diseases; Alves-Santos, F.M., Diez, J.J., Eds.; Research Signpost: Kerala, India, 2011; p. 250. [Google Scholar]
- Harman, G.E.; Howell, C.R.; Viterbo, A.; Chet, I.; Lorito, M. Trichoderma species--opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2004, 2, 43–56. [Google Scholar] [CrossRef] [PubMed]
- Howell, C.R. Mechanisms Employed by Trichoderma Species in the biological control of plant diseases: The history and evolution of current concepts. Plant Dis. 2003, 87, 4–10. [Google Scholar] [CrossRef]
- Benítez, T.; Rincón, A.M.; Limón, M.C.; Codón, A.C. Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 2004, 7, 249–260. [Google Scholar] [PubMed]
- Mitchell, R.G.; Zwolinski, J.; Jones, N.; Coutinho, T. The effect of applying prophylactic measures on the post-planting survival of Pinus patula in South Africa. South. Afr. For. J. 2004, 200, 51–58. [Google Scholar]
- Martínez-Alvarez, P.; Alves-Santos, F.M.; Diez, J.J. In vitro and in vivo interactions between Trichoderma viride and Fusarium circinatum. Silv. Fenn. 2012, 46, 303–316. [Google Scholar] [CrossRef]
- Moraga-Suazo, P.; Opazo, A.; Zaldúa, S.; González, G.; Sanfuentes, E. Evaluation of Trichoderma spp. and Clonostachys spp. strains to control Fusarium circinatum in Pinus radiata seedlings. Chil. J. Agric. Res. 2011, 71, 412–417. [Google Scholar] [CrossRef]
- López-López, N.; Segarra, G.; Vergara, O.; López-fabal, A.; Trillas, M.I. Compost from forest cleaning green waste and Trichoderma asperellum strain T34 reduced incidence of Fusarium circinatum in Pinus radiata seedlings. Biol. Control 2016, 95, 31–39. [Google Scholar] [CrossRef]
- Martínez-Álvarez, P.; Fernández-González, R.A.; Sanz-Ros, A.V.; Pando, V.; Diez, J.J. Two fungal endophytes reduce the severity of pitch canker disease in Pinus radiata seedlings. Biol. Control 2016, 94, 1–10. [Google Scholar] [CrossRef]
- Cerqueira, A.; Alves, A.; Berenguer, H.; Correia, B. Phosphite shifts physiological and hormonal profile of Monterey pine and delays Fusarium circinatum progression. Plant Physiol. Biochem. 2017, 114, 88–99. [Google Scholar] [CrossRef] [PubMed]
- Kaplan, E.L.; Meier, P. Nonparametric estimation from incomplete observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Therneau, T. A Package for Survival Analysis in S. R Package Version 2.38; R Foundation for Statistical Computing: Vienna, Austria, 2017; Volume 143. [Google Scholar]
- European Commission. Invasive Alien Species: A European Response; European Commission: Luxembourg, 2014. [Google Scholar] [CrossRef]
- Santini, A.; Ghelardini, L.; de Pace, C.; Desprez-Loustau, M.L.; Capretti, P.; Chandelier, A.; Cech, T.; Chira, D.; Diamandis, S.; Gnitniekis, T.; et al. Biogeographical patterns and determinants of invasion by forest pathogens in Europe. New Phytol. 2013, 197, 238–250. [Google Scholar] [CrossRef] [PubMed]
- Eschen, R.; Britton, K.; Brockerhoff, E.; Burgess, T.; Dalley, V.; Epanchin-Niell, R.S.; Gupta, K.; Hardy, G.; Huang, Y.; Kenis, M.; et al. International variation in phytosanitary legislation and regulations governing importation of plants for planting. Environ. Sci. Policy 2015, 51, 228–237. [Google Scholar] [CrossRef]
- Stenlid, J.; Oliva, J. Phenotypic interactions between tree hosts and invasive forest pathogens in the light of globalization and climate change. Philos. Trans. R. Soc. B 2016, 371, 20150455. [Google Scholar] [CrossRef] [PubMed]
- Rikala, R. Production and quality requirements of forest tree seedlings in Finland. Tree Plant Notes 2000, 49, 56–60. [Google Scholar]
- Menkis, A.; Burokien, D.; Stenlind, J.; Stenström, E. High-throughput sequencing shows high fungal diversity and community segregation in the rhizospheres of container-grown conifer seedlings. Forests 2016, 7, 44. [Google Scholar] [CrossRef]
- Prus-Glowacki, W.; Stephan, B.R. Genetic variation of Pinus sylvestris from Spain in relation to other European populations. Silv. Fenn. 1994, 43, 7–14. [Google Scholar]
- Cheddadi, R.; Vendramin, G.G.; Litt, T.; Kageyama, M.; Lorentz, S.; Laurent, J.; François, L.; De Beaulieu, J.; Sadoril, L.; Jost, A.; et al. Imprints of glacial refugia in the modern genetic diversity of Pinus sylvestris. Glob. Ecol. Biogeogr. 2006, 15, 271–282. [Google Scholar] [CrossRef]
- Dvorak, W.; Hodge, G.; Kietzka, J. Genetic variation in survival, growth, and stem form of Pinus leiophylla in Brazil and South Africa and provenance resistance to pitch canker. South. Hemisphere For. J. 2007, 69, 125–135. [Google Scholar] [CrossRef]
- Dvorak, W.S.; Potter, K.M.; Hipkins, V.D.; Hodge, G.R. Genetic diversity and gene exchange in Pinus oocarpa, a Mesoamerican pine with resistance to the pitch canker fungus (Fusarium circinatum). Int. J. Plant Sci. 2009, 170, 609–626. [Google Scholar] [CrossRef]
- Mitchell, R.G.; Wingfield, M.J.; Hodge, G.R.; Steenkamp, E.T.; Coutinho, T.A. Selection of Pinus spp. in South Africa for tolerance to infection by the pitch canker fungus. New For. 2012, 43, 473–489. [Google Scholar] [CrossRef]
- Hodge, G.R.; Dvorak, W.S. Variation in pitch canker resistance among provenances of Pinus patula and Pinus tecunumanii from Mexico and Central America. New For. 2007, 33, 193–206. [Google Scholar] [CrossRef]
- Elvira-Recuenco, M.; Iturritxa, E.; Majada, J.; Alia, R.; Raposo, R. Adaptive potential of maritime pine (Pinus pinaster) populations to the emerging pitch canker pathogen, Fusarium circinatum. PLoS ONE 2014, 9, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Vivas, M.; Zas, R.; Solla, A. Screening of Maritime pine (Pinus pinaster) for resistance to Fusarium circinatum, the causal agent of Pitch Canker disease. Forestry 2012, 85, 185–192. [Google Scholar] [CrossRef]
- Iturritxa, E.; Mesanza, N.; Elvira-Recuenco, M.; Serrano, Y.; Quintana, E.; Raposo, R. Evaluation of genetic resistance in Pinus to pitch canker in Spain. Australas. Plant Pathol. 2012, 41, 601–607. [Google Scholar] [CrossRef]
- Panter, S.N.; Jones, D.A. Age-related resistance to plant pathogens. Adv. Bot. Res. 2002, 38, 251–280. [Google Scholar]
- Vinale, F.; Sivasithamparam, K.; Ghisalberti, E.L.; Marra, R.; Woo, S.L.; Lorito, M. Trichoderma–plant–pathogen interactions. Soil Biol. Biochem. 2008, 40, 1–10. [Google Scholar] [CrossRef]
- Lo, T. General mechanisms of action of microbial biocontrol agents. Plant Pathol. Bull. 1998, 7, 155–166. [Google Scholar]
- Iturritxa, E.; Desprez-loustau, M.L.; García-serna, I.; Quintana, E.; Mesanza, N.; Aitken, J. Effect of alternative disinfection treatments against fungal canker in seeds of Pinus radiata. Seed Technol. 2011, 33, 88–110. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Provenance | Coordinates | Altitude | Climatic Data | ||
|---|---|---|---|---|---|---|
| North | West | MASL | Precipitation (mm) | Mean Temperature (°C) | ||
| Pinus sylvestris | PI-A210-2 | 47°40′00″ | 25°20′00″ | 1100–1400 | 840 | 5.6 |
| Pinus mugo | Muntele Oslea | 45°13′00″ | 22°53′00″ | 1850–1900 | 1310 | 3.8 |
| Picea abies | MO C-210-2 | 45°15′00″ | 24°45′00″ | 1350–1650 | 800 | 4.3 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-García, J.; Paraschiv, M.; Flores-Pacheco, J.A.; Chira, D.; Diez, J.J.; Fernández, M. Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol. Forests 2017, 8, 318. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090318
Martín-García J, Paraschiv M, Flores-Pacheco JA, Chira D, Diez JJ, Fernández M. Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol. Forests. 2017; 8(9):318. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090318
Chicago/Turabian StyleMartín-García, Jorge, Marius Paraschiv, Juan Asdrúbal Flores-Pacheco, Danut Chira, Julio Javier Diez, and Mercedes Fernández. 2017. "Susceptibility of Several Northeastern Conifers to Fusarium circinatum and Strategies for Biocontrol" Forests 8, no. 9: 318. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090318