Excessive Accumulation of Chinese Fir Litter Inhibits Its Own Seedling Emergence and Early Growth—A Greenhouse Perspective

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Collection of Leaf Litter and Seeds
2.2. Greenhouse Experiments
2.3. Statistical Analysis
3. Results
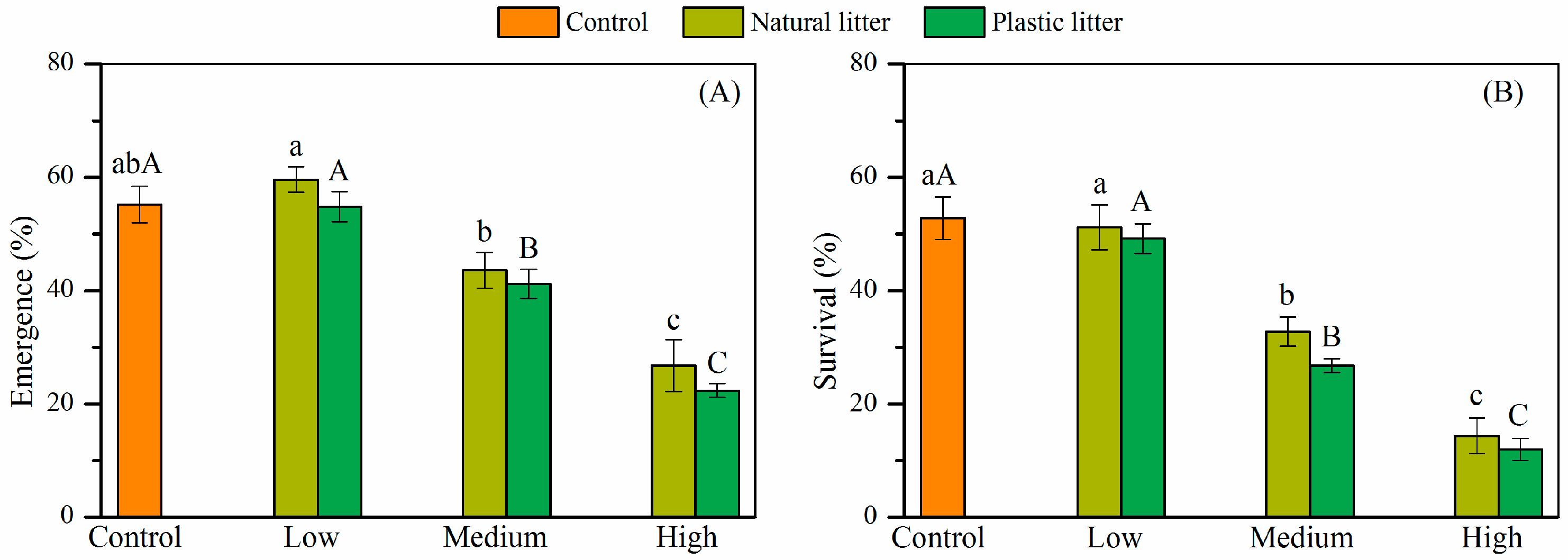
3.1. Effect of Litter on Seed Emergence and Survival
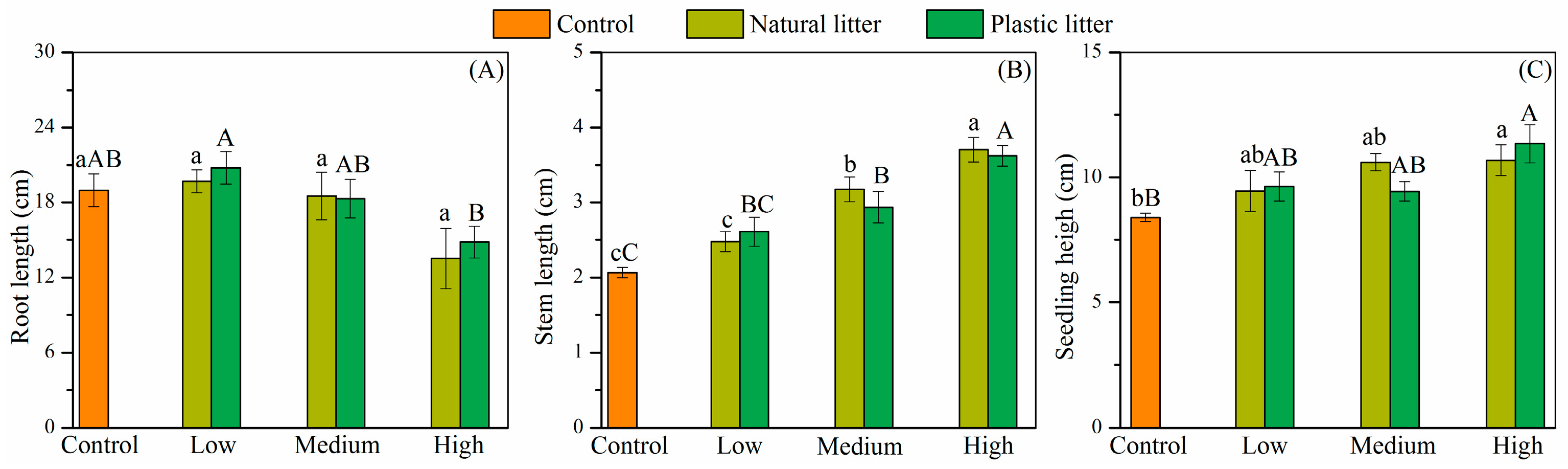
3.2. Effect of Litter on Seedling Growth
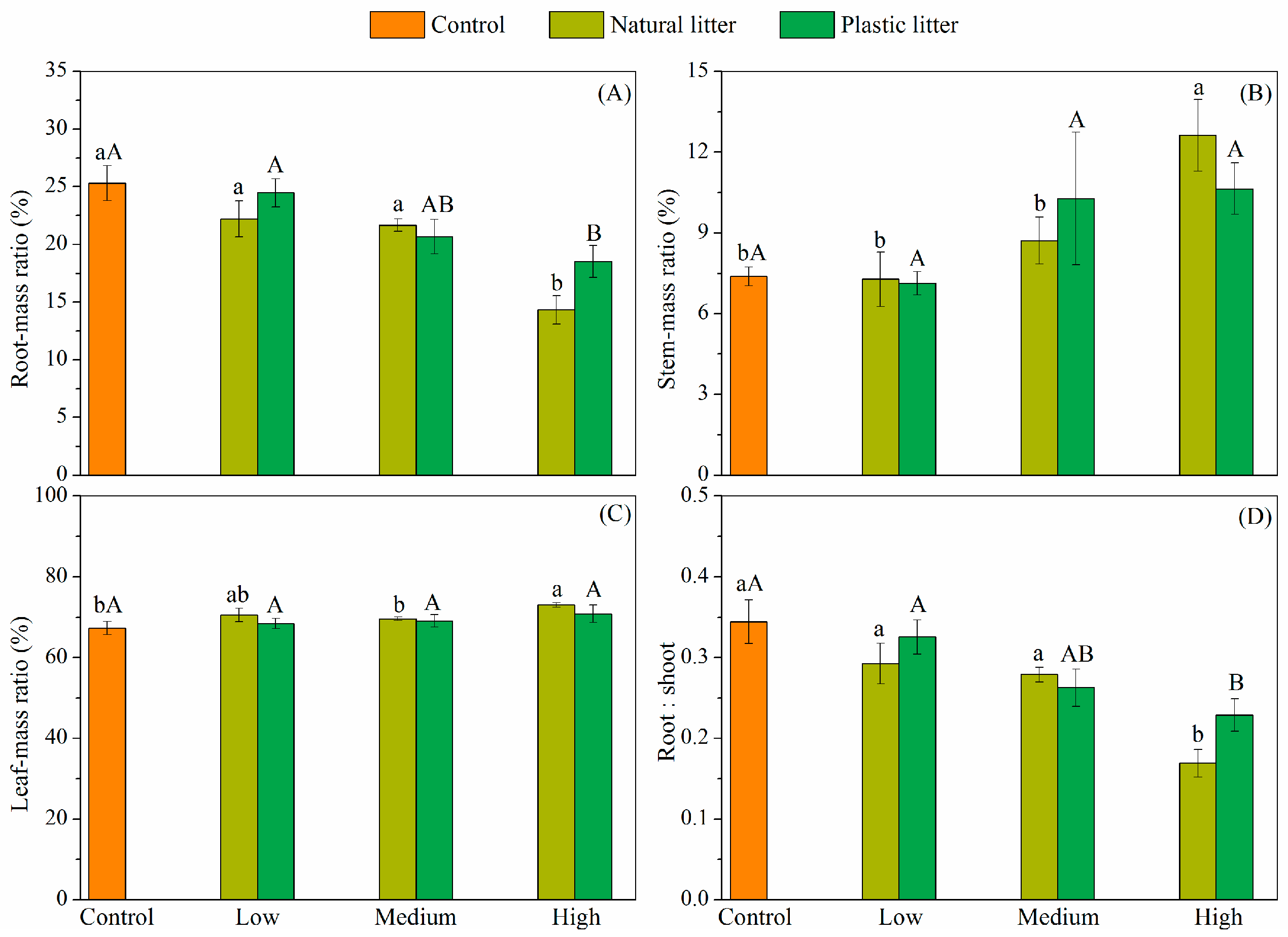
3.3. Effect of Litter on Seedling Biomass and Biomass Allocation
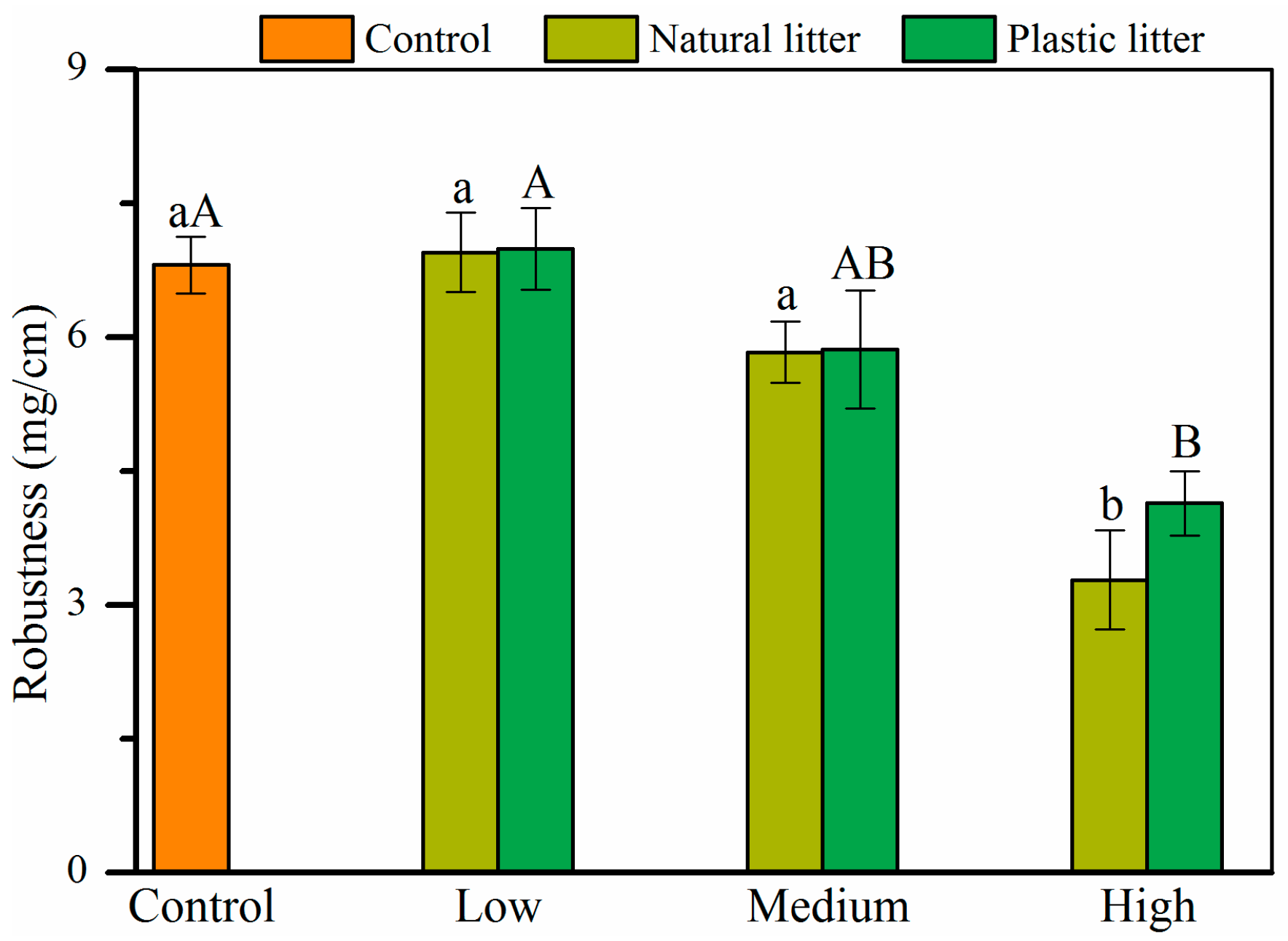
3.4. Effect of Litter on Robustness
4. Discussion
4.1. Physical versus Chemical Effects
4.2. Effects of Litter Amount on Seedling Emergence and Survival
4.3. Effects of Litter Amount on Seedling Biomass Allocation and Robustness
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Aubin, I.; Messier, C.; Bouchard, A. Can plantation develop understory biological and physical attributes of naturally regenerated forests? Biol. Conserv. 2008, 141, 2461–2476. [Google Scholar] [CrossRef] [Green Version]
- Manson, D.G.; Schmidt, S.; Bristow, M.; Erskine, P.D.; Vanclay, J.K. Species-site matching in mixed species plantations of native trees in tropical Australia. Agrofor. Syst. 2013, 87, 233–250. [Google Scholar] [CrossRef]
- Chazdon, R.L.; Guarlguata, M.R. Natural regeneration as a tool for large-scale forest restoration in the tropics: Prospects and challenges. Biotropica 2016, 48, 716–730. [Google Scholar] [CrossRef]
- Zhang, C.L.; Li, X.W.; Chen, Y.Q.; Zhao, J.; Wan, S.Z.; Lin, Y.B.; Fu, S.L. Effects of Eucalyptus litter and roots on the establishment of native tree species in Eucalyptus plantations in South China. For. Ecol. Manag. 2016, 375, 76–83. [Google Scholar] [CrossRef]
- Li, W.H. Degradation and restoration of forests ecosystems in China. For. Ecol. Manag. 2004, 201, 33–41. [Google Scholar] [CrossRef]
- Rodriguez-Loinaz, G.; Amezaga, I.; Onaindia, M. Use of native species to improve carbon sequestration and contribute towards solving the environmental problems of the timberlands in Biscay, Northern Spain. J. Environ. Manag. 2013, 120, 18–26. [Google Scholar] [CrossRef] [PubMed]
- Robert, H. Conservation through intensification? The effects of plantations on natural forests. Ecol. Econ. 2014, 105, 204–210. [Google Scholar] [CrossRef]
- Pirard, R.; Secco, L.D.; Warman, R. Do timber plantations contribute to forest conservation? Environ. Sci. Policy 2016, 57, 122–130. [Google Scholar] [CrossRef]
- Wang, J.; Ren, H.; Yang, L.; Li, D.; Guo, Q. Soil seed banks in four 22-year-old plantations in South China: Implications for restoration. For. Ecol. Manag. 2009, 258, 2000–2006. [Google Scholar] [CrossRef]
- Bertacchi, M.I.F.; Amazonas, N.T.; Brancalion, P.H.S.; Brondani, G.E.; de Oliveira, A.C.S.; de Pascoa, M.A.R.; Rodrigues, R.R. Establishment of tree seedlings in the understory of restoration plantations: Natural regeneration and enrichment plantings. Restor. Ecol. 2016, 24, 100–108. [Google Scholar] [CrossRef]
- Carnevale, N.J.; Montagnini, F. Facilitating regeneration of secondary forests with the use of mixed and pure plantations of indigenous tree species. For. Ecol. Manag. 2002, 163, 217–227. [Google Scholar] [CrossRef]
- Dupuy, J.M.; Chazdon, R.L. Interacting effects of canopy gap, understory vegetation and leaf litter on tree seedling recruitment and composition in tropical secondary forests. For. Ecol. Manag. 2008, 255, 3716–3725. [Google Scholar] [CrossRef]
- Hu, Z.H.; He, Z.M.; Huang, Z.Q.; Fan, S.H.; Yu, Z.P.; Wang, M.H.; Zhou, X.H.; Fang, C.M. Effects of harvest residue management on soil carbon and nitrogen processes in a Chinese fir plantation. For. Ecol. Manag. 2014, 326, 163–170. [Google Scholar] [CrossRef]
- Chen, C.Y.; Liao, L.P.; Wang, S.L. Ecology of Chinese Fir Plantation Forest; China Science Publishing House: Beijing, China, 2000. [Google Scholar]
- Yang, Y.S.; Guo, J.F.; Chen, G.S.; Lin, P. Litterfall, nutrient return, and leaf-litter decomposition in four plantations compared with a natural forest in subtropical China. Ann. For. Sci. 2004, 61, 465–476. [Google Scholar] [CrossRef]
- Bi, J.; Blanco, J.A.; Seely, B.; Kimmins, J.P.; Ding, Y.; Welham, C. Yield decline in Chinese fir plantations: A simulation investigation with implications for model complexity. Can. J. For. Res. 2007, 37, 1615–1630. [Google Scholar] [CrossRef]
- Ma, X.Q.; Heal, K.V.; Liu, A.Q.; Jarvis, P.G. Nutrient cycling and distribution in different-aged plantations of Chinese fir in southern China. For. Ecol. Manag. 2007, 243, 61–74. [Google Scholar] [CrossRef]
- Chen, L.C.; Wang, S.L.; Wang, P.; Kong, C.H. Autoinhibition and soil allelochemical (cyclic dipeptide) levels in replanted Chinese fir (Cunninghamia lanceolata) plantations. Plant Soil 2014, 374, 793–801. [Google Scholar] [CrossRef]
- Zhu, N.H.; Yao, J.Y.; Zhou, G.Y.; Li, J.X.; Li, C.; Jing, M.Q.; Zhao, S.M. Study on natural vegetation regeneration from over maturity plantation of Chinese fir. J. Cent. South Univ. For. Technol. 2011, 31, 106–110. [Google Scholar] [CrossRef]
- Luo, J.; Tian, Y.X.; Zhou, X.L.; Chen, J.H.; Zeng, Z.Q.; Li, X.Q.; Yao, M. Studies on the characteristics of soil seed banks under main forest types of close-to-nature forest management in Hunan. J. Cent. South Univ. For. Technol. 2014, 34, 56–61. [Google Scholar] [CrossRef]
- Walck, J.L.; Hidayati, S.N.; Dixon, K.W.; Thompson, K.; Poschlod, P. Climate change and plant regeneration from seed. Glob. Chang. Biol. 2011, 17, 2145–2161. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; Fan, B.; Yu, X.J. Litter production, leaf litter decomposition and nutrient return in Cunninghamia lanceolata plantations in south China: Effect of planting conifers with broadleaved species. Plant Soil 2007, 297, 201–211. [Google Scholar] [CrossRef]
- Wang, Q.K.; Wang, S.L.; Xu, G.B.; Fan, B. Conversion of secondary broadleaved forest into Chinese fir plantation alters litter production and potential nutrient returns. Plant Ecol. 2010, 209, 269–278. [Google Scholar] [CrossRef]
- Facelli, J.M. Multiple indirect effects of plant litter affect the establishment of woody seedlings in old fields. Ecology 1994, 75, 1727–1735. [Google Scholar] [CrossRef]
- Nakamura, T. Ecological relationship between seedling emergence and litter cover in the earliest stage of plant succession on sandy soil. Ecol. Res. 1996, 11, 105–110. [Google Scholar] [CrossRef]
- Eckstein, R.L.; Donath, T.W. Interactions between litter and water availability affect seedling emergence in four familial pairs of floodplain species. J. Ecol. 2005, 93, 807–816. [Google Scholar] [CrossRef]
- Puhlick, J.J.; Laughlin, D.C.; Moore, M.M. Factors influencing ponderosa pine regeneration in the southwestern USA. For. Ecol. Manag. 2012, 264, 10–19. [Google Scholar] [CrossRef]
- Loydi, A.; Donath, T.W.; Otte, A.; Eckstein, R.L. Negative and positive interactions among plants: Effects of competitors and litter on seedling emergence and growth of forest and grassland species. Plant Soil 2015, 17, 667–675. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, P.; Antunes, C.; Pinho, P.; Máguas, C.; Correia, O. Natural regeneration of Pinus pinaster, and Eucalyptus globulus, from plantation into adjacent natural habitats. For. Ecol. Manag. 2016, 378, 91–102. [Google Scholar] [CrossRef]
- Zhang, R.; Hu, X.; Baskin, J.M.; Baskin, C.C.; Wang, Y. Effects of litter on seedling emergence and seed persistence of three common species on the Loess Plateau in Northwestern China. Front. Plant Sci. 2017, 8, 103. [Google Scholar] [CrossRef] [PubMed]
- Kostel-Hughes, F.; Young, T.P.; Wehr, J.D. Effects of leaf litter depth on the emergence and seedling growth of deciduous forest tree species in relation to seed size. J. Torrey Bot. Soc. 2005, 132, 50–61. [Google Scholar] [CrossRef]
- Quddus, M.S.; Bellairs, S.M.; Wurm, P.A.S. Acacia holosericea (Fabaceae) litter has allelopathic and physical effects on mission grass (Cenchrus pedicellatus and C. polystachios) (Poaceae) seedling establishment. Aust. J. Bot. 2014, 62, 189–195. [Google Scholar] [CrossRef]
- Rotundo, J.L.; Aguiar, M.R. Litter effects on plant regeneration in arid lands: A complex balance between seed retention, seed longevity and soil–seed contact. J. Ecol. 2005, 93, 829–838. [Google Scholar] [CrossRef]
- Donath, T.W.; Eckstein, R.L. Grass and oak litter exert different effects on seedling emergence of herbaceous perennials from grasslands and woodlands. J. Ecol. 2008, 96, 272–280. [Google Scholar] [CrossRef]
- Ruprecht, E.; Józsa, J.; Ölvedi, T.B.; Simon, J. Differential effects of several “litter” types on the germination of dry grassland species. J. Veg. Sci. 2010, 21, 1069–1081. [Google Scholar] [CrossRef]
- Ellsworth, J.W.; Harrington, R.A.; Fownes, J.H. Seedling emergence, growth, and allocation of Oriental bittersweet: Effects of seed input, seed bank, and forest floor litter. For. Ecol. Manag. 2004, 190, 255–264. [Google Scholar] [CrossRef]
- Peterson, C.J.; Facelli, J.M. Contrasting germination and seedling growth of Betula alleghaniensis and Rhus typhina subjected to various amounts and types of plant litter. Am. J. Bot. 1992, 79, 1209–1216. [Google Scholar] [CrossRef]
- Bosy, J.T.; Reader, R.J. Mechanisms underlying the suppression of forb seedling emergence by grass (Poa pratensis) litter. Funct. Ecol. 1995, 9, 635–639. [Google Scholar] [CrossRef]
- Olson, B.E.; Wallander, R.T. Effects of invasive forb litter on seed germination, seedling growth and survival. Basic Appl. Ecol. 2002, 3, 309–317. [Google Scholar] [CrossRef]
- Muturi, G.M.; Poorter, L.; Bala, P.; Mohren, G.M.J. Unleached Prosopis litter inhibits germination but leached stimulates seedling growth of dry woodland species. J. Arid Environ. 2017, 138, 44–50. [Google Scholar] [CrossRef]
- Hovstad, K.A.; Ohlson, M. Physical and chemical effects of litter on plant establishment in semi-natural grasslands. Plant Ecol. 2008, 196, 251–260. [Google Scholar] [CrossRef]
- Myster, R.W. Contrasting litter effects on old field tree germination and emergence. Plant Ecol. 1994, 114, 169–174. [Google Scholar] [CrossRef]
- Stinchcombe, J.R.; Schmitt, J. Ecosystem engineers as selective agents: The effects of leaf litter on emergence time and early growth in Impatiens capensis. Ecol. Lett. 2006, 9, 258–270. [Google Scholar] [CrossRef] [PubMed]
- Loydi, A.; Eckstein, R.L.; Otte, A.; Donath, T.W. Effects of litter on seedling establishment in natural and semi-natural grasslands: A meta-analysis. J. Ecol. 2013, 101, 454–464. [Google Scholar] [CrossRef]
- Zhuang, Z.; Li, Y.J.; Liu, Q.Q.; Yang, Z.; Liu, B.; Liu, A.Q. Effects of Chinese fir litter extracts on the seed germination and seedlings. J. For. Environ. 2017, 37, 29–33. [Google Scholar] [CrossRef]
- Xia, Z.C.; Kong, C.H.; Chen, L.C.; Wang, S.L. Allelochemical-mediated soil microbial community in long-term monospecific Chinese fir forest plantations. Appl. Soil Ecol. 2015, 96, 52–59. [Google Scholar] [CrossRef]
- Yang, Y.S.; Xie, J.S.; Chen, Y.X.; He, Z.M.; Yu, B.N. Amount, composition and seasonal dynamics of litterfall in mixed forest of Cunninghamia Lanceolata and Tsoongiodenaron odorum. Sci. Silvae Sin. 2001, 37, 30–34. [Google Scholar] [CrossRef]
- Wen, Y.G.; Wei, S.Z.; Qing, W.M. Analysis on the litter-fall dynamics of Chinese fir plantation and its correlation to climatic factors in North Guangxi. Acta Ecol. Sin. 1990, 10, 367–372. [Google Scholar] [CrossRef]
- Gao, S.L.; He, Z.M.; Huang, Z.Q.; Lin, S.Z.; Liu, Z.M.; Zheng, L.J. Amount, composition and seasonal dynamics of litterfall in different ages of Cunninghamia lanceolata plantations. Aata Agric. Univ. Jiangxiensis 2015, 37, 638–644. [Google Scholar] [CrossRef]
- Barritt, A.R.; Facelli, J.M. Effects of Casuarina pauper litter and grove soil on emergence and growth of understorey species in arid lands of South Australia. J. Arid Environ. 2001, 49, 569–579. [Google Scholar] [CrossRef]
- Liu, G.X.; Wan, L.Q.; He, F.; Tong, Z.Y.; Liu, Z.K.; Li, X.L. Effects of litter, seed position, and water availability on establishment of seedling for two semiarid grass species. Plant Ecol. 2016, 217, 277–287. [Google Scholar] [CrossRef]
- Harris, M.R.; Lamb, D.; Erskine, P.D. An investigation into the possible inhibitory effects of white cypress pine (Callitris glaucophylla) litter on the germination and growth of associated ground cover species. Aust. J. Bot. 2003, 51, 93–102. [Google Scholar] [CrossRef]
- Amatangelo, K.L.; Dukes, J.S.; Field, C.B. Responses of a California annual grassland to litter manipulation. J. Veg. Sci. 2008, 19, 605–612. [Google Scholar] [CrossRef]
- Yanes, C.V.; Segovia, A.O. Effects of litter from a tropical rainforest on tree seed germination and establishment under controlled conditions. Tree Physiol. 1992, 11, 391–400. [Google Scholar] [CrossRef]
- Burgos, J.V.; Barradas, M.C.D.; Zunzunegui, M. Effects of Pinus pinea litter on seed germination and seedling performance of three Mediterranean shrub species. Plant Growth Regul. 2012, 66, 285–292. [Google Scholar] [CrossRef]
- Facelli, J.M.; Pickett, S.T.A. Plant litter: Light interception and effects on an old-field plant community. Ecology 1991, 72, 1024–1031. [Google Scholar] [CrossRef]
- Qi, S.S.; Dai, Z.C.; Miao, S.L.; Zhai, D.L.; Si, C.C.; Huang, P.; Wang, R.P.; Du, D.L. Light limitation and litter of an invasive clonal plant, Wedelia trilobata, inhibit its seedling recruitment. Ann. Bot. 2014, 114, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Enquist, B.J.; Niklas, K.J. Global allocation rules for patterns of biomass partitioning in seed plants. Science 2006, 295, 1517–1520. [Google Scholar] [CrossRef] [PubMed]
- Liu, B.; Liu, Z.M.; Wang, L.X.; Wang, Z.N. Responses of rhizomatous grass Phragmites communis to wind erosion: Effects on biomass allocation. Plant Soil 2014, 380, 389–398. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Model | df | M.S. | F | p | Model | df | M.S. | F | p |
|---|---|---|---|---|---|---|---|---|---|
| Emergence | Leaf mass | ||||||||
| Amount | 3 | 2252.37 | 49.99 | 0.000 | Amount | 3 | 1190.252 | 7.112 | 0.001 |
| Type | 1 | 84.100 | 1.870 | 0.181 | Type | 1 | 1.770 | 0.011 | 0.919 |
| Amount × Type | 3 | 12.100 | 0.270 | 0.848 | Amount × Type | 3 | 124.838 | 0.746 | 0.533 |
| Survival | Total mass | ||||||||
| Amount | 3 | 3470.53 | 76.69 | 0.000 | Amount | 3 | 2812.985 | 8.756 | 0.000 |
| Type | 1 | 67.60 | 1.490 | 0.231 | Type | 1 | 19.101 | 0.059 | 0.809 |
| Amount × Type | 3 | 15.60 | 0.350 | 0.793 | Amount × Type | 3 | 252.388 | 0.786 | 0.511 |
| Root length | Root-mass ratio | ||||||||
| Amount | 3 | 68.816 | 5.805 | 0.003 | Amount | 3 | 146.160 | 16.516 | 0.000 |
| Type | 1 | 2.973 | 0.251 | 0.620 | Type | 1 | 18.150 | 2.051 | 0.162 |
| Amount × Type | 3 | 1.434 | 0.121 | 0.947 | Amount × Type | 3 | 12.962 | 1.465 | 0.243 |
| Stem length | Stem-mass ratio | ||||||||
| Amount | 3 | 4.693 | 44.789 | 0.000 | Amount | 3 | 43.210 | 7.864 | 0.000 |
| Type | 1 | 0.021 | 0.201 | 0.657 | Type | 1 | 0.200 | 0.036 | 0.850 |
| Amount × Type | 3 | 0.056 | 0.535 | 0.662 | Amount × Type | 3 | 5.007 | 0.911 | 0.447 |
| Seedling height | Leaf-mass ratio | ||||||||
| Amount | 3 | 11.762 | 8.198 | 0.000 | Amount | 3 | 36.173 | 3.427 | 0.029 |
| Type | 1 | 0.065 | 0.046 | 0.832 | Type | 1 | 14.552 | 1.379 | 0.249 |
| Amount ×Type | 3 | 1.400 | 0.976 | 0.417 | Amount ×Type | 3 | 3.024 | 0.286 | 0.835 |
| Root mass | Root/shoot | ||||||||
| Amount | 3 | 317.197 | 12.928 | 0.000 | Amount | 3 | 0.039 | 16.697 | 0.000 |
| Type | 1 | 3.234 | 0.132 | 0.719 | Type | 1 | 0.004 | 1.608 | 0.214 |
| Amount × Type | 3 | 16.020 | 0.653 | 0.587 | Amount × Type | 3 | 0.002 | 1.034 | 0.391 |
| Stem mass | Robustness | ||||||||
| Amount | 3 | 4.833 | 2.301 | 0.097 | Amount | 3 | 22.473 | 24.392 | 0.000 |
| Type | 1 | 1.542 | 0.734 | 0.398 | Type | 1 | 0.521 | 0.565 | 0.458 |
| Amount × Type | 3 | 0.890 | 0.424 | 0.737 | Amount × Type | 3 | 0.438 | 0.476 | 0.702 |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, B.; Daryanto, S.; Wang, L.; Li, Y.; Liu, Q.; Zhao, C.; Wang, Z. Excessive Accumulation of Chinese Fir Litter Inhibits Its Own Seedling Emergence and Early Growth—A Greenhouse Perspective. Forests 2017, 8, 341. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090341
Liu B, Daryanto S, Wang L, Li Y, Liu Q, Zhao C, Wang Z. Excessive Accumulation of Chinese Fir Litter Inhibits Its Own Seedling Emergence and Early Growth—A Greenhouse Perspective. Forests. 2017; 8(9):341. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090341
Chicago/Turabian StyleLiu, Bo, Stefani Daryanto, Lixin Wang, Yanjuan Li, Qingqing Liu, Chong Zhao, and Zhengning Wang. 2017. "Excessive Accumulation of Chinese Fir Litter Inhibits Its Own Seedling Emergence and Early Growth—A Greenhouse Perspective" Forests 8, no. 9: 341. https://0-doi-org.brum.beds.ac.uk/10.3390/f8090341