Comparing Methods for Monitoring Establishment of the Emerald Ash Borer (Agrilus planipennis, Coleoptera: Buprestidae) Egg Parasitoid Oobius agrili (Hymenoptera: Encyrtidae) in Maryland, USA

Abstract
:1. Introduction
2. Materials and Methods
2.1. Parasitoid Releases
2.2. Sampling Methods
2.3. Data Analyses
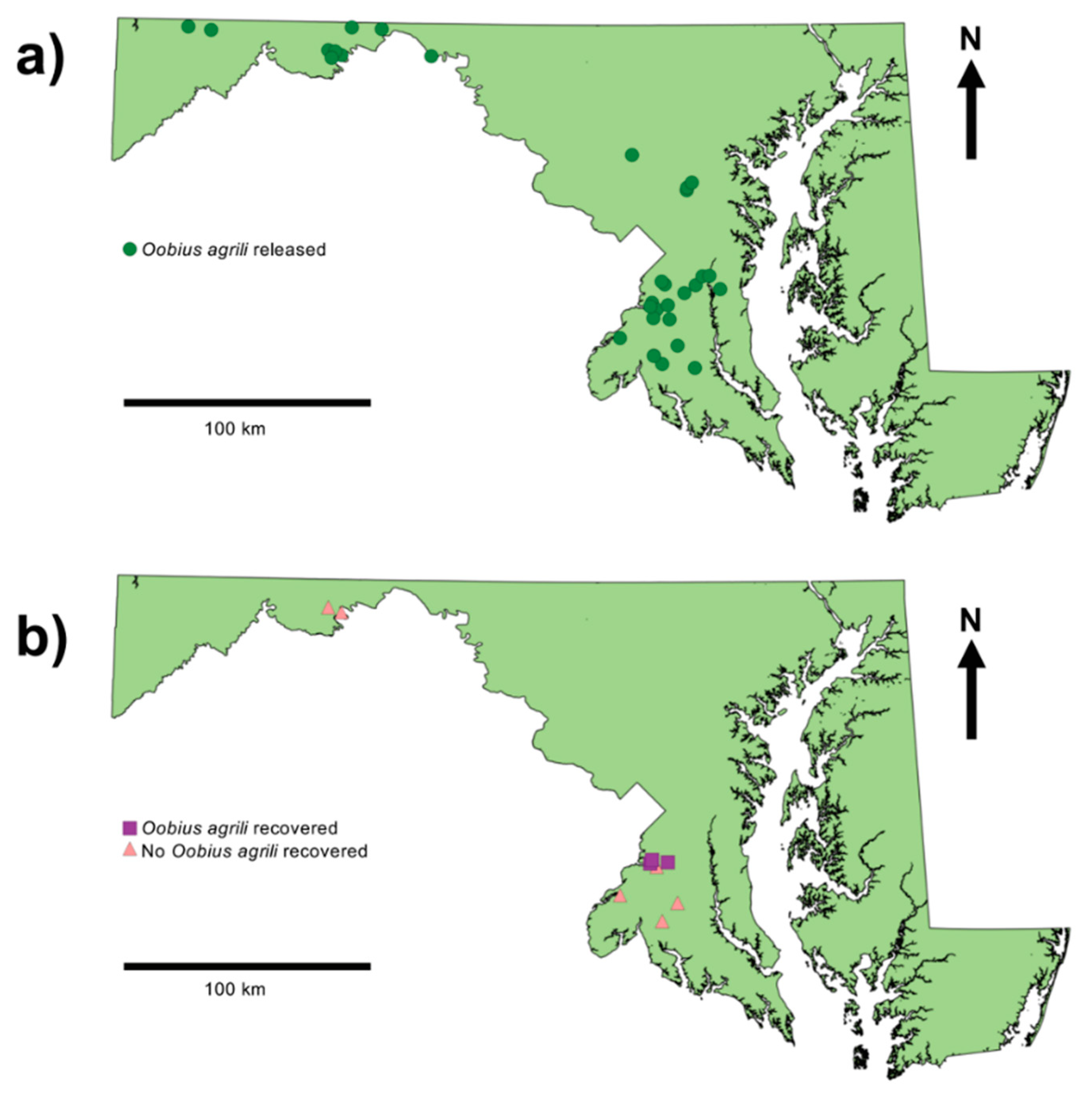
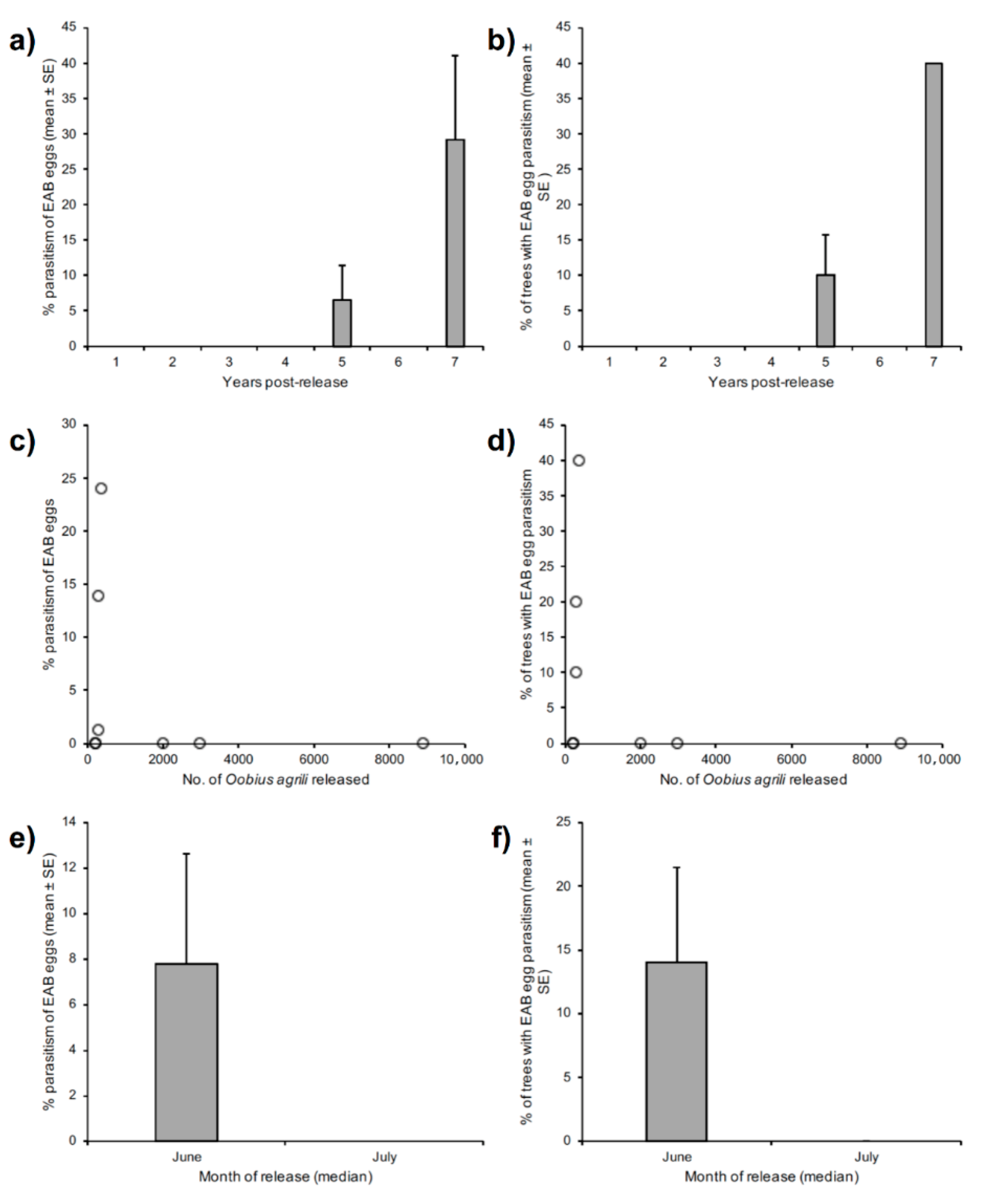
3. Results
3.1. Summary
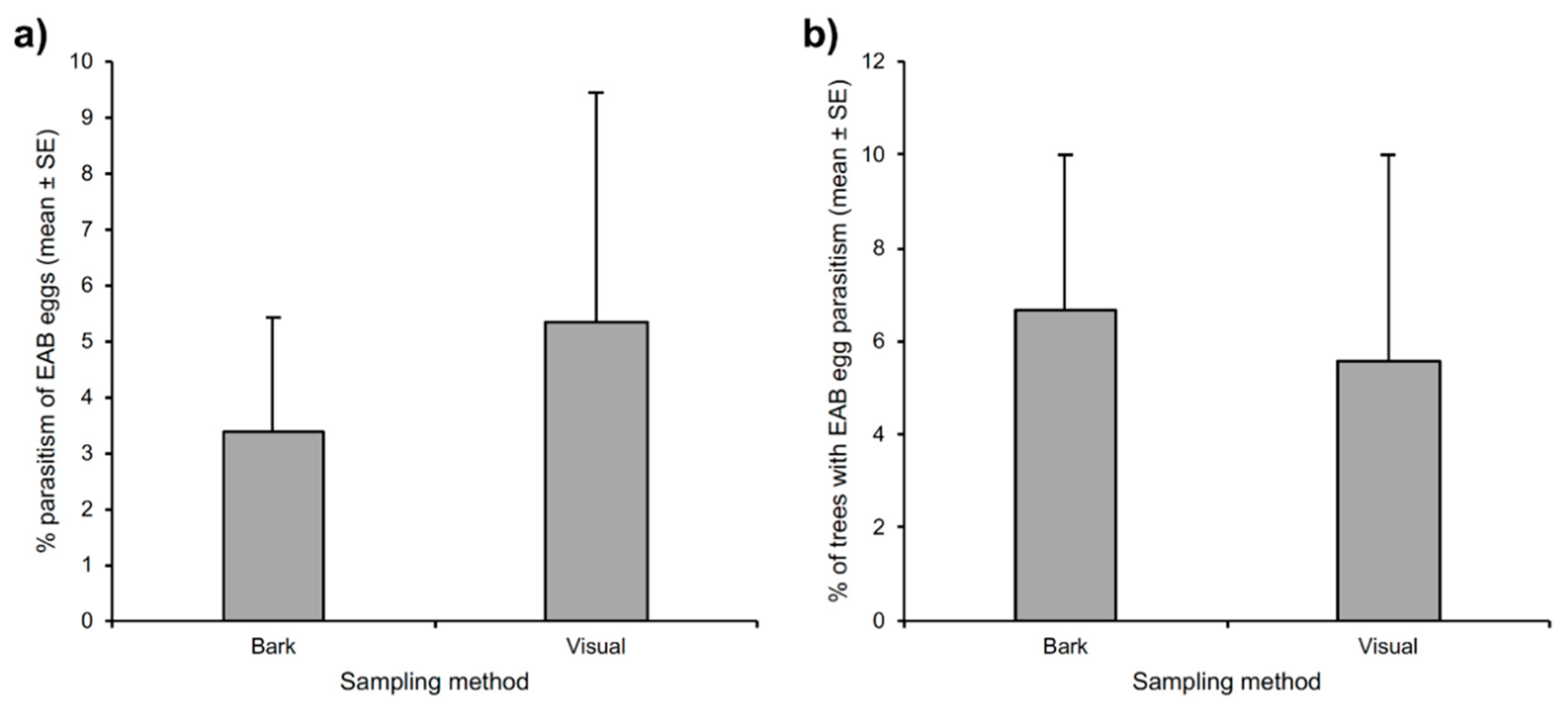
3.2. Comparison of Sampling Method Efficiency
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Langor, D.W.; DeHass, L.J.; Foottit, R.G. Diversity of non-native terrestrial arthropods on woody plants in Canada. Biol. Invasions 2009, 11, 5–19. [Google Scholar] [CrossRef]
- Lovett, G.M.; Weiss, M.; Liebhold, A.M.; Holmes, T.P.; Leung, B.; Lambert, K.F.; Orwig, D.A.; Campbell, F.T.; Rosenthal, J.; McCullough, D.G.; et al. Nonnative forest insects and pathogens in the United States: Impacts and policy options. Ecol. Appl. 2016, 26, 1437–1455. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wan, F.H.; Yang, N.W. Invasion and management of agricultural alien insects in China. Annu. Rev. Entomol. 2016, 61, 77–98. [Google Scholar] [CrossRef] [PubMed]
- Aukema, J.E.; McCullough, D.G.; Von Holle, B.; Liebhold, A.M.; Britton, K.; Frankel, S.J. Historical accumulation of nonindigenous forest pests in the continental United States. BioScience 2010, 60, 886–897. [Google Scholar] [CrossRef]
- Aukema, J.E.; Leung, B.; Kovacs, K.; Chivers, C.; Britton, K.O.; Englin, J.; Frankel, S.J.; Haight, R.G.; Holmes, T.P.; Liebhold, A.M.; et al. Economic impacts of non-native forest insects in the continental United States. PLoS ONE 2011, 6, e24587. [Google Scholar] [CrossRef] [PubMed]
- Gandhi, K.J.K.; Herms, D.A. North American arthropods at risk due to widespread Fraxinus mortality caused by the alien emerald ash borer. Biol. Invasions 2010, 12, 1839–1846. [Google Scholar] [CrossRef]
- Kovacs, K.F.; Haight, R.G.; McCullough, D.G.; Mercader, R.J.; Siegert, N.W.; Liebhold, A.M. Cost of potential emerald ash borer damage in U.S. communities, 2009–2019. Ecol. Econ. 2010, 69, 569–578. [Google Scholar] [CrossRef]
- Kovacs, K.F.; Mercader, R.J.; Haight, R.G.; Siegert, N.W.; McCullough, D.G.; Liebhold, A.M. The influence of satellite populations of emerald ash borer on projected economic costs in U.S. communities, 2010–2020. J. Environ. Manag. 2011, 92, 2170–2181. [Google Scholar] [CrossRef] [PubMed]
- Jennings, D.E.; Duan, J.J.; Bean, D.; Rice, K.A.; Williams, G.L.; Bell, S.K.; Shurtleff, A.S.; Shrewsbury, P.M. Effects of the emerald ash borer invasion on the community composition of arthropods associated with ash tree boles in Maryland, USA. Agric. For. Entomol. 2017, 19, 122–129. [Google Scholar] [CrossRef]
- Klooster, W.S.; Gandhi, K.J.K.; Long, L.C.; Perry, K.I.; Rice, K.B.; Herms, D.A. Ecological impacts of emerald ash borer in forests at the epicenter of the invasion in North America. Forests 2018, 9, 250. [Google Scholar] [CrossRef]
- Herms, D.A.; McCullough, D.G. Emerald ash borer invasion of North America: History, biology, ecology, impacts, and management. Annu. Rev. Entomol. 2014, 59, 13–30. [Google Scholar] [CrossRef] [PubMed]
- Siegert, N.W.; McCullough, D.G.; Liebhold, A.M.; Telewski, F.W. Dendrochronological reconstruction of the epicentre and early spread of emerald ash borer in North America. Divers. Distrib. 2014, 20, 847–858. [Google Scholar] [CrossRef]
- Bauer, L.S.; Duan, J.J.; Gould, J.R.; Van Driesche, R. Progress in the classical biocontrol of Agrilus planipennis Fairmaire (Coleoptera: Buprestidae) in North America. Can. Entomol. 2015, 147, 300–317. [Google Scholar] [CrossRef]
- McCullough, D.G.; Mercader, R.J.; Siegert, N.W. Developing and integrating tactics to slow ash (Oleaceae) mortality caused by emerald ash borer (Coleoptera: Buprestidae). Can. Entomol. 2015, 147, 349–358. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.G.; Gould, J.R. Progress and challenges of protecting North American ash trees from the emerald ash borer using biological control. Forests 2018, 9, 142. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Abell, K.J.; Lelito, J.P.; Van Driesche, R. Establishment and abundance of Tetrastichus planipennisi (Hymenoptera: Eulophidae) in Michigan: Potential for success in classical biocontrol of the invasive emerald ash borer (Coleoptera: Buprestidae). J. Econ. Entomol. 2013, 106, 1145–1154. [Google Scholar] [CrossRef] [PubMed]
- Hooie, N.A.; Wiggins, G.J.; Lambdin, P.L.; Grant, J.F.; Powell, S.D.; Lelito, J.P. Native parasitoids and recovery of Spathius agrili from areas of release against emerald ash borer in eastern Tennessee, USA. Biocontrol Sci. Technol. 2015, 25, 345–351. [Google Scholar] [CrossRef]
- Davidson, W.; Rieske, L.K. Establishment of classical biological control targeting emerald ash borer is facilitated by use of insecticides, with little effect on native arthropod communities. Biol. Control 2016, 101, 78–86. [Google Scholar] [CrossRef]
- Jennings, D.E.; Duan, J.J.; Bean, D.; Gould, J.R.; Rice, K.A.; Shrewsbury, P.M. Monitoring the establishment and abundance of introduced parasitoids of emerald ash borer larvae in Maryland, USA. Biol. Control 2016, 101, 138–144. [Google Scholar] [CrossRef]
- Johnson, T.D.; Lelito, J.P.; Pfammatter, J.A.; Raffa, K.F. Evaluation of tree mortality and parasitoid recoveries on the contiguous western invasion front of emerald ash borer. Agric. For. Entomol. 2016, 18, 327–339. [Google Scholar] [CrossRef]
- Abell, K.J.; Bauer, L.S.; Duan, J.J.; Van Driesche, R. Long-term monitoring of the introduced emerald ash borer (Coleoptera: Buprestidae) egg parasitoid, Oobius agrili (Hymenoptera: Encyrtidae), in Michigan, USA and evaluation of a newly developed monitoring technique. Biol. Control 2014, 79, 36–42. [Google Scholar] [CrossRef]
- Abell, K.J.; Bauer, L.S.; Miller, D.L.; Duan, J.J.; Van Driesche, R.G. Monitoring the establishment and flight phenology of parasitoids of emerald ash borer (Coleoptera: Buprestidae) in Michigan by using sentinel eggs and larvae. Fla. Entomol. 2016, 99, 667–672. [Google Scholar] [CrossRef]
- Parisio, M.S.; Gould, J.R.; Vandenberg, J.D.; Bauer, L.S.; Fierke, M.K. Evaluation of recovery and monitoring methods for parasitoids released against emerald ash borer. Biol. Control 2017, 106, 45–53. [Google Scholar] [CrossRef]
- Bauer, L.S.; Gould, J.R.; Duan, J.J.; Hansen, J.A.; Cossé, A.; Miller, D.; Abell, K.J.; Van Driesche, R.; Lelito, J.P.; Poland, T. Sampling methods for recovery of exotic emerald ash borer parasitoids after environmental release. In Proceedings of the 22nd USDA Interagency Research Forum on Invasive Species, Annapolis, MD, USA, 11–14 January 2011; McManus, K., Gottschalk, K.W., Eds.; United States Department of Agriculture, Forest Service: Morgantown, WV, USA, 2012; pp. 2–4. [Google Scholar]
- Duan, J.J.; Bauer, L.S.; Ulyshen, M.D.; Gould, J.R.; Van Driesche, R. Development of methods for the field evaluation of Oobius agrili (Hymenoptera: Encyrtidae) in North America, a newly introduced egg parasitoid of the emerald ash borer (Coleoptera: Buprestidae). Biol. Control 2011, 56, 170–174. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Hansen, J.A.; Abell, K.J.; Van Driesche, R. An improved method for monitoring parasitism and establishment of Oobius agrili (Hymenoptera: Encyrtidae), an egg parasitoid introduced for biological control of the emerald ash borer (Coleoptera: Buprestidae) in North America. Biol. Control 2012, 60, 255–261. [Google Scholar] [CrossRef]
- Jennings, D.E.; Duan, J.J.; Larson, K.M.; Lelito, J.P.; Shrewsbury, P.M. Evaluating a new method for monitoring the field establishment and parasitism of Oobius agrili (Hymenoptera: Encyrtidae), an egg parasitoid of emerald ash borer (Coleoptera: Buprestidae). Fla. Entomol. 2014, 97, 1263–1265. [Google Scholar] [CrossRef]
- Jennings, D.E.; Duan, J.J.; Abell, K.J.; Bauer, L.S.; Gould, J.R.; Shrewsbury, P.M.; Van Driesche, R.G. Life table evaluation of change in emerald ash borer populations due to biological control. In Biology and Control of Emerald Ash Borer; Van Driesche, R.G., Reardon, R.C., Eds.; United States Department of Agriculture, Forest Service: Morgantown, WV, USA, 2015; pp. 139–151. [Google Scholar]
- Bauer, L.S.; Liu, H. Oobius agrili (Hymenoptera: Encyrtidae), a solitary egg parasitoid of emerald ash borer from China. In Proceedings of the Emerald Ash Borer and Asian Longhorned Beetle Research and Technology Development Meeting, Cincinnati, OH, USA, 29 October–2 November 2006; Mastro, V., Lance, D., Reardon, R., Parra, G., Eds.; United States Department of Agriculture, Forest Service: Morgantown, WV, USA, 2007; pp. 63–64. [Google Scholar]
- Abell, K.J.; Poland, T.; Cossé, A.; Bauer, L. Trapping techniques for emerald ash borer and its introduced parasitoids. In Biology and Control of Emerald Ash Borer; Van Driesche, R.G., Reardon, R.C., Eds.; United States Department of Agriculture, Forest Service: Morgantown, WV, USA, 2015; pp. 113–127. [Google Scholar]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2016; Available online: http://www.r-project.org (accessed on 15 November 2016).
- Abell, K.J.; Duan, J.J.; Shrewsbury, P.M. Determining optimal parasitoid release timing for the biological control of emerald ash borer (Coleoptera: Buprestidae). Fla. Entomol. 2018. under review. [Google Scholar]
- Hoban, J.; Duan, J.J.; Hough-Goldstein, J. Effects of temperature and photoperiod on the reproductive biology and diapause of Oobius agrili (Hymenoptera: Encyrtidae), an egg parasitoid of emerald ash borer (Coleoptera: Buprestidae). Environ. Entomol. 2016, 45, 726–731. [Google Scholar] [CrossRef] [PubMed]
- Larson, K.M.; Duan, J.J. Differences in the reproductive biology and diapause of two congeneric species of egg parasitoids (Hymenoptera: Encyrtidae) from northeast Asia: Implications for biological control of the invasive emerald ash borer (Coleoptera: Buprestidae). Biol. Control 2016, 103, 39–45. [Google Scholar] [CrossRef]
- Wetherington, M.T.; Jennings, D.E.; Shrewsbury, P.M.; Duan, J.J. Climate variation alters the synchrony of host-parasitoid interactions. Ecol. Evol. 2017, 7, 8578–8587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, A.L.; Jennings, D.E.; Hooks, C.R.R.; Shrewsbury, P.M. Field surveys of egg mortality and indigenous egg parasitoids of the brown marmorated stink bug, Halyomorpha halys, in ornamental nurseries in the mid-Atlantic region of the USA. J. Pest Sci. 2017, 90, 1159–1168. [Google Scholar] [CrossRef] [PubMed]
- Hanks, L.M.; Gould, J.R.; Paine, T.D.; Millar, J.G.; Wang, Q. Biology and host relations of Avetianella longoi (Hymenoptera: Encyrtidae), an egg parasitoid of the eucalyptus longhorned borer (Coleoptera: Cerambycidae). Ann. Entomol. Soc. Am. 1995, 88, 666–671. [Google Scholar] [CrossRef]
- Duan, J.J.; Bauer, L.S.; Van Driesche, R.G. Emerald ash borer biocontrol in ash saplings: The potential for early stage recovery of North American ash trees. For. Ecol. Manag. 2017, 394, 64–72. [Google Scholar] [CrossRef]
- Margulies, E.; Bauer, L.; Ibáñez, I. Buying time: Preliminary assessment of biocontrol in the recovery of native forest vegetation in the aftermath of the invasive emerald ash borer. Forests 2017, 8, 369. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Year | No. of Releases | Mean (±SE) | Total | Earliest | Latest |
|---|---|---|---|---|---|
| 2009 | 2 | 182.0 ± 39.0 | 364 | 3 June | 1 July |
| 2010 | 5 | 145.2 ± 28.5 | 726 | 26 May | 22 June |
| 2011 | 16 | 167.0 ± 6.8 | 2672 | 25 May | 30 June |
| 2012 | 10 | 87.9 ± 36.9 | 2286 | 8 June | 14 September |
| 2013 | 10 | 181.1 ± 75.1 | 6518 | 30 May | 3 July |
| 2014 | 24 | 675.4 ± 96.2 | 16,210 | 20 June | 11 September |
| 2015 | 39 | 702.6 ± 62.0 | 27,400 | 18 June | 10 September |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jennings, D.E.; Duan, J.J.; Shrewsbury, P.M. Comparing Methods for Monitoring Establishment of the Emerald Ash Borer (Agrilus planipennis, Coleoptera: Buprestidae) Egg Parasitoid Oobius agrili (Hymenoptera: Encyrtidae) in Maryland, USA. Forests 2018, 9, 659. https://0-doi-org.brum.beds.ac.uk/10.3390/f9100659
Jennings DE, Duan JJ, Shrewsbury PM. Comparing Methods for Monitoring Establishment of the Emerald Ash Borer (Agrilus planipennis, Coleoptera: Buprestidae) Egg Parasitoid Oobius agrili (Hymenoptera: Encyrtidae) in Maryland, USA. Forests. 2018; 9(10):659. https://0-doi-org.brum.beds.ac.uk/10.3390/f9100659
Chicago/Turabian StyleJennings, David E., Jian J. Duan, and Paula M. Shrewsbury. 2018. "Comparing Methods for Monitoring Establishment of the Emerald Ash Borer (Agrilus planipennis, Coleoptera: Buprestidae) Egg Parasitoid Oobius agrili (Hymenoptera: Encyrtidae) in Maryland, USA" Forests 9, no. 10: 659. https://0-doi-org.brum.beds.ac.uk/10.3390/f9100659