First Isolation and Characterization of a Group C Banna Virus (BAV) from Anopheles sinensis Mosquitoes in Hubei, China


Abstract
:1. Introduction
2. Materials and Methods
2.1. Mosquito Collection, Treatment, and PCR Screening
2.2. Virus Isolation, Purification, and Electron Microscopy
2.3. Plaque Assay and Growth Kinetics
2.4. Genome Sequencing and Polymerase Chain Reaction (PCR) Confirmation
2.5. Bioinformatics Analyses
3. Results
3.1. BAV Detection and the Genome of BAV HB14-71-01
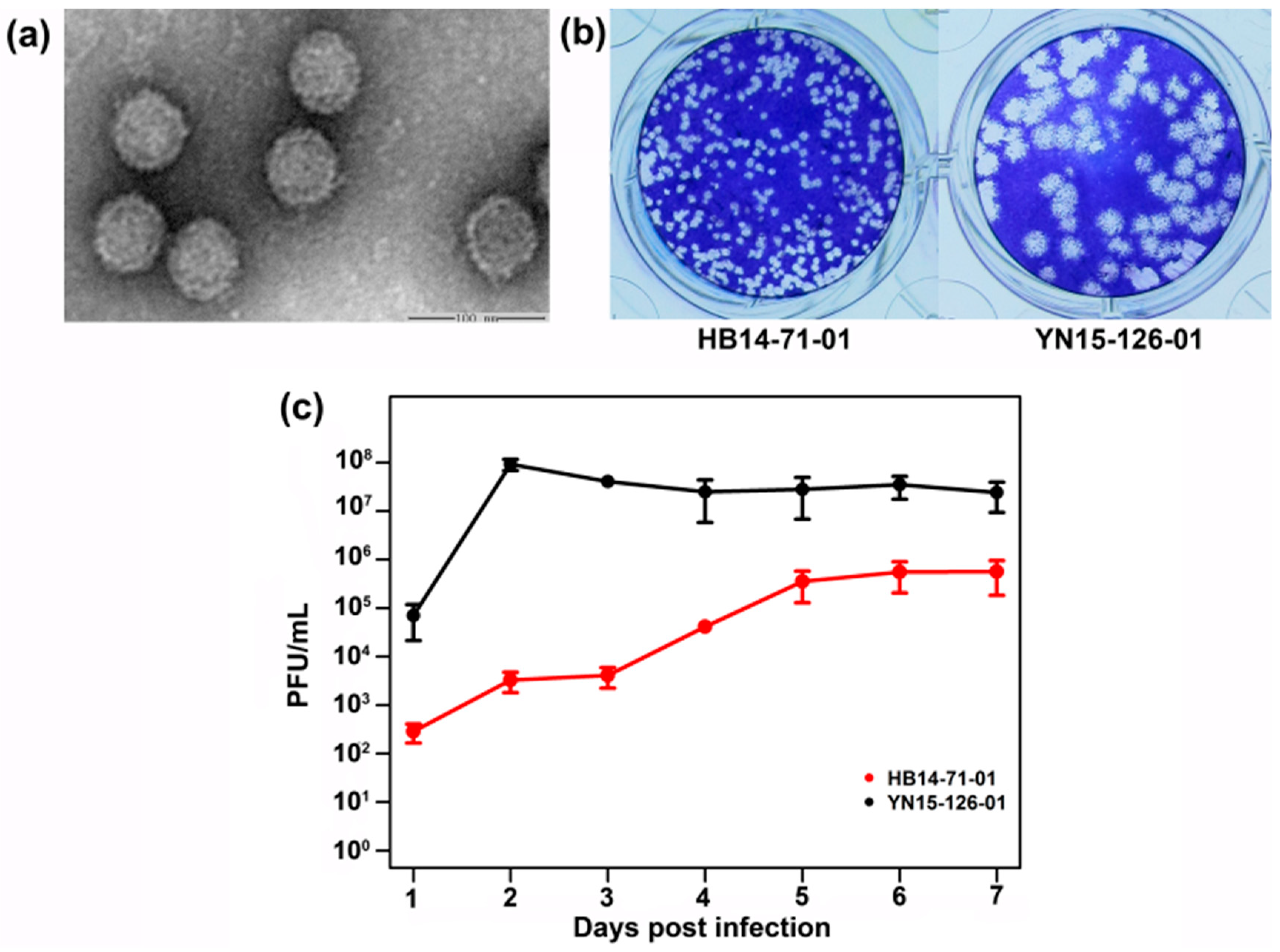
3.2. Viral Plaque and Growth Kinetics for BAV Strain HB14-71-01
3.3. Gene Sequence Comparison of BAVs with Full Genomes
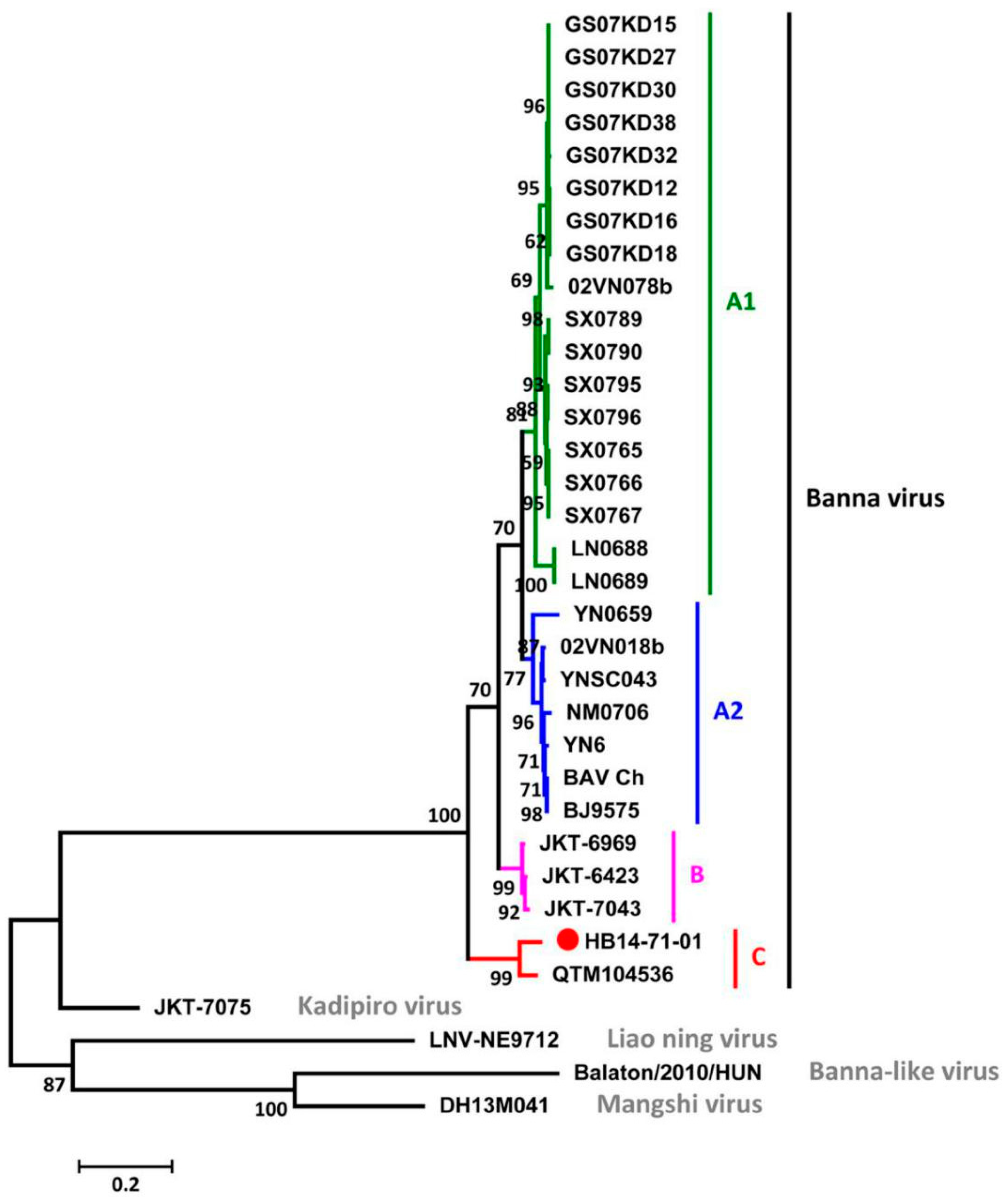
3.4. Phylogenetic Analysis among BAVs and Other Members of the Seadornavirus Genus
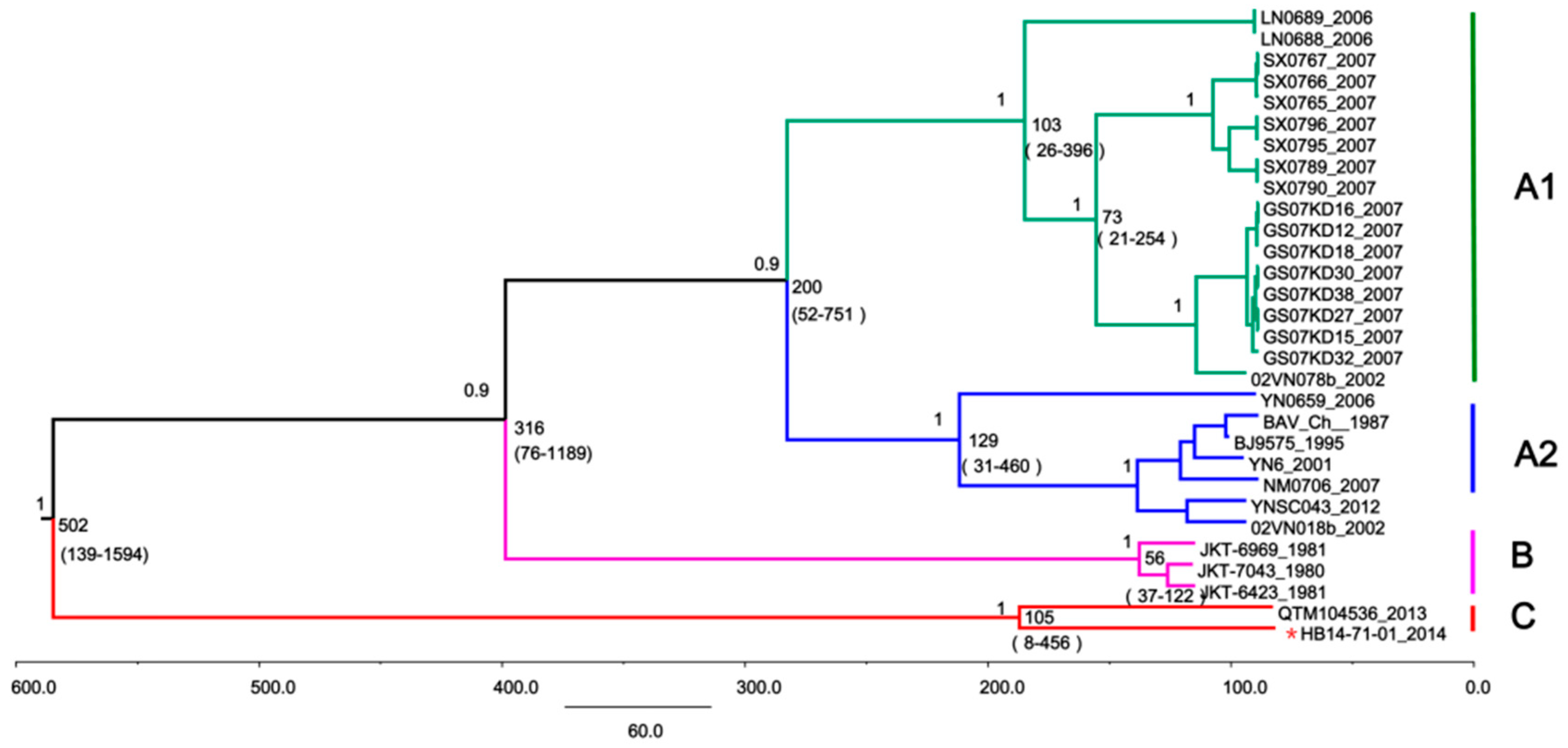
3.5. Time-Scaled Evolutionary Analysis of BAVs
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Attoui, H.; Billoir, F.; Biagini, P.; de Micco, P.; de Lamballerie, X. Complete sequence determination and genetic analysis of Banna virus and Kadipiro virus: Proposal for assignment to a new genus (Seadornavirus) within the family Reoviridae. J. Gen. Virol. 2000, 81, 1507–1515. [Google Scholar] [CrossRef] [PubMed]
- Xu, P.; Wang, Y.; Zuo, J.; Lin, J.; Xu, P. New orbiviruses isolated from patients with unknown fever and encephalitis in Yunnan province. Chin. J. Virol. 1990, 6, 27–33. [Google Scholar]
- Liu, H.; Li, M.H.; Zhai, Y.G.; Meng, W.S.; Sun, X.H.; Cao, Y.X.; Fu, S.H.; Wang, H.Y.; Xu, L.H.; Tang, Q.; et al. Banna virus, China, 1987. Emerg. Infect. Dis. 2010, 16, 514–517. [Google Scholar] [CrossRef] [PubMed]
- Nabeshima, T.; Thi Nga, P.; Guillermo, P.; Parquet Mdel, C.; Yu, F.; Thanh Thuy, N.; Minh Trang, B.; Tran Hien, N.; Sinh Nam, V.; Inoue, S.; et al. Isolation and molecular characterization of Banna virus from mosquitoes, Vietnam. Emerg. Infect. Dis. 2008, 14, 1276–1279. [Google Scholar] [CrossRef] [PubMed]
- Attoui, H.; Jaafar, F.M.; de Micco, P.; de Lamballerie, X. Coltiviruses and Seadornaviruses in North America, Europe, and Asia. Emerg. Infect. Dis. 2005, 11, 1673–1679. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, F.M.; Attoui, H.; Mertens, P.P.C.; de Micco, P.; de Lamballerie, X. Structural organization of an encephalitic human isolate of Banna virus (genus Seadornavirus, family Reoviridae). J. Gen. Virol. 2005, 86, 1147–1157. [Google Scholar] [CrossRef] [PubMed]
- Jaafar, F.M.; Attoui, H.; Bahar, M.W.; Siebold, C.; Sutton, G.; Mertens, P.P.C.; de Micco, P.; Stuart, D.I.; Grimes, J.M.; de Lamballerie, X. The structure and function of the outer coat protein VP9 of Banna virus. Structure 2005, 13, 17–28. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Gao, X.Y.; Fu, S.H.; Li, M.H.; Zhai, Y.G.; Meng, W.S.; Sun, X.X.; Wang, H.Y.; Lyu, Z.; Shen, X.X.; et al. Analysis on molecular genetic evolution of Banna virus based on the 12(th) segment. Zhonghua Liu Xing Bing Xue Za Zhi 2016, 37, 1277–1282. [Google Scholar] [PubMed]
- Liu, H.; Gao, X.Y.; Fu, S.H.; Li, M.H.; Zhai, Y.G.; Meng, W.S.; Sun, X.H.; Lv, Z.; Wang, H.Y.; Shen, X.X.; et al. Molecular evolution of emerging Banna virus. Infect. Genet. Evol. 2016, 45, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Li, Y.; Fu, S.; Lei, W.; Guo, X.; Feng, Y.; Gao, X.; Li, X.; Yang, Z.; Xu, Z.; et al. Genome sequencing and phylogenetic analysis of Banna virus (genus Seadornavirus, family Reoviridae) isolated from Culicoides. Sci. China Life Sci. 2017, 60, 1372–1382. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.X.; Fu, S.H.; Tian, Z.F.; He, Y.; Wang, H.Q.; Wang, H.Y.; Yang, H.M.; Tao, B.; Liang, G.D. Isolation and identification of Banna virus from mosquito for the first time in Inner Mongolia. Chin. J. Exp. Clin. Virol. 2009, 23, 106–108. [Google Scholar]
- Meng, W.S.; Zhang, J.B.; Sun, X.H.; Liu, Q.N.; Chen, Z.; Zhai, Y.G.; Fu, S.H.; Cao, Y.X.; Wang, H.Y.; Ding, J.; et al. Isolation and identification of arboviruses from mosquito pools in some regions of Liaoning province, China. Chin. J. Epidemiol. 2009, 1, 50–54. [Google Scholar]
- Sun, X.H.; Fu, S.H.; Wang, J.L.; Lü, X.J.; Wang, H.Q.; He, Y.; Zhai, Y.G.; Liang, G.D. First isolation of Banna virus in northwestern part of Yunnan province. Chin. J. Microbiol. Immunol. 2009, 29, 495–498. [Google Scholar]
- Zhai, Y.; Wang, H.; Xu, H.; Meng, W.; Chao, Y.; Fu, S.; Liang, G. Investigation on arboviruses in Tianshui and Longnan regions of Gansu province. Chin. J. Zoonoses 2008, 2, 95–99. [Google Scholar]
- Li, M.H.; Meng, W.S.; Fu, S.H.; Cheng, J.X.; Zhao, J.Y.; Kong, X.S.; Dai, P.F.; Liang, G.D. Arbovirus investigation in some regions of Shanxi province in 2007. Chin. J. Exp. Clin. Virol. 2009, 23, 32–34. [Google Scholar]
- Shi, C.; Liu, Y.; Hu, X.; Xiong, J.; Zhang, B.; Yuan, Z. A metagenomic survey of viral abundance and diversity in mosquitoes from hubei province. PLoS ONE 2015, 10, e0129845. [Google Scholar] [CrossRef] [PubMed]
- Billoir, F.; Attoui, H.; Simon, S.; Gallian, P.; de Micco, P.; De Lamballerie, X. Molecular diagnosis of group B coltiviruses infections. J. Virol. Methods 1999, 81, 39–45. [Google Scholar] [CrossRef]
- Abyss. Available online: http://www.bcgsc.ca/platform/bioinfo/software/abyss (accessed on 10 December 2018).
- SOAP. Available online: http://soap.genomics.org.cn/soapdenovo.html (accessed on 10 December 2017).
- BioEdit. Available online: http://www.mbio.ncsu.edu/bioedit/bioedit.html (accessed on 10 December 2017).
- Attoui, H.; Jaafar, F.M.; Belhouchet, M.; Tao, S.; Chen, B.; Liang, G.; Tesh, R.B.; de Micco, P.; de Lamballerie, X. Liao ning virus, a new Chinese seadornavirus that replicates in transformed and embryonic mammalian cells. J. Gen. Virol. 2006, 87, 199–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reuter, G.; Boros, Á.; Delwart, E.; Pankovics, P. Novel Seadornavirus (family Reoviridae) related to Banna virus in Europe. Arch. Virol. 2013, 158, 2163–2167. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, H.; He, Y.; Zhou, Y.; Meng, J.; Zhu, W.; Chen, H.; Liao, D.; Man, Y. Isolation and genetic characterization of Mangshi virus: A newly discovered Seadornavirus of the Reoviridae family found in Yunnan Province, China. PLoS ONE 2015, 10, e0143601. [Google Scholar] [CrossRef] [PubMed]
- MEGA. Available online: https://www.megasoftware.net/ (accessed on 10 December 2017).
- BEAST. Available online: http://beast.community/index.html (accessed on 3 April 2018).
- MrModeltest2. Available online: https://github.com/nylander/MrModeltest2/tree/master (accessed on 3 April 2018).
- Shi, M.; Lin, X.D.; Tian, J.H.; Chen, L.J.; Chen, X.; Li, C.X.; Qin, X.C.; Li, J.; Cao, J.P.; Eden, J.S.; et al. Redefining the invertebrate RNA virosphere. Nature 2016, 540, 539–543. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Virus | Isolate | Source | Location | Accession Number | Time of Collection | |
|---|---|---|---|---|---|---|
| 1st to 11th segments | 12th segment | |||||
| Banna virus | 02VN018b | Culex Annulus | Vietnam: Quang Binh | EU265683; EU265684; EU265685; EU265686; EU265687; EU265688; EU265689; EU265690; EU265691; EU265692; EU265693 | EU265694 | 2002 |
| 02VN078b | Cx. tritaeniorhynchus | Vietnam: Ha Tay | EU265695; EU265696; EU265697; EU265698; EU265699; EU265700; EU265701; EU265702; EU265703; EU265704 | EU265705 | 2002 | |
| BAV_CH | Human | China: Yunnan | AF168005; AF134526; AY549307; AY549308; AY549309; AF168006; AF052035; AF052034; AF052033; AF052032; AF052031 | AF052030 | 1987 | |
| BJ9575 | Unidentified mosquito | China: Beijing | AY568289 | 1995 | ||
| GS07-KD12 | Anopheles sinensis | China: Gansu | GQ331954 | 2007 | ||
| GS07-KD15 | Cx. tritaeniorhynchus | China: Gansu | GQ331955 | 2007 | ||
| GS07-KD16 | Cx. pipiens pallens | China: Gansu | GQ331956 | 2007 | ||
| GS07-KD18 | An. sinensis | China: Gansu | GQ331957 | 2007 | ||
| GS07-KD27 | Cx. tritaeniorhyn | China: Gansu | GQ331958 | 2007 | ||
| GS07-KD30 | Cx. pipiens pallens | China: Gansu | GQ331960 | 2007 | ||
| GS07-KD32 | Cx. pipiens pallens | China: Gansu | GQ331961 | 2007 | ||
| GS07-KD38 | Cx. pipiens pallens | China: Gansu | GQ331962 | 2007 | ||
| JKT-6423 | Cx. pseudovishnui | Indonesia: Java | NC_004198 | 1981 | ||
| JKT-6969 | Aedes vagus | Indonesia: Java | AF052008 | 1981 | ||
| JKT-7043 | Cx. pipiens pallens | Indonesia: Java | AF052024 | 1980 | ||
| LN0688 | An. sinensis | China: Liaoning | FJ217990 | 2006 | ||
| LN0689 | An. sinensis | China: Liaoning | FJ217991 | 2006 | ||
| NM0706 | Cx. modestus | China: Inner Mongolia | CQ331973 | 2007 | ||
| SX0765 | Cx. pipiens pallens | China: Shanxi | CQ331963 | 2007 | ||
| SX0766 | Cx. pipiens pallens | China: Shanxi | CQ331964 | 2007 | ||
| SX0767 | Ae. vexans | China: Shanxi | CQ331965 | 2007 | ||
| SX0789 | Ae. dorsalis | China: Shanxi | CQ331967 | 2007 | ||
| SX0790 | Ae. vexans | China: Shanxi | CQ331968 | 2007 | ||
| SX0795 | Cx. pipiens pallens | China: Shanxi | CQ331971 | 2007 | ||
| SX0796 | Cx. pipiens pallens | China: Shanxi | CQ331972 | 2007 | ||
| YN0659 | An. sinensis | China: Yunnan | FJ161965 | 2006 | ||
| YN6 | Unidentified mosquito | China: Yunnan | AY568290 | 2001 | ||
| YNSC043 | Culicoides sp. | China: Yunnan | KC954611; KC954612; KC954613; KC954614; KC954615; KC954616; KC954617; KC954618; KC954619; KC954620; KC954621 | KC954622 | 2004 | |
| HB14-71-01 | An. sinensis | China: Hubei | MH521264; MH521265; MH521266; MH521267; MH521268; MH521269; MH521270; MH521271; MH521272; MH521273; MH521274 | MH521275 | 2014 | |
| QTM104536 1 | Odonata | China: Hubei | KX884638; KX884639; KX884640; KX884641; KX884642; KX884643; KX884644; KX884645; KX884646; KX884647; KX884649 | KX884648 | 2013 | |
| Banna-like virus | Balaton/2010/HUN 2 | Cyprinus carpio | Hungary: Veszpre | JX947850 | 2010 | |
| Mangshi virus | DH13M041 | Cx. tritaeniorhynchus | China: Yunnan | KR349198 | 2013 | |
| Kadipiro virus | JKT-7075 | Cx. fuscocephalus | Indonesia: Java | NC_004199 | 1981 | |
| Liao ning virus | LNV-NE9712 | Ae. dorsalis | China: Liaoning | NC_007747 | 1997 |
| Banna Virus Isolates | VP1 (Pol/Core) | VP2 (T2/Core) | VP3 (Cap/Core) | VP4 (Outer Coat) | VP5 (NS) | VP6-(NS) | VP7-(NS) | VP8 (T13-Core) | VP9 (Outer Coat) | VP10 (Core) | VP11-(NS) | VP12-(NS) |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HB14-71-01 | ||||||||||||
| YNSC043 | 0.755 | 0.748 | 0.717 | 0.766 | 0.715 | 0.766 | 0.739 | 0.790 | 0.652 | 0.696 | 0.829 | 0.780 |
| 02VN078b | 0.757 | 0.744 | 0.704 | 0.760 | 0.697 | 0.761 | 0.744 | 0.772 | 0.639 | 0.698 | 0.815 | 0.778 |
| 02VN018b | 0.762 | 0.740 | 0.702 | 0.762 | 0.713 | 0.747 | 0.728 | 0.790 | 0.645 | 0.696 | 0.830 | 0.784 |
| JKT-6423 | 0.766 | 0.733 | 0.700 | 0.765 | 0.701 | 0.762 | 0.743 | 0.781 | 0.650 | 0.678 | 0.831 | 0.790 |
| QTM104536 | 0.881 | 0.879 | 0.904 | 0.834 | 0.895 | 0.875 | 0.908 | 0.879 | 0.848 | 0.798 | 0.907 | 0.925 |
| BAV_Ch | 0.757 | 0.721 | 0.661 | 0.672 | 0.645 | 0.679 | 0.736 | 0.788 | 0.652 | 0.695 | 0.827 | 0.775 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xia, H.; Liu, H.; Zhao, L.; Atoni, E.; Wang, Y.; Yuan, Z. First Isolation and Characterization of a Group C Banna Virus (BAV) from Anopheles sinensis Mosquitoes in Hubei, China. Viruses 2018, 10, 555. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100555
Xia H, Liu H, Zhao L, Atoni E, Wang Y, Yuan Z. First Isolation and Characterization of a Group C Banna Virus (BAV) from Anopheles sinensis Mosquitoes in Hubei, China. Viruses. 2018; 10(10):555. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100555
Chicago/Turabian StyleXia, Han, Hong Liu, Lu Zhao, Evans Atoni, Yujuan Wang, and Zhiming Yuan. 2018. "First Isolation and Characterization of a Group C Banna Virus (BAV) from Anopheles sinensis Mosquitoes in Hubei, China" Viruses 10, no. 10: 555. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100555