Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Ethics Statement
2.2. EV71 Virus and Cell Treatment
2.3. Animals
2.4. RNA Isolation
2.5. RNA Library Construction and Sequencing
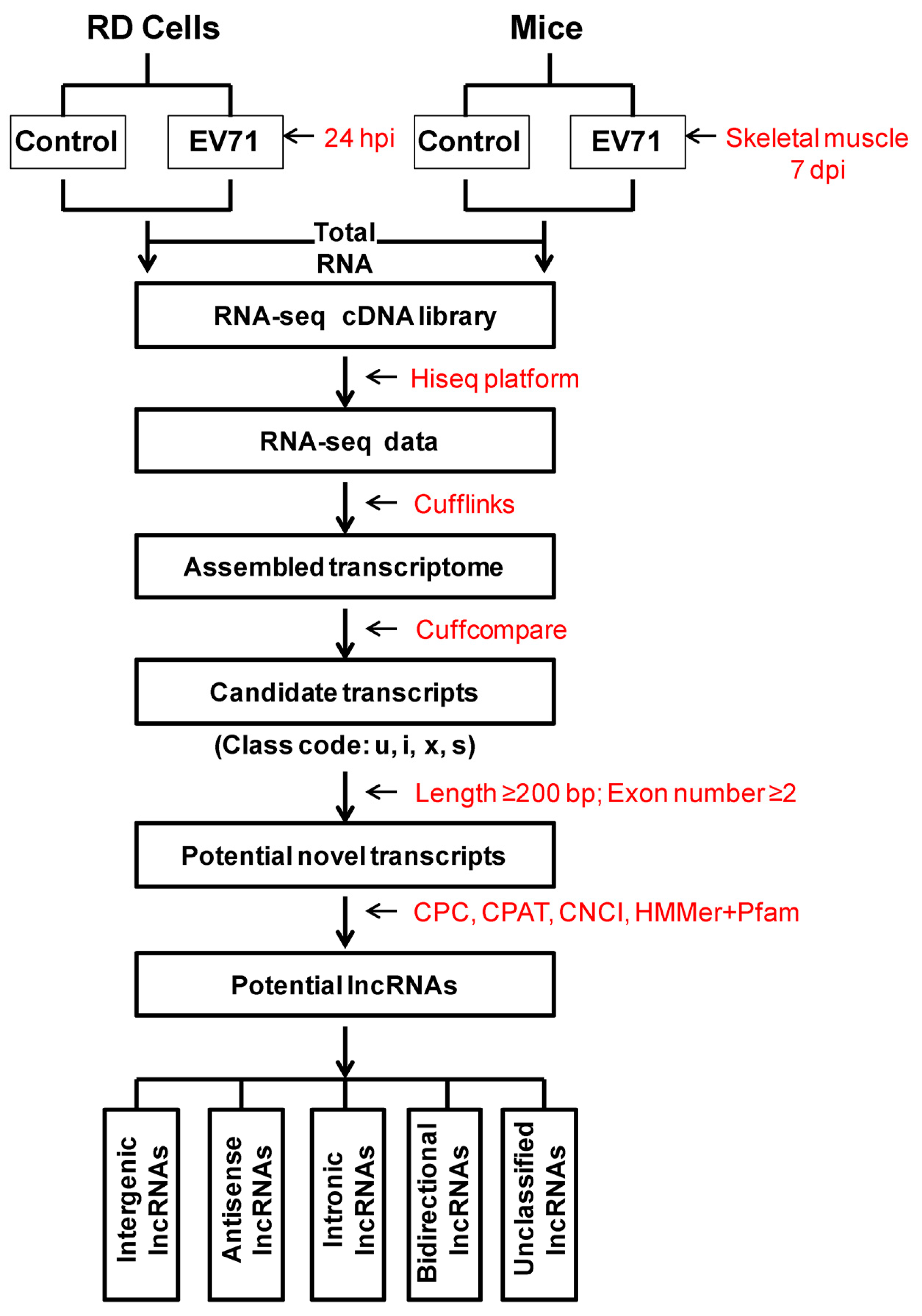
2.6. The Prediction and Classification of New lncRNAs
2.7. Identification of Differentially-Expressed Genes
2.8. Prediction of Target Genes of Differentially-Expressed lncRNAs
2.9. GO Annotations and KEGG Enrichment
2.10. qPCR Confirmation
2.11. Statistical Analysis
3. Results
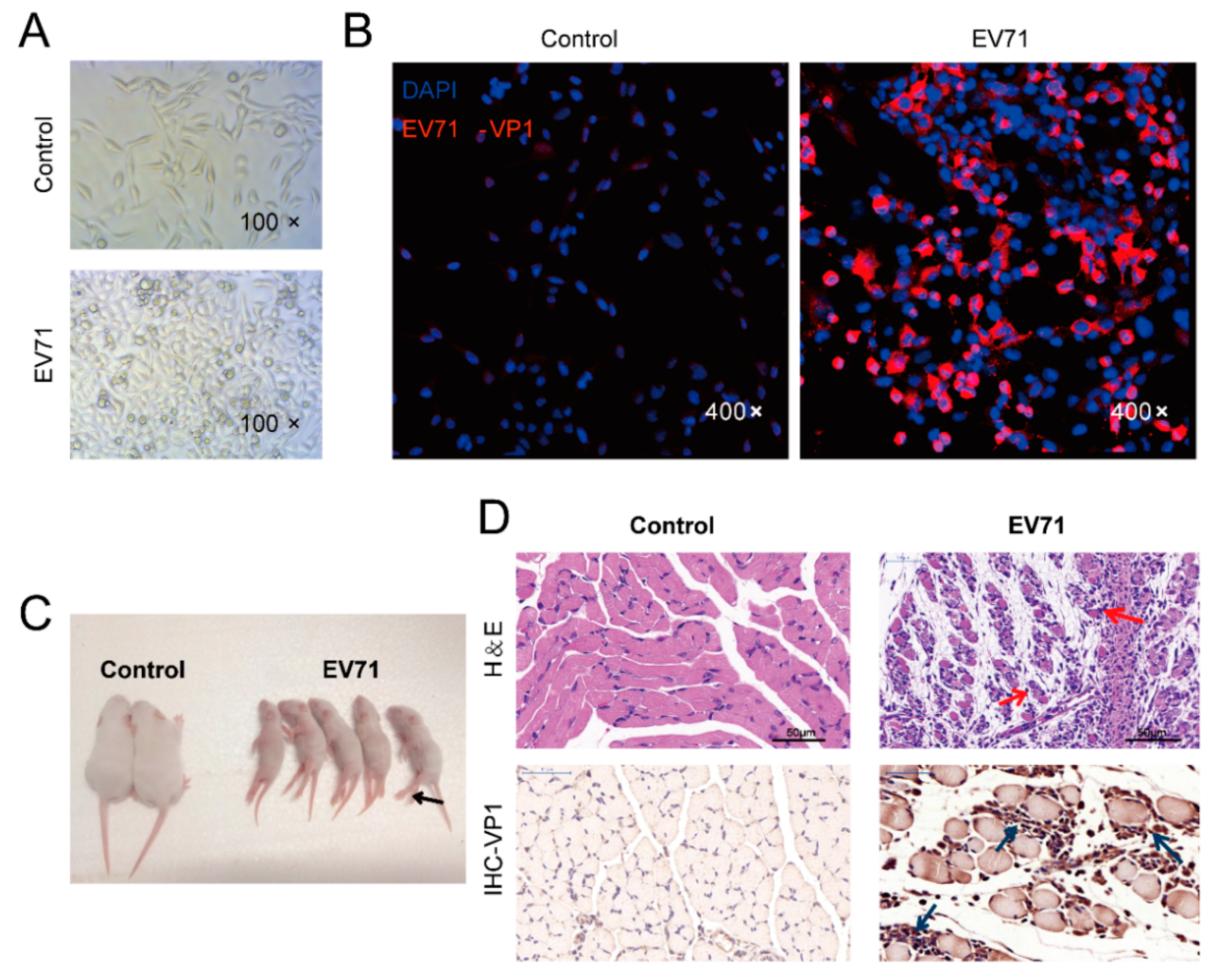
3.1. In Vitro and In Vivo Models of EV71 Infection
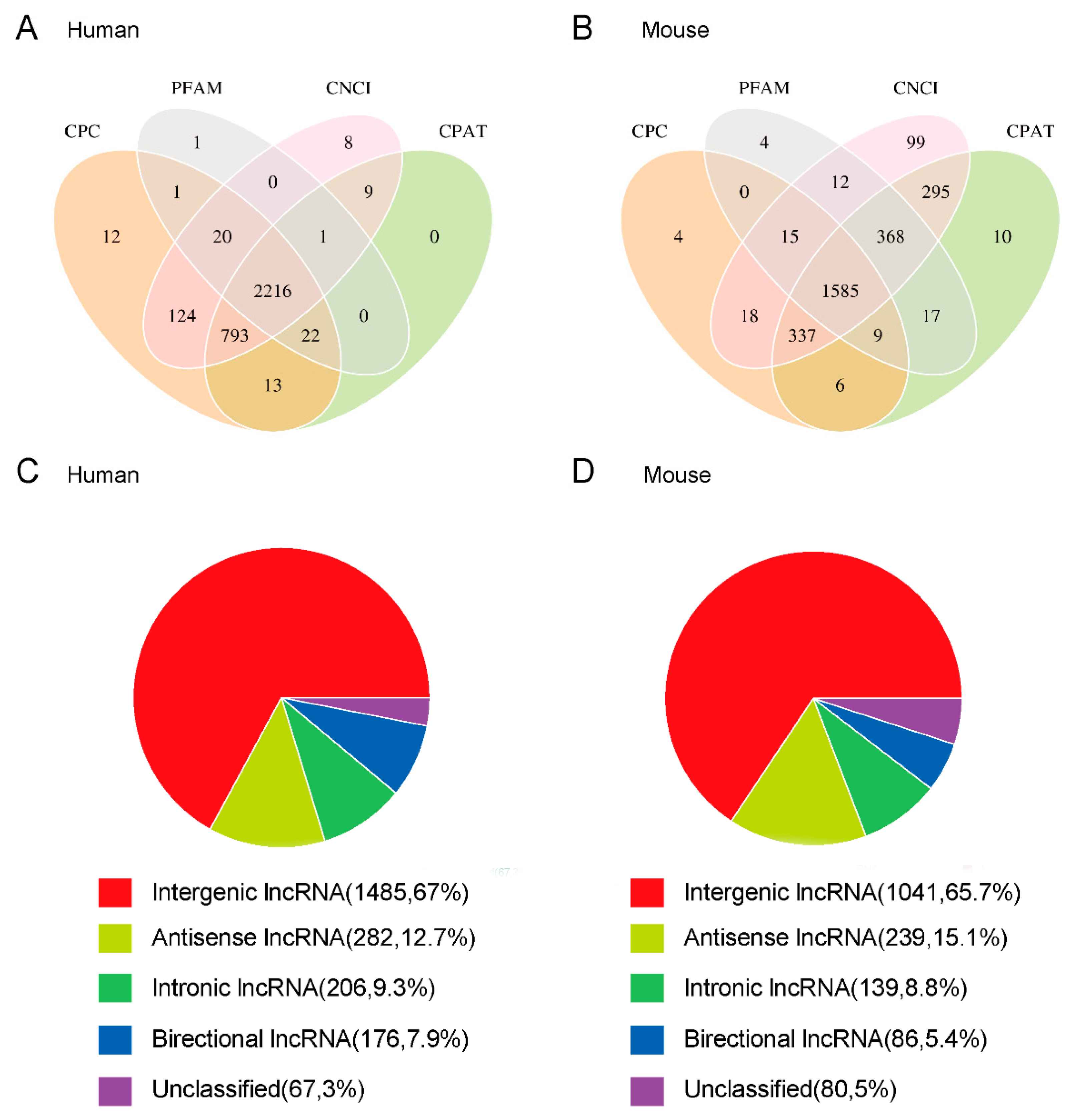
3.2. Identification and Classification of Newly Predicted lncRNAs
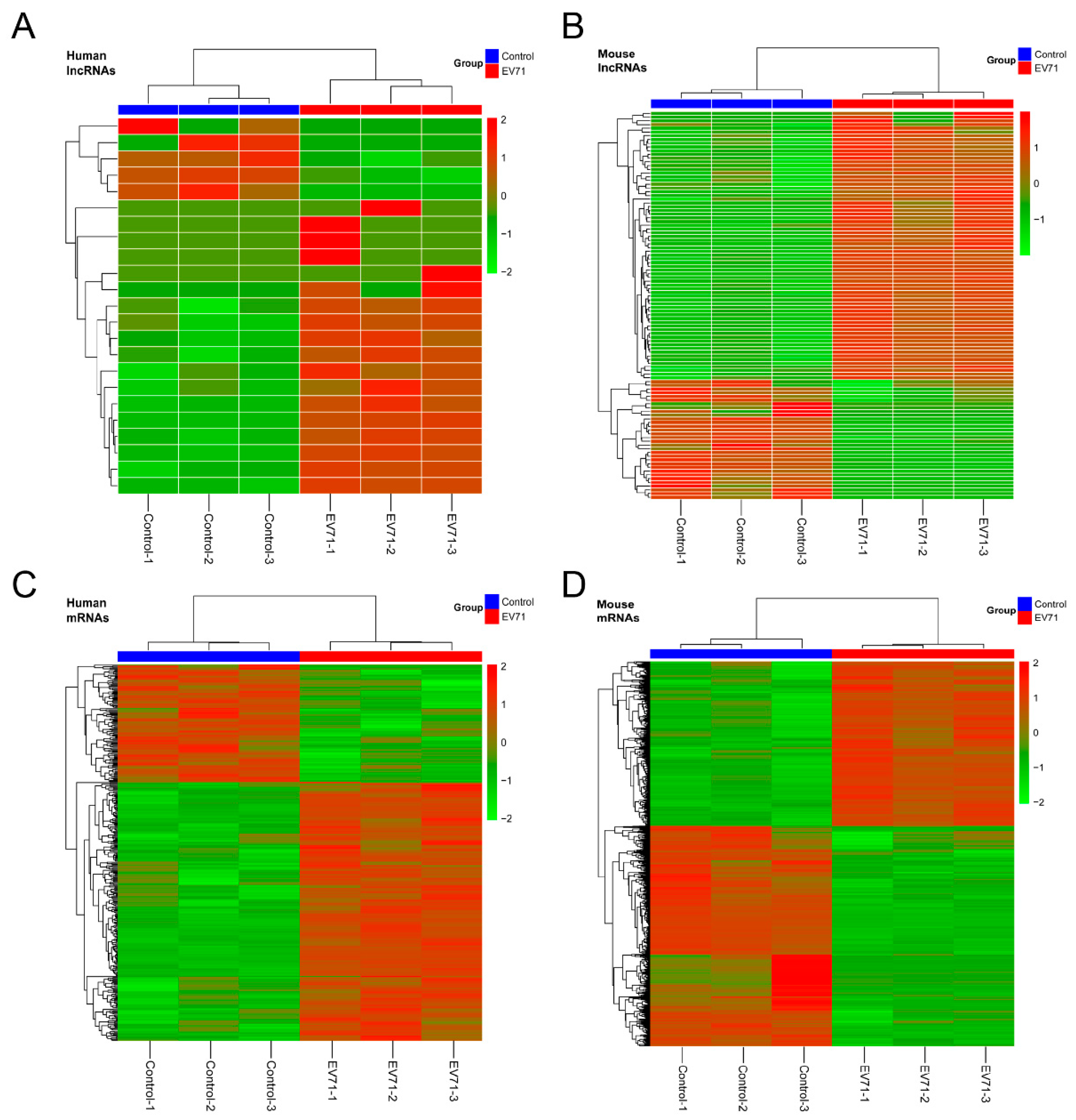
3.3. Profiles of Differentially-Expressed lncRNAs and mRNAs
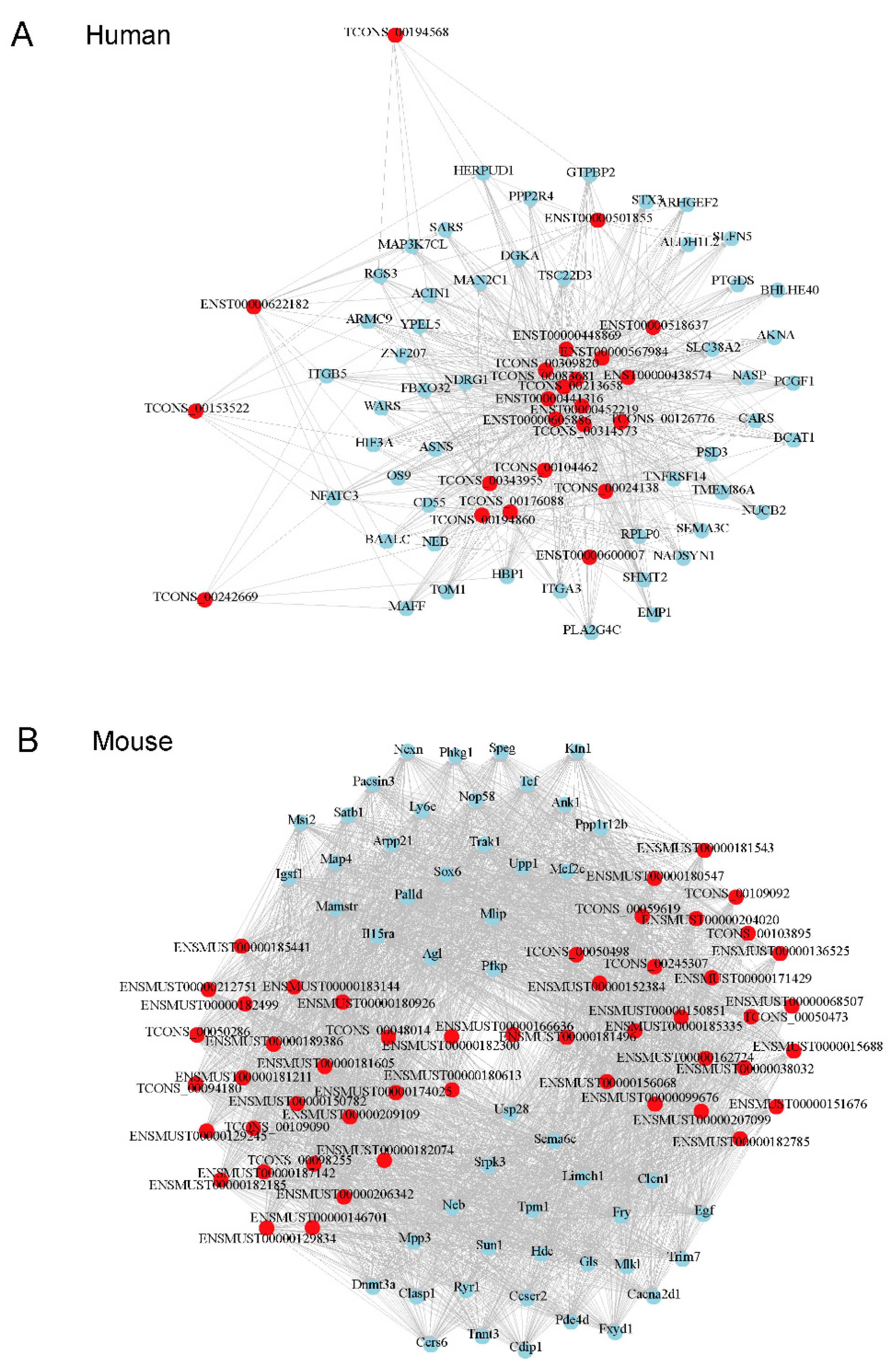
3.4. Prediction of Target Protein-Coding RNA
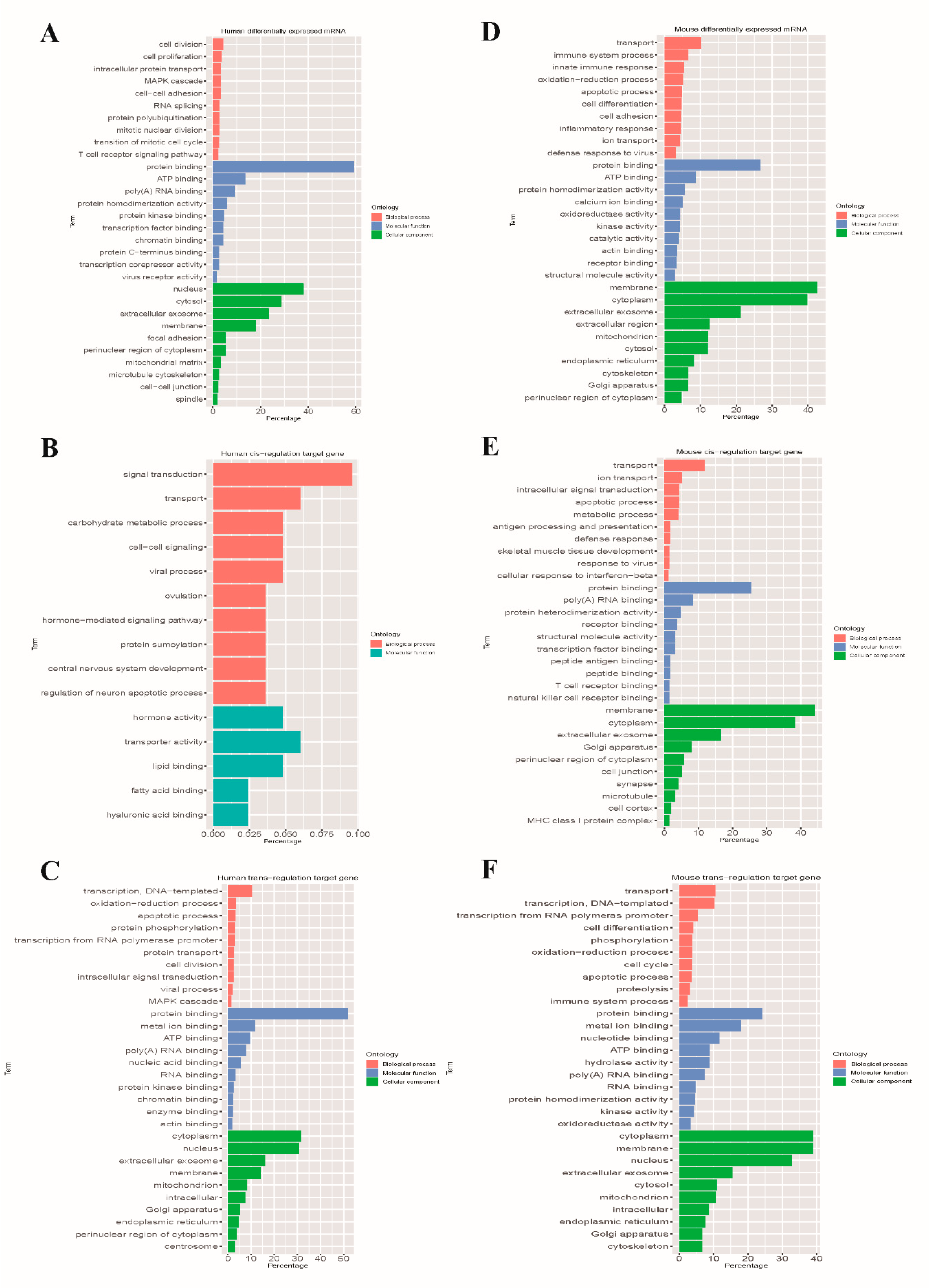
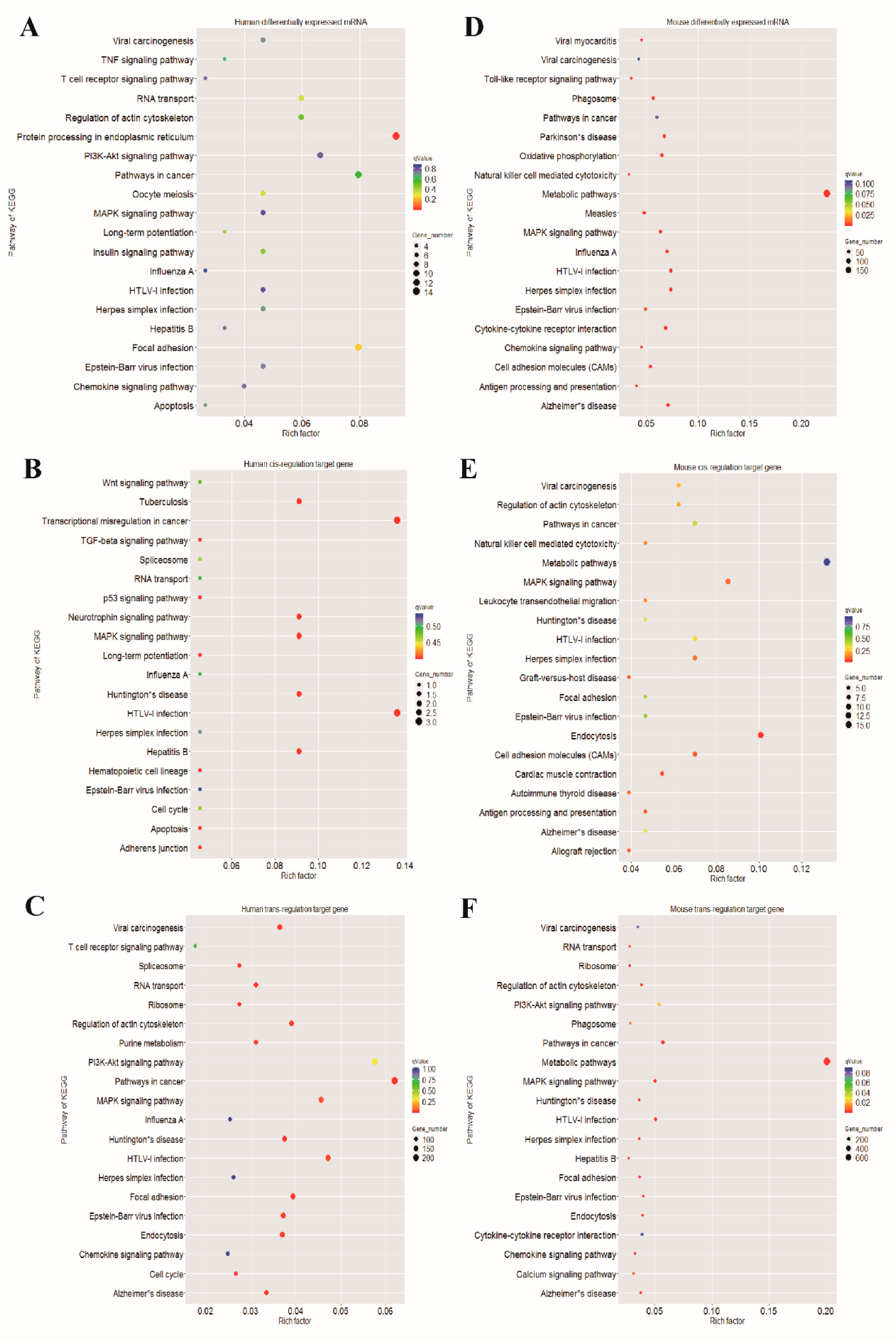
3.5. GO Annotation and KEGG Pathway Analysis
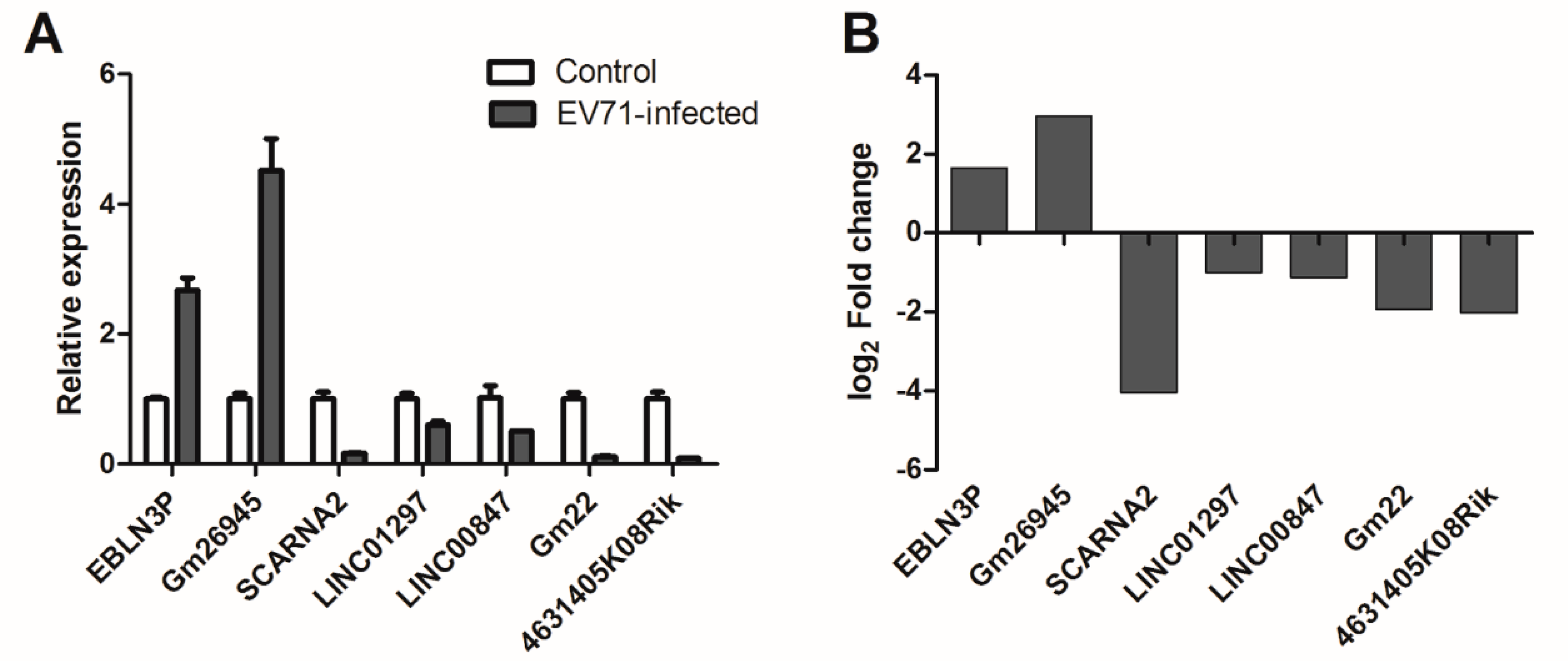
3.6. qPCR Validation
4. Discussion
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Lum, L.C.; Wong, K.T.; Lam, S.K.; Chua, K.B.; Goh, A.Y.; Lim, W.L.; Ong, B.B.; Paul, G.; AbuBakar, S.; Lambert, M. Fatal enterovirus 71 encephalomyelitis. J. Pediatr. 1998, 133, 795–798. [Google Scholar] [CrossRef]
- Stock, I. Hand, foot and mouth disease—More than a harmless “childhood disease”. Med. Monatsschr. Pharm. 2014, 37, 4–10, quiz 11–12. [Google Scholar] [PubMed]
- Solomon, T.; Lewthwaite, P.; Perera, D.; Cardosa, M.J.; McMinn, P.; Ooi, M.H. Virology, epidemiology, pathogenesis, and control of enterovirus 71. Lancet Infect. Dis. 2010, 10, 778–790. [Google Scholar] [CrossRef] [Green Version]
- Pathinayake, P.S.; Hsu, A.C.; Wark, P.A. Innate Immunity and Immune Evasion by Enterovirus 71. Viruses 2015, 7, 6613–6630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hung, T.; Chang, H.Y. Long noncoding RNA in genome regulation: Prospects and mechanisms. RNA Biol. 2010, 7, 582–585. [Google Scholar] [CrossRef] [PubMed]
- Mercer, T.R.; Dinger, M.E.; Mattick, J.S. Long non-coding RNAs: Insights into functions. Nat. Rev. Genet. 2009, 10, 155–159. [Google Scholar] [CrossRef] [PubMed]
- Emmrich, S.; Streltsov, A.; Schmidt, F.; Thangapandi, V.R.; Reinhardt, D.; Klusmann, J. LincRNAs MONC and MIR100HG act as oncogenes in acute megakaryoblastic leukemia. Mol. Cancer 2014, 13, 171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponting, C.P.; Oliver, P.L.; Reik, W. Evolution and functions of long noncoding RNAs. Cell 2009, 136, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.T. Epigenetic regulation by long noncoding RNAs. Science 2012, 338, 1435–1439. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Bai, J.; Zhang, X.; Lv, Y.; Gong, Y.; Liu, L.; Zhao, H.; Yu, F.; Ping, Y.; Zhang, G.; et al. A comprehensive overview of lncRNA annotation resources. Brief. Bioinform. 2017, 18, 236–249. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Kuang, H.; Xue, J.; Liao, L.; Yin, F.; Zhou, X. LncRNA AB073614 regulates proliferation and metastasis of colorectal cancer cells via the PI3K/AKT signaling pathway. Biomed. Pharmacother. 2017, 93, 1230–1237. [Google Scholar] [CrossRef] [PubMed]
- Montes, M.; Lund, A.H. Emerging roles of lncRNAs in senescence. FEBS J. 2016, 283, 2414–2426. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zacharopoulou, E.; Gazouli, M.; Tzouvala, M.; Vezakis, A.; Karamanolis, G. The contribution of long non-coding RNAs in Inflammatory Bowel Diseases. Digest. Liver Dis. 2017, 49, 1067–1072. [Google Scholar] [CrossRef] [PubMed]
- Wen, K.; Yang, L.; Xiong, T.; Di, C.; Ma, D.; Wu, M.; Xue, Z.; Zhang, X.; Long, L.; Zhang, W.; et al. Critical roles of long noncoding RNAs in Drosophila spermatogenesis. Genome Res. 2016, 26, 1233–1244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, X.; Gralinski, L.; Armour, C.D.; Ferris, M.T.; Thomas, M.J.; Proll, S.; Bradel-Tretheway, B.G.; Korth, M.J.; Castle, J.C.; Biery, M.C.; et al. Unique signatures of long noncoding RNA expression in response to virus infection and altered innate immune signaling. MBio 2010, 1, e00206-10. [Google Scholar] [CrossRef] [PubMed]
- Dang, D.; Zhang, C.; Zhang, R.; Wu, W.; Chen, S.; Ren, J.; Zhang, P.; Zhou, G.; Feng, D.; Sun, T.; et al. Involvement of inducible nitric oxide synthase and mitochondrial dysfunction in the pathogenesis of enterovirus 71 infection. Oncotarget 2017, 8, 81014–81026. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, C.; Zhang, R.; Ren, J.; Chen, S.; Sui, M.; Zhou, G.; Dang, D.; Zhu, J.; Feng, H.; et al. Pulmonary edema following central nervous system lesions induced by a non- mouse-adapted EV71 strain in neonatal BALB/c mice. Virol. J. 2017, 14, 243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhan, S.; Dong, Y.; Zhao, W.; Guo, J.; Zhong, T.; Wang, L.; Li, L.; Zhang, H. Genome-wide identification and characterization of long non-coding RNAs in developmental skeletal muscle of fetal goat. BMC Genom. 2016, 17, 666. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Bai, J.; Wu, A.; Wang, Y.; Zhang, J.; Wang, Z.; Li, Y.; Xu, J.; Li, X. Co-LncRNA: Investigating the lncRNA combinatorial effects in GO annotations and KEGG pathways based on human RNA-Seq data. Database 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Weng, K.F.; Chen, L.L.; Huang, P.N.; Shih, S.R. Neural pathogenesis of enterovirus 71 infection. Microbes Infect. 2010, 12, 505–510. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, R.; Wu, W.; Duan, G. Antiviral and Inflammatory Cellular Signaling Associated with Enterovirus 71 Infection. Viruses 2018, 10, 155. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.S.; Yao, Y.C.; Lin, S.C.; Lee, Y.P.; Wang, Y.F.; Wang, J.R.; Liu, C.C.; Lei, H.Y.; Yu, C.K. Retrograde axonal transport: A major transmission route of enterovirus 71 in mice. J. Virol. 2007, 81, 8996–9003. [Google Scholar] [CrossRef] [PubMed]
- Zou, R.; Zhang, G.; Li, S.; Wang, W.; Yuan, J.; Li, J.; Wang, Y.; Lin, Y.; Deng, Y.; Zhou, B.; et al. A functional polymorphism in IFNAR1 gene is associated with susceptibility and severity of HFMD with EV71 infection. Sci. Rep. 2015, 5, 18541. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.; Cheng, X.; Yang, X.; Zhao, R.; Wang, P.; Han, Y.; Luo, Z.; Cao, Y.; Zhu, C.; Xiong, Y.; et al. Early growth response-1 facilitates enterovirus 71 replication by direct binding to the viral genome RNA. Int. J. Biochem. Cell Biol. 2015, 62, 36–46. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.P.; Dai, H.J.; Yang, Y.H.; Zhuang, Y.; Sheng, R. MicroRNA-23b inhibits enterovirus 71 replication through downregulation of EV71 VPl protein. Intervirology 2013, 56, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Yin, Z.; Guan, D.; Fan, Q.; Su, J.; Zheng, W.; Ma, W.; Ke, C. lncRNA expression signatures in response to enterovirus 71 infection. Biochem. Biophys. Res. Commun. 2013, 430, 629–633. [Google Scholar] [CrossRef] [PubMed]
- Meng, J.; Yao, Z.; He, Y.; Zhang, R.; Yang, H.; Yao, X.; Chen, L.; Zhang, H.; Cheng, J. Long non-coding RNA expression profiles in different severity EV71-infected hand foot and mouth disease patients. Biochem. Biophys. Res. Commun. 2017, 493, 1594–1600. [Google Scholar] [CrossRef] [PubMed]
- White, R.R.; Milholland, B.; MacRae, S.L.; Lin, M.; Zheng, D.; Vijg, J. Comprehensive transcriptional landscape of aging mouse liver. BMC Genom. 2015, 16, 899. [Google Scholar] [CrossRef] [PubMed]
- Sommer, C.A.; Christodoulou, C.; Gianotti-Sommer, A.; Shen, S.S.; Sailaja, B.S.; Hezroni, H.; Spira, A.; Meshorer, E.; Kotton, D.N.; Mostoslavsky, G. Residual expression of reprogramming factors affects the transcriptional program and epigenetic signatures of induced pluripotent stem cells. PLoS ONE 2012, 7, e51711. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Duan, X.; Holmes, J.A.; Li, W.; Lee, S.H.; Tu, Z.; Zhu, C.; Salloum, S.; Lidofsky, A.; Schaefer, E.A.; et al. A novel lncRNA regulates HCV infection through IFI6. Hepatology 2018. [Google Scholar] [CrossRef] [PubMed]
- Vikram, R.; Ramachandran, R.; Abdul, K.S. Functional significance of long non-coding RNAs in breast cancer. Breast Cancer 2014, 21, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Crea, F.; Watahiki, A.; Quagliata, L.; Xue, H.; Pikor, L.; Parolia, A.; Wang, Y.; Lin, D.; Lam, W.L.; Farrar, W.L.; et al. Identification of a long non-coding RNA as a novel biomarker and potential therapeutic target for metastatic prostate cancer. Oncotarget 2014, 5, 764–774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, M.; Zhao, H.; Wang, Z.; Cheng, L.; Yang, L.; Shi, H.; Yang, H.; Sun, J. Identification and validation of potential prognostic lncRNA biomarkers for predicting survival in patients with multiple myeloma. J. Exp. Clin. Cancer Res. 2015, 34, 102. [Google Scholar] [CrossRef] [PubMed]
- He, Q.Q.; Ren, S.; Xia, Z.C.; Cheng, Z.K.; Peng, N.F.; Zhu, Y. Fibronectin Facilitates Enterovirus 71 Infection by Mediating Viral Entry. J. Virol. 2018, 92. [Google Scholar] [CrossRef] [PubMed]
- Leong, P.W.; Liew, K.; Lim, W.; Chow, V.T. Differential display RT-PCR analysis of enterovirus-71-infected rhabdomyosarcoma cells reveals mRNA expression responses of multiple human genes with known and novel functions. Virology 2002, 295, 147–159. [Google Scholar] [CrossRef] [PubMed]
- Lu, C.; Liu, X.; Ding, X.; Chen, X.; Fan, H.; Liu, Y.; Xie, N.; Tan, Y.; Ko, J.; Zhang, W.; et al. A Metabolomics Profiling Study in Hand-Foot-and-Mouth Disease and Modulated Pathways of Clinical Intervention Using Liquid Chromatography/Quadrupole Time-of-Flight Mass Spectrometry. Evid. Based Complement. Altern. Med. 2013, 2013, 647452. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Zhang, C.; Wang, H.; Zhou, G.; Wang, X.; Zhang, R.; Chen, S.; Ren, J.; Chen, L.; Dang, D.; et al. Mast cells contribute to Enterovirus 71 infection-induced pulmonary edema in neonatal mice. Lab. Investig. 2018, 98, 1039–1051. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Peng, X.; Xie, T.; Lu, X.; Liu, F.; Wu, H.; Yang, Z.; Wang, J.; Cheng, L.; Wu, N. Detection of the long noncoding RNAs nuclear-enriched autosomal transcript 1 (NEAT1) and metastasis associated lung adenocarcinoma transcript 1 in the peripheral blood of HIV-1-infected patients. HIV Med. 2016, 17, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Morchikh, M.; Cribier, A.; Raffel, R.; Amraoui, S.; Cau, J.; Severac, D.; Dubois, E.; Schwartz, O.; Bennasser, Y.; Benkirane, M. HEXIM1 and NEAT1 Long Non-coding RNA Form a Multi-subunit Complex that Regulates DNA-Mediated Innate Immune Response. Mol. Cell 2017, 67, 387–399. [Google Scholar] [CrossRef] [PubMed]
- Paraskevopoulou, M.D.; Hatzigeorgiou, A.G. Analyzing MiRNA-LncRNA Interactions. Methods Mol. Biol. 2016, 1402, 271–286. [Google Scholar] [PubMed]
- Keniry, A.; Oxley, D.; Monnier, P.; Kyba, M.; Dandolo, L.; Smits, G.; Reik, W. The H19 lincRNA is a developmental reservoir of miR-675 that suppresses growth and Igf1r. Nat. Cell Biol. 2012, 14, 659–665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anderson, D.M.; Anderson, K.M.; Chang, C.L.; Makarewich, C.A.; Nelson, B.R.; McAnally, J.R.; Kasaragod, P.; Shelton, J.M.; Liou, J.; Bassel-Duby, R.; et al. A micropeptide encoded by a putative long noncoding RNA regulates muscle performance. Cell 2015, 160, 595–606. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, A.; Pasut, A.; Matsumoto, M.; Yamashita, R.; Fung, J.; Monteleone, E.; Saghatelian, A.; Nakayama, K.I.; Clohessy, J.G.; Pandolfi, P.P. mTORC1 and muscle regeneration are regulated by the LINC00961-encoded SPAR polypeptide. Nature 2017, 541, 228–232. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| lncRNA Symbol | Chromosome Location | Primer Sequences (5′-3′) | Products (bp) | |
|---|---|---|---|---|
| EBLN3P | 9 | Forward | GGACGCGGCCTCCTTACCTC | 189 |
| Reverse | GGCACGCGCTAGTTCCACAG | 189 | ||
| Gm26945 | 12 | Forward | GGTGGCTCATGTGGATCACTGATG | 87 |
| Reverse | ACGATGGACCTCCTGAGTGACAG | 87 | ||
| SCARNA2 | 1 | Forward | CTGTGGCGTCGCGTGTGAG | 97 |
| Reverse | CGCACGCACTCGCCTAACAC | 97 | ||
| LINC01297 | 22 | Forward | ACGTCTCCATCCAGGACTGAGG | 141 |
| Reverse | AAGACTGCCATCCAAGGAAGCG | 141 | ||
| LINC00847 | 5 | Forward | GGACTTGCCAGCCTTCAGAACTC | 181 |
| Reverse | GGTGATTGGATAGTGCCTGCCTAC | 181 | ||
| Gm22 | 8 | Forward | GCCACAGCCATCAGCACAGC | 127 |
| Reverse | CGCAACACCACTGACATGGAGAG | 127 | ||
| 4631405K08Rik | 1 | Forward | AGAGTCACTGTCCTCCGCTTATCC | 199 |
| Reverse | GCTGGCTCTCCTCTTCAATGAGTG | 199 | ||
| β-actin | Forward | AGCGAGCATCCCCCAAAGTT | 285 | |
| (Homo sapiens) | Reverse | GGGCACGAAGGCTCATCATT | 285 | |
| β-actin | Forward | GTGACGTTGACATCCGTAAAG | 287 | |
| (Mus musculus) | Reverse | GTAACAGTCCGCCTAGAAGCAC | 287 | |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhang, C.; Qin, L.; Li, D.; Zhou, G.; Dang, D.; Chen, S.; Sun, T.; Zhang, R.; Wu, W.; et al. Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq. Viruses 2018, 10, 556. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100556
Li Y, Zhang C, Qin L, Li D, Zhou G, Dang D, Chen S, Sun T, Zhang R, Wu W, et al. Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq. Viruses. 2018; 10(10):556. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100556
Chicago/Turabian StyleLi, Ying, Chao Zhang, Luwei Qin, Dong Li, Guangyuan Zhou, Dejian Dang, Shuaiyin Chen, Tiantian Sun, Rongguang Zhang, Weidong Wu, and et al. 2018. "Characterization of Critical Functions of Long Non-Coding RNAs and mRNAs in Rhabdomyosarcoma Cells and Mouse Skeletal Muscle Infected by Enterovirus 71 Using RNA-Seq" Viruses 10, no. 10: 556. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100556