Bowman‒Birk Inhibitor Suppresses Herpes Simplex Virus Type 2 Infection of Human Cervical Epithelial Cells

 and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Virus
2.2. BBI
2.3. Reagents
2.4. Cell Viability Assay
2.5. In Vitro Antiviral Assay
2.6. RNA Extraction and Real-Time PCR
2.7. Western Blot Analysis
2.8. Statistical Analysis
3. Results
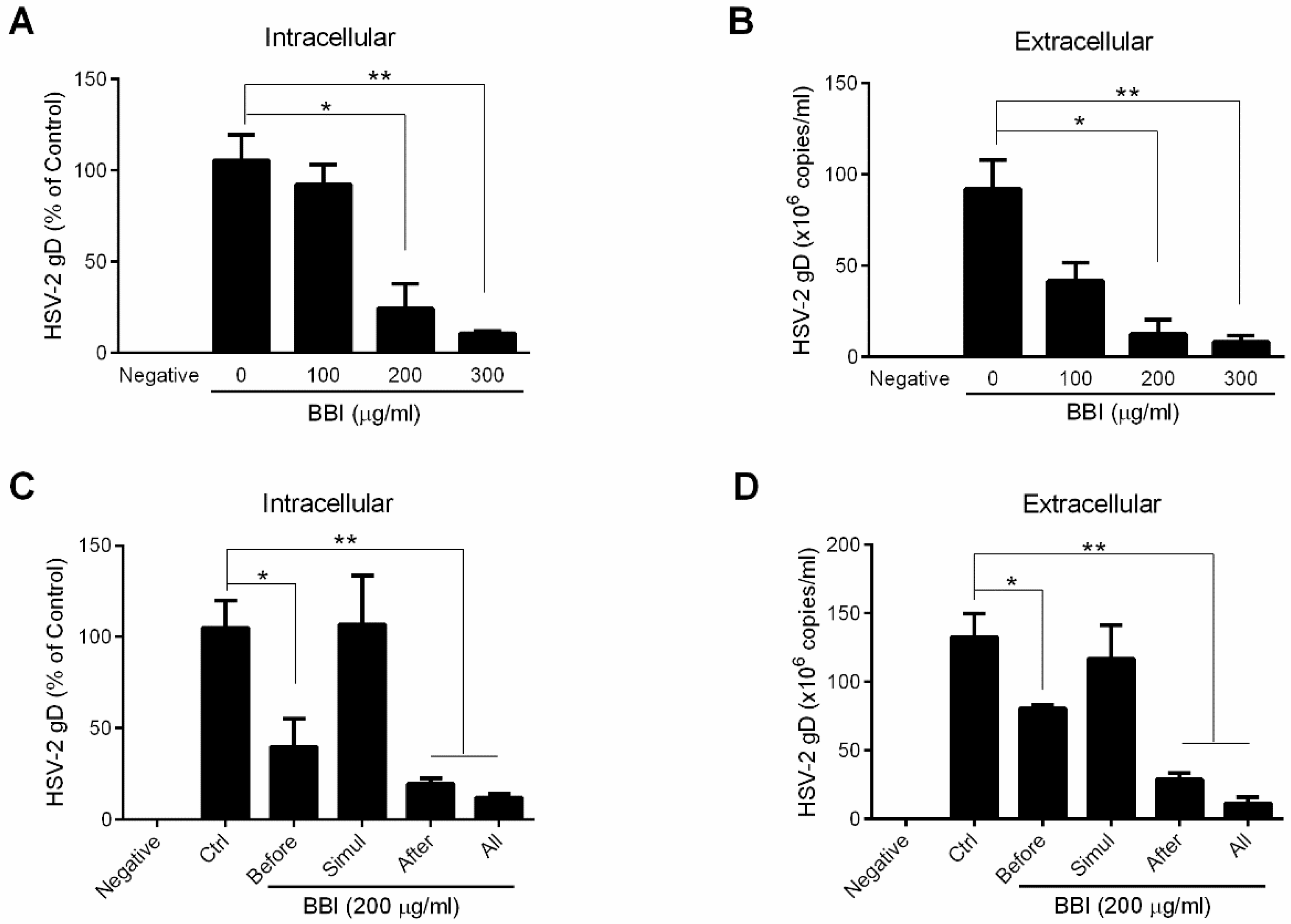
3.1. BBI Inhibits HSV-2 Infection of End1/E6E7 Cells
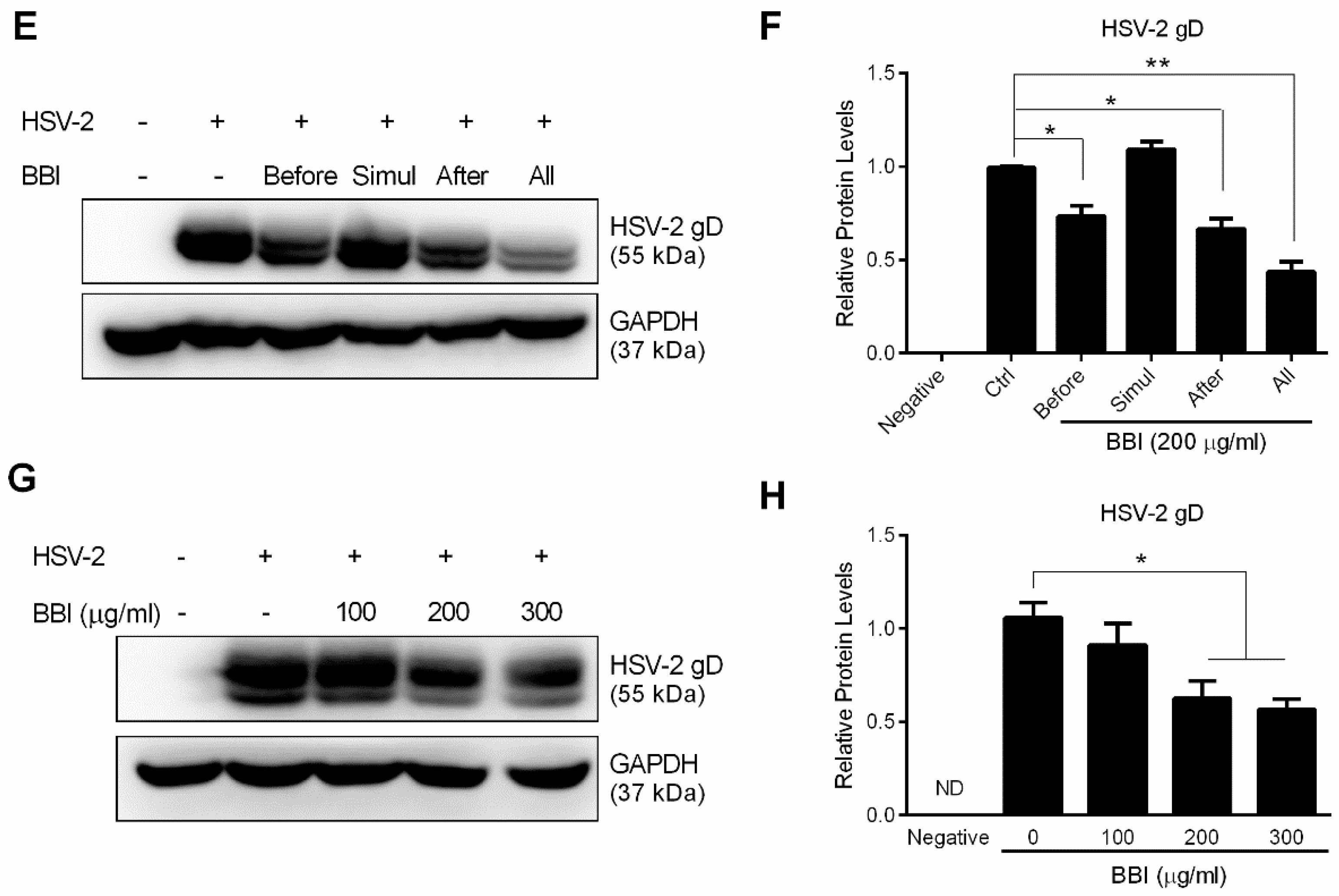
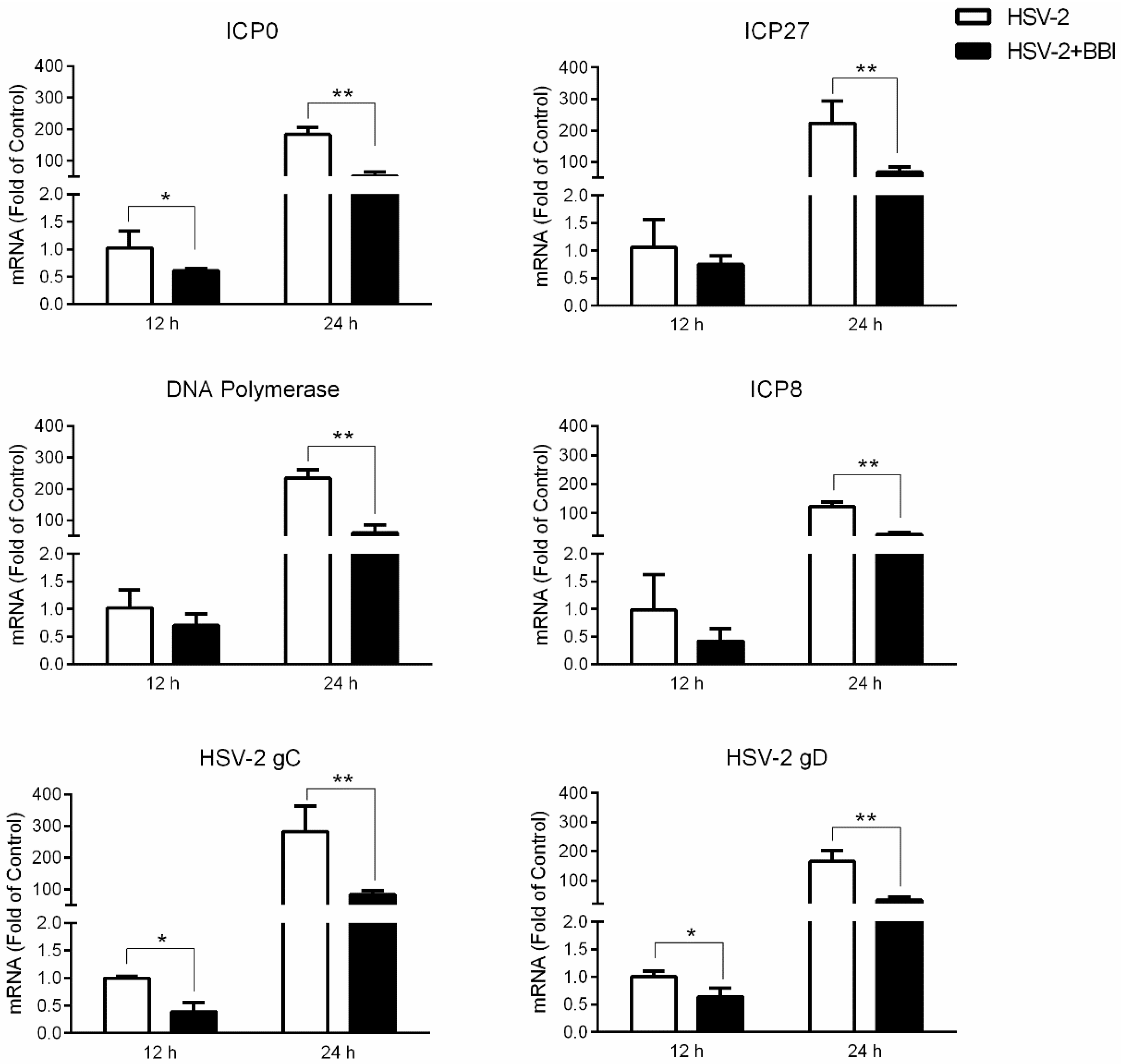
3.2. BBI Suppresses HSV-2 Gene Expression
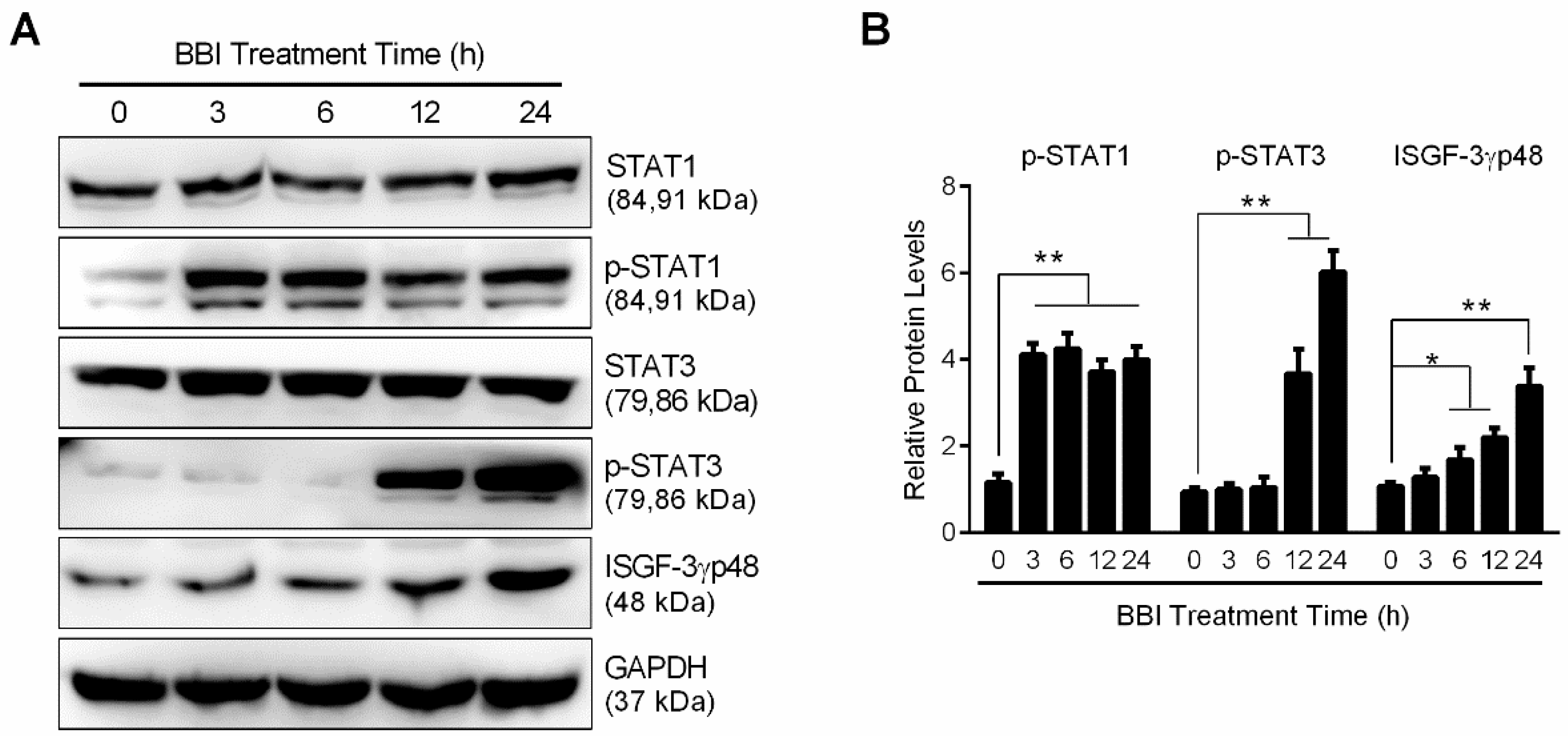
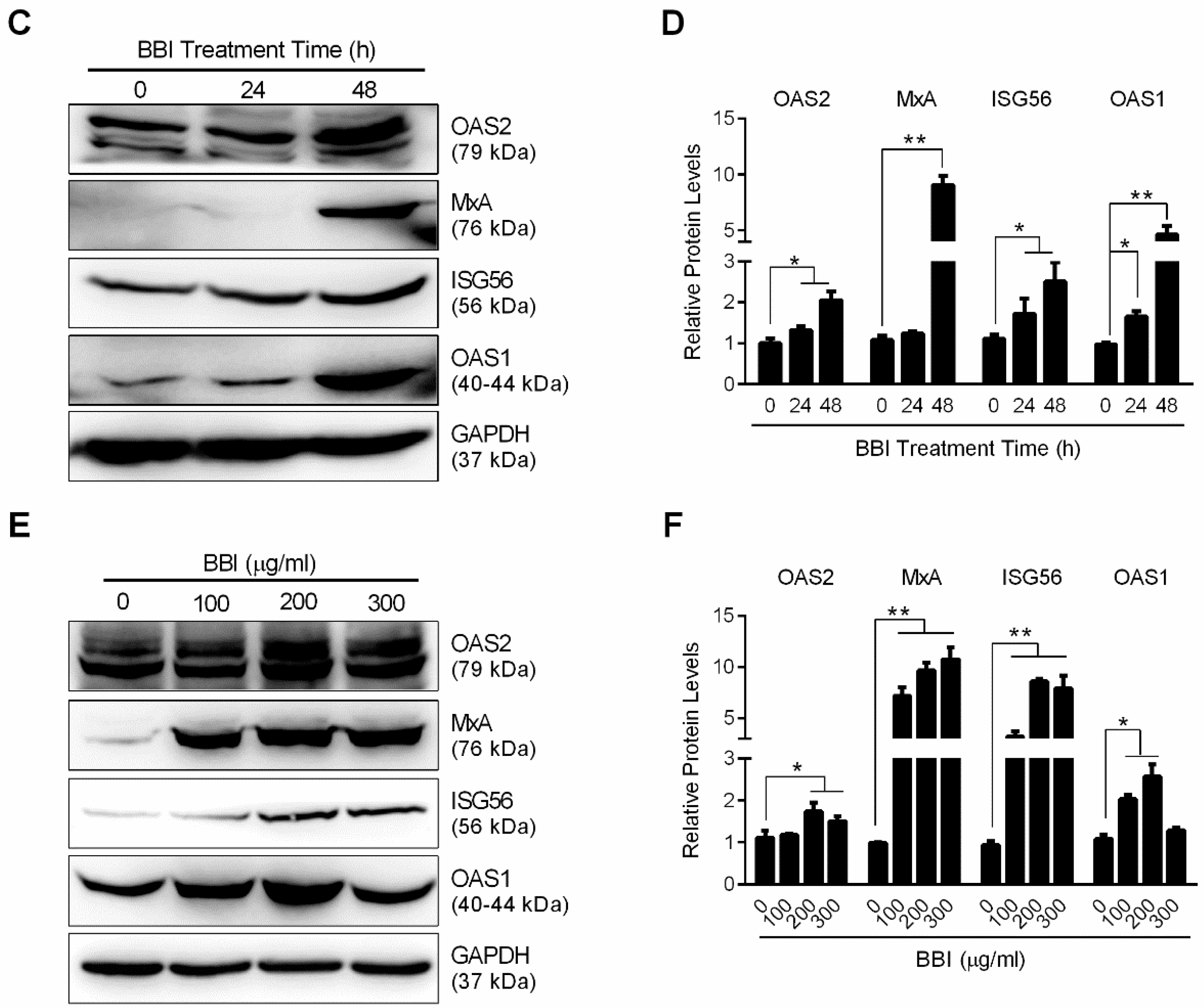
3.3. BBI Activates the JAK/STAT Signaling Pathway
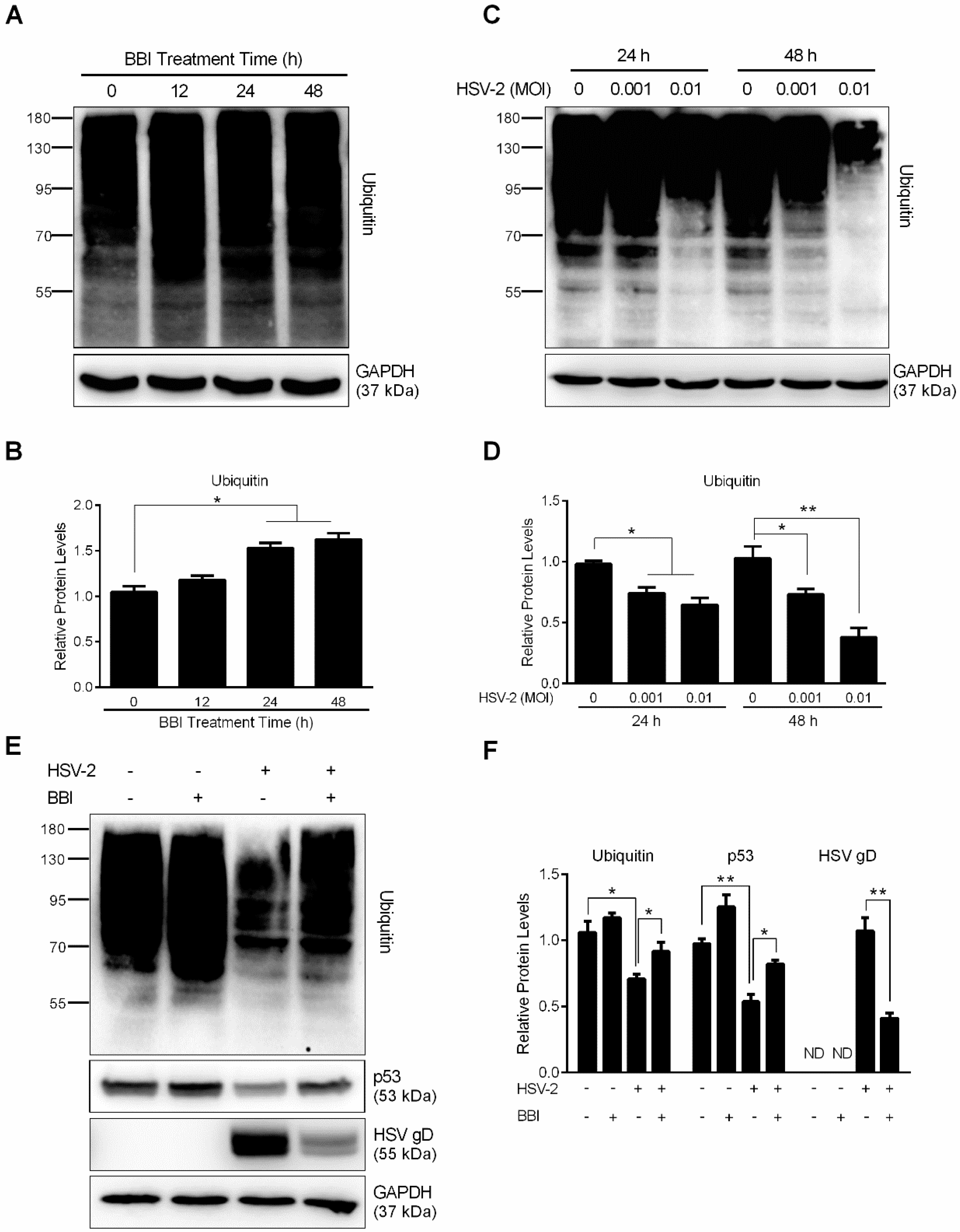
3.4. BBI Inhibits the Cellular UPS
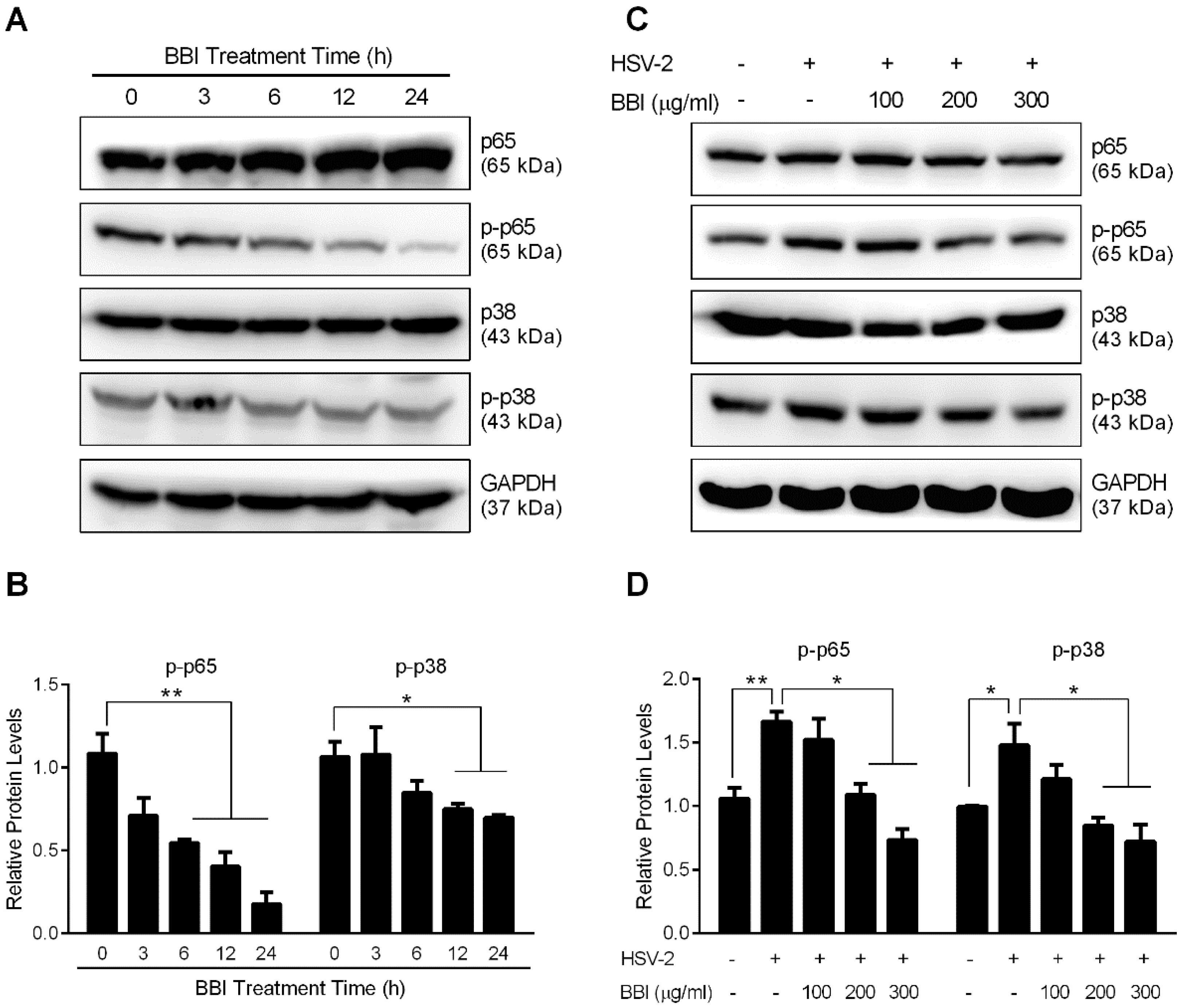
3.5. BBI Inhibits HSV-2-Induced Activation of NF-κB and MAPK
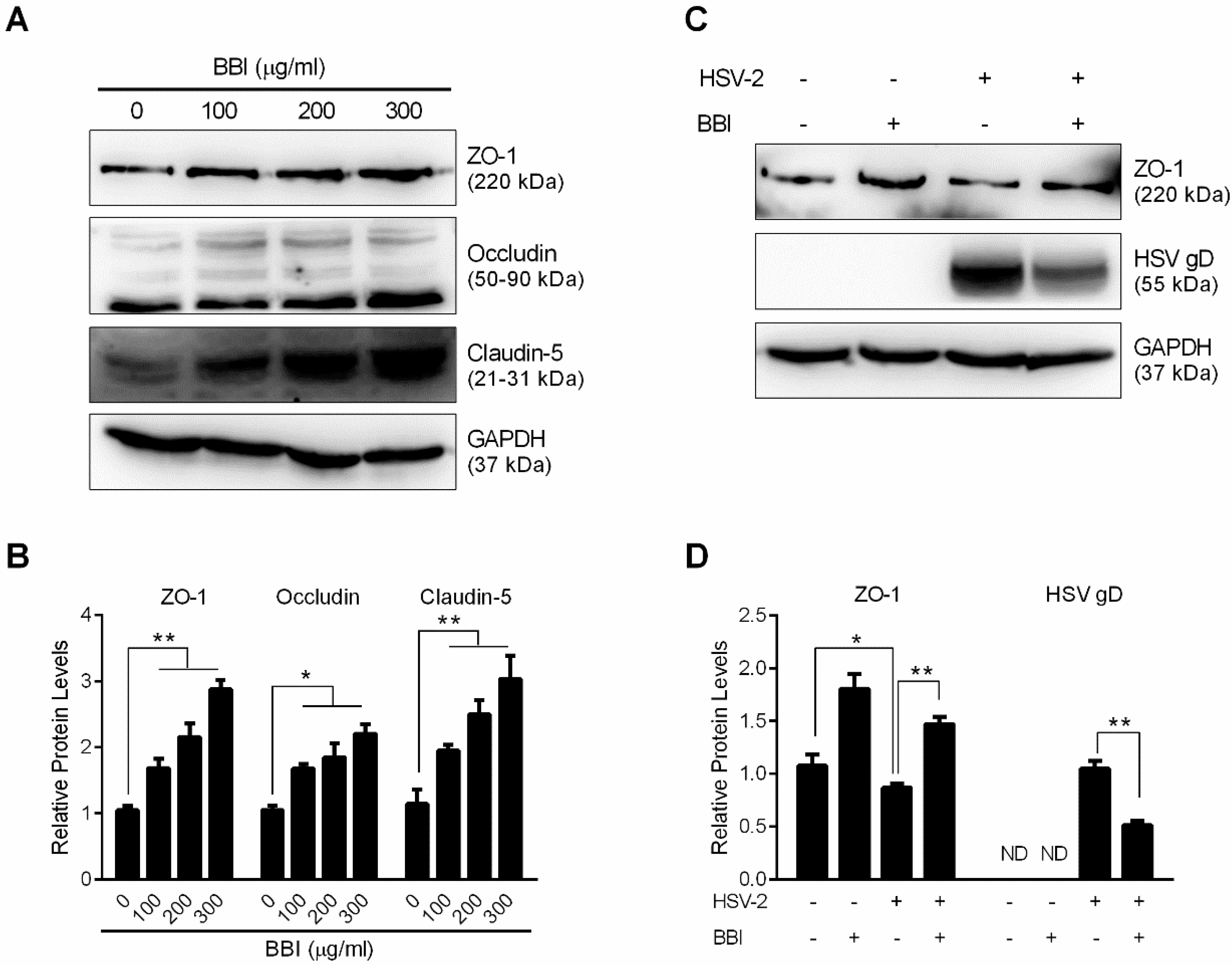
3.6. BBI Suppresses HSV-2-Induced Downregulation of Tight Junction Proteins
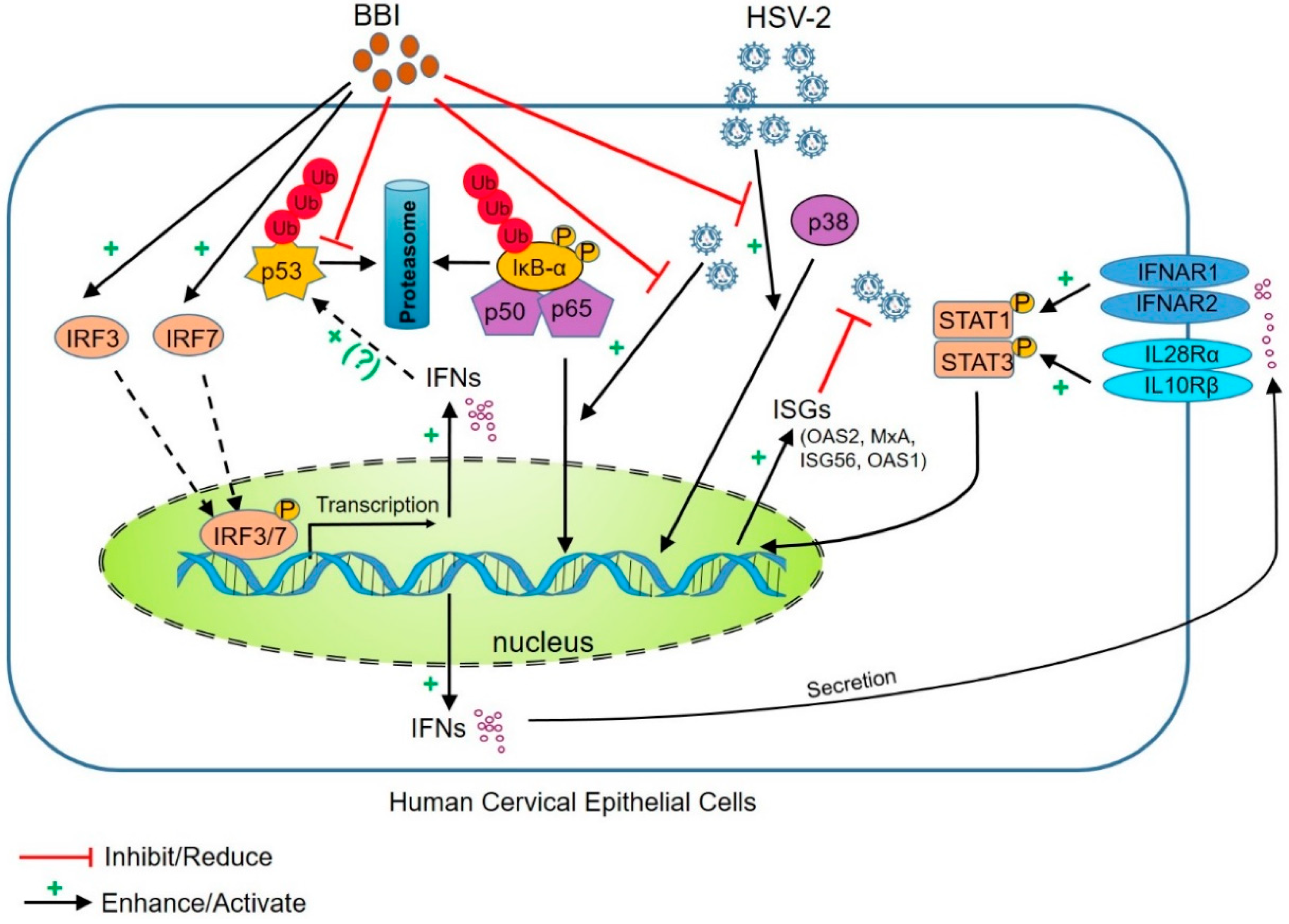
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Pinninti, S.G.; Kimberlin, D.W. Maternal and neonatal herpes simplex virus infections. Am. J. Perinatol. 2013, 30, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Looker, K.J.; Magaret, A.S.; May, M.T.; Turner, K.M.E.; Vickerman, P.; Newman, L.M.; Gottlieb, S.L. First estimates of the global and regional incidence of neonatal herpes infection. Lancet Glob. Health 2017, 5, e300–e309. [Google Scholar] [CrossRef] [Green Version]
- Fearon, E.; Wiggins, R.D.; Pettifor, A.E.; MacPhail, C.; Kahn, K.; Selin, A.; Gomez-Olive, F.X.; Delany-Moretlwe, S. Associations between friendship characteristics and HIV and HSV-2 status amongst young South African women in HPTN-068. J. Int. AIDS Soc. 2017, 20, e25029. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rollenhagen, C.; Lathrop, M.J.; Macura, S.L.; Doncel, G.F.; Asin, S.N. Herpes simplex virus type-2 stimulates HIV-1 replication in cervical tissues: Implications for HIV-1 transmission and efficacy of anti-HIV-1 microbicides. Mucosal Immunol. 2014, 7, 1165–1174. [Google Scholar] [CrossRef] [PubMed]
- Freeman, E.E.; Weiss, H.A.; Glynn, J.R.; Cross, P.L.; Whitworth, J.A.; Hayes, R.J. Herpes simplex virus 2 infection increases HIV acquisition in men and women: Systematic review and meta-analysis of longitudinal studies. AIDS 2006, 20, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Sheth, P.M.; Sunderji, S.; Shin, L.Y.; Rebbapragada, A.; Huibner, S.; Kimani, J.; Macdonald, K.S.; Ngugi, E.; Bwayo, J.J.; Moses, S.; et al. Coinfection with herpes simplex virus type 2 is associated with reduced HIV-specific T cell responses and systemic immune activation. J. Infect. Dis. 2008, 197, 1394–1401. [Google Scholar] [CrossRef] [PubMed]
- Drannik, A.G.; Nag, K.; Sallenave, J.M.; Rosenthal, K.L. Antiviral activity of trappin-2 and elafin in vitro and in vivo against genital herpes. J. Virol. 2013, 87, 7526–7538. [Google Scholar] [CrossRef] [PubMed]
- Cannella, F.; Scagnolari, C.; Selvaggi, C.; Stentella, P.; Recine, N.; Antonelli, G.; Pierangeli, A. Interferon lambda 1 expression in cervical cells differs between low-risk and high-risk human papillomavirus-positive women. Med. Microbiol. Immunol. 2014, 203, 177–184. [Google Scholar] [CrossRef] [PubMed]
- Ank, N.; Paludan, S.R. Type III IFNs: New layers of complexity in innate antiviral immunity. Biofactors 2009, 35, 82–87. [Google Scholar] [CrossRef] [PubMed]
- Fichorova, R.N.; Cronin, A.O.; Lien, E.; Anderson, D.J.; Ingalls, R.R. Response to Neisseria gonorrhoeae by cervicovaginal epithelial cells occurs in the absence of toll-like receptor 4-mediated signaling. J. Immunol. 2002, 168, 2424–2432. [Google Scholar] [CrossRef] [PubMed]
- Zhou, L.; Li, J.L.; Zhou, Y.; Liu, J.B.; Zhuang, K.; Gao, J.F.; Liu, S.; Sang, M.; Wu, J.G.; Ho, W.Z. Induction of interferon-lambda contributes to TLR3 and RIG-I activation-mediated inhibition of herpes simplex virus type 2 replication in human cervical epithelial cells. Mol. Hum. Reprod. 2015, 21, 917–929. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Q.; Guo, L.; Wang, X.; Liu, Y.; Liu, H.; Zhou, R.H.; Gu, J.; Liu, J.B.; Xu, P.; Zhou, L.; et al. Human Cervical Epithelial Cells Release Antiviral Factors and Inhibit HIV Replication in Macrophages. J. Innate Immun. 2018, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Klysik, K.; Pietraszek, A.; Karewicz, A.; Nowakowska, M. Acyclovir in the Treatment of Herpes Viruses—A Review. Curr. Med. Chem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.; Wald, A.; Krantz, E.; Selke, S.; Warren, T.; Vargas-Cortes, M.; Miller, G.; Corey, L. Valacyclovir and acyclovir for suppression of shedding of herpes simplex virus in the genital tract. J. Infect. Dis. 2004, 190, 1374–1381. [Google Scholar] [CrossRef] [PubMed]
- Karrasch, M.; Liermann, K.; Betz, B.B.; Wagner, S.; Scholl, S.; Dahms, C.; Sauerbrei, A.; Kunze, A. Rapid acquisition of acyclovir resistance in an immunodeficient patient with herpes simplex encephalitis. J. Neurol. Sci. 2018, 384, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Scieux, C.; Garrait, V.; Socie, G.; Rocha, V.; Molina, J.M.; Thouvenot, D.; Morfin, F.; Hocqueloux, L.; Garderet, L.; et al. Resistant herpes simplex virus type 1 infection: An emerging concern after allogeneic stem cell transplantation. Clin. Infect. Dis. 2000, 31, 927–935. [Google Scholar] [CrossRef] [PubMed]
- Piret, J.; Boivin, G. Antiviral drug resistance in herpesviruses other than cytomegalovirus. Rev. Med. Virol. 2014, 24, 186–218. [Google Scholar] [CrossRef] [PubMed]
- Smirnoff, P.; Ramachandran, J.; Birk, Y. Preparation of photoreactive derivatives of trypsin-chymotrypsin inhibitors from soybeans and chick peas by selective modification of lysine residues. Int. J. Pept. Protein Res. 1985, 26, 274–278. [Google Scholar] [CrossRef] [PubMed]
- Fereidunian, A.; Sadeghalvad, M.; Oscoie, M.O.; Mostafaie, A. Soybean Bowman-Birk protease inhibitor (BBI): Identification of the mechanisms of BBI suppressive effect on growth of two adenocarcinoma cell lines: AGS and HT29. Arch. Med. Res. 2014, 45, 455–461. [Google Scholar] [CrossRef] [PubMed]
- Witschi, H.; Kennedy, A.R. Modulation of lung tumor development in mice with the soybean-derived Bowman-Birk protease inhibitor. Carcinogenesis 1989, 10, 2275–2277. [Google Scholar] [CrossRef] [PubMed]
- Gran, B.; Tabibzadeh, N.; Martin, A.; Ventura, E.S.; Ware, J.H.; Zhang, G.X.; Parr, J.L.; Kennedy, A.R.; Rostami, A.M. The protease inhibitor, Bowman-Birk Inhibitor, suppresses experimental autoimmune encephalomyelitis: A potential oral therapy for multiple sclerosis. Mult. Scler. 2006, 12, 688–697. [Google Scholar] [CrossRef] [PubMed]
- Routray, D.S. Bowman Birk Inhibitors (BBI) in interception of inflammation and malignant transformation of OPMDs. Oral Oncol. 2018, 78, 220–221. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Ye, L.; Cook, D.R.; Wang, X.; Liu, J.; Kolson, D.L.; Persidsky, Y.; Ho, W.Z. Soybean-derived Bowman-Birk inhibitor inhibits neurotoxicity of LPS-activated macrophages. J. Neuroinflamm. 2011, 8, 15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Larionova, N.V.; Malykh, E.V.; Villemson, A.L.; Krasota, A.J.; Duchene, D.; Ollivon, M.; Gernet, M.V.; Belousova, R.V.; Shen, W.C.; Larionova, N.I. Effect of membranotropic and mucoadhesive formulations of protein proteinase inhibitors on bovine herpes virus-1 reproduction. Int. J. Pharm. 2003, 256, 191–198. [Google Scholar] [CrossRef]
- Ma, T.C.; Le, G.; Zhou, R.H.; Wang, X.; Liu, J.B.; Li, J.L.; Zhou, Y.; Hou, W.; Ho, W.Z. Soybean-derived Bowman-Birk inhibitor (BBI) blocks HIV entry into macrophages. Virology 2018, 513, 91–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, T.C.; Zhou, R.H.; Wang, X.; Li, J.L.; Sang, M.; Zhou, L.; Zhuang, K.; Hou, W.; Guo, D.Y.; Ho, W.Z. Soybean-derived Bowman-Birk Inhibitor (BBI) Inhibits HIV Replication in Macrophages. Sci. Rep. 2016, 6, 34752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fichorova, R.N.; Rheinwald, J.G.; Anderson, D.J. Generation of papillomavirus-immortalized cell lines from normal human ectocervical, endocervical, and vaginal epithelium that maintain expression of tissue-specific differentiation proteins. Biol. Reprod. 1997, 57, 847–855. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, M.; Chen, Y.; Cheng, L.; Chu, Y.; Song, H.Y.; Wu, Z.W. Pyrrolidine dithiocarbamate inhibits herpes simplex virus 1 and 2 replication, and its activity may be mediated through dysregulation of the ubiquitin-proteasome system. J. Virol. 2013, 87, 8675–8686. [Google Scholar] [CrossRef] [PubMed]
- Teresa Sciortino, M.; Medici, M.A.; Marino-Merlo, F.; Zaccaria, D.; Giuffre, M.; Venuti, A.; Grelli, S.; Mastino, A. Signaling pathway used by HSV-1 to induce NF-κB activation: Possible role of herpes virus entry receptor A. Ann. N. Y. Acad. Sci. 2007, 1096, 89–96. [Google Scholar] [CrossRef] [PubMed]
- Gillis, P.A.; Okagaki, L.H.; Rice, S.A. Herpes simplex virus type 1 ICP27 induces p38 mitogen-activated protein kinase signaling and apoptosis in HeLa cells. J. Virol. 2009, 83, 1767–1777. [Google Scholar] [CrossRef] [PubMed]
- Blaskewicz, C.D.; Pudney, J.; Anderson, D.J. Structure and function of intercellular junctions in human cervical and vaginal mucosal epithelia. Biol. Reprod. 2011, 85, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xu, X.Q.; Zhou, L.; Zhou, R.H.; Wang, X.; Li, J.L.; Liu, J.B.; Liu, H.; Zhang, B.; Ho, W.Z. Human Intestinal Epithelial Cells Release Antiviral Factors That Inhibit HIV Infection of Macrophages. Front. Immunol. 2018, 9, 247. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Wang, Y.; Wang, X.; Ye, L.; Zhou, Y.; Persidsky, Y.; Ho, W. Immune activation of human brain microvascular endothelial cells inhibits HIV replication in macrophages. Blood 2013, 121, 2934–2942. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, A.; Zhang, J.; Yu, F.S. Toll-like receptor 3 agonist poly(I:C)-induced antiviral response in human corneal epithelial cells. Immunology 2006, 117, 11–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, W.; Schaffer, P.A. Herpes simplex virus type 1 ICP0 regulates expression of immediate-early, early, and late genes in productively infected cells. J. Virol. 1992, 66, 2904–2915. [Google Scholar] [PubMed]
- Bryant, H.E.; Wadd, S.E.; Lamond, A.I.; Silverstein, S.J.; Clements, J.B. Herpes simplex virus IE63 (ICP27) protein interacts with spliceosome-associated protein 145 and inhibits splicing prior to the first catalytic step. J. Virol. 2001, 75, 4376–4385. [Google Scholar] [CrossRef] [PubMed]
- Gottlieb, J.; Marcy, A.I.; Coen, D.M.; Challberg, M.D. The herpes simplex virus type 1 UL42 gene product: A subunit of DNA polymerase that functions to increase processivity. J. Virol. 1990, 64, 5976–5987. [Google Scholar] [PubMed]
- Gao, M.; Knipe, D.M. Potential role for herpes simplex virus ICP8 DNA replication protein in stimulation of late gene expression. J. Virol. 1991, 65, 2666–2675. [Google Scholar] [PubMed]
- Spear, P.G. Herpes simplex virus: Receptors and ligands for cell entry. Cell. Microbiol. 2004, 6, 401–410. [Google Scholar] [CrossRef] [PubMed]
- Sainz, B., Jr.; Halford, W.P. Alpha/Beta interferon and gamma interferon synergize to inhibit the replication of herpes simplex virus type 1. J. Virol. 2002, 76, 11541–11550. [Google Scholar] [CrossRef] [PubMed]
- Honda, K.; Takaoka, A.; Taniguchi, T. Type I inteferon gene induction by the interferon regulatory factor family of transcription factors. Immunity 2006, 25, 349–360. [Google Scholar] [CrossRef] [PubMed]
- Lazear, H.M.; Nice, T.J.; Diamond, M.S. Interferon-lambda: Immune Functions at Barrier Surfaces and Beyond. Immunity 2015, 43, 15–28. [Google Scholar] [CrossRef] [PubMed]
- Kristiansen, H.; Scherer, C.A.; McVean, M.; Iadonato, S.P.; Vends, S.; Thavachelvam, K.; Steffensen, T.B.; Horan, K.A.; Kuri, T.; Weber, F.; et al. Extracellular 2′-5′ oligoadenylate synthetase stimulates RNase L-independent antiviral activity: A novel mechanism of virus-induced innate immunity. J. Virol. 2010, 84, 11898–11904. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Pan, Q.; Rong, L.; He, W.; Liu, S.L.; Liang, C. The IFITM proteins inhibit HIV-1 infection. J. Virol. 2011, 85, 2126–2137. [Google Scholar] [CrossRef] [PubMed]
- Yao, X.D.; Rosenthal, K.L. Herpes simplex virus type 2 virion host shutoff protein suppresses innate dsRNA antiviral pathways in human vaginal epithelial cells. J. Gen. Virol. 2011, 92 Pt 9, 1981–1993. [Google Scholar] [CrossRef]
- Christensen, M.H.; Jensen, S.B.; Miettinen, J.J.; Luecke, S.; Prabakaran, T.; Reinert, L.S.; Mettenleiter, T.; Chen, Z.J.; Knipe, D.M.; Sandri-Goldin, R.M.; et al. HSV-1 ICP27 targets the TBK1-activated STING signalsome to inhibit virus-induced type I IFN expression. EMBO J. 2016, 35, 1385–1399. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; You, H.; Su, C.; Li, Y.; Chen, S.; Zheng, C. Herpes Simplex Virus 1 Tegument Protein VP22 Abrogates cGAS/STING-Mediated Antiviral Innate Immunity. J. Virol. 2018, 92, JVI-00841. [Google Scholar] [CrossRef] [PubMed]
- Lin, L.; Qin, Y.; Wu, H.; Chen, Y.; Wu, S.; Si, X.; Wang, H.; Wang, T.; Zhong, X.; Zhai, X.; et al. Pyrrolidine dithiocarbamate inhibits enterovirus 71 replication by down-regulating ubiquitin-proteasome system. Virus Res. 2015, 195, 207–216. [Google Scholar] [CrossRef] [PubMed]
- Si, X.; McManus, B.M.; Zhang, J.; Yuan, J.; Cheung, C.; Esfandiarei, M.; Suarez, A.; Morgan, A.; Luo, H. Pyrrolidine dithiocarbamate reduces coxsackievirus B3 replication through inhibition of the ubiquitin-proteasome pathway. J. Virol. 2005, 79, 8014–8023. [Google Scholar] [CrossRef] [PubMed]
- Kanarek, N.; Ben-Neriah, Y. Regulation of NF-κB by ubiquitination and degradation of the IκBs. Immunol. Rev. 2012, 246, 77–94. [Google Scholar] [CrossRef] [PubMed]
- Kwon, S.K.; Saindane, M.; Baek, K.H. p53 stability is regulated by diverse deubiquitinating enzymes. Biochim. Biophys. Acta 2017, 1868, 404–411. [Google Scholar] [CrossRef] [PubMed]
- Rivas, C.; Aaronson, S.A.; Munoz-Fontela, C. Dual Role of p53 in Innate Antiviral Immunity. Viruses 2010, 2, 298–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taura, M.; Eguma, A.; Suico, M.A.; Shuto, T.; Koga, T.; Komatsu, K.; Komune, T.; Sato, T.; Saya, H.; Li, J.D.; et al. p53 regulates Toll-like receptor 3 expression and function in human epithelial cell lines. Mol. Cell. Biol. 2008, 28, 6557–6567. [Google Scholar] [CrossRef] [PubMed]
- Gregory, D.; Hargett, D.; Holmes, D.; Money, E.; Bachenheimer, S.L. Efficient replication by herpes simplex virus type 1 involves activation of the IκB kinase-IκB-p65 pathway. J. Virol. 2004, 78, 13582–13590. [Google Scholar] [CrossRef] [PubMed]
- Hargett, D.; McLean, T.; Bachenheimer, S.L. Herpes simplex virus ICP27 activation of stress kinases JNK and p38. J. Virol. 2005, 79, 8348–8360. [Google Scholar] [CrossRef] [PubMed]
- Song, S.; Qiu, M.; Chu, Y.; Chen, D.; Wang, X.; Su, A.; Wu, Z. Downregulation of cellular c-Jun N-terminal protein kinase and NF-κB activation by berberine may result in inhibition of herpes simplex virus replication. Antimicrob. Agents Chemother. 2014, 58, 5068–5078. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Su, A.; Fu, Y.; Wang, X.; Lv, X.; Xu, W.; Xu, S.; Wang, H.; Wu, Z. Harmine blocks herpes simplex virus infection through downregulating cellular NF-κB and MAPK pathways induced by oxidative stress. Antivir. Res. 2015, 123, 27–38. [Google Scholar] [CrossRef] [PubMed]
- Yoon, M.; Spear, P.G. Disruption of adherens junctions liberates nectin-1 to serve as receptor for herpes simplex virus and pseudorabies virus entry. J. Virol. 2002, 76, 7203–7208. [Google Scholar] [CrossRef] [PubMed]
- Horbul, J.E.; Schmechel, S.C.; Miller, B.R.; Rice, S.A.; Southern, P.J. Herpes simplex virus-induced epithelial damage and susceptibility to human immunodeficiency virus type 1 infection in human cervical organ culture. PLoS ONE 2011, 6, e22638. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Sequence | |
|---|---|---|
| Forward (5′-3′) | Reverse (5′-3′) | |
| GAPDH | GGTGGTCTCCTCTGACTTCAACA | GTTGCTGTAGCCAAATTCGTTGT |
| IFN-α | TTTCTCCTGCCTGAAGAACAG | GCTCATGATTTCTGCTCTGACA |
| IFN-β | GCCGCATTGACCATCTATGAGA | GAGATCTTCAGTTTCGGAGGTAAC |
| IFN-λ1 | CTTCCAAGCCCACCCCAACT | GGCCTCCAGGACCTTCAGC |
| IFN-λ2/3 | TTTAAGAGGGCCAAAGATGC | TGGGCTGAGGCTGGATACAG |
| IRF3 | ACCAGCCGTGGACCAAGAG | TACCAAGGCCCTGAGGCAC |
| IRF7 | TGGTCCTGGTGAAGCTGGAA | GATGTCGTCATAGAGGCTGTTGG |
| HSV-2 ICP0 | GTGCATGAAGACCTGGATTCC | GGTCACGCCCACTATCAGGTA |
| HSV-2 ICP27 | TTCTGCGATCCATATCCGAGC | AAACGGCATCCCGCCAAA |
| HSV-2 ICP8 | AGGACATAGAGACCATCGCGTTCA | TGGCCAGTTCGCTCACGTTATT |
| HSV-2 gC | AAATCCGATGCCGGTTTCCCAA | TTACCATCACCTCCTCTAAGCTAGGC |
| HSV-2 gD | ATCCGAACGCAGCCCCGC | TCTCCGTCCAGTCGTTTAT |
| HSV-2 DNA polymerase | GCTCGAGTGCGAAAAAACGTTC | CGGGGCGCTCGGCTAAC |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Y.; Xu, X.-Q.; Zhang, B.; Gu, J.; Meng, F.-Z.; Liu, H.; Zhou, L.; Wang, X.; Hou, W.; Ho, W.-Z. Bowman‒Birk Inhibitor Suppresses Herpes Simplex Virus Type 2 Infection of Human Cervical Epithelial Cells. Viruses 2018, 10, 557. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100557
Liu Y, Xu X-Q, Zhang B, Gu J, Meng F-Z, Liu H, Zhou L, Wang X, Hou W, Ho W-Z. Bowman‒Birk Inhibitor Suppresses Herpes Simplex Virus Type 2 Infection of Human Cervical Epithelial Cells. Viruses. 2018; 10(10):557. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100557
Chicago/Turabian StyleLiu, Yu, Xi-Qiu Xu, Biao Zhang, Jun Gu, Feng-Zhen Meng, Hang Liu, Li Zhou, Xu Wang, Wei Hou, and Wen-Zhe Ho. 2018. "Bowman‒Birk Inhibitor Suppresses Herpes Simplex Virus Type 2 Infection of Human Cervical Epithelial Cells" Viruses 10, no. 10: 557. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100557