Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti

 , and
, and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Mosquitoes
2.2. Infection of HWE Mosquitoes with Viruses
2.3. Generation of cDNA Templates
2.4. Taqman qPCR Assays for the Virus-Specific Detection of Plus- and Minus-Strand RNAs
2.5. Midgut Sample Preparation for SEM
2.6. Midgut Sample Preparation for TEM
2.7. Analyzing CHIK Virion Density in Midgut Epithelial Cells and Measuring Midgut BL Thickness in TEM Images
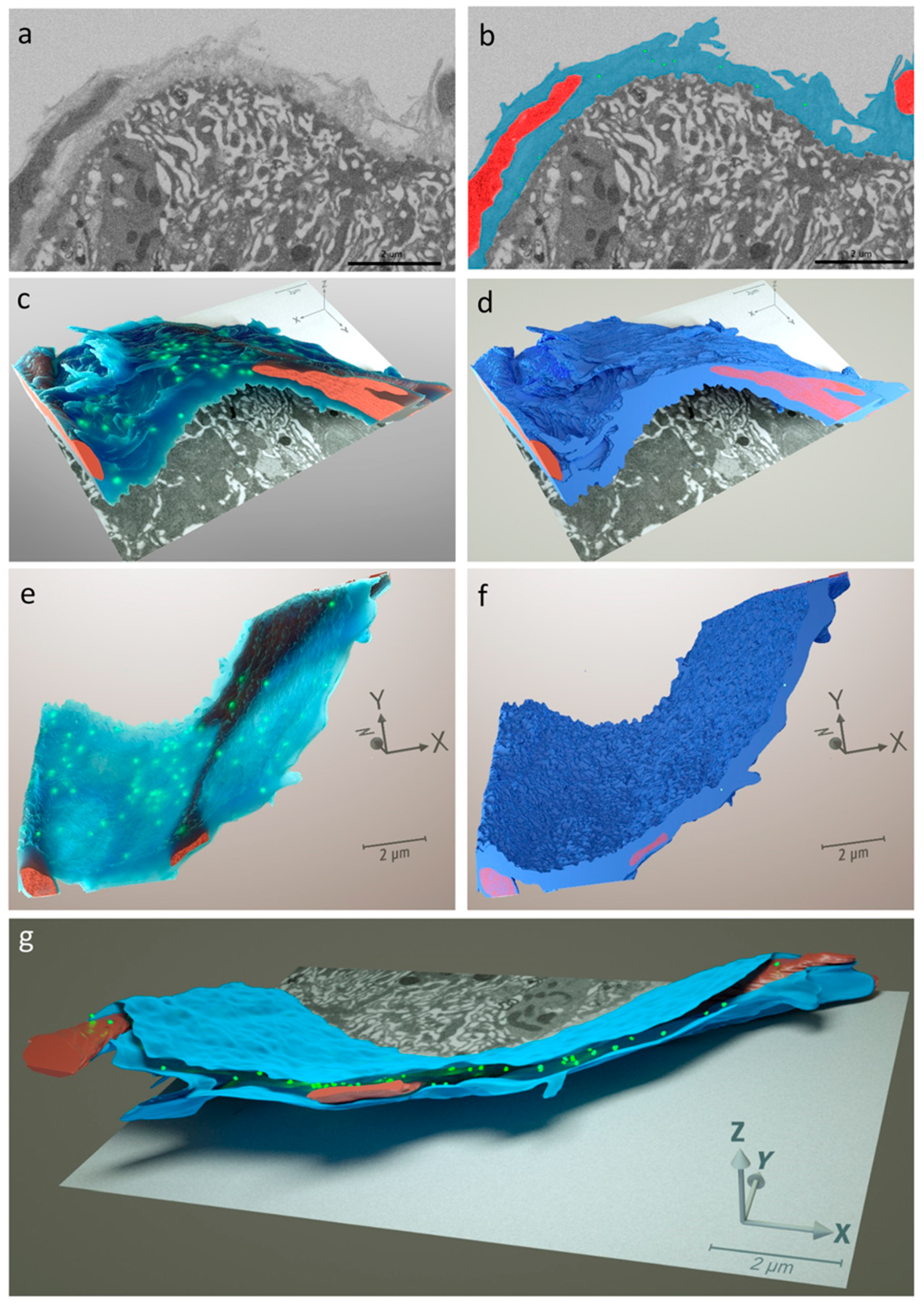
2.8. Midgut Sample Preparation for Serial Block Face Imaging via FIB-SEM
3. Results
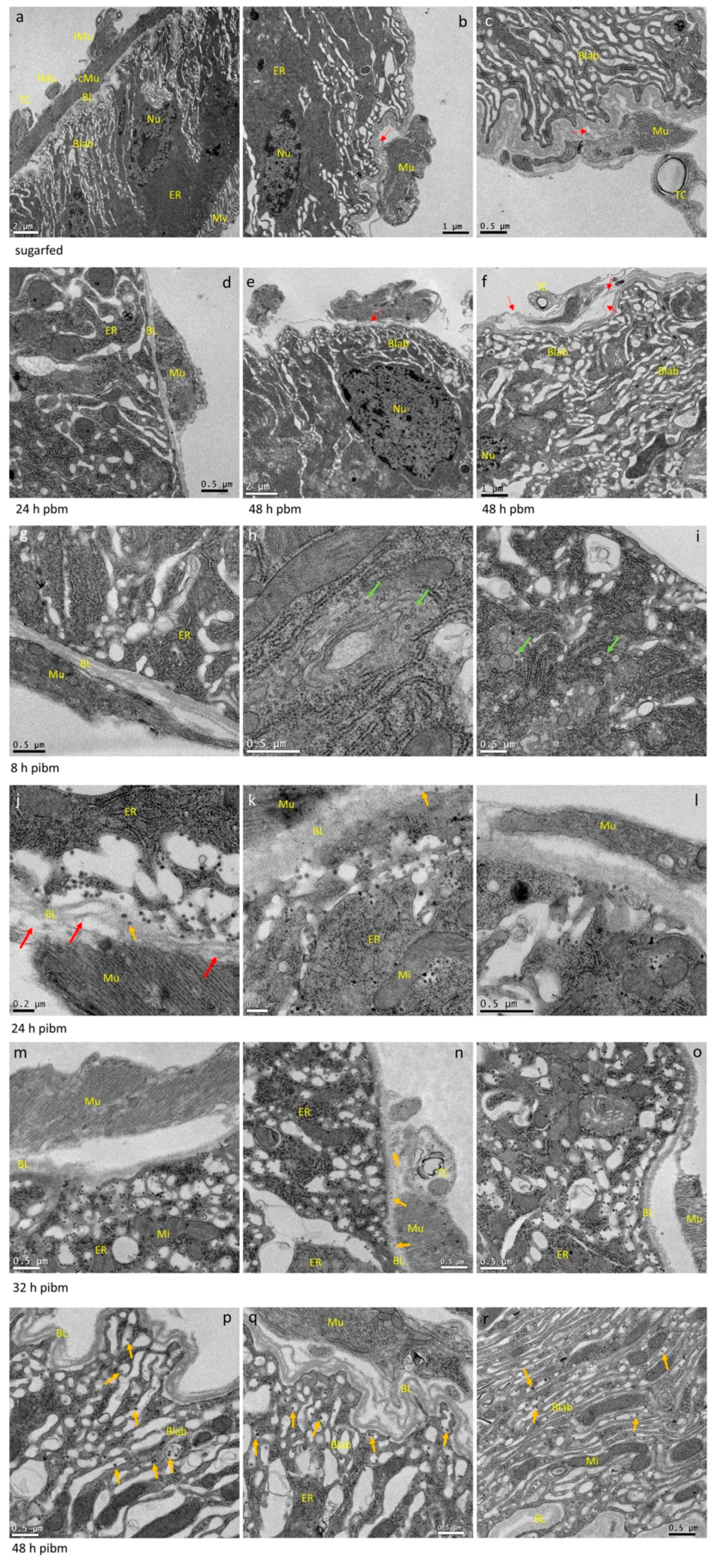
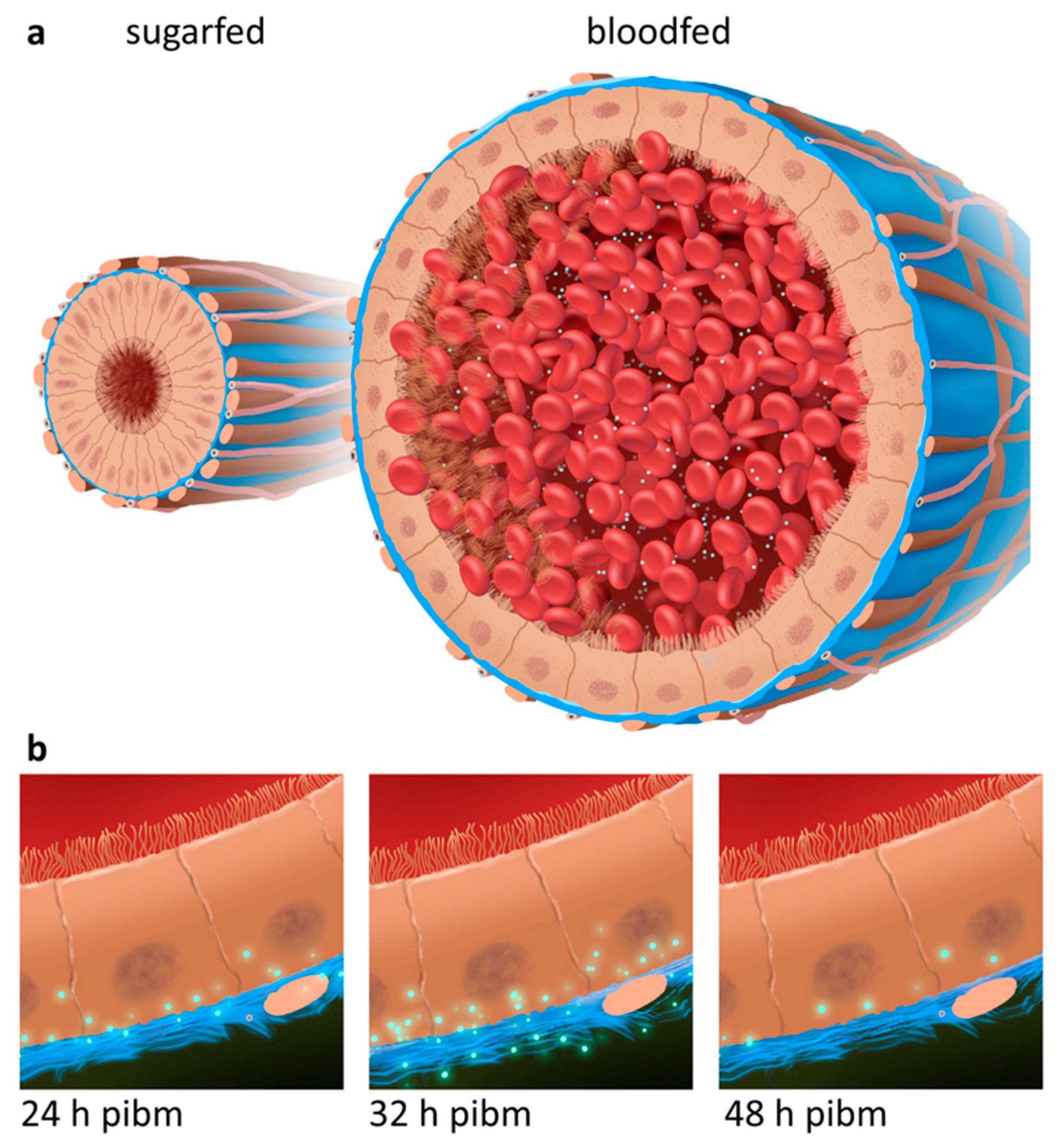
3.1. Bloodmeal Ingestion Causes Temporal Midgut Tissue Expansion and Damage to the BL
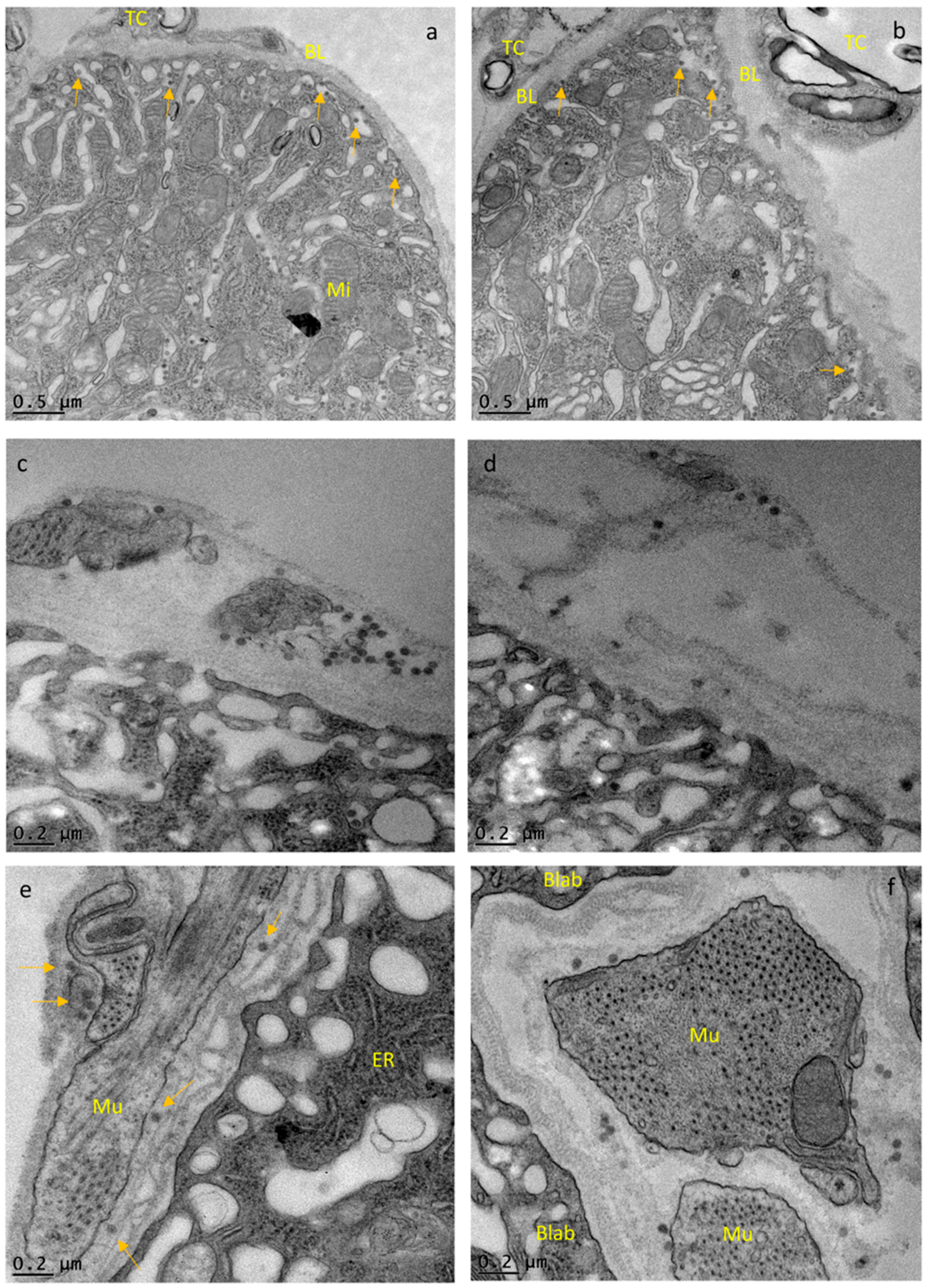
3.2. CHIKV Disseminates from the Midgut within a Narrow Time Window during Bloodmeal Digestion
3.3. Repeated Bloodmeals Enhance Virion Dissemination from the Midgut Epithelium
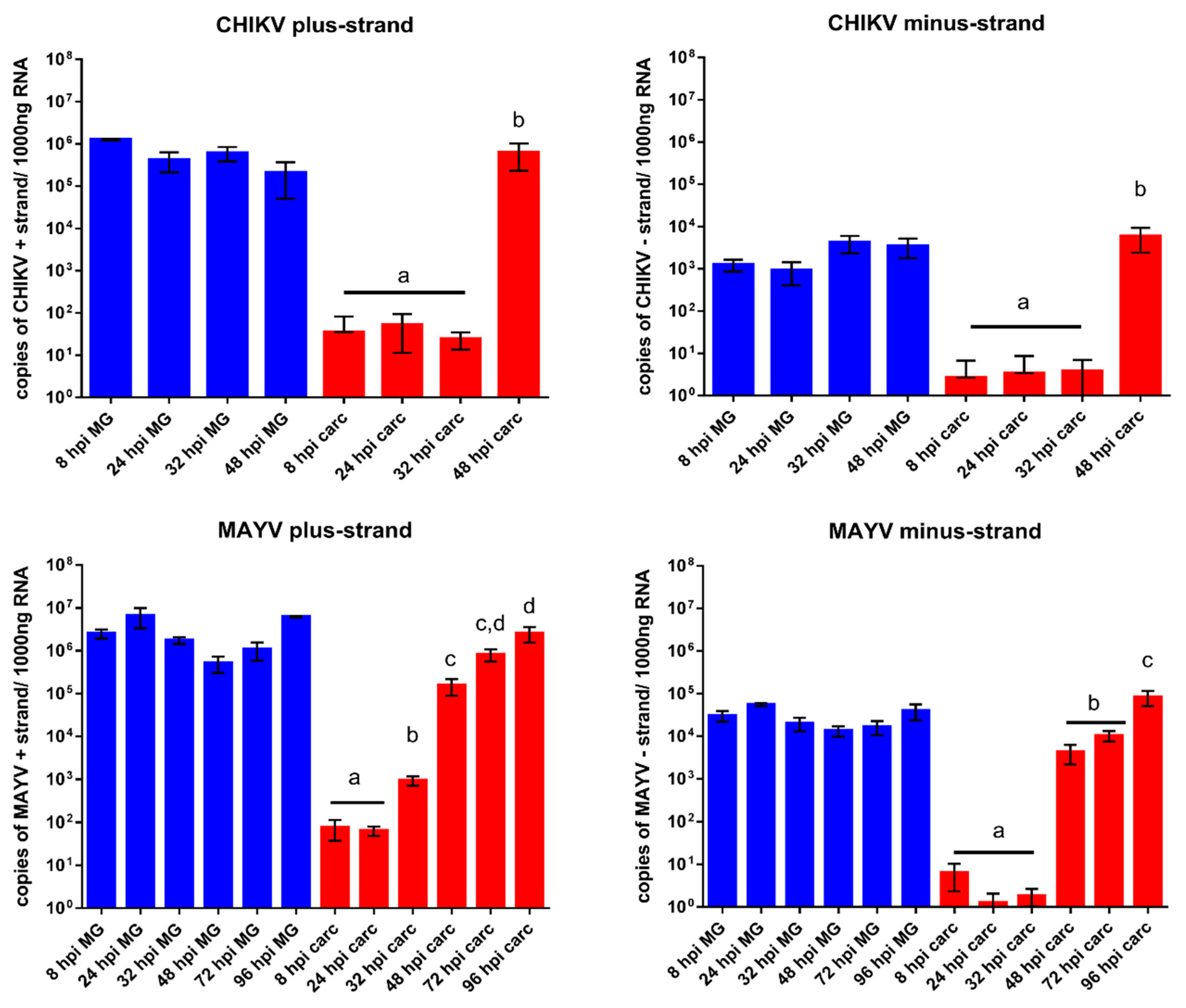
3.4. Quantification of Alphavirus RNA in Infected Midguts and Carcasses over Time Indicates that Virus Dissemination from the Midgut Occurs within 48 h pibm
4. Discussion
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Powers, A.M.; Roehrig, J.T. Alphaviruses. Methods Mol. Biol. 2011, 665, 17–38. [Google Scholar] [PubMed]
- Powers, A.M. Chikungunya. Clin. Lab. Med. 2010, 30, 209–219. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Reisen, W.K. Present and future arboviral threats. Antiviral Res. 2010, 85, 328–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fischer, M.; Staples, J.E.; Arboviral Diseases Branch, National Center for Emerging and Zoonotic Infectious Diseases, CDC. Notes from the field: Chikungunya virus spreads in the Americas-Caribbean and South America, 2013–2014. MMWR Morb. Mortal. Wkly. Rep. 2014, 63, 500–501. [Google Scholar] [PubMed]
- Morrison, T.E. Reemergence of chikungunya virus. J. Virol. 2014, 88, 11644–11647. [Google Scholar] [CrossRef] [PubMed]
- Hecker, H. Structure and function of midgut epithelial cells in Culicidae mosquitoes (Insecta, Diptera). Cell Tissue Res. 1977, 184, 321–341. [Google Scholar] [CrossRef] [PubMed]
- Houk, E.J. Midgut ultrastructure of Culex tarsalis (Diptera: Culicidae) before and after a blood meal. Tissue Cell 1977, 9, 103–118. [Google Scholar] [CrossRef]
- Franz, A.W.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue barriers to arbovirus infection in mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [Green Version]
- Solignat, M.; Gay, B.; Higgs, S.; Briant, L.; Devaux, C. Replication cycle of chikungunya: A re-emerging arbovirus. Virology 2009, 393, 183–197. [Google Scholar] [CrossRef] [PubMed]
- Hecker, H.; Rudin, W. Morphometric parameters of the midgut cells of Aedes aegypti L. (Insecta, Diptera) under various conditions. Cell Tissue Res. 1981, 219, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Okuda, K.; de Souza Caroci, A.; Ribolla, P.E.; de Bianchi, A.G.; Bijovsky, A.T. Functional morphology of adult female Culex quinquefasciatus midgut during blood digestion. Tissue Cell 2002, 34, 210–219. [Google Scholar] [CrossRef]
- Wengler, G. The regulation of disassembly of alphavirus cores. Arch. Virol. 2009, 154, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Strauss, J.H.; Strauss, E.G. The alphaviruses: Gene expression, replication, and evolution. Microbiol. Rev. 1994, 58, 491–562. [Google Scholar]
- Smith, D.R.; Arrigo, N.C.; Leal, G.; Muehlberger, L.E.; Weaver, S.C. Infection and dissemination of Venezuelan equine encephalitis virus in the epidemic mosquito vector, Aedes taeniorhynchus. Am. J. Trop. Med. Hyg. 2007, 77, 176–187. [Google Scholar] [CrossRef] [PubMed]
- Myles, K.M.; Pierro, D.J.; Olson, K.E. Comparison of the transmission potential of two genetically distinct Sindbis viruses after oral infection of Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2004, 41, 95–106. [Google Scholar] [CrossRef] [PubMed]
- Parikh, G.R.; Oliver, J.D.; Bartholomay, L.C. A haemocyte tropism for an arbovirus. J. Gen. Virol. 2009, 90, 292–296. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Houk, E.J.; Hardy, J.L.; Chiles, R.E. Permeability of the midgut basal lamina in the mosquito, Culex tarsalis Coquillett (Insecta, Diptera). Acta. Trop. 1981, 38, 163–171. [Google Scholar]
- Dong, S.; Balaraman, V.; Kantor, A.M.; Lin, J.; Grant, D.G.; Held, N.L.; Franz, A.W.E. Chikungunya virus dissemination from the midgut of Aedes aegypti is associated with temporal basal lamina degradation during bloodmeal digestion. PLoS Negl. Trop. Dis. 2017, 11, e0005976. [Google Scholar] [CrossRef]
- Houk, E.J.; Kramer, L.D.; Hardy, J.L.; Chiles, R.E. Western equine encephalomyelitis virus: In vivo infection and morphogenesis in mosquito mesenteronal epithelial cells. Virus Res. 1985, 2, 123–138. [Google Scholar] [CrossRef]
- Hardy, J.L.; Houk, E.J.; Kramer, L.D.; Reeves, W.C. Intrinsic factors affecting vector competence of mosquitoes for arboviruses. Annu. Rev. Entomol. 1983, 28, 229–262. [Google Scholar] [CrossRef] [PubMed]
- Romoser, W.S.; Wasieloski, L.P., Jr.; Pushko, P.; Kondig, J.P.; Lerdthusnee, K.; Neira, M.; Ludwig, G.V. Evidence for arbovirus dissemination conduits from the mosquito (Diptera: Culicidae) midgut. J. Med. Entomol. 2004, 41, 467–475. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, S.G.; Murphy, F.A.; Sudia, W.D. St. Louis encephalitis virus: An ultrastructural study of infection in a mosquito vector. Virology 1973, 56, 70–87. [Google Scholar] [CrossRef]
- Weaver, S.C. Electron microscopic analysis of infection patterns for Venezuelan equineencephalomyelitis virus in the vector mosquito, Culex (Melanoconion) taeniopus. Am. J. Trop. Med. Hyg. 1986, 35, 624–631. [Google Scholar] [CrossRef] [PubMed]
- Lerdthusnee, K.; Romoser, W.S.; Faran, M.E.; Dohm, D.J. Rift Valley fever virus in the cardia of Culex pipiens: An immunocytochemical and ultrastructural study. Am. J. Trop. Med. Hyg. 1995, 53, 331–337. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Popov, V.; Wen, J.; Han, V.; Higgs, S. Ultrastructural study of West Nile virus pathogenesis in Culex pipiens quinquefasciatus (Diptera: Culicidae). J. Med. Entomol. 2005, 42, 429–444. [Google Scholar] [CrossRef] [PubMed]
- Hernandez, R.; Brown, D.T.; Paredes, A. Structural differences observed in arboviruses of the alphavirus and flavivirus genera. Adv. Virol. 2014, 2014, 259382. [Google Scholar] [CrossRef] [PubMed]
- Fernandez de Castro, I.; Risco, C. Imaging RNA virus replication assemblies: Bunyaviruses and reoviruses. Future Virol. 2014, 9, 1089–1104. [Google Scholar] [CrossRef]
- Scott, T.W.; Hildreth, S.W.; Beaty, B.J. The distribution and development of eastern equine encephalitis virus in its enzootic mosquito vector, Culiseta melanura. Am. J. Trop. Med. Hyg. 1984, 33, 300–310. [Google Scholar] [CrossRef] [PubMed]
- Larsen, J.R.; Ashley, R.F. Demonstration of Venezuelan equine encephalomyelitis virus in tissues of Aedes aegypti. Am. J. Trop. Med. Hyg. 1971, 20, 754–760. [Google Scholar] [CrossRef] [PubMed]
- Scott, T.W.; Burrage, T.G. Rapid infection of salivary glands in Culiseta melanura with eastern equine encephalitis virus: An electron microscopic study. Am. J. Trop. Med. Hyg. 1984, 33, 961–964. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H.; Lerdthusnee, K.; Romoser, W.S. Togavirus-associated pathologic changes in the midgut of a natural mosquito vector. J. Virol. 1988, 62, 2083–2090. [Google Scholar] [PubMed]
- Weaver, S.C.; Lorenz, L.H.; Scott, T.W. Pathologic changes in the midgut of Culex tarsalis following infection with Western equine encephalomyelitis virus. Am. J. Trop. Med. Hyg. 1992, 47, 691–701. [Google Scholar] [CrossRef] [PubMed]
- Vega-Rúa, A.; Schmitt, C.; Bonne, I.; Krijnse Locker, J.; Failloux, A.B. Chikungunya virus replication in salivary glands of the mosquito Aedes albopictus. Viruses 2015, 7, 5902–5907. [Google Scholar] [CrossRef] [PubMed]
- Soekiman, S.; Matsumura, T.; Yamanishi, H. Multiplication of chikungunya virus in salivary glands of Aedes albopictus (Oahu strain) mosquitoes: An electron microscopic study. Jpn. J. Med. Sci. Biol. 1986, 39, 207–211. [Google Scholar] [CrossRef] [PubMed]
- Janzen, H.G.; Rhodes, A.J.; Doane, F.W. Chikungunya virus in salivary glands of Aedes aegypti (L.): An electron microscope study. Can. J. Microbiol. 1970, 16, 581–586. [Google Scholar] [CrossRef] [PubMed]
- Romoser, W.S.; Faran, M.E.; Bailey, C.L. Newly recognized route of arbovirus dissemination from the mosquito (Diptera: Culicidae) midgut. J. Med. Entomol. 1987, 24, 431–432. [Google Scholar] [CrossRef] [PubMed]
- Romoser, W.S.; Faran, M.E.; Bailey, C.L.; Lerdthusnee, K. An immunocytochemical study of the distribution of Rift Valley fever virus in the mosquito Culex pipiens. Am. J. Trop. Med. Hyg. 1992, 46, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Foy, B.D.; Myles, K.M.; Pierro, D.J.; Sanchez-Vargas, I.; Uhlírová, M.; Jindra, M.; Beaty, B.J.; Olson, K.E. Development of a new Sindbis virus transducing system and its characterization in three Culicine mosquitoes and two Lepidopteran species. Insect Mol. Biol. 2004, 13, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H.; Repik, P.M. Detection of eastern equine encephalomyelitis virus deposition in Culiseta melanura following ingestion of radiolabeled virus in blood meals. Am. J. Trop. Med. Hyg. 1991, 44, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Brault, A.C.; Foy, B.D.; Myles, K.M.; Kelly, C.L.; Higgs, S.; Weaver, S.C.; Olson, K.E.; Miller, B.R.; Powers, A.M. Infection patterns of o’nyong nyong virus in the malaria-transmitting mosquito, Anopheles gambiae. Insect Mol. Biol. 2004, 13, 625–635. [Google Scholar] [CrossRef] [PubMed]
- Salazar, M.I.; Richardson, J.H.; Sánchez-Vargas, I.; Olson, K.E.; Beaty, B.J. Dengue virus type 2: Replication and tropisms in orally infected Aedes aegypti mosquitoes. BMC Microbiol. 2007, 7, 9. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B.; Houk, E.J. Dissemination barriers for western equine encephalomyelitis virus in Culex tarsalis infected after ingestion of low viral doses. Am. J. Trop. Med. Hyg. 1981, 30, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Flick, D.; Fleming, K.H.; Jochim, R.; Beaty, B.J.; Black, W.C., 4th. Quantitative trait loci that control dengue-2 virus dissemination in the mosquito Aedes aegypti. Genetics 2005, 170, 185–194. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Beaty, B.J.; Black, W.C., 4th. Selection of D2S3, an Aedes aegypti (Diptera: Culicidae) strain with high oral susceptibility to Dengue 2 virus and D2MEB, a strain with a midgut barrier to Dengue 2 escape. J. Med. Entomol. 2005, 42, 110–119. [Google Scholar] [PubMed]
- Khoo, C.C.; Doty, J.B.; Held, N.L.; Olson, K.E.; Franz, A.W. Isolation of midgut escape mutants of two American genotype dengue 2 viruses from Aedes aegypti. Virol. J. 2013, 10, 257. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Behura, S.K.; Franz, A.W.E. The midgut transcriptome of Aedes aegypti fed with saline or protein meals containing chikungunya virus reveals genes potentially involved in viral midgut escape. BMC Genom. 2017, 18, 382. [Google Scholar] [CrossRef] [PubMed]
- Hardy, J.L. Susceptibility and resistance of vector mosquitoes, In The Arboviruses: Ecology and Epidemiology, Vol. I; Monath, T.P., Ed.; CRC Press: Boca Raton, FL, USA, 1988; pp. 87–126. [Google Scholar]
- Reddy, J.T.; Locke, M. The size limited penetration of gold particles through insect basal lamina. J. Insect Physiol. 1990, 36, 397–407. [Google Scholar] [CrossRef]
- Kantor, A.M.; Dong, S.; Held, N.L.; Ishimwe, E.; Passarelli, A.L.; Clem, R.J.; Franz, A.W. Identification and initial characterization of matrix metalloproteinases in the yellow fever mosquito, Aedes aegypti. Insect Mol. Biol. 2017, 26, 113–126. [Google Scholar] [CrossRef] [PubMed]
- Drobne, D.; Milani, M.; Zrimec, A.; Zrimec, M.B.; Tatti, F.; Draslar, K. Focused ion beam/scanning electron microscopy studies of Porcellio scaber (Isopoda, Crustacea) digestive gland epithelium cells. Scanning 2005, 27, 30–34. [Google Scholar] [CrossRef] [PubMed]
- Heymann, J.A.; Hayles, M.; Gestmann, I.; Giannuzzi, L.A.; Lich, B.; Subramaniam, S. Site-specific 3D imaging of cells and tissues with a dual beam microscope. J. Struct. Biol. 2006, 155, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carpenter, K.J.; Weber, P.K.; Davisson, M.L.; Pett-Ridge, J.; Haverty, M.I.; Keeling, P.J. Correlated SEM, FIB-SEM, TEM, and NanoSIMS imaging of microbes from the hindgut of a lower termite: Methods for in situ functional and ecological studies of uncultivable microbes. Microsc. Microanal. 2013, 19, 1490–1501. [Google Scholar] [CrossRef] [PubMed]
- Kizilyaprak, C.; Daraspe, J.; Humbel, B.M. Focused ion beam scanning electron microscopy in biology. J. Microsc. 2014, 254, 109–114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Giulio, A.; Muzzi, M. Two novel approaches to study arthropod anatomy by using dualbeam FIB/SEM. Micron 2018, 106, 21–26. [Google Scholar] [CrossRef] [PubMed]
- Long, K.C.; Ziegler, S.A.; Thangamani, S.; Hausser, N.L.; Kochel, T.J.; Higgs, S.; Tesh, R.B. Experimental transmission of Mayaro virus by Aedes aegypti. Am. J. Trop. Med. Hyg. 2011, 85, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Auguste, A.J.; Liria, J.; Forrester, N.L.; Giambalvo, D.; Moncada, M.; Long, K.C.; Morón, D.; de Manzione, N.; Tesh, R.B.; Halsey, E.S.; et al. Evolutionary and Ecological Characterization of Mayaro Virus Strains Isolated during an Outbreak, Venezuela, 2010. Emerg. Infect. Dis. 2015, 21, 1742–1750. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Kantor, A.M.; Lin, J.; Passarelli, A.L.; Clem, R.J.; Franz, A.W. Infection pattern and transmission potential of chikungunya virus in two New World laboratory-adapted Aedes aegypti strains. Sci. Rep. 2016, 6, 24729. [Google Scholar] [CrossRef] [PubMed]
- Anderson, C.R.; Downs, W.G.; Wattley, G.H.; Ahin, N.W.; Reese, A.A. Mayaro virus: A new human disease agent. II. Isolation from blood of patients in Trinidad, B.W.I. Am. J. Trop. Med. Hyg. 1957, 6, 1012–1016. [Google Scholar] [CrossRef] [PubMed]
- Deerinck, T.J.; Bushong, E.; Thor, A.; Ellisman, M.H. NCMIR Methods for 3D EM: A New Protocol for Preparation of Biological Specimens for Serial Block-Face SEM. SBEM Protocol v7_01_2010. Available online: https://ncmir.ucsd.edu/sbem-protocol (accessed on 17 October 2018).
- Villinger, C.; Schauflinger, M.; Gregorius, H.; Kranz, C.; Höhn, K.; Nafeey, S.; Walther, P. Three-dimensional imaging of adherent cells using FIB/SEM and STEM. Methods Mol. Biol. 2014, 1117, 617–638. [Google Scholar] [PubMed]
- Yao, J.S.; Strauss, E.G.; Strauss, J.H. Interactions between PE2, E1, and 6K required for assembly of alphaviruses studied with chimeric viruses. J. Virol. 1996, 70, 7910–7920. [Google Scholar] [PubMed]
- Lanciotti, R.S.; Kosoy, O.L.; Laven, J.J.; Panella, A.J.; Velez, J.O.; Lambert, A.J.; Campbell, G.L. Chikungunya virus in US travelers returning from India, 2006. Emerg. Infect. Dis. 2007, 13, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Mukhopadhyay, S.; Chipman, P.R.; Hong, E.M.; Kuhn, R.J.; Rossmann, M.G. In vitro-assembled alphavirus core-like particles maintain a structure similar to that of nucleocapsid cores in mature virus. J. Virol. 2002, 76, 11128–11132. [Google Scholar] [CrossRef] [PubMed]
- Farjana, T.; Tuno, N. Multiple blood feeding and host-seeking behavior in Aedes aegypti and Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2013, 50, 838–846. [Google Scholar] [CrossRef] [PubMed]
- Weaver, S.C.; Scott, T.W.; Lorenz, L.H. Patterns of eastern equine encephalomyelitis virus infection in Culiseta melanura (Diptera: Culicidae). J. Med. Entomol. 1990, 27, 878–891. [Google Scholar] [CrossRef] [PubMed]
- Romoser, W.S.; Turell, M.J.; Lerdthusnee, K.; Neira, M.; Dohm, D.; Ludwig, G.; Wasieloski, L. Pathogenesis of Rift Valley fever virus in mosquitoes-tracheal conduits and the basal lamina as an extra-cellular barrier. In Infectious Diseases from Nature: Mechanisms of Viral Emergence and Persistence; Peters, C.J., Calisher, C.H., Eds.; Springer: Vienna, Austria, 2005; pp. 89–100. [Google Scholar]
- Bowers, D.F.; Abell, B.A.; Brown, D.T. Replication and tissue tropism of the alphavirus Sindbis in the mosquito Aedes albopictus. Virology 1995, 212, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, R.; Scott, T.W. Apoptosis in mosquito midgut epithelia associated with West Nile virus infection. Apoptosis 2006, 11, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Gort, T.; Boyle, D.L.; Clem, R.J. Effects of manipulating apoptosis on Sindbis virus infection of Aedes aegypti mosquitoes. J. Virol. 2012, 86, 6546–6554. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Clem, R.J. Defining the core apoptosis pathway in the mosquito disease vector Aedes aegypti the roles of iap1, ark, dronc, and effector caspases. Apoptosis 2011, 16, 105–113. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, K.; Olson, B.J.; Huang, N.; Unis, D.; Clem, R.J. Rapid selection against arbovirus-induced apoptosis during infection of a mosquito vector. Proc. Natl. Acad. Sci. USA 2015, 112, E1152–E1161. [Google Scholar] [CrossRef] [PubMed]








© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kantor, A.M.; Grant, D.G.; Balaraman, V.; White, T.A.; Franz, A.W.E. Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti. Viruses 2018, 10, 571. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100571
Kantor AM, Grant DG, Balaraman V, White TA, Franz AWE. Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti. Viruses. 2018; 10(10):571. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100571
Chicago/Turabian StyleKantor, Asher M., DeAna G. Grant, Velmurugan Balaraman, Tommi A. White, and Alexander W. E. Franz. 2018. "Ultrastructural Analysis of Chikungunya Virus Dissemination from the Midgut of the Yellow Fever Mosquito, Aedes aegypti" Viruses 10, no. 10: 571. https://0-doi-org.brum.beds.ac.uk/10.3390/v10100571