Complete Nucleotide Sequence of a Partitivirus from Rhizoctonia solani AG-1 IA Strain C24


Guangdong Province Key Laboratory of Microbial Signals and Disease Control, College of Agriculture, South China Agricultural University, Guangzhou 510642, China
*
Authors to whom correspondence should be addressed.
Viruses 2018, 10(12), 703; https://0-doi-org.brum.beds.ac.uk/10.3390/v10120703
Submission received: 29 September 2018
/
Revised: 21 November 2018
/
Accepted: 7 December 2018
/
Published: 11 December 2018
(This article belongs to the Special Issue Mycoviruses)
{kind=link}
{kind=link}
Abstract
:The complete genome of a novel double-stranded (ds) RNA mycovirus, named as Rhizoctonia solani partitivirus 5 (RsPV5), isolated from rice sheath blight fungus R. solani AG-1 IA strain C24, was sequenced and analysed. RsPV5 consists of two segments, dsRNA-1 (1899 nucleotides) and dsRNA-2 (1787 nucleotides). DsRNA-1 has an open reading frame (ORF) 1 that potentially codes for a protein of 584 amino acid (aa) containing the conserved motifs of a RNA-dependent RNA polymerase (RdRp), and dsRNA-2 also contains a ORF 2, encoding a putative capsid protein (CP) of 513 aa. Phylogenetic analysis revealed that RsPV5 clustered together with six other viruses in an independent clade of the genus Alphapartitivirus, indicating that RsPV5 was a new member of the genus Alphapartitivirus, within the family Partitiviridae.
1. Introduction
Mycoviruses (fungal viruses or viruses of fungi) are widely distributed in fungi, of which, only a few affect their fungal hosts resulting in alterations of growth rate or enhanced virulence or hypovirulence [1,2]. Mycoviruses with hypovirulent traits are anticipated to be important biological control agents against plant fungal diseases in the future [3]. Since the successful application of a mycovirus for biological control of chestnut blight [4], mycovirus research from major groups of fungi has attracted attention. A wide range of mycoviruses, such as Rosellinia necatrix partitivirus 1-W8 [5], Botrytis cinerea mitovirus 1 [6], and Rhizoctonia solani partitivirus 2 [7], have been discovered in different fungi. Currently, mycoviruses are mainly classified into 14 families, including 7 families of double-stranded (dsRNA) viruses (Partitiviridae, Totiviridae, Reoviridae, Chrysoviridae, Quadriviridae, Megabirnaviridae, Endornaviridae), 5 families of positive-strand RNA (+ssRNA) viruses (Barnaviridae, Alphaflexiviridae, Hypoviridae, Narnaviridae, Gammaflexiviridae), 1 family of negative-strand RNA (-ssRNA) viruses (Mycomononegaviridae) and family Amalgamaviridae [8,9]. However many mycoviruses remain unclassified [8], and the first circular ssDNA mycovirus from the phytopathogenic fungus Sclerotinia sclerotiorum was discovered in 2010 [10].
Rhizoctonia solani Kühn [teleomorph: Thanatephorus cucumeris (Frank) Donk] is an economically important soil-borne fungal pathogen that causes severe plant diseases and disastrous economic losses in a wide variety of commercial crops including rice, maize and wheat [11,12]. R. solani is a common mycovirus host [7,13]. Investigations concerning the association of dsRNA with Rhizoctonia decline revitalised research on R. solani mycoviruses [14] which in turn revealed that mycoviruses are ubiquitous in natural R. solani isolates [7,13,15,16,17,18,19]. Subsequently complete genome sequences of several R. solani mycoviruses have been documented and their sequence properties and phylogene have been analysed. Thus far the R. solani mycoviruses described mainly belong to the genera Partitivirus [13,18,19,20], Mitovirus [12,21] and Endornavirus [22,23], along with some unclassified mycoviruses [7,24,25]. To date, the complete genomes of six mycoviruses, i.e. Rhizoctonia solani dsRNA virus 1 (RsRV1) [7], Rhizoctonia solani partitivirus 2 (RsPV2) [13], Rhizoctonia solani RNA virus HN008 (RsRV-HN008) [24], Rhizoctonia solani dsRNA virus 3 (RsRV3) [18], Rhizoctonia solani partitivirus 3 (RsPV3) [19] and Rhizoctonia solani partitivirus 4 (RsPV4) [19], from R. solani AG-1 IA have been reported, of which, RsRV1 and RsRV-HN008 are unclassified, while RsPV2, RsRV3, RsPV3 and RsPV4 belong to the genus Alphapartitivirus.
Investigations on the AG-1 IA isolate of R. solani, the causal agent of rice sheath blight revealed the presence of three novel mycoviruses in the authors’ laboratory [7,13,18]. In this study we describe the complete nucleotide sequence of another partitivirus nominated Rhizoctonia solani partitivirus 5 (RsPV5), isolated from R. solani AG-1 IA strain C24. The sequences of the two genomic components of RsPV5 were analysed and a phylogenetic tree was constructed based on the derived amino acid sequence of the putative RNA-dependent RNA polymerase (RdRp) to clarify the phylogenetic status of RsPV5. The phylogenetic analysis indicated that RsPV5 has the closest relationship with members of the genus Alphapartitivirus.
2. Materials and Methods
2.1. Fungal Strain
The C24 strain of R. solani AG-1 IA was used in this study, which was isolated from rice leaves with typical symptoms of rice sheath blight collected from Zhangzhou city, Fujian province, China, in 1999 and stored at −20 °C.
2.2. Isolation and Sequencing of Mycovirus dsRNA
Mycelia of the strain C24 were cultured on cellophane covered on potato dextrose agar (PDA) plates at 28 °C. After cultivation for 5 days, the mycelia were harvested and stored at −80 °C for further use. The lyophilized mycelia were ground into a fine powder with a mortar and pestle in the presence of liquid nitrogen. Viral dsRNAs were extracted using a slightly modified version of a CF-11 cellulose chromatography method as described by Morris and Dodds [26]. To remove contaminating DNA and single stranded RNA, the extracts were treated with DNase I and SI nuclease, and viral dsRNAs were separated and analysed by gel electrophoresis and visualization with ethidium bromide staining. The cDNA library was constructed using random primer (5′-CCTGAATTCGGATCCTCCNNNNNN-3′) along with reverse transcriptase, and amplified with specific primer (5′-CCTGAATTCGGATCCTCC-3′). To sequence the 5′ and 3′-termini of the dsRNA, a RACE procedure modified from that described by Potgieter et al. [27] was used. All PCR amplicons were cloned into the pMD18-T vector and transformed into Escherichia coli strain JM109. Plasmid DNA from recombinant clones was isolated and at least three clones for each fragment of sequence were sequenced in both directions. The complete nucleotide sequences of the two genomic components of RsPV5 were assembled and deposited in GenBank database with the accession numbers of MH715946 and MH715947.
2.3. Data Analysis
Sequence analysis and multiple alignments were actualized by DNAMAN and ClustalX. A phylogenetic tree was constructed on the basis of neighbor-joining (NJ) method using MEGA 6 with 1000 replicates.
3. Results
3.1. Genomic Structure Analysis
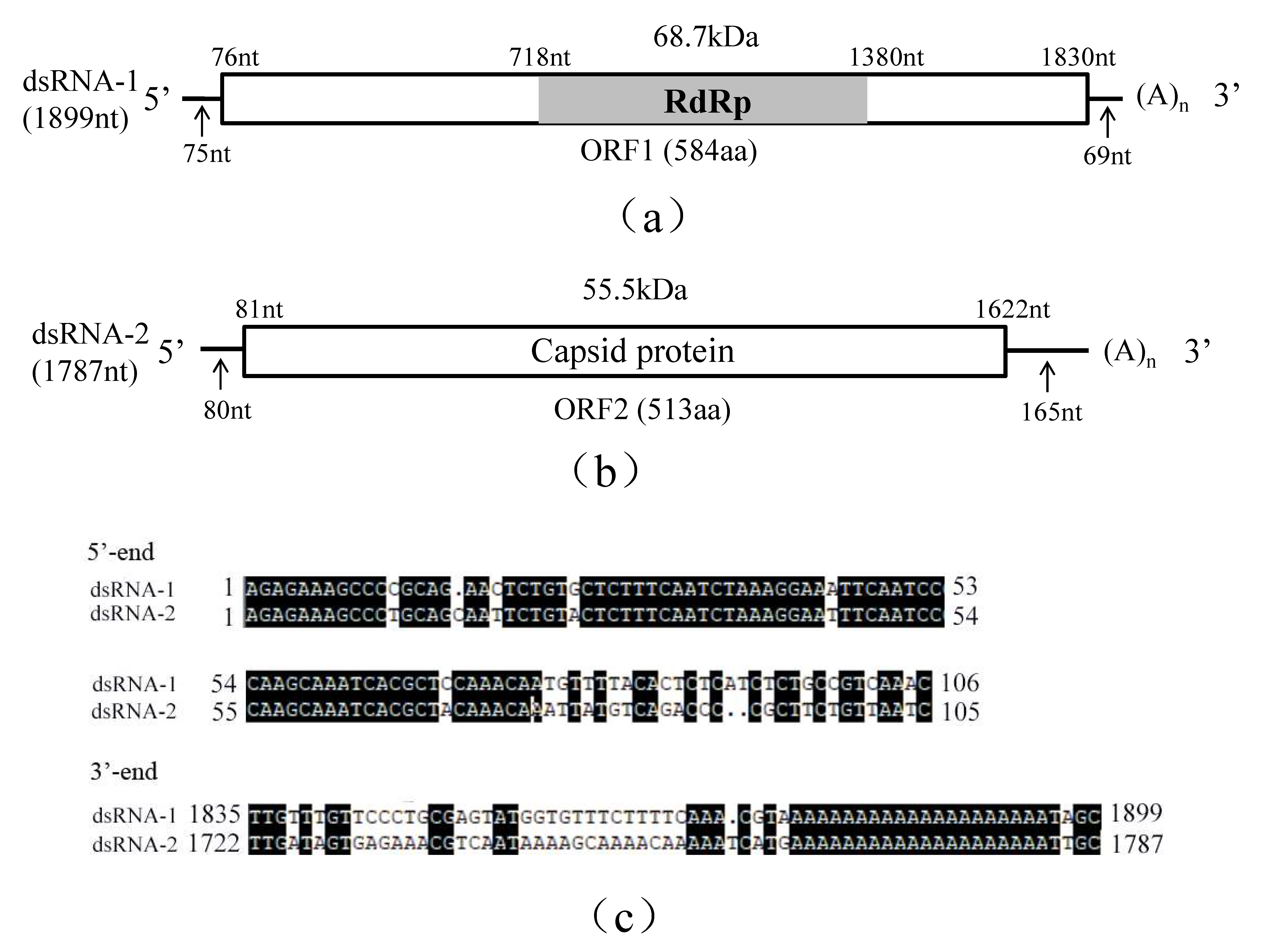
Sequence analysis revealed that R. solani strain C24 was infected by a novel virus, RsPV5, belonging the family Partitiviridae. The complete genome of RsPV5 is composed of two segments, designated dsRNA-1 and dsRNA-2, respectively (Figure 1a,b). A comparison of both dsRNA segments demonstrated that both 5′- and 3′-termini are conserved (Figure 1c). Additionally the 3′-ends of both dsRNAs were interrupted by poly(A) tails, a feature similar to some other members in the family Partitiviridae [13,28].
Analysis of the full-length cDNA sequence of dsRNA-1 indicated that it comprises 1899 nucleotides (nt), with a GC content of 45.98%, and contains an open reading frame 1 (ORF1), starting at nt 76 and terminating at nt 1830. ORF1 potentially encodes a 68.7 kDa protein of 584 amino acids (aa) containing sequence-conserved motifs characteristic for RNA-dependent RNA polymerase (RdRp; Figure 1a). The 5′-untranslated region (UTR) and 3′-UTR of dsRNA-1 consists of 75 nt and 69 nt, respectively. Homology searches with BLASTp confirmed that the protein was closely related to the RdRps of partitiviruses including Rhizoctonia solani dsRNA virus 3 (RsRV3, GenBank accession number: YP_009329886.2) with an aa identity of 82%, Heterobasidion partitivirus 12 (HetPV12, GenBank accession number: YP_009508051.1) with an aa identity of 61%, Heterobasidion partitivirus 13 (HetPV13, GenBank accession number: AHL25155.1) with an aa identity of 59%.
Analysis of the full-length cDNA sequence of dsRNA-2 indicated that it is 1787 bp in length with a GC content of 52.20% containing a single open reading frame 2 (ORF2) starting at nt 81 and terminating at nt 1622. DsRNA-2 potentially encodes a putative capsid protein (CP) of 513 aa that has an estimated molecular mass of 55.5 kDa (Figure 1b). The 5′-UTR and 3′-UTR of dsRNA2 are respectively 80 nt and 165 nt in length. BLASTp search revealed that this protein has 65% identity to RsRV3 CP gene (GenBank accession number: YP_009329885.2) and 30% identity to the HetPV13 CP gene (GenBank accession number: AHL25156.1).
3.2. Phylogenetic Analysis
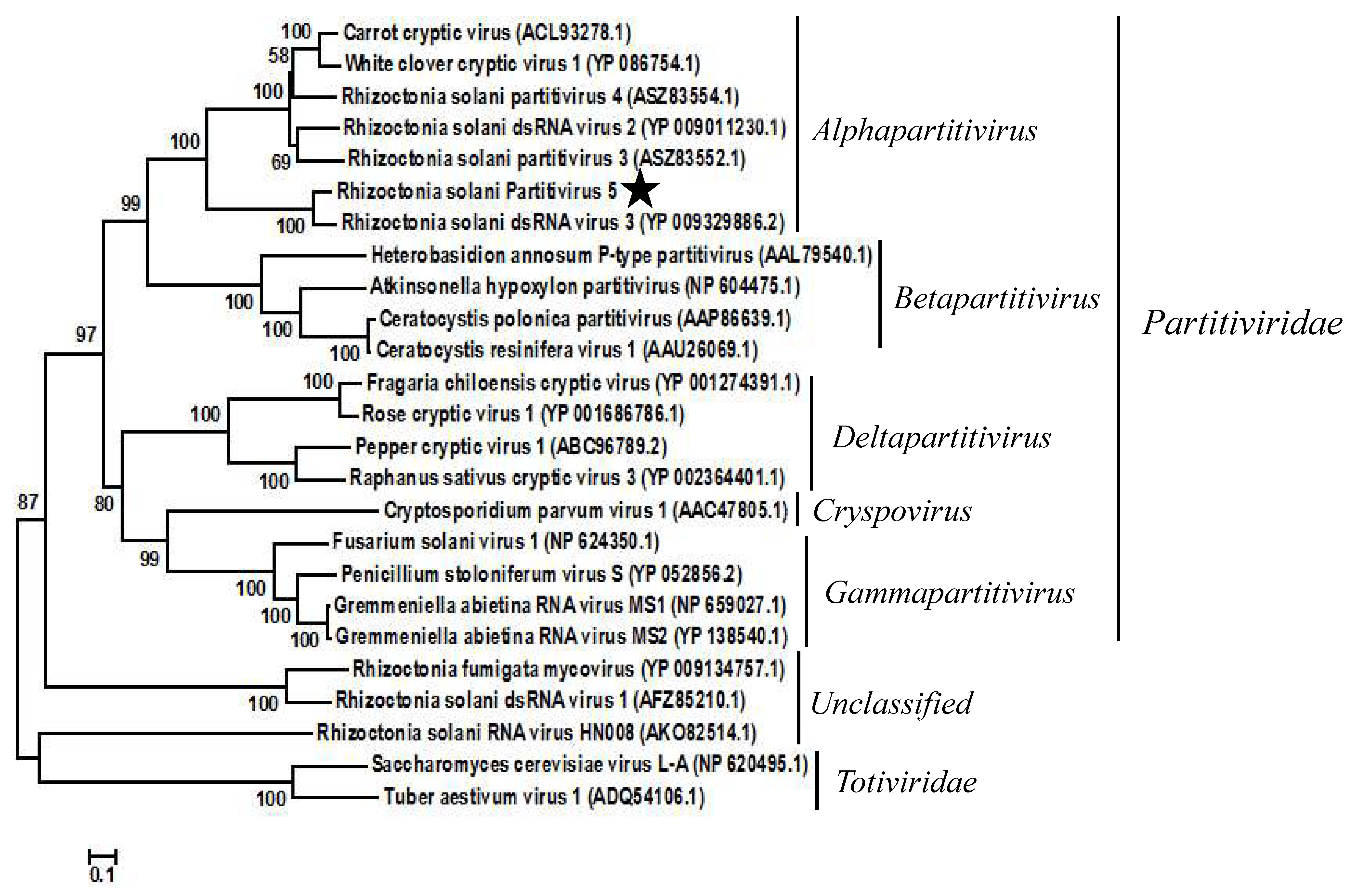
To confirm the taxonomic status of RsPV5, a phylogenetic tree was constructed based on the aa sequences of RdRp regions of RsPV5 and 24 other selected viruses in the families Partitiviridae and Totiviridae as well as the unclassified viruses (Figure 2). The result of phylogenetic analysis showed that RsPV5, Rhizoctonia solani dsRNA virus 3, Rhizoctonia solani dsRNA virus 2, Rhizoctonia solani partitivirus 3, Rhizoctonia solani partitivirus 4, White clover cryptic virus 1 and Carrot cryptic virus were clustered together in a distinct group belonging to the genus Alphapartitivirus. The phylogenetic tree illustrated that RsPV5 is a new member of the genus Alphapartitivirus in the family Partitiviridae. In addition, RsPV5 was placed in the same clade with RsRV3, a mycovirus of R. solani AG-1 IA previously identified in our laboratory [18], indicating that these two viruses have a close relationship. Furthermore, RsRV1 [7] and RsRV-HN008 [24] belong to the subclade of unclassified family. R. solani AG-1 IA appears to have an extensive virome which might expand further in the future with the advent of next generation sequencing.
4. Discussion
It can be seen from the above study, RsPV5, which infects R. solani AG-1 IA, is a novel dsRNA mycovirus of the genus Alphapartitivirus in the family Partitiviridae. So far, seven mycoviruses four of them were found in the authors’ laboratory [7,13,18] infecting Rhizoctonia solani AG1-IA have been reported and have proved to belong to different viral family [7,13,18,19,24], indicating a rich diversity of mycoviruses in R. solani AG-1 IA.
Author Contributions
C.L. performed the experiments, analysed the data, sequenced and analysed the virus, wrote the draft paper; M.Z. (Miaolin Zeng) and M.Z. (Meiling Zhang) provided advices for experimental operation, assisted in data analysis and paper revision; C.S. and E.Z. conceived, designed and supervised the experiments, reviewed and revised the paper.
Funding
This study is supported by the National Natural Science Foundation of China, “Genome structure and function analysis of dsRNA mycoviruses of Rhizoctonia solani AG-1 IA, the causal agent of rice sheath blight” (No. 31470247).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Pearson, M.N.; Beever, R.E.; Boine, B.; Arthur, K. Mycoviruses of filamentous fungi and their relevance to plant pathology. Mol. Plant Path. 2009, 10, 115–128. [Google Scholar] [CrossRef] [PubMed]
- Herrero, N.; Márquez, S.S.; Zabalgogeazcoa, I. Mycoviruses are common among different species of endophytic fungi of grasses. Arch. Virol. 2009, 154, 327–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nuss, D.L. Hypovirulence: mycoviruses at the fungal-plant interface. Nat. Rev. Microbiol. 2005, 3, 632–642. [Google Scholar] [CrossRef] [PubMed]
- Grente, J.; Berthelay-Sauret, S. Biological control of chestnut blight in France. In Proceedings of the American Chestnut Symposium, Morgantown, WV, USA, 4–5 January 1978; pp. 30–34. [Google Scholar]
- Sasaki, A.; Miyanishi, M.; Ozaki, K.; Onoue, M.; Yoshida, K. Molecular characterization of a partitivirus from the plant pathogenic ascomycete Rosellinia necatrix. Arch. Virol. 2005, 150, 1069–1083. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.; Zhang, L.; Li, G.; Jiang, D.; Ghabrial, S.A. Genome characterization of a debilitation-associated mitovirus infecting the phytopathogenic fungus Botrytis cinerea. Virology 2010, 406, 117–126. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Liu, H.; Zhang, M.; Cao, X.; Zhou, E. The complete genomic sequence of a novel mycovirus from Rhizoctonia solani AG-1 IA strain B275. Arch. Virol. 2013, 158, 1609–1612. [Google Scholar] [CrossRef] [PubMed]
- Ghabrial, S.A.; Castón, J.R.; Jiang, D.; Nibert, M.L.; Suzuki, N. 50-plus years of fungal viruses. Virology 2015, 479, 356–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Puchades, A.V.; Carpino, C.; Alfaro-Fernandez, A.; Font-San-Ambrosio, M.I.; Davin, S.; Guerri, J.; Rubio, L.; Galipienso, L. Detection of Southern tomato virus by molecular hybridisation. Ann. Appl. Biol. 2017, 171, 172–178. [Google Scholar] [CrossRef]
- Yu, X.; Li, B.; Fu, Y.; Jiang, D.; Ghabrial, S.A.; Li, G.; Peng, Y.; Xie, J.; Cheng, J.; Huang, J.; et al. A geminivirus-related DNA mycovirus that confers hypovirulence to a plant pathogenic fungus. Proc. Natl. Acad. Sci. USA 2010, 107, 8387–8392. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cubeta, M.A.; Vilgalys, R. Population biology of the Rhizoctonia solani complex. Phytopathology 1997, 87, 480–484. [Google Scholar] [CrossRef]
- Abdoulaye, A.H.; Cheng, J.; Jiang, D.; Xie, J. Complete genome sequence of a novel mitovirus from the phytopathogenic fungus Rhizoctonia oryzae-sativae. Arch. Virol. 2017, 162, 1409–1412. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Zhang, M.; Chen, Q.; Zhu, M.; Zhou, E. A novel mycovirus closely related to viruses in the genus Alphapartitivirus confers hypovirulence in the phytopathogenic fungus Rhizoctonia solani. Virology 2014, 456, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Castanho, B.; Butler, E.E.; Shepherd, R.J. Association of double-stranded-RNA with Rhizoctonia decline. Phytopathology 1978, 68, 1515–1519. [Google Scholar] [CrossRef]
- Bharathan, N.; Tavantzis, S.M. Genetic diversity of double-stranded RNA from Rhizoctonia solani. Phytopathology 1990, 80, 631–635. [Google Scholar] [CrossRef]
- Robinson, H.L.; Deacon, J.W. Double-stranded RNA elements in Rhizoctonia solani AG 3. Mycol. Res. 2002, 106, 12–22. [Google Scholar] [CrossRef]
- Bharathan, N.; Saso, H.; Gudipati, L.; Bharathan, S.; Whited, K.; Anthony, K. Double-stranded RNA: distribution and analysis among isolates of Rhizoctonia solani AG-2 to-13. Plant Pathol. 2005, 54, 196–203. [Google Scholar] [CrossRef]
- Zhang, M.; Zheng, L.; Liu, C.; Shu, C.W.; Zhou, E.X. Characterization of a novel dsRNA mycovirus isolated from strain A105 of Rhizoctonia solani AG-1 IA. Arch. Virol. 2018, 163, 427–430. [Google Scholar] [CrossRef] [PubMed]
- Lyu, R.L.; Zhang, Y.; Tang, Q.; Li, Y.; Cheng, J.; Fu, Y.; Chen, T.; Jiang, D.; Xie, J. Two alphapartitiviruses co-infecting a single isolate of the plant pathogenic fungus Rhizoctonia solani. Arch. Virol. 2018, 163, 515–520. [Google Scholar] [CrossRef]
- Strauss, E.E.; Lakshman, D.K.; Tavantzis, S.M. Molecular characterization of the genome of a partitivirus from the basidiomycete Rhizoctonia solani. J. General Virol. 2000, 81, 549–555. [Google Scholar] [CrossRef]
- Lakshman, D.K.; Jian, J.; Tavantzis, S.M. A double-stranded RNA element from a hypovirulent strain of Rhizoctonia solani occurs in DNA form and is genetically related to the pentafunctional AROM protein of the shikimate pathway. Proc. Natl. Acad. Sci. USA 1998, 95, 6425–6429. [Google Scholar] [CrossRef]
- Li, W.; Zhang, T.; Sun, H.; Deng, Y.; Zhang, A.; Chen, H.; Wang, K. Complete genome sequence of a novel endornavirus in the wheat sharp eyespot pathogen Rhizoctonia cerealis. Arch. Virol. 2014, 159, 1213–1216. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Falloon, R.E.; Stewart, A.; Pitman, A.R. Molecular characterisation of an endornavirus from Rhizoctonia solani AG-3PT infecting potato. Fungal Biol. 2014, 118, 924–934. [Google Scholar] [CrossRef] [PubMed]
- Zhong, J.; Chen, C.Y.; Gao, B.D. Genome sequence of a novel mycovirus of Rhizoctonia solani, a plant pathogenic fungus. Virus Genes 2015, 51, 167–170. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, P.; Zhang, L.; Xia, Z.; Qin, X.; Yang, G.; Mo, X. Molecular characterization of a novel mycovirus from Rhizoctonia fumigata AG-Ba isolate C-314 Baishi. Arch. Virol. 2015, 160, 2371–2374. [Google Scholar] [CrossRef] [PubMed]
- Morris, T.; Dodds, J. Isolation and analysis of double-stranded RNA from virus-infected plant and fungal tissue. Phytopathology 1979, 69, 854–858. [Google Scholar] [CrossRef]
- Potgieter, A.C.; Page, N.A.; Liebenberg, J.; Wright, I.M.; Landt, O.; van Dijk, A.A. Improved strategies for sequence-independent amplification and sequencing of viral dsRNA genomes. J. General Virol. 2009, 90, 1423–1432. [Google Scholar] [CrossRef] [PubMed]
- Lim, W.S.; Jeong, J.H.; Jeong, R.D.; Yoo, Y.B.; Yie, S.W.; Kim, K.H. Complete nucleotide sequence and genome organization of a dsRNA partitivirus infecting Pleurotus ostreatus. Virus Res. 2005, 108, 111–119. [Google Scholar] [CrossRef]
Figure 1.
Schematic representation of the genomic organization of RsPV5 isolated from R. solani AG-1 IA strain C24, the causal agent of rice sheath blight. (a) Schematic representation of the genomic organization of dsRNA-1. The rectangle represents open reading frame (ORF 1) and the nucleotide positions of the start and end codons are listed above the box. The gray bar represents the conserved RNA-dependent RNA polymerase (RdRp), the predicted molecular masse and the nucleotide positions of the start and termination codons are listed above the bar. The arrows under the single lines represent the length of the non-coding sequence. (b) Schematic representation of the genomic organization of dsRNA-2. The rectangle represents the open reading frame (ORF 2) and its encoded protein, capsid protein (CP), the nucleotide positions of the start and end codons are listed above the box. The arrows under the single lines represent the length of the non-coding sequences. (c) Alignments of 5′- and 3′-untranslated regions (UTRs) of RsPV5 dsRNA-1 and dsRNA-2. The letters with black shading represent conserved sequences at both ends.
Figure 1.
Schematic representation of the genomic organization of RsPV5 isolated from R. solani AG-1 IA strain C24, the causal agent of rice sheath blight. (a) Schematic representation of the genomic organization of dsRNA-1. The rectangle represents open reading frame (ORF 1) and the nucleotide positions of the start and end codons are listed above the box. The gray bar represents the conserved RNA-dependent RNA polymerase (RdRp), the predicted molecular masse and the nucleotide positions of the start and termination codons are listed above the bar. The arrows under the single lines represent the length of the non-coding sequence. (b) Schematic representation of the genomic organization of dsRNA-2. The rectangle represents the open reading frame (ORF 2) and its encoded protein, capsid protein (CP), the nucleotide positions of the start and end codons are listed above the box. The arrows under the single lines represent the length of the non-coding sequences. (c) Alignments of 5′- and 3′-untranslated regions (UTRs) of RsPV5 dsRNA-1 and dsRNA-2. The letters with black shading represent conserved sequences at both ends.

Figure 2.
Phylogenetic analysis of RsPV5. A phylogenetic tree was generated for the putative amino acid sequences of the deduced RdRp proteins using the neighbor-joining method with the program MEGA 6.0 and Bootstrap 1000 replicates. The RdRp sequences were obtained from GenBank and the accession numbers of viruses are given in the brackets behind the virus names. The scale means a genetic distance of 0.1 amino acid substitutions per site. Viral lineages are marked based on their taxonomic status.
Figure 2.
Phylogenetic analysis of RsPV5. A phylogenetic tree was generated for the putative amino acid sequences of the deduced RdRp proteins using the neighbor-joining method with the program MEGA 6.0 and Bootstrap 1000 replicates. The RdRp sequences were obtained from GenBank and the accession numbers of viruses are given in the brackets behind the virus names. The scale means a genetic distance of 0.1 amino acid substitutions per site. Viral lineages are marked based on their taxonomic status.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Liu, C.; Zeng, M.; Zhang, M.; Shu, C.; Zhou, E. Complete Nucleotide Sequence of a Partitivirus from Rhizoctonia solani AG-1 IA Strain C24. Viruses 2018, 10, 703. https://0-doi-org.brum.beds.ac.uk/10.3390/v10120703
AMA Style
Liu C, Zeng M, Zhang M, Shu C, Zhou E. Complete Nucleotide Sequence of a Partitivirus from Rhizoctonia solani AG-1 IA Strain C24. Viruses. 2018; 10(12):703. https://0-doi-org.brum.beds.ac.uk/10.3390/v10120703
Chicago/Turabian StyleLiu, Chen, Miaolin Zeng, Meiling Zhang, Canwei Shu, and Erxun Zhou. 2018. "Complete Nucleotide Sequence of a Partitivirus from Rhizoctonia solani AG-1 IA Strain C24" Viruses 10, no. 12: 703. https://0-doi-org.brum.beds.ac.uk/10.3390/v10120703
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.