Natural Variation in Resistance to Virus Infection in Dipteran Insects


1
Institute of Evolutionary Biology and Centre for Infection, Evolution and Immunity, University of Edinburgh, Edinburgh EH9 3FL UK
2
Department of Medical Microbiology, Radboud University Medical Center, Radboud Institute for Molecular Life Sciences, P.O. Box 9101, Nijmegen 6500 HB, The Netherlands
3
Radboud Center for Infectious Diseases, Radboud University Medical Center, Nijmegen 6525 GA, The Netherlands
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this manuscript.
Viruses 2018, 10(3), 118; https://0-doi-org.brum.beds.ac.uk/10.3390/v10030118
Submission received: 10 February 2018
/
Revised: 28 February 2018
/
Accepted: 8 March 2018
/
Published: 9 March 2018
(This article belongs to the Special Issue Antiviral Defense in Invertebrates)
Abstract
:The power and ease of Drosophila genetics and the medical relevance of mosquito-transmitted viruses have made dipterans important model organisms in antiviral immunology. Studies of virus–host interactions at the molecular and population levels have illuminated determinants of resistance to virus infection. Here, we review the sources and nature of variation in antiviral immunity and virus susceptibility in model dipteran insects, specifically the fruit fly Drosophila melanogaster and vector mosquitoes of the genera Aedes and Culex. We first discuss antiviral immune mechanisms and describe the virus-specificity of these responses. In the following sections, we review genetic and microbiota-dependent variation in antiviral immunity. In the final sections, we explore less well-studied sources of variation, including abiotic factors, sexual dimorphism, infection history, and endogenous viral elements. We borrow from work on other pathogen types and non-dipteran species when it parallels or complements studies in dipterans. Understanding natural variation in virus–host interactions may lead to the identification of novel restriction factors and immune mechanisms and shed light on the molecular determinants of vector competence.
1. Introduction
The vast majority of research on invertebrate antiviral immunity has used dipteran systems, including the fruit fly Drosophila and the medically important vector mosquitoes of the genera Culex, Aedes, and Anopheles. These fields have complemented each other, with interest in mosquito-vectored viruses guiding studies in Drosophila, and the well-annotated genome and genetic tools of Drosophila providing a model to guide laboratory experiments in the less tractable mosquito. The comparison of antiviral processes in these dipterans highlights a conserved yet dynamic immune system, with some antiviral mechanisms conserved from flies to mammals, whereas others have evolved since the expansion of dipterans.
Studies from both fruit flies and mosquitoes have uncovered variation in host–virus interactions. These studies have focused primarily on differences in immune mechanisms between virus species within a host species, or on variation in resistance to a particular virus within or between populations of host species. Insight into virus-specific mechanisms has been gleaned through the use of a diverse set of natural and non-natural viruses in Drosophila, and through comparisons of vectored flaviviruses, rhabdoviruses, alphaviruses, and bunyaviruses—part of a larger group referred to as arthropod-borne viruses (arboviruses). These studies reveal a general requirement for antiviral RNAi in dipterans, virus-specific contributions of canonical antimicrobial pathways (such as Toll, Imd, or JAK-STAT), a diverse transcriptional response, and emerging antiviral processes that have yet to be studied in a comparative context (reviewed in [1,2,3,4,5,6,7]). In addition, there is tremendous variation in virus susceptibility between hosts, where the resistance of an individual host is dependent on factors such as genotype, microbiota, infection history, mating status, and the environment. Variation in resistance is further complicated in mosquitoes, where the phenotype of interest is vector competence, the ability of a mosquito to transmit an arbovirus after it had crossed multiple infection barriers (midgut infection barrier (MIB), midgut escape barrier (MEB), salivary gland infection barrier, and salivary gland escape barrier) (reviewed in [8]).
Viral genetic variation, both within and between hosts, is likely to contribute to infection outcomes [9,10]. Arbovirus genetic variation is dynamic within a single host, and is shaped by natural selection and population bottlenecks and expansions at each infection barrier [11,12,13,14]. In some cases, viral variants affecting vector competence have spread through a population, leading to outbreaks of arboviral disease in these regions. For example, single amino acid substitutions in Venezuelan equine encephalitis virus (VEEV) and Chikungunya virus (CHIKV) envelope proteins are associated with increased infectivity in Ochlerotatus taeniorhynchus and Ae. albopictus, respectively [15,16,17,18]. This aspect of pathogen-associated natural variation is beyond the scope of this review and is discussed in detail in [12,19].
The nature of variation in virus–host interactions is of great medical and evolutionary importance. Discerning general immune mechanisms and identifying determinants behind resistant individuals can inform the development of broadly acting intervention strategies to reduce arbovirus transmission and arboviral disease. Naturally segregating polymorphisms associated with resistance to viral infection can provide mechanistic insights into the control of viral infection, and identify loci likely under pathogen-mediated selection. These loci sometimes exhibit signatures of adaptive evolution or balancing selection, giving credence to the hypothesis of antagonistic coevolution between viruses and host immune genes. In this review, we provide a general description of antiviral immune mechanisms, and their impact across virus species studied in dipterans. We cover the nature and sources of variation in resistance among host individuals, particularly focusing on population studies of resistance polymorphisms and microbiota. For a list of viruses discussed in this review, see Table 1.
2. Antiviral Immune Processes
Numerous conserved signaling pathways and cellular processes mediate antiviral immunity in dipterans (Figure 1) (reviewed in [1,2,4,5,6]). A subset of these pathways is generally antiviral (e.g., RNA interference), whereas others provide antiviral defense against only specific viruses (e.g., autophagy). In some cases, variation in viral resistance or tolerance can be mapped directly to genetic variation within these pathways. Although our focus is on dipteran immunity, we note that most of these processes are widely conserved, from flies to mammals. In this section, we briefly review known antiviral immune mechanisms in dipterans, which we will later discuss in a comparative context in Section 3.
2.1. RNA Interference
RNA silencing pathways utilizes short RNA sequences bound to an Argonaute-family protein to transcriptionally or post-transcriptionally silence complementary “target” sequences. These include the microRNA (miRNA) pathway, which primarily regulates endogenous gene expression, the PIWI-interacting RNA (piRNA) pathway, which regulates transposons, and the small interfering RNA pathway (siRNA), which serves as one of the most important antiviral defenses in invertebrates. Briefly, Dicer-2 (Dcr-2), an RNase III family endonuclease, recognizes viral-derived dsRNA and cuts it into 21 nucleotide siRNA duplexes. A single strand of the siRNA duplex is loaded into the Argonaute-2 (Ago2)-containing RNAi Induced Silencing Complex (RISC), which then slices any viral sequence that is complementary to the loaded siRNA, thereby controlling virus infection. In Drosophila, the siRNA pathway provides broad protection against both RNA and DNA viruses [20,21]. The siRNA pathway has been shown to be antiviral against arboviruses in relevant vector mosquitoes as well [22]. Recently, “secondary siRNAs” have been described, which are produced from reverse-transcribed viral DNA circles. These secondary siRNAs are excreted by hemocytes in exosomes and are proposed to mediate a non-cell autonomous, systemic RNAi-based immune response [23,24,25] (further discussed in Section 6.3).
In addition to viral siRNAs, Aedes mosquito species produce virus-derived piRNAs. This pathway is crucial for transposon suppression in the germline and does not have an antiviral role in Drosophila [26]. However, the PIWI genes have undergone expansion in mosquitoes and generate virus-derived piRNAs in the mosquito soma [27,28,29]. Depletion of viral piRNAs in mosquito cells by knocking down the associated proteins (Ago3 and Piwi5) seemed to have no effect on viral replication [30]. However, silencing Ago3 in An. gambiae mosquitoes led to increased O’nyong-nyong virus (ONNV) replication [31]. In addition, another piRNA pathway component, Piwi4 has been shown to be antiviral against Semliki forest virus (SFV) and Zika virus [32,33].
2.2. Other RNA Processes
In addition to RNAi, other RNA-based cellular processes have been implicated in antiviral defense. The RNA decay pathway protects against defective cytoplasmic RNAs, including those without 5′ caps or polyadenylated tails, lacking stop codons (nonstop decay), encoding early stop codons (nonsense-mediated decay), or stalled in ribosomes (no go decay) (reviewed in [34]). Offending RNA molecules are deadenylated and either degraded 3′ to 5′ by the RNA exosome, or decapped and degraded by the 5′ to 3′ exonuclease Xrn1 (reviewed in [35]). Viral RNA may include a number of hallmarks of aberrant cellular transcripts, including 5′ triphosphate groups, limited poly-A tails, or interior stop codons, making them susceptible to the RNA decay machinery [36,37]. Consistently, in Drosophila, the decapping enzymes, DCP1 and DCP2, and the RNA exosome have antiviral activity, and RISC-mediated silencing is partially dependent on Xrn1 and the exosome [38,39,40,41]. Additionally, there is evidence of viral modulation of processing body (P body) and stress granule formation. For example, Cricket paralysis virus (CrPV) infection leads to dispersal of P body components, which are sites that may be associated with organized decapping and 5′ to 3′ decay [42]. In mosquitoes, evidence for antiviral RNA decay comes from the flavivirus noncoding RNA, sfRNA, which is produced by stalling of Xrn1 on structured RNA elements in the 3′ UTR [43].
RNA editing by adenosine deaminases that act on RNA (ADAR) occurs during the mammalian innate immune response, where the replacement of adenosines with inosines reduces the stability of dsRNA and the accuracy of replication and translation, due to inosines being read as guanosines (reviewed in [44]). Evidence in support of an antiviral activity of ADAR in insects comes mainly from mutation and substitution patterns in a subset of RNA viruses. Two sequenced strains of sigma virus (DMelSV) appeared to have been hypermutated from A to G [45], and analysis of DMelSV variation within flies and across populations found that ADAR-resistant sites are less likely to be polymorphic [46]. Similar patterns are also observed in Zika virus, although it is unknown whether this is insect-mediated [47].
2.3. Nuclear Factor κB Pathways
There are two Nuclear factor κB (NF-κB) signaling cascades in flies: the Toll and immune deficiency (IMD) pathways (reviewed in [48,49]). Generally, these pathways are activated when an upstream pattern recognition receptor interacts with a pathogen-associated molecular pattern. This leads to phosphorylation of IκB (inhibitor of κB; encoded by cactus for the Toll signaling and by the C-terminal domain of Relish (Rel2 in mosquitoes) in IMD signaling) by an IκB kinase (IKK, encoded by pelle in Toll signaling and kenny in IMD signaling). This results in degradation of IκB and subsequent release of the NF-κB transcription factor (encoded by dorsal (Rel1 in mosquitoes) or Dorsal-related immune factor for Toll signaling and Relish for IMD signaling). These transcription factors then translocate into the nucleus to induce transcription of immune effectors, including antimicrobial peptides [48,49]. Activation of IMD may also signal through the JNK pathway [50,51]. Ostensibly, the primary functions of these pathways are in anti-bacterial and anti-fungal defense, although both have been implicated in defense against various dipteran viruses [21,52,53,54,55,56,57]. In agreement with an antiviral function, some insect viruses encode suppressors of the NF-κB pathway [58,59].
The antiviral effectors downstream of NF-κB are, for the most part, unknown, besides a couple of examples. The NF-κB-responsive antimicrobial peptides are slightly upregulated after viral challenge, and some seem to have antiviral properties [60,61]. However, the antiviral activity of the Toll and IMD pathways may be based on mechanisms independent from antimicrobial peptides. For example, Toll signaling seems to be involved in differentiation of hemocytes, phagocytic cells in the hemolymph of invertebrates (see Section 2.6 and Section 3.3 for the role of phagocytosis in immunity) [62]. IMD signaling can be pro-apoptotic, which itself can have antiviral functions [63,64], and is involved in Pvf2 upregulation which activates antiviral ERK signaling [65,66].
2.4. JAK-STAT Pathway
In insects, the JAK-STAT pathway is activated upon binding of one of the unpaired ligands (upd, upd2, or upd3) to the pathway receptor, domeless. This interaction results in activation of the JAK kinase hopscotch, which then phosphorylates Stat92E, resulting in its dimerization and translocation into the nucleus where it induces transcription of JAK-STAT dependent genes (reviewed in [67]). The antiviral gene Vago, which is induced downstream of dsRNA recognition by Dcr-2, may also activate JAK-STAT signaling in mosquitoes, although possibly through a different receptor [21,68,69,70]. During viral infection, Vago and the upd ligands are upregulated, as well as a subset of known STAT-regulated genes [71,72]. This activation ostensibly results in an antiviral transcriptional program, and altering pathway activity in infected flies and mosquitoes affects resistance to some viruses [21,69,70,71,73]. Similar to antiviral NF-κB signaling, there are virus-specific differences in STAT-responsive transcriptional output, and STAT-responsive antiviral effectors are still mostly unknown. However, attC is upregulated downstream of JAK-STAT following Sindbis virus (SINV) infection in Drosophila, and heterozygous attC mutations lead to increased viral replication [61]. Additionally, two JAK-STAT pathway candidate effectors have been identified in Dengue virus (DENV)-infected Ae. aegypti. These DENV restriction factors (DVRF) include DVRF1 (CG15168 in D. melanogaster), a putative transmembrane protein which could serve as a receptor, and DVRF2 (CG8541 in D. melanogaster), a predicted secreted protein with anti-freezing and allergen domains [73]. Neither of the Drosophila DVRF homologs have been implicated in antiviral immunity.
2.5. Nutrient Signaling
Broadly speaking, diet and nutrient availability are intricately linked to immune variation and function (reviewed in [74,75]). Known diet-responsive signaling pathways have been implicated in antiviral immune modulation in dipterans, including ERK and PI3K/Akt signaling. Both ERK and PI3K/Akt are downstream of insulin receptor signaling, and PI3K/Akt has crosstalk with the amino acid-sensing TOR signaling pathway [65,76]. The ERK pathway can restrict viral infection in Drosophila midguts and mosquito cells [65,66]. In addition, downregulation of PI3K/Akt signaling can enhance antiviral defense, at least partially through releasing its inhibition of antiviral autophagy [77,78,79]. Akt also inhibits FOXO, a virus-induced transcription factor downstream of Toll, which upregulates antimicrobial peptides and possibly RNAi pathway components [80,81,82]. Notably, PI3K/Akt activity is proviral independent of its role in autophagy regulation, and is activated by SINV to promote cap-dependent translation [83].
2.6. Apoptosis, Phagocytosis, and Autophagy
Apoptosis, phagocytosis, and autophagy make important contributions to dipteran antiviral defense. Apoptosis is long known to be an important host defense response in vertebrates (reviewed in [84]). In Drosophila, viral infection induces p53-mediated transcription of RHG genes (reaper, hid, grim, and sickle), which promote degradation of Drosophila inhibitor of apoptosis 1 (DIAP1) and consequent activation of the initiator caspase Dronc [64,85]. This response occurs rapidly after infection, effectively reducing the duration a virus can access host factors that are crucial for replication [64]. Additionally, an N-terminal degron (a protein domain that regulates protein turnover rate) renders DIAP1 inherently unstable [86], allowing the promotion of apoptosis during virus-mediated translational inhibition [87]. Phosphatidylserine on apoptotic cells is recognized by the engulfment receptors draper and Integrin βν on phagocytes, which eliminate infected cells [88]. Individual viral particles, including Drosophila C Virus (DCV) and White spot syndrome virus (a natural shrimp nimavirus) may also be direct targets of phagocytosis in cell culture [89,90].
Autophagy, a process in which intracellular particles are enveloped by membrane crescents and shuttled to lysosomes for degradation, is also antiviral in some contexts [77,79,91]. For example, noncanonical Toll-7 signaling may be responsible for activating antiviral autophagy during rhabdovirus and bunyavirus infection, although autophagy is dispensable against most other viruses [56,78,79].
3. Virus-Specific Responses
Virus–host interactions include both general and virus-specific components. For example, a genome-wide RNAi screen of West Nile virus (WNV) uncovered 50 restriction factors, over half of which were antiviral across flaviviruses, whereas only seven were broadly antiviral across disparate viral families [92]. This indicates that hosts have evolved antiviral mechanisms in response to virus-specific infection cues (e.g., pathogen associated molecular patterns, pathogenesis, or affected tissues), and/or that viruses are able to evade a subset of the host immune responses. Comparative antiviral immunity experiments have primarily focused on RNAi, induced immune pathways and their downstream transcriptional responses (i.e., NF-κB and JAK-STAT pathways), and cellular responses. These studies have used a collection of native and non-native viruses (Table 1) and shown considerable variation in the efficacy of cellular and humoral immunity across viruses, concomitant with highly virus-specific transcriptional responses to infection [6]. Notably, there has been recent interest in identifying and isolating native dipteran viruses, focusing both on viruses from diverse families and those with nearby relatives of medical importance, representing a clear potential for dipterans to become a powerful in vivo comparative system to study the diversity of antiviral immune responses [93,94,95,96].
3.1. RNA Interference
The RNA interference pathway is often referred to as being the most general and important innate antiviral immune pathway in insects. Indeed, RNAi is demonstrably antiviral against a spectrum of RNA and DNA viruses across insects [3,22]. The broad importance of antiviral RNAi is underlined by the prevalence of viral suppressors of RNAi (VSRs), often found in insect-infecting viruses, which inhibit processing of viral dsRNA by RNAi enzymes [97,98,99,100,101,102]. These VSRs may vary in potency, host-specificity, and mechanism, although commonly act by binding either short or long dsRNA to shield it from Dcr-2 or Ago2 processing [102,103]. In some cases, a VSR may block RNAi function so completely that removing key RNAi components appears not to have an effect on virus replication, and thus variation in VSR efficacy could lead to observed virus-specific variation in the requirement of antiviral RNAi. For example, Nora virus VP1 encodes an Ago2 suppressor, implying that RNAi poses an antiviral threat to Nora virus [20]. However, Nora virus titers remain unchanged in Dcr-2, r2d2, and Ago2 mutants during persistent infection [104].
VSR potency may also be reflected in length distributions of virus-derived small RNAs. Canonically, virus-derived small RNAs are overwhelmingly 21 nucleotides in length, because Dcr-2 cleaves at 21 base intervals. However, a metagenomic survey of viruses in D. melanogaster found variation in virus-mapping small RNA length distributions [94]. Four D. melanogaster-infecting picorna-like viruses (DCV, Nora virus, Kilifi virus, and Thika virus) have broad length distributions, with similar amounts of 22–29 nucleotide viral-derived small RNAs as 21 nucleotide species. Because two of the four viruses with broad siRNA length distributions are known to suppress RNAi, a viable hypothesis is that especially potent VSRs may reduce the number of Dcr-2-generated siRNAs, although increased viral degradation products and the concomitant increase in 22–29 nucleotide species could also explain the pattern [94]. Regardless, the prevalence of VSRs underlines the importance of considering virus-mediated subversion of host immunity when studying virus-specific responses.
3.2. Induced Immune Responses
Viruses trigger a rapid transcriptional response in infected flies [105]. This depends on recognition of virus-associated molecular patterns, and on other infection-derived cues such as host manipulation or damage (reviewed in [4]). As in mammals, NF-κB and JAK-STAT pathways help coordinate these inducible responses, and a subset of upstream signaling pathway regulators are often differentially expressed during infection with different viruses [21,71,72,106,107]. In Drosophila, the Toll and Imd pathways are antiviral against a broad panel of RNA viruses. Toll signaling is antiviral upon oral, but not systemic, viral infection, and mutants are less resistant to DCV, CrPV, Nora virus, Drosophila X virus (DXV), and Flock House virus (FHV) [52,57]. Likewise, IMD has been proposed to be antiviral against DCV, SINV, and CrPV [54,55,66]. The antiviral role of NF-κB pathways in mosquitoes is not as apparently widespread. The Toll pathway was found to have an antiviral role only against the flavivirus DENV in Ae. aegypti mosquitoes [53,108], whereas no role of this pathway was observed for the alphaviruses SFV and ONNV [109,110]. On the other hand, the IMD pathway was found to be effective against SFV and ONNV when stimulated prior to infection in cell culture [109,111]. Even though some IMD pathway components and effector genes were found to be upregulated during DENV infection, transient activation of the pathway did not affect viral titers [53].
The antiviral role of JAK-STAT signaling has also been investigated across Drosophila viruses, where it is crucial in CrPV and DCV infections, but exerts a minimal effect on FHV, SINV and vesicular stomatitis virus (VSV) [21,71]. However, JAK-STAT controls replication of a SINV replicon, indicating there may be JAK-STAT-dependent responses specific to tissues or the route of infection [54,112]. In mosquitoes, the antiviral effect of the JAK-STAT pathway has been seen in DENV-infected Ae. aegypti mosquitoes, where silencing components of the pathway resulted in higher viral titers and activating the pathway had the opposite effect [70,73]. Similar to the IMD pathway, stimulating the JAK-STAT pathway prior to infection had an antiviral effect on SFV [109]. However, the antiviral role of this pathway in other virus–vector combinations remains unclear, where no upregulation of JAK-STAT pathway components was observed upon infection (reviewed in [113]).
The resulting downstream transcriptional response across viruses is remarkably variable [4,21]. Comparison of the differentially expressed genes following infection with DCV, SINV, or FHV resulted in the identification of 601 genes, only 42 of which are shared between the three viruses [21]. An exemplar of this trend is an analysis of JAK-STAT responsive genes vir-1 and TotM, which show an almost mutually exclusive expression pattern in response to a panel of viruses, whereby vir-1 is strongly induced in response to CrPV, DCV, and FHV, while TotM is induced after SINV, VSV, or DXV infection [21]. Neither gene is strongly upregulated in response to the DNA viruses IIV6 or Kallithea virus [21,107]. Likewise, the heatshock pathway is induced in vivo in response to DCV and CrPV, but not IIV6 [72]. Diedel, a negative regulator of Imd signaling, also has highly virus-specific regulation, and is induced over 100-fold during SINV and VSV infection, but less than 5-fold in response to CrPV, DCV, and FHV [59]. However, there may be responses common to a particular virus family or even between viruses of different families. A comparative transcriptomic analysis of infections by the flaviviruses DENV, Yellow fever virus (YFV) and WNV infection in Ae. aegypti mosquitoes found 35 genes that were commonly differentially expressed, suggesting a transcriptomic signature unique to flaviviruses. Antimicrobial effectors of the JAK-STAT pathway (four Cecropin A-like genes and one defensin gene) as well as components of the Toll pathway were found to be downregulated by all three viruses [114]. A more recent study compared transcriptional responses between flaviviruses, alphaviruses and bunyaviruses upon infection of Ae. aegypti mosquitoes. Only 19 genes were found to be upregulated by all tested viruses and no commonly downregulated genes were found. Among the 19, the gene responsible for GABA signaling was found to be connected to blood feeding and responsible for enhancing virus replication [115]. Together, these studies indicate the transcriptional response to a particular virus is unique, and may be the amalgamation of host adaptation to a specific virus, virus-mediated damage and immune subversion, cellular and tissue tropism, and virus replication kinetics.
3.3. Autophagy, Phagocytosis, and Apoptosis
A recent study using Drosophila has formally compared the requirement of phagocytosis, apoptosis, and autophagy using a panel of six viruses (DCV, CrPV, FHV, VSV, SINV, and IIV6) [79]. Interference with phagocytosis by latex bead injection or genetic ablation of hemocytes led to increased susceptibility to CrPV, FHV, and VSV, but not to IIV6, DCV, or SINV [79]. Antiviral phagocytosis likely operates through different mechanisms. CrPV and FHV induce apoptosis, and phagocytes were recruited to dying cells to remove them [79,116]. However, VSV did not induce apoptosis, indicating the effect from phagocytes may be through direct clearance of viral particles, or, more speculatively, via the recently described secondary siRNA pathway [25] (described in more detail in Section 6.3). Additionally, DCV infection induced apoptosis and subsequent phagocytic clearance, although phagocytosis-deficient flies were not more susceptible to infection [79,88]. Finally, this study found that autophagy-deficient flies were more susceptible to VSV [77], but not to DCV, CrPV, FHV, and IIV6 infections [79]. Additionally, autophagy appears to be proviral in FHV infection, where flies mutant for Atg7, a gene required for autophagosome formation, had lower FHV titer and less virus-induced mortality [79].
4. Genetic Variation in Antiviral Immunity in Dipterans
The evolutionary relationship between host and pathogens is often referred to as antagonistic coevolution, because an increase in the fitness of one often corresponds to a decrease in the fitness of the other. Framing host–parasite interactions in an evolutionary context is critical, as variation in epidemiological and population genetic parameters (e.g., host costs of resistance, demographic population structure, and pathogen virulence and transmissibility) is expected to result in different types of selection, which in turn affects the architecture of genetic variation in resistance [117]. Even with relatively simple genetic architectures, where resistance and susceptibility are controlled by few host and parasite genotypes at single loci, antagonistic coevolution can result in a resistant allele being maintained at a stable frequency, changing in frequency chaotically or cyclically, or fixing in a population (e.g., [117,118,119]). In reality, host–parasite interactions can be immensely complex [120], and most of the loci under pathogen-mediated selection are unknown. Identification of these loci is of great medical and evolutionary importance, and efforts have been made to characterize genetic variation in host resistance, and to determine the effects of parasite mediated selection on host gene evolution.
4.1. Segregating Genetic Variants Associated with Viral Resistance in Drosophila
In some cases, genetic variation has been mapped to discrete loci in the host genome. In D. melanogaster, the most extensively studied host–virus system at the population level is DmelSV, a negative-sense ssRNA rhabdovirus (reviewed in [121]). DmelSV is transmitted vertically to offspring through eggs and sperm, although some fly strains are not permissive to DmelSV replication [122]. This resistance was first mapped to a complex amino acid substitution in the N-terminal PB1 domain of ref(2)P (Table 2), a gene now known to be involved in autophagy and Toll signaling [122,123,124,125,126]. Viral replication and transmission is reduced in homozygous ref(2)P mutants or trans-heterozygotes bearing a mutant ref(2)P and the refractory allele, indicating that ref(2)P is a proviral host factor in DmelSV infection [127].
More recently, two additional loci have been mapped with segregating variants associated with resistance to DmelSV. The first is a triallelic polymorphism at the CHKov1 and CHKov2 paralogs [131], where an insertion of a transposable element (Doc element) in CHKov1 provides resistance to DmelSV, and a following complex (and rare) rearrangement is associated with even greater DmelSV resistance [131] (Table 2). The Doc insertion also confers resistance to pesticides [139]. Although the functional relevance of the CHKov genes during viral infection remains untested, the resistance mutations could exert an effect on viral entry, as CHKov is a predicted acetylcholine esterase, and Rabies virus (also a rhabdovirus) uses the acetylcholine receptor as a cell entry point [140]. In addition to ref(2)P and CHKov, a large deletion in the serine-rich linker region of Ge-1 results in a 10-fold reduction in viral titer and lower infection rates (Table 2) [40,128]. Ge-1 is a conserved adaptor bridge between Decapping protein 1 and Decapping protein 2 that helps localize these enzymes to processing bodies [141,142], and thus could plausibly exert its antiviral effect through the RNA decay pathway [40]. Finally, up to 7 QTLs associated with DmelSV replication or transmission have been identified in genetic mapping experiments, however the exact loci responsible have yet to be reported [128,132,143,144].
Natural variation has also been mapped for two other Drosophila viruses: DCV and Kallithea virus. Resistance to DCV has been mapped to complex polymorphisms at the pastrel (pst) locus [130,133]. The primary pst resistance mutation is due to a single amino acid change, although there are multiple structural alleles and cis-regulatory changes that may enhance resistance, resulting in seven alleles with four distinct phenotypes [134]. Although the function of pst is unknown, overexpression of the susceptible allele provides protection against DCV, and pst is upregulated after intra-abdominal viral injection, indicating that pst is an induced antiviral factor [134]. In addition to pst, polymorphism in Anaphase promoting complex 7, Ubiquitin conjugating enzyme E2H, and 2 QTLs may also underlie genetic variation in DCV resistance, although these await more extensive characterization [130,133].
A genome-wide association study has found multiple loci that are associated with either viral titer or mortality following Kallithea virus infection, a dsDNA nudivirus of D. melanogaster [94,107]. The most confident association was a nonsynonymous polymorphism affecting a subset of splice variants in Cdc42-interacting protein 4 (Cip4), a gene involved in membrane trafficking [107,145]. Many other loci were found significantly associated to Kallithea virus resistance (>50 genes), in stark contrast to the above RNA viruses, where variation in resistance is explained by few large effect loci. This could reflect general differences in coevolution between hosts and their DNA and RNA viruses, such as the greater complexity (Kallithea has approximately 100 genes) and reduced substitution rate of DNA viruses [146].
4.2. Evolution of Resistance Loci
The genetic architecture of resistance to viruses in Drosophila often seems to include large-effect polymorphisms at an intermediate frequency in populations (Table 2). These variants are likely under pathogen-mediated selection, and surrounding patterns of polymorphism and divergence have been compared to expected patterns of balancing selection (i.e., selection that maintains genetic variation) or recurrent positive selection (i.e., selection on an advantageous allele), which would be compatible with their involvement in antagonistic coevolution (reviewed in [117,147,148]). Although none of the identified resistance loci display significantly increased diversity that is a hallmark of balancing selection, ref(2)P and pastrel have relatively high levels of nonsynonymous or structural polymorphism [123,130,134,149]. This may be due, in part, to incomplete selective sweeps, whereby a resistance mutation rises to high frequency and subsequently loses the selective benefit through viral counter-adaptation. This was witnessed in the 1980s, when a DmelSV strain that regained transmission advantage in flies with a resistant ref(2)P allele swept through the population [150,151,152,153,154]. Additionally, some dipteran immune genes exhibit unusually high diversity. NF-κB-responsive AMPs appear to be under balancing selection and show high levels of nonsynonymous polymorphism, with some convergently maintained in different species, likely due to their role in anti-bacterial or anti-fungal immunity [155,156]. The N-terminal glutamine-rich repeat region of Ago2 is also hypervariable either due to diversifying selection or high mutation rates and low constraint [157,158].
In addition to maintaining polymorphism, pathogen-mediated arms races may be expected to fix adaptive mutations. This can occur quickly on an evolutionary timescale, and these beneficial mutations will only be briefly visible as variation within populations. However, they can be recognized as elevated divergence between populations or species. This is evident in genes with segregating resistance polymorphism, and ref(2)P, CHKov1/2, and Ge-1 show signs of recent or recurrent positive selection [40,131,153]. These patterns are also apparent in some immune genes, and are most striking in the RNAi pathway, which evolves rapidly due to adaptive evolution, particularly in genes mediating defense against transposons and viruses, including antiviral effectors Dcr-2 and Ago2 [159,160,161]. In addition, RNAi genes are more likely to show diversity patterns consistent with positive selection driving new mutations to fixation [161,162,163,164]. Although less likely to be virus-mediated selection, genes in the Toll and IMD signaling pathways may also have elevated levels of positive selection, with the most convincing evidence in Relish and its interactors [165,166,167]. Additionally, genes encoding pathogen recognition proteins appear to evolve rapidly, especially those involved in phagocytosis [166,168].
4.3. Genetic Variation Associated with Viral Resistance in Mosquitoes
Although the genomic tools of mosquitoes are not as advanced as those for Drosophila, the sequencing of the genome of Ae. aegypti (the principal vector of Dengue, Chikungunya and Zika viruses) [169] has enabled the investigation of the precise genetic determinants of vector competence. Prior to the genomics age, Ae. aegypti strains with differences in midgut and disseminated DENV infections were identified [170,171] and used to map alleles associated with susceptible or refractory phenotypes. Quantitative-trait loci (QTL) mapping allowed the determination of several loci that act additively to determine midgut infection and dissemination [137,171,172]. Two QTLs were identified that significantly associated with DENV-2 midgut infection [137]. One of these contained early trypsin, a female-specific gene induced shortly after blood-feeding, and required for proteolytic digestion of the bloodmeal [173]. A subsequent fine-scale mapping study of the segregating sites in the early trypsin coding sequence in four Mexican populations was unable to find a causal variant, indicating that the locus may be linked to early trypsin in the original QTL, or that polymorphic cis-acting elements are causing differences in early trypsin expression [174]. Regardless, there is evidence that trypsins are important regulators of DENV infection, framing this QTL in an interesting context. For example, addition of a soybean trypsin inhibitor to an infectious blood meal significantly reduced DENV midgut infection and dissemination [175]. In contrast, another group showed that adding soybean trypsin inhibitor as well as RNAi knock-down of the late trypsin gene 5G1 led to higher DENV infectivity of Ae. aegypti mosquitoes [176]. An expressed sequence tag (EST) with high homology to a trypsin inhibitor was found in refractory Ae. aegypti populations, implying further that midgut proteolytic activity could limit DENV infection [177].
Another QTL mapping study found four genomic regions associated with variation in body size that may additionally affect vector competence of Ochlerotatus triseriatus mosquitoes for La Crosse encephalitis virus (LACV) [178]. This virus belonging to the California serogroup of bunyaviruses is actively maintained by transovarial transmission from infected females to their progeny in the intervening periods between epizootic outbreaks, during which the virus maintains low level replication that is not detrimental to the overwintering embryos [179]. Three genetic loci were found to additively determine transovarial transmission rates in Ocherotatus triseriatus mosquitoes for LACV (Table 2) [138].
Genetic differentiation among various Ae. aegypti strains from Vietnam and Thailand, possibly caused by insecticide use, were also found to be associated with vector competence for DENV [180,181]. Lambrechts and colleagues determined that there is an active interaction and ongoing local adaptation between vector and virus genotypes, resulting in genotype-by-genotype (GxG) interactions that affect vector competence phenotypes [9]. Population genetic studies to map the mosquito loci underlying these GxG interactions identified QTLs that conferred resistance to several DENV viral strains and others where a specific QTL varied based on the DENV isolate or serotype used [136]. Further fine-scale mapping of these GxG interactions revealed that polymorphisms in the Dcr-2 locus correlated with DENV susceptibility in an isolate specific manner. Thus, variation in dsRNA binding or cleavage by Dcr-2, coupled with variation in the relative importance of antiviral RNAi across DENV strains could explain the observed G×G interactions, although the precise mechanism remains to be characterized [135]. As in Drosophila, RNAi genes in Aedes may have higher rates of protein evolution [182]. Mosquitoes do not have an overtly antagonistic relationship with the arboviruses they transmit, leading to the speculation that it is not arboviruses, but insect-specific viruses [183] or transposon movement that could drive such evolution.
Lately, transcriptomic differences between refractory and susceptible vector populations have been used to identify causal determinants of variation in vector competence, with most studies focusing on Ae. aegypti and DENV. Behura et al. identified different gene networks being activated in susceptible versus refractory populations [184]. In particular, apoptosis-related genes were expressed at higher levels in refractory populations, which may be a defense response that impedes a productive DENV infection. For example, a mosquito ortholog of the Drosophila pro-apoptotic gene reaper, termed michelob_x, as well as other related genes, were upregulated upon DENV infection only in a refractory Ae. aegypti strain [64,177]. In agreement, knock-down of the initiator apoptotic caspase AeDronc in a refractory Ae. aegypti strain increased its permissiveness to DENV infection, whereas suppressing the caspase inhibitor AeIAP1 in susceptible strains made them refractory [185]. Differential regulation of apoptosis in mosquitoes with a refractory phenotype has been documented in other vector-virus combinations as well [5,186]. For example, WNV infection showed apoptotic effects in the midguts of a refractory population of Cx. pipiens mosquitoes, and clearance of WNV infection in the salivary glands over time correlated with levels of apoptosis in Cx. quinquefasciatus [187,188].
In addition to apoptosis-related factors, expression of furin-like genes were higher in the susceptible strains, suggesting that they may aid in DENV maturation [184]. Expression of many cuticle protein genes were comparatively lower in the susceptible population, suggesting that they may possess thinner anatomical barriers that enhance midgut infection and escape [184]. A putative 67 kDa DENV receptor protein (R67/R64) on Ae. aegypti midgut epithelial cells was found to be expressed at significantly higher levels in the midguts of strains susceptible to DENV infection as compared to refractory ones [189]. Interestingly, a glucosyl/glucuronosyl transferase (AAEL003099) was found to be downregulated in Ae. aegypti populations refractory to DENV [190] and the same gene was found to be upregulated by a Talaromyces species of fungus that increases the mosquito’s susceptibility to DENV [191]. These studies suggest that the relative expression levels of these genes in mosquito populations could be a source of natural variation that influences vector competence.
Several genes belonging to innate immune pathways (Toll, JAK/STAT, IMD) were selectively upregulated in refractory Ae. aegypti strains, suggesting that the basal expression level of immune response genes may modulate susceptibility to DENV infection [190]. The same study also highlighted the importance of differential vATPase subunit expression in influencing vector competence. These multi-subunit enzymes drive proton transport to acidify organelles like endosomes and lysosomes [192] and is required during a key step in the DENV life cycle, low pH-mediated viral and cellular membrane fusion [193]. Indeed, RNAi-mediated knock-down of the vATPase G subunit gene (AAEL012819) rendered susceptible strains more refractory to DENV [190].
5. Microbe-Dependent Variation in Dipteran Antiviral Immunity
In addition to genetic determinants in the host genome, the fly microbial community may influence resistance and tolerance to some viruses, where inter-strain variation in antiviral protection can manifest itself as variation among fly individuals. The effect of Wolbachia infection has been the primary focus of these studies, although the importance of the gut microbiome is beginning to be understood.
5.1. Wolbachia in Drosophila
Wolbachia is a α-proteobacterium widespread among insects that infects reproductive tissues and induces a diverse array of phenotypes across insect hosts, including male-killing, male feminization, cytoplasmic incompatibility, and increased resistance to RNA viruses [194,195,196]. The exact mechanism behind Wolbachia-mediated antiviral protection has remained elusive. Wolbachia titer is generally correlated with the magnitude of antiviral protection [197,198,199,200], which has been explained by competition between viruses and Wolbachia for resources or by Wolbachia-mediated immune priming [201]. A cholesterol-rich diet ameliorates Wolbachia pathogen interference against DCV in Drosophila, but altering nutrient availability has no effect on DENV in mosquitoes [202,203]. Although Ago2, reactive oxygen species, certain AMPs, and Mt2 (a methyltransferase with antiviral or proviral activity in different contexts) are differentially regulated during Wolbachia infection [204,205,206,207,208], the effects are small, or species specific, making it unlikely that immune priming is the sole mechanism of viral interference [207,209,210,211,212]. Regardless of mechanism, different Wolbachia strains have different propensities toward pathogen interference [200]. Natural variation in antiviral protection of Wolbachia has been investigated in D. melanogaster and D. simulans, where Wolbachia infection is generally protective against RNA (but not DNA) viruses [195,199].
Three genotypes of the single Wolbachia pipientis strain that infects D. melanogaster have been extensively characterized for antiviral protection. The most prevalent genotype in the wild, wMel, provides limited antiviral protection, whereas the other two, wMelCS and wMelPop, provide intermediate and strong antiviral protection, respectively [213]. Antiviral protection in these strains is positively correlated with Wolbachia titer and virulence, with wMelPop causing considerable mortality in infected flies [213,214]. The extreme phenotypes of wMelPop have been mapped to a copy number variant encompassing eight Wolbachia genes, called Octomom [213,215,216]. Octomom copy number varies within and across flies infected with wMelPop, where higher copy number wMelPop genotypes replicate more rapidly, have a higher virulence, and provide greater protection against RNA viruses [213,216,217]. Wolbachia-mediated antiviral protection is likely more complicated than this single locus, as wMelCS and wMel3562 (another highly virulent Wolbachia strain) have single or low Octomom copy numbers [213,218], and Wolbachia with Octomom deletions retains its pathogenicity (although this genotype has not been assayed for pathogen interference) [215,219]. Notably, the wMelPop strain was recovered from laboratory flies, and has not been found in the wild [213,214].
Many Wolbachia strains are able to subvert host reproduction, benefiting infected females and driving the infection through a population [220]. Because antiviral protection and reproductive subversion appears to be uncorrelated [221], this could lead to waves of viral resistance and susceptibility in fly populations. For example, comparison of Wolbachia and mitochondrial sequences indicates that the wMel strain has recently swept to high frequency at the expense of the wMelCS strain, suggesting that worldwide populations of D. melanogaster may have recently become more susceptible to viral infection [222,223,224]. In addition, one of the four native Wolbachia strains that infect Drosophila simulans (of which two exhibit pathogen interference) has rapidly (within approximately 10 years) swept through populations in California and Australia [197,225,226,227]. This strain, wRi, exhibits pathogen interference, suggesting that these D. simulans populations may have recently become more resistant to RNA viruses. As an estimated 40% of insect species may host Wolbachia and approximately half of Drosophila-infecting Wolbachia strains provide antiviral resistance [199], Wolbachia is likely to be a dynamic and sizable source of variation for virus susceptibility among individuals and species.
5.2. Wolbachia in Mosquitoes
Wolbachia is not found to be naturally associated with Ae. aegypti, although an Ae. albopictus strain from La Réunion Island naturally harbors two different strains of Wolbachia (wAlbA and wAlbB) [228]. These bacteria could reduce DENV transmission by limiting the amount of infectious virus particles secreted into the saliva, perhaps contributing to the reduced vector competence in this species relative to Ae. aegypti [228]. Artificial introduction of the Wolbachia wMel strain into Ae. aegypti limits DENV and CHIKV infection without adversely affecting mosquito life span [229]. This may allow the bacterium to spread and stably establish in the population, making it a promising candidate for introgression into Ae. aegypti [230]. Since 2011, there have been several efforts to release Wolbachia-infected Ae. aegypti into wild mosquito populations in Australia [231,232], showing the successful establishment of Wolbachia in Aedes populations and reduction of DENV transmission potential in field conditions [233]. Recently, the Eliminate Dengue Program, now known as the World Mosquito Program (https://www.worldmosquitoprogram.org), in partnership with organizations and governments of countries with high arbovirus transmission has initiated trials to evaluate the potential of wMel infected Ae. aegypti to suppress arbovirus transmission. As it constitutes an artificial intervention, variation in antiviral immunity introduced by transinfecting Ae. aegypti with Wolbachia will not be discussed further in this review.
5.3. Gut Microbiota in Drosophila
The interplay between gut commensals and viruses has been best studied in mammals, where the microbiota can be pro- or anti-viral, dependent on the virus [234]. This appears to be true in insects as well. For example, baculoviruses reach higher titers and are more virulent in the moth Spodoptera exigua when the microbiota is present [235]. Conversely, in D. melanogaster, the microbiota signals through Relish to upregulate Pvf2, which in turn activates antiviral ERK signaling in the gut [65,66]. Antibiotic treatment leads to increased replication rates of DCV and VSV [66], although this does not affect virus-induced mortality [57]. Upregulation of Pvf2 was strictly mediated by gram-negative bacteria (E. coli or Acetobacter species), although some bacterial species may be more potent activators [66]. Older flies have an altered microbiota and concomitantly higher levels of reactive oxygen species, which renders them unable to upregulate Pvf2 [236]. These phenotypes can be reversed by a faecal transplant from young to old flies, or with experimental association of old flies with Lactobacillus fructivorans and heat-killed Acetobacter pomorum [236]. The microbiota also influences gut environmental factors that likely have a role in antiviral immunity, including gut renewal rate and basal levels of JAK-STAT and IMD signaling [237]. Thus, it seems probable that the community structure of commensals plays an integral role in shaping the gut environment that enteric viruses encounter, and that variation in gut bacterial strains may translate into an important source of variation in natural populations. Indeed, microbiota is variable in wild flies [238,239]; however, frequencies of strains and species across worldwide populations is unknown, and little has been done to link specific commensals to antiviral resistance in flies [66,236].
5.4. Gut Microbiota in Mosquitoes
The mosquito midgut flora affects several mosquito traits, including development, nutrition, reproductive capacity and vector competence. Mosquitoes start acquiring their gut flora at the larval stage, the composition of which will be defined by the microbial flora of the larval aquatic habitat. The gut microflora is not static and changes later in the adult stage based on its sex, the flora acquired from nectar feeding, gut environmental changes through blood meals, and sometimes from venereal transmission (reviewed in [240]). As in Drosophila, the microbiota in mosquitoes may have complex and virus-specific effects on immunity and vector competence. The presence of different Gram-negative bacterial species such as Proteus, Paenibacillus and Chromobacterium in the Ae. aegypti midgut protects against DENV infection, whereas an opposite effect was seen for ONNV in its vector, Anopheles gambiae, in which treatment with antibiotics decreased susceptibility to ONNV [111,241,242]. Oral co-infection of a naturally occurring midgut bacterium of Ae. aegypti, Serratia odorifera, along with DENV or CHIKV increased its susceptibility to both arboviruses, presumably through the interaction of bacterial and mosquito midgut proteins [243,244]. In turn, the microbial community structure in the midguts of Ochlerotatus triseriatus and Aedes japonicus mosquitoes were significantly changed after a LACV infectious blood meal, suggesting an interaction between the vector gut microbiota and the infecting arbovirus, possibly influencing vector competence phenotypes [245].
These microbes could influence vector competence through different mechanisms, including innate immune priming and secretion of antiviral effectors. For example, the microbiota upregulates genes encoding antimicrobial peptides such as cecropins, defensins and lysozyme C that have known antiviral activity against DENV [53,60,241,242]. In addition, certain bacterial isolates belonging to the Enterobacteriaceae family, isolated from the midgut of Ae. albopictus were found to suppress replication of LACV in vertebrate cells, and a bacteria-free biofilm from Chromobacterium (isolate Csp_P) could inhibit DENV replication in vertebrate cells [242,246]. More recently, Talaromyces species of fungus isolated from the midgut of field-caught Ae. aegypti was shown to increase the permissiveness of the mosquito to DENV by secreting a hitherto unknown factor that downregulates trypsin gene expression and enzymatic activity in the gut [191]. This is line with the observation that a trypsin inhibitor or genetic knock-down of a late trypsin gene increased susceptibility to DENV [176]. Thus, the gut microbial flora is likely to be a dynamic source of natural variation for susceptibility to arboviral infection.
6. Other Sources of Variation
The resistance of an individual host is dependent on several other factors besides host genotype and the associated microbiota. The abiotic environment, sexual dimorphism, mating, infection history and endogenous viral elements have each been identified to influence infection outcomes. However, most of these factors have received disproportionate attention in either fruit flies or mosquitoes, and there have been few comparative studies of these effects across viruses, making generalizations between hosts and among viruses difficult.
6.1. Abiotic Environment
Variation in the abiotic environment, such as temperature, time of infection, or nutrient availability, can alter infection outcomes, either by interacting with the host immune system or the pathogen life cycle [74]. These factors have been investigated in laboratory experiments in Drosophila, although the ecological importance of each is not directly clear, as wild Drosophila have not been analyzed. For example, temperature influences the virulence of DmelSV and CrPV, and the titer of DmelSV and Kallithea virus in D. melanogaster [55,107,143]. Infection success may also depend on the time of day during which an infection occurs, as the antibacterial immune response appears to be more potent at night, in part due to a circadian rhythm-mediated increase in phagocytosis, although this has not been investigated in the context of viral infection [247,248,249]. Nutrient signaling also has substantial crosstalk with antiviral defense pathways, and starvation and macronutrient availability are known modulators of innate immunity [80,81,82,250,251]. Although associations between macronutrient profiles and efficacy of antiviral defense has not been assessed in Drosophila, African armyworms “self-medicate” with high protein, low carbohydrate diets after baculovirus infection and a similar diet leads to greater antibacterial defense in D. melanogaster [251,252]. Additionally, there is substantial genotype-by-nutrition variation in antibacterial immunity, and polymorphisms have been mapped which resistance to Providencia rettgeri in a diet-dependent manner [253]. Interestingly, most of these variants are in genes without a known role in immunity [253].
The interaction between immunity and the abiotic environment has been much better studied in mosquitoes, where vector competence is known to be modulated by variables emerging from different mosquito habitats. For example, larval environmental stressors like starvation, competition, elevated rearing temperatures, and low dose insecticide exposure were shown to increase the susceptibility of Ae. aegypti to SINV infection. This observation was attributed to the higher expression of antimicrobial peptides like cecropin and defensin as well as different stress-response genes like HSP70, HSP83, transferrin and CYP6Z6 [254].
Reduced nutrient availability or larval overcrowding may lead to stress, starvation, and a smaller adult size in mosquitoes, although the ultimate effect of these variables on vector competence is not clear. Nutritionally deprived and smaller-sized Ochlerotatus triseriatus females were better at transmitting LACV due to the thinner basal lamina layer of their midguts, thereby allowing more viruses to disseminate into the hemocoel and reach the salivary glands and be subsequently transmitted [255]. Similarly, laboratory-derived smaller Ae. aegypti mosquitoes were more susceptible to DENV infection, resulting in higher dissemination rates (frequency at which an arbovirus can infect and replicate in the midgut epithelial cells and cross over into the hemocoel) ([256], but see [257]). In contrast, a recent vector competence study of LACV in field collected Ochlerotatus triseriatus mosquitoes showed the opposite result, where infected females were larger than uninfected ones [258]. Increased larval competition also resulted in increased DENV infection rates in Ae. albopictus, although no such effects were observed with the lab-adapted Rockefeller strain of Ae. aegypti [259]. However, in a recent investigation, smaller-sized larvae from a Trinidadian strain of Ae. aegypti exhibited reduced susceptibility to DENV [260].
For many virus–vector combinations, it has been observed that mosquitoes held at lower temperatures during the extrinsic incubation period (EIP, time period between the acquisition of a virus through an infectious blood meal to its subsequent transmission to a susceptible vertebrate host) show reduced infection and dissemination and/or transmission rates [261,262,263,264]. A higher temperature could lead to a shorter extrinsic incubation period which in turn could lead to an increased transmission potential, as has been reported in DENV-infected Ae. aegypti and Ae. albopictus mosquitoes [265,266]. However, the relationship is not so straightforward and an inverse effect has been seen with WEEV-infected Cx. tarsalis mosquitoes and CHIKV-infected Ae. albopictus mosquitoes [267,268].
A mechanistic explanation for the effect of temperature on viral infection was proposed by Adelman et al. These authors found that although siRNA production was unaffected, RNAi efficiency was lower when Ae. aegypti mosquitoes were reared at a cooler temperature of 18 °C, which in turn could have led to increased replication of CHIKV and YFV in these mosquitoes [269]. The authors hypothesized that the differential effect of low temperature on virus replication and the RNAi machinery determines the eventual outcome on virus transmission [269,270].
Several studies have shown that large fluctuations in diurnal temperatures at a higher mean temperature (26°C–30°C) have a detrimental impact on development of immature stages, fecundity, survival and susceptibility to DENV infection by prolonging the EIP [271,272,273]. These results correlate well with the seasonal variation associated with DENV transmission in an endemic area like Thailand, where smaller variations in temperature are seen during the high transmission season as opposed to larger fluctuations during the low transmission season [271]. However, the same authors found that similar fluctuations at a lower mean temperature (20 °C) result in shorter EIPs and increased potential for DENV transmission [272]. It is thus evident that variation induced by temperature has a significant effect on vector competence. This environmental variable is multi-factorial and given the importance of climate change on the global spread of mosquitoes and arboviruses, it is paramount to model its effect on virus transmission using realistic and natural parameters.
6.2. Sexual Dimorphism and Mating
Sexual dimorphism in immunocompetence is common across animals, where there is a trend towards males suffering higher costs of infection [274]. In Drosophila, this may be true of viral infections, which seem to cause more harm in males (although female flies are more affected by bacterial infections) [275]. For example, the dicistroviruses DCV and CrPV are associated with higher mortality rates in males, and DCV grows to a higher titer, indicating that males are less resistant to these viruses [55,276]. Additionally, males have a lower tolerance to Kallithea virus infection, and die at a much higher rate than females, even though Kallithea virus replicates to a lower titer in males. Sex-specific immunity has not been well investigated in mosquitoes, because males are not hematophagous and unable to vector viruses of medical concern.
Although reduced male immune function can be theoretically explained by sex-specific investment in mating success and survival, the molecular mechanisms remain unclear, and may be specific to any given host–parasite combination [277,278,279]. Basic differences in immune-related traits between uninfected males and females may, in part, help to explain the observed sexual dimorphism in resistance and tolerance. For instance, females may be more developmentally resilient against infection. There is a higher basal rate of intestinal stem cell proliferation and gut renewal, more circulating phagocytes, and higher phenoloxidase activity in females [275,280,281]. Conversely, males may be more primed for resisting infection on a transcriptional level, and genes encoding for Toll pathway components tend to have higher basal expression in males [275].
In Drosophila, mating induces an immunosuppressive state, marked by variable AMP expression and reduced resistance to some bacterial infections [282,283,284,285]. Although this may occur in other insect species as well [286], it has not been well-studied in mosquitoes, presumably because blood-feeding is contingent on copulation. The reduction in immune function in female flies has been linked with increased synthesis of juvenile hormone (JH) [285,287,288], a multifunctional hormone required for oogenesis [289]. JH signals through the germ cells expressed (gce) receptor, and is itself activated by a male seminal protein, Sex Peptide (SP), that is transferred to the female during mating [290]. Removal of the corpus allatum (the endocrine gland where JH is synthesized) or genetic ablation of gce or JH in female flies, or SP in male flies, restored virgin levels of immunity post-copulation [285]. Although the effect of mating on antiviral immune function has not been studied in flies, a Heliothis zea (the corn earworm) nudivirus and a Mythimna separata (the northern armyworm) entomopoxvirus encode JH regulatory genes likely obtained by horizontal transfer, indicating JH synthesis may be beneficial to some DNA viruses [291,292].
6.3. Infection History
Infection with other viruses in the past and present may also influence the efficacy of antiviral responses or replication of cohabiting viruses. Co-infection may occur frequently in D. melanogaster, although interactions between viruses in this host have not been tested. In a Japanese population of singly collected flies, 42% were uninfected, 13% were infected with a single virus, whereas 19% had two infections, and the rest had multiple infections (n = 31) [293]. A similar pattern has been observed in honey bees [294]. This could either imply that there are flies with general susceptibility to diverse viruses, or that virus-by-virus interactions can promote infection in some cases, possibly through expression of virally-encoded immune inhibitors [295,296]. A consistent observation has been described in Cx. tritaeniorhynchus cells persistently infected with Culex flavivirus (CxFV), which led to slight enhancement in Japanese encephalitis virus (JEV) and DENV replication [297].
In areas endemic for arbovirus transmission, multiple arboviruses may be co-circulating at the same time. There have been several reports of DENV and CHIKV co-infections in humans during these outbreaks [298,299,300]. Recently, with outbreaks of DENV, CHIKV and ZIKV coinciding in Latin America, cases of triple co-infection have been detected [301]. The potential of Ae. aegypti and Ae. albopictus mosquitoes to simultaneously transmit both or all three viruses in their saliva was investigated by different groups. Indeed, multiple studies showed that a combination of two different viruses or all three could be transmitted at the same time in the mosquito saliva. Only minor interference effects were detected due to co-infection or infection of one virus prior to another [302,303,304]. As such, co-infection or dual infection of Aedes spp. mosquitoes with other co-circulating arboviruses do not seem to influence vector competence.
Alternatively, superinfection exclusion has been described in mosquito cells infected by mosquito-specific viruses (i.e., non-vectored mosquito viruses), whereby a virus-infected cell is refractory towards a secondary infection by a similar virus, although by an unknown mechanism [183,305]. For example, prior infection of Ae. albopictus C6/36 cells by Palm Creek virus (PCV) was found to moderately suppress the replication of two different flaviviruses—Murray Valley encephalitis virus and WNV [306]. In Cx. annulirostris, high PCV titers could suppress subsequent WNV infection when the virus was introduced through an infectious blood meal, although not when intra-thoracically inoculated. PCV was found to localize to mosquito midgut cells and may potentially interfere with midgut infection or escape [307]. Similarly, the insect-specific Nhumirim virus (NHUMV) was found to significantly interfere with the replication of WNV and SLEV in C6/36 cells, and to reduce WNV transmission by 40% in Cx. quinquefasciatus in vivo [308,309]. Finally, Eilat virus (EILV), a mosquito-specific alphavirus, interfered with a panel of alphaviruses in vitro, and delayed CHIKV infection in Ae. aegypti [310], similar to early CxFV inhibition of WNV in Cx. pipiens [311]. Despite considerable evidence that mosquito-specific viruses can modulate vector competence of several mosquito species, world-wide distributions and frequencies of co-infections are unknown, and no association of these viruses with refractory or susceptible populations of a particular mosquito species has been found.
Recently, a mechanism for antiviral immune memory has been described in flies, hinting that past infections could contribute to variation in defense among individuals. During infection, hemocytes take up virus and exogenous viral dsRNA, which are reverse transcribed into DNA molecules [23,24,25]. These vDNAs serve as a template for de novo synthesis of secondary siRNA, which are packaged into exosomes and are proposed to mediate a systemic immune response [25]. Although virus-specific immune memory has not been observed for DCV infections [312], immune memory has been observed in the Indian mealmoth, Plodia interpuctella, in response to a DNA virus, where the memory benefit can be trans-generational [313]. Therefore, successful mitigation of virus infection may render the host refractory towards future infection with similar strains.
6.4. Endogenous Viral Elements—Non-Retroviral Integrated RNA Virus Sequences
Arboviruses are non-retroviral positive or negative-sense RNA viruses that do not produce a DNA intermediate in their replication cycle. Surprisingly, however, arbovirus-derived viral DNA (vDNA) forms have been detected in infected mosquitoes and mosquito cells [314,315]. These vDNA forms are thought to be byproducts of reverse transcriptase activity by cellular retrotransposons [23,24]. Indeed, treating infected cells and mosquitoes with azidothymidine (a reverse transcriptase inhibitor) led to a dose-dependent reduction in vDNA and reduced production of virus-derived siRNAs in CHIKV-infected Ae. albopictus mosquitoes. Most importantly, loss of vDNAs reduced the survival of infected mosquitoes, suggesting that vDNA production is essential for tolerance to infection, possibly by modifying small RNA production [24]. If confirmed in other virus–vector combinations, these results suggest that a complex interplay between viruses, transposable elements, vDNA production and RNAi responses are crucial for sustaining a stable infection and thus for transmission to a susceptible mammalian host.
During the course of acute infection, vDNAs were also found in the wings and legs of infected mosquitoes, suggesting that these DNA elements may be episomal [24]. Although no experimental evidence exists at the moment, it is quite possible that these viral DNA forms are the precursors to endogenous viral elements (EVEs, also referred to as non-retroviral integrated RNA virus sequences), found integrated in the genomes of several vector mosquito species [314,316,317,318,319]. Recently, a comprehensive bioinformatic analysis revealed EVEs from different arboviral families (Flaviviridae, Rhabdoviridae, Reoviridae and members from the Order Bunyavirales) predominantly in the genomes of Ae. aegypti and Ae. albopictus and to a much lower extent in Cx. quinquefasciatus and Anopheline mosquitoes [320]. Most of these sequences belong to insect-specific viruses, which are likely maintained in nature by transovarial transmission and would therefore be more likely to infect germline cells, produce viral DNA forms, and be stably inherited [320]. EVEs were found to be significantly enriched in piRNA clusters, but were also found in genic and intergenic regions. More importantly, EVEs produced piRNAs in an antisense orientation to viral mRNA sequences, suggesting that they have antiviral potential. There is variation in the EVE content in the genomes from geographic populations of mosquitoes, suggesting that the acquisition of EVEs is a highly variable process and that the EVE repertoire could be an archive of past infections [320]. An exciting possibility is that integration of viral sequences in mosquito genomes coupled with their production of antisense piRNAs serve as an RNAi-based adaptive immune system and could potentially either target or help tolerate infections with cognate viruses [321]. These new insights into the dynamic composition of mosquito genomes coupled with the compelling phenotype of vDNA-associated tolerance provide a new perspective on the molecular basis for variation in virus–vector interactions.
7. Concluding Remarks
The outcome of viral infection is characterized, in part, by general immune responses, virus-specific responses, and variation among host individuals. The mechanisms and relative importance of these factors in host–virus interactions have been the focus of many studies on Drosophilidae and Culicidae insects and their infecting viruses. D. melanogaster has been an instrumental model in discerning molecular mechanisms of immunity, and large infection experiments across genotypes have led to the identification of antiviral resistance loci. However, only five natural viruses have been isolated, and, of these, three have been used in genome-wide association studies (GWAS) or mapping studies. GWAS for resistance to non-natural viruses found no associations, underlining the importance of shared coevolutionary history in maintaining genetic variation for resistance [130]. Thus, the isolation of new Drosophila viruses will enable a more powerful comparative framework to assess the generality of immune responses and patterns of genetic variation in resistance. In contrast, there are many isolates of native mosquito viruses, but very few studies focusing on identifying host loci underlying resistance to viral infection. Although large-effect QTL have been mapped for DENV and LACV transmission (Table 2), difficult lab rearing, a lack of genomic tools, highly structured populations, and an incomplete understanding of arbovirus host range has largely precluded fine scale mapping experiments or GWAS of viral resistance (Table 2) [10,137]. The recent release of the complete Ae. aegypti genome marks an important step in overcoming these hurdles, and may make quantitative genetics in this species more feasible [322]. Comparative transcriptomic studies in Ae. aegypti populations susceptible and refractory to DENV infection have led to the identification of a few host genes that may contribute to differential vector competence [184,185]. However, many of these genes need to be validated in multiple susceptible versus refractory populations and their precise molecular mechanisms need to be characterized. The further implementation of gene-knockout techniques in live mosquitoes will undoubtedly shed further light on the role of these factors in determining vector competence.
Acknowledgments
This work is financially supported by a Consolidator Grant from the European Research Council under the European Union’s Seventh Framework Programme (Grant Number ERC CoG 615680) and a VICI grant from the Netherlands Organization for Scientific Research (Grant Number 016.VICI.170.090) to Ronald P. van Rij. William H. Palmer is supported in the laboratory of Ronald P. van Rij by a Short Term Fellowship from the a European Molecular Biology Organization (Grant Number 7095). We thank Darren Obbard for helpful discussions during the preparation of this manuscript. The figure in this article was created using BioRender.
Conflicts of Interest
The authors declare no conflicts of interest.
References
- Merkling, S.H.; van Rij, R.P. Beyond RNAi: Antiviral defense strategies in Drosophila and mosquito. J. Insect Physiol. 2013, 59, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Kingsolver, M.B.; Huang, Z.; Hardy, R.W. Insect antiviral innate immunity: Pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, A.W.; van Rij, R.P. The long and short of antiviral defense: Small RNA-based immunity in insects. Curr. Opin. Virol. 2014, 7, 19–28. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, O.; Imler, J.L. Induced antiviral innate immunity in Drosophila. Curr. Opin. Microbiol. 2014, 20, 62–68. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Dimopoulos, G. Mosquito immunity against arboviruses. Viruses 2014, 6, 4479–4504. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Cherry, S. Viruses and antiviral immunity in Drosophila. Dev. Comp. Immunol. 2014, 42, 67–84. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Antiviral Immunity and Virus-Mediated Antagonism in Disease Vector Mosquitoes. Trends Microbiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Franz, A.W.E.; Kantor, A.M.; Passarelli, A.L.; Clem, R.J. Tissue Barriers to Arbovirus Infection in Mosquitoes. Viruses 2015, 7, 3741–3767. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambrechts, L.; Chevillon, C.; Albright, R.G.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Scott, T.W. Genetic specificity and potential for local adaptation between dengue viruses and mosquito vectors. BMC Evol. Biol. 2009, 9, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L. Quantitative genetics of Aedes aegypti vector competence for dengue viruses: Towards a new paradigm? Trends Parasitol. 2011, 27, 111–114. [Google Scholar] [CrossRef] [PubMed]
- Brackney, D.E.; Beane, J.E.; Ebel, G.D. RNAi Targeting of West Nile Virus in Mosquito Midguts Promotes Virus Diversification. PLoS Pathog. 2009, 5, e1000502. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.L.; Coffey, L.L.; Weaver, S.C. Arboviral bottlenecks and challenges to maintaining diversity and fitness during mosquito transmission. Viruses 2014, 6, 3991–4004. [Google Scholar] [CrossRef] [PubMed]
- Lequime, S.; Fontaine, A.; Ar Gouilh, M.; Moltini-Conclois, I.; Lambrechts, L. Genetic Drift, Purifying Selection and Vector Genotype Shape Dengue Virus Intra-host Genetic Diversity in Mosquitoes. PLoS Genet. 2016, 12, e1006111. [Google Scholar] [CrossRef] [PubMed]
- Grubaugh, N.D.; Weger-Lucarelli, J.; Murrieta, R.A.; Fauver, J.R.; Garcia-Luna, S.M.; Prasad, A.N.; Black, W.C.; Ebel, G.D. Genetic Drift during Systemic Arbovirus Infection of Mosquito Vectors Leads to Decreased Relative Fitness during Host Switching. Cell Host Microbe 2016, 19, 481–492. [Google Scholar] [CrossRef] [PubMed]
- Brault, A.C.; Powers, A.M.; Ortiz, D.; Estrada-Franco, J.G.; Navarro-Lopez, R.; Weaver, S.C. Venezuelan equine encephalitis emergence: Enhanced vector infection from a single amino acid substitution in the envelope glycoprotein. Proc. Natl. Acad. Sci. USA 2004, 101, 11344–11349. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Vanlandingham, D.L.; McGee, C.E.; Higgs, S. A single mutation in Chikungunya virus affects vector specificity and epidemic potential. PLoS Pathog. 2007, 3, 1895–1906. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Weaver, S.C. Sequential adaptive mutations enhance efficient vector switching by chikungunya virus and its epidemic emergence. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Tsetsarkin, K.A.; Chen, R.; Leal, G.; Forrester, N.; Higgs, S.; Huang, J.; Weaver, S.C. Chikungunya virus emergence is constrained in Asia by lineage-specific adaptive landscapes. Proc. Natl. Acad. Sci. USA 2011, 108, 7872–7877. [Google Scholar] [CrossRef] [PubMed]
- Vignuzzi, M.; Higgs, S. The Bridges and Blockades to Evolutionary Convergence on the Road to Predicting Chikungunya Virus Evolution. Annu. Rev. Virol. 2017, 4, 181–200. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, J.T.; Bronkhorst, A.W.; Overheul, G.J.; Sadanandan, S.A.; Ekström, J.O.; Heestermans, M.; Hultmark, D.; Antoniewski, C.; van Rij, R.P. Convergent Evolution of Argonaute-2 Slicer Antagonism in Two Distinct Insect RNA Viruses. PLoS Pathog. 2012, 8, e1002872. [Google Scholar] [CrossRef] [PubMed]
- Kemp, C.; Mueller, S.; Goto, A.; Barbier, V.; Paro, S.; Bonnay, F.; Dostert, C.; Troxler, L.; Hetru, C.; Meignin, C.; et al. Broad RNA Interference-Mediated Antiviral Immunity and Virus-Specific Inducible Responses in Drosophila. J. Immunol. 2013, 190, 650–658. [Google Scholar] [CrossRef] [PubMed]
- Olson, K.E.; Blair, C.D. Arbovirus-mosquito interactions: RNAi pathway. Curr. Opin. Virol. 2015, 15, 119–126. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Vodovar, N.; Mondotte, J.A.; Monot, C.; Frangeul, L.; Blanc, H.; Gausson, V.; Vera-Otarola, J.; Cristofari, G.; Saleh, M.C. RNA-mediated interference and reverse transcription control the persistence of RNA viruses in the insect model Drosophila. Nat. Immunol. 2013, 14, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Goic, B.; Stapleford, K.A.; Frangeul, L.; Doucet, A.J.; Gausson, V.; Blanc, H.; Schemmel-Jofre, N.; Cristofari, G.; Lambrechts, L.; Vignuzzi, M.; et al. Virus-derived DNA drives mosquito vector tolerance to arboviral infection. Nat. Commun. 2016, 7, 12410. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassetto, M.; Kunitomi, M.; Andino, R. Circulating Immune Cells Mediate a Systemic RNAi-Based Adaptive Antiviral Response in Drosophila. Cell 2017, 169, 314–325. [Google Scholar] [CrossRef] [PubMed]
- Petit, M.; Mongelli, V.; Frangeul, L.; Blanc, H.; Jiggins, F.; Saleh, M.-C. piRNA pathway is not required for antiviral defense in Drosophila melanogaster. Proc. Natl. Acad. Sci. USA 2016, 113, E4218–E4227. [Google Scholar] [CrossRef] [PubMed]
- Vodovar, N.; Bronkhorst, A.W.; van Cleef, K.W.R.; Miesen, P.; Blanc, H.; van Rij, R.P.; Saleh, M.-C. Arbovirus-Derived piRNAs Exhibit a Ping-Pong Signature in Mosquito Cells. PLoS ONE 2012, 7, e30861. [Google Scholar] [CrossRef] [PubMed]
- Morazzani, E.M.; Wiley, M.R.; Murreddu, M.G.; Adelman, Z.N.; Myles, K.M. Production of Virus-Derived Ping-Pong-Dependent piRNA-like Small RNAs in the Mosquito Soma. PLoS Pathog. 2012, 8, e1002470. [Google Scholar] [CrossRef] [PubMed]
- Campbell, C.L.; Black, W.C.; Hess, A.M.; Foy, B.D. Comparative genomics of small RNA regulatory pathway components in vector mosquitoes. BMC Genom. 2008, 9, 425. [Google Scholar] [CrossRef] [PubMed]
- Miesen, P.; Girardi, E.; van Rij, R.P. Distinct sets of PIWI proteins produce arbovirus and transposon-derived piRNAs in Aedes aegypti mosquito cells. Nucleic Acids Res. 2015, 43, 6545–6556. [Google Scholar] [CrossRef] [PubMed]
- Keene, K.M.; Foy, B.D.; Sanchez-Vargas, I.; Beaty, B.J.; Blair, C.D.; Olson, K.E. RNA interference acts as a natural antiviral response to O’nyong-nyong virus (Alphavirus; Togaviridae) infection of Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2004, 101, 17240–17245. [Google Scholar] [CrossRef] [PubMed]
- Schnettler, E.; Donald, C.L.; Human, S.; Watson, M.; Siu, R.W.C.; McFarlane, M.; Fazakerley, J.K.; Kohl, A.; Fragkoudis, R. Knockdown of piRNA pathway proteins results in enhanced semliki forest virus production in mosquito cells. J. Gen. Virol. 2013, 94, 1680–1689. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Varjak, M.; Maringer, K.; Watson, M.; Sreenu, V.B.; Fredericks, A.C.; Pondeville, E.; Donald, C.L.; Sterk, J.; Kean, J.; Vazeille, M.; et al. Aedes aegypti Piwi4 Is a Noncanonical PIWI Protein Involved in Antiviral Responses. mSphere 2017, 2, e00144-17. [Google Scholar] [CrossRef] [PubMed]
- Shoemaker, C.J.; Green, R. Translation drives mRNA quality control. Nat. Struct. Mol. Biol. 2012, 19, 594–601. [Google Scholar] [CrossRef] [PubMed]
- Schoenberg, D.R.; Maquat, L.E. Regulation of cytoplasmic mRNA decay. Nat. Rev. Genet. 2012, 13, 246–259. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Wilusz, J. Cytoplasmic Viruses: Rage against the (Cellular RNA Decay) Machine. PLoS Pathog. 2013, 9, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Molleston, J.M.; Cherry, S. Attacked from all sides: RNA decay in antiviral defense. Viruses 2017, 9. [Google Scholar] [CrossRef] [PubMed]
- Orban, T.I.; Izaurralde, E. Decay of mRNAs targeted by RISC requires XRN1, the Ski complex, and the exosome. RNA 2005, 11, 459–469. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, K.C.; McLane, L.M.; Maqbool, T.; Panda, D.; Gordesky-Gold, B.; Cherry, S. A genome-wide RNAi screen reveals that mRNA decapping restricts bunyaviral replication by limiting the pools of dcp2-accessible targets for cap-snatching. Genes Dev. 2013, 27, 1511–1525. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Magwire, M.M.; Bayer, F.; Jiggins, F.M. A Polymorphism in the Processing Body Component Ge-1 Controls Resistance to a Naturally Occurring Rhabdovirus in Drosophila. PLoS Pathog. 2016, 12. [Google Scholar] [CrossRef]
- Molleston, J.M.; Sabin, L.R.; Moy, R.H.; Menghani, S.V.; Rausch, K.; Gordesky-Gold, B.; Hopkins, K.C.; Zhou, R.; Jensen, T.H.; Wilusz, J.E.; et al. A conserved virus-induced cytoplasmic TRAMP-like complex recruits the exosome to target viral RNA for degradation. Genes Dev. 2016, 30, 1658–1670. [Google Scholar] [CrossRef] [PubMed]
- Khong, A.; Jan, E. Modulation of Stress Granules and P Bodies during Dicistrovirus Infection. J. Virol. 2011, 85, 1439–1451. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.L.; Anderson, J.R.; Kumagai, Y.; Wilusz, C.J.; Akira, S.; Khromykh, A.A.; Wilusz, J. A noncoding RNA produced by arthropod-borne flaviviruses inhibits the cellular exoribonuclease XRN1 and alters host mRNA stability. RNA 2012, 18, 2029–2040. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Samuel, C.E. ADARs: Viruses and innate immunity. Curr. Top. Microbiol. Immunol. 2012, 353, 163–195. [Google Scholar] [CrossRef] [PubMed]
- Carpenter, J.A.; Keegan, L.P.; Wilfert, L.; O’Connell, M.A.; Jiggins, F.M. Evidence for ADAR-induced hypermutation of the Drosophila sigma virus (Rhabdoviridae). BMC Genet. 2009, 10, 75. [Google Scholar] [CrossRef] [PubMed]
- Piontkivska, H.; Matos, L.F.; Paul, S.; Scharfenberg, B.; Farmerie, W.G.; Miyamoto, M.M.; Wayne, M.L. Role of host-driven mutagenesis in determining genome evolution of sigma virus (DMelSV; Rhabdoviridae) in Drosophila melanogaster. Genome Biol. Evol. 2016, 8, 2952–2963. [Google Scholar] [CrossRef] [PubMed]
- Piontkivska, H.; Frederick, M.; Miyamoto, M.M.; Wayne, M.L. RNA editing by the host ADAR system affects the molecular evolution of the Zika virus. Ecol. Evol. 2017, 7, 4475–4485. [Google Scholar] [CrossRef] [PubMed]
- Valanne, S.; Wang, J.-H.; Ramet, M. The Drosophila Toll Signaling Pathway. J. Immunol. 2011, 186, 649–656. [Google Scholar] [CrossRef] [PubMed]
- Myllymaki, H.; Valanne, S.; Ramet, M. The Drosophila Imd Signaling Pathway. J. Immunol. 2014, 192, 3455–3462. [Google Scholar] [CrossRef] [PubMed]
- Silverman, N.; Zhou, R.; Erlich, R.L.; Hunter, M.; Bernstein, E.; Schneider, D.; Maniatis, T. Immune activation of NF-kappaB and JNK requires Drosophila TAK1. J. Biol. Chem. 2003, 278, 48928–48934. [Google Scholar] [CrossRef] [PubMed]
- Delaney, J.R.; Stöven, S.; Uvell, H.; Anderson, K.V.; Engström, Y.; Mlodzik, M. Cooperative control of Drosophila immune responses by the JNK and NF-κB signaling pathways. EMBO J. 2006, 25, 3068–3077. [Google Scholar] [CrossRef] [PubMed]
- Zambon, R.A.; Nandakumar, M.; Vakharia, V.N.; Wu, L.P. The Toll pathway is important for an antiviral response in Drosophila. Proc. Natl. Acad. Sci. USA 2005, 102, 7257–7262. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.; Ramirez, J.L.; Dimopoulos, G. The Aedes aegypti toll pathway controls dengue virus infection. PLoS Pathog. 2008, 4, e1000098. [Google Scholar] [CrossRef] [PubMed]
- Avadhanula, V.; Weasner, B.P.; Hardy, G.G.; Kumar, J.P.; Hardy, R.W. A novel system for the launch of alphavirus RNA synthesis reveals a role for the Imd pathway in arthropod antiviral response. PLoS Pathog. 2009, 5, e1000582. [Google Scholar] [CrossRef] [PubMed]
- Costa, A.; Jan, E.; Sarnow, P.; Schneider, D. The Imd pathway is involved in antiviral immune responses in Drosophila. PLoS ONE 2009, 4, e7436. [Google Scholar] [CrossRef] [PubMed]
- Nakamoto, M.; Moy, R.H.; Xu, J.; Bambina, S.; Yasunaga, A.; Shelly, S.S.; Gold, B.; Cherry, S. Virus Recognition by Toll-7 Activates Antiviral Autophagy in Drosophila. Immunity 2012, 36, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, Á.G.; Naylor, H.; Esteves, S.S.; Pais, I.S.; Martins, N.E.; Teixeira, L. The Toll-Dorsal Pathway Is Required for Resistance to Viral Oral Infection in Drosophila. PLoS Pathog. 2014, 10, e1004507. [Google Scholar] [CrossRef] [PubMed]
- Thoetkiattikul, H.; Beck, M.H.; Strand, M.R. Inhibitor kappaB-like proteins from a polydnavirus inhibit NF-kappaB activation and suppress the insect immune response. Proc. Natl. Acad. Sci. USA 2005, 102, 11426–11431. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, O.; Kellenberger, C.; Kemp, C.; Troxler, L.; Pelte, N.; Boutros, M.; Marques, J.T.; Daeffler, L.; Hoffmann, J.A.; Roussel, A.; et al. Cytokine Diedel and a viral homologue suppress the IMD pathway in Drosophila. Proc. Natl. Acad. Sci. USA 2016, 113, 698–703. [Google Scholar] [CrossRef] [PubMed]
- Luplertlop, N.; Surasombatpattana, P.; Patramool, D.; Dumas, E.; Wasinpiyamongkol, L.; Saune, L.; Hamel, R.; Bernard, E.; Sereno, D.; Thomas, F.R.; et al. Induction of a peptide with activity against a broad spectrum of pathogens in the Aedes aegypti salivary gland, following infection with Dengue Virus. PLoS Pathog. 2011, 7, e1001252. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Kingsolver, M.B.; Avadhanula, V.; Hardy, R.W. An Antiviral Role for Antimicrobial Peptides during the Arthropod Response to Alphavirus Replication. J. Virol. 2013, 87, 4272–4280. [Google Scholar] [CrossRef] [PubMed]
- Qiu, P.; Pan, P.C.; Govind, S. A role for the Drosophila Toll/Cactus pathway in larval hematopoiesis. Development 1998, 125, 1909–1920. [Google Scholar] [PubMed]
- Georgel, P.; Naitza, S.; Kappler, C.; Ferrandon, D.; Zachary, D.; Swimmer, C.; Kopczynski, C.; Duyk, G.; Reichhart, J.M.; Hoffmann, J.A. Drosophila Immune Deficiency (IMD) Is a Death Domain Protein that Activates Antibacterial Defense and Can Promote Apoptosis. Dev. Cell 2001, 1, 503–514. [Google Scholar] [CrossRef]
- Liu, B.; Behura, S.K.; Clem, R.J.; Schneemann, A.; Becnel, J.; Severson, D.W.; Zhou, L. P53-Mediated Rapid Induction of Apoptosis Conveys Resistance to Viral Infection in Drosophila melanogaster. PLoS Pathog. 2013, 9, e1003137. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hopkins, K.; Sabin, L.; Yasunaga, A.; Subramanian, H.; Lamborn, I.; Gordesky-Gold, B.; Cherry, S. ERK signaling couples nutrient status to antiviral defense in the insect gut. Proc. Natl. Acad. Sci. USA 2013, 110, 15025–15030. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.L.; Cohen, J.; Yasunaga, A.; Xu, J.; Osborn, G.; Subramanian, H.; Gold, B.; Buchon, N.; Cherry, S. Microbiota-dependent priming of antiviral intestinal immunity in Drosophila. Cell Host Microbe 2015, 18, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Arbouzova, N.I.; Zeidler, M.P. JAK/STAT signalling in Drosophila: Insights into conserved regulatory and cellular functions. Development 2006, 133, 2605–2616. [Google Scholar] [CrossRef] [PubMed]
- Deddouche, S.; Matt, N.; Budd, A.; Mueller, S.; Kemp, C.; Galiana-Arnoux, D.; Dostert, C.; Antoniewski, C.; Hoffmann, J.A.; Imler, J.L. The DExD/H-box helicase Dicer-2 mediates the induction of antiviral activity in Drosophila. Nat. Immunol. 2008, 9, 1425–1432. [Google Scholar] [CrossRef] [PubMed]
- Paradkar, P.N.; Trinidad, L.; Voysey, R.; Duchemin, J.-B.; Walker, P.J. Secreted Vago restricts West Nile virus infection in Culex mosquito cells by activating the Jak-STAT pathway. Proc. Natl. Acad. Sci. USA 2012, 109, 18915–18920. [Google Scholar] [CrossRef] [PubMed]
- Jupatanakul, N.; Sim, S.; Angleró-Rodríguez, Y.I.; Souza-Neto, J.; Das, S.; Poti, K.E.; Rossi, S.L.; Bergren, N.; Vasilakis, N.; Dimopoulos, G. Engineered Aedes aegypti JAK/STAT Pathway-Mediated Immunity to Dengue Virus. PLoS Negl. Trop. Dis. 2017, 11, e0005187. [Google Scholar] [CrossRef] [PubMed]
- Dostert, C.; Jouanguy, E.; Irving, P.; Troxler, L.; Galiana-Arnoux, D.; Hetru, C.; Hoffmann, J.A.; Imler, J.L. The Jak-STAT signaling pathway is required but not sufficient for the antiviral response of Drosophila. Nat. Immunol. 2005, 6, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Merkling, S.H.; Bronkhorst, A.W.; Kramer, J.M.; Overheul, G.J.; Schenck, A.; Van Rij, R.P. The Epigenetic Regulator G9a Mediates Tolerance to RNA Virus Infection in Drosophila. PLoS Pathog. 2015, 11, e1004692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Souza-Neto, J.A.; Sim, S.; Dimopoulos, G. An evolutionary conserved function of the JAK-STAT pathway in anti-dengue defense. Proc. Natl. Acad. Sci. USA 2009, 106, 17841–17846. [Google Scholar] [CrossRef] [PubMed]
- Lazzaro, B.P.; Little, T.J. Immunity in a variable world. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 15–26. [Google Scholar] [CrossRef] [PubMed]
- Ponton, F.; Wilson, K.; Holmes, A.J.; Cotter, S.C.; Raubenheimer, D.; Simpson, S.J. Integrating nutrition and immunology: A new frontier. J. Insect Physiol. 2013, 59, 130–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Partridge, L.; Alic, N.; Bjedov, I.; Piper, M.D.W. Ageing in Drosophila: The role of the insulin/Igf and TOR signalling network. Exp. Gerontol. 2011, 46, 376–381. [Google Scholar] [CrossRef] [PubMed]
- Shelly, S.; Lukinova, N.; Bambina, S.; Berman, A.; Cherry, S. Autophagy Is an Essential Component of Drosophila Immunity against Vesicular Stomatitis Virus. Immunity 2009, 30, 588–598. [Google Scholar] [CrossRef] [PubMed]
- Moy, R.H.; Gold, B.; Molleston, J.M.; Schad, V.; Yanger, K.; Salzano, M.V.; Yagi, Y.; Fitzgerald, K.A.; Stanger, B.Z.; Soldan, S.S.; et al. Antiviral autophagy restricts rift valley fever virus infection and is conserved from flies to mammals. Immunity 2014, 40, 51–65. [Google Scholar] [CrossRef] [PubMed]
- Lamiable, O.; Arnold, J.; da Silva de Faria, I.J.; Proveti Olmo, R.; Bergami, F.; Meignin, C.; Hoffmann, J.A.; Marques, J.T.; Imler, J.-L. Analysis of the contribution of hemocytes and autophagy to Drosophila antiviral immunity. J. Virol. 2016, 90, JVI.00238-16. [Google Scholar] [CrossRef] [PubMed]
- DiAngelo, J.R.; Bland, M.L.; Bambina, S.; Cherry, S.; Birnbaum, M.J. The immune response attenuates growth and nutrient storage in Drosophila by reducing insulin signaling. Proc. Natl. Acad. Sci. USA 2009, 106, 20853–20858. [Google Scholar] [CrossRef] [PubMed]
- Becker, T.; Loch, G.; Beyer, M.; Zinke, I.; Aschenbrenner, A.C.; Carrera, P.; Inhester, T.; Schultze, J.L.; Hoch, M. FOXO-dependent regulation of innate immune homeostasis. Nature 2010, 463, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Spellberg, M.J.; Marr, M.T. FOXO regulates RNA interference in Drosophila and protects from RNA virus infection. Proc. Natl. Acad. Sci. USA 2015, 112, 14587–14592. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.K.; Hardy, R.W. Role for the Phosphatidylinositol 3-Kinase-Akt-TOR Pathway during Sindbis Virus Replication in Arthropods. J. Virol. 2012, 86, 3595–3604. [Google Scholar] [CrossRef] [PubMed]
- Everett, H.; McFadden, G. Apoptosis: An innate immune response to virus infection. Trends Microbiol. 1999, 7, 160–165. [Google Scholar] [CrossRef]
- Steller, H. Regulation of apoptosis in Drosophila. Cell Death Differ. 2008, 15, 1132–1138. [Google Scholar] [CrossRef] [PubMed]
- Vandergaast, R.; Mitchell, J.K.; Byers, N.M.; Friesen, P.D. Insect Inhibitor-of-Apoptosis (IAP) Proteins Are Negatively Regulated by Signal-Induced N-Terminal Degrons Absent within Viral IAP Proteins. J. Virol. 2015, 89, 4481–4493. [Google Scholar] [CrossRef] [PubMed]
- Vandergaast, R.; Schultz, K.L.W.; Cerio, R.J.; Friesen, P.D. Active Depletion of Host Cell Inhibitor-of-Apoptosis Proteins Triggers Apoptosis upon Baculovirus DNA Replication. J. Virol. 2011, 85, 8348–8358. [Google Scholar] [CrossRef] [PubMed]
- Nainu, F.; Tanaka, Y.; Shiratsuchi, A.; Nakanishi, Y. Protection of Insects against Viral Infection by Apoptosis-Dependent Phagocytosis. J. Immunol. 2015, 195, 5696–5706. [Google Scholar] [CrossRef] [PubMed]
- Ye, T.; Zhang, X. Involvement of Ran in the regulation of phagocytosis against virus infection in S2 cells. Dev. Comp. Immunol. 2013, 41, 491–497. [Google Scholar] [CrossRef] [PubMed]
- Zhu, F.; Zhang, X. The Wnt signaling pathway is involved in the regulation of phagocytosis of virus in Drosophila. Sci. Rep. 2013, 3, 2069. [Google Scholar] [CrossRef] [PubMed]
- Moy, R.H.; Cherry, S. Antimicrobial autophagy: A conserved innate immune response in Drosophila. J. Innate Immun. 2013, 5, 444–455. [Google Scholar] [CrossRef] [PubMed]
- Yasunaga, A.; Hanna, S.L.; Li, J.; Cho, H.; Rose, P.P.; Spiridigliozzi, A.; Gold, B.; Diamond, M.S.; Cherry, S. Genome-Wide RNAi Screen Identifies Broadly-Acting Host Factors That Inhibit Arbovirus Infection. PLoS Pathog. 2014, 10, e1003914. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Weaver, S.C.; Tesh, R.B.; Vasilakis, N. Insect-specific virus discovery: Significance for the arbovirus community. Viruses 2015, 7, 4911–4928. [Google Scholar] [CrossRef] [PubMed]
- Webster, C.L.; Waldron, F.M.; Robertson, S.; Crowson, D.; Ferrari, G.; Quintana, J.F.; Brouqui, J.-M.; Bayne, E.H.; Longdon, B.; Buck, A.H.; et al. The Discovery, Distribution, and Evolution of Viruses Associated with Drosophila melanogaster. PLoS Biol. 2015, 13, e1002210. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Webster, C.L.; Longdon, B.; Lewis, S.H.; Obbard, D.J. Twenty-Five New Viruses Associated with the Drosophilidae (Diptera). Evol. Bioinform. Online 2016, 12, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Medd, N.C.; Fellous, S.; Nakai, M.; Xuereb, A.; Cross, J.V.; Obbard, D.J. The virome of Drosophila suzukii, an invasive pest of soft fruit. bioRxiv 2017, 190322. [Google Scholar] [CrossRef]
- Li, H.; Li, W.X.; Ding, S.W. Induction and suppression of RNA silencing by an animal virus. Science 2002, 296, 1319–1321. [Google Scholar] [CrossRef] [PubMed]
- Van Rij, R.P.; Saleh, M.C.; Berry, B.; Foo, C.; Houk, A.; Antoniewski, C.; Andino, R. The RNA silencing endonuclease Argonaute 2 mediates specific antiviral immunity in Drosophila melanogaster. Genes Dev. 2006, 20, 2985–2995. [Google Scholar] [CrossRef] [PubMed]
- Nayak, A.; Berry, B.; Tassetto, M.; Kunitomi, M.; Acevedo, A.; Deng, C.; Krutchinsky, A.; Gross, J.; Antoniewski, C.; Andino, R. Cricket paralysis virus antagonizes Argonaute 2 to modulate antiviral defense in Drosophila. Nat. Struct. Mol. Biol. 2010, 17, 547–554. [Google Scholar] [CrossRef] [PubMed]
- Bronkhorst, A.W.; Van Cleef, K.W.R.; Venselaar, H.; Van Rij, R.P. A dsRNA-binding protein of a complex invertebrate DNA virus suppresses the Drosophila RNAi response. Nucleic Acids Res. 2014, 42, 12237–12248. [Google Scholar] [CrossRef] [PubMed]
- Van Cleef, K.W.R.; Van Mierlo, J.T.; Miesen, P.; Overheul, G.J.; Fros, J.J.; Schuster, S.; Marklewitz, M.; Pijlman, G.P.; Junglen, S.; Van Rij, R.P. Mosquito and Drosophila entomobirnaviruses suppress dsRNA- and siRNA-induced RNAi. Nucleic Acids Res. 2014, 42, 8732–8744. [Google Scholar] [CrossRef] [PubMed]
- Van Mierlo, J.T.; Overheul, G.J.; Obadia, B.; van Cleef, K.W.R.; Webster, C.L.; Saleh, M.C.; Obbard, D.J.; van Rij, R.P. Novel Drosophila Viruses Encode Host-Specific Suppressors of RNAi. PLoS Pathog. 2014, 10, e1004256. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Csorba, T.; Kontra, L.; Burgyán, J. Viral silencing suppressors: Tools forged to fine-tune host-pathogen coexistence. Virology 2015, 479–480, 85–103. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Habayeb, M.S.; Ekström, J.O.; Hultmark, D. Nora virus persistent infections are not affected by the RNAi machinery. PLoS ONE 2009, 4, e5731. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Grant, G.; Sabin, L.R.; Gordesky-Gold, B.; Yasunaga, A.; Tudor, M.; Cherry, S. Transcriptional pausing controls a rapid antiviral innate immune response in Drosophila. Cell Host Microbe 2012, 12, 531–543. [Google Scholar] [CrossRef] [PubMed]
- Tsai, C.W.; McGraw, E.A.; Ammar, E.-D.; Dietzgen, R.G.; Hogenhout, S.A. Drosophila melanogaster Mounts a Unique Immune Response to the Rhabdovirus Sigma virus. Appl. Environ. Microbiol. 2008, 74, 3251–3256. [Google Scholar] [CrossRef] [PubMed]
- Palmer, W.H.; Medd, N.; Beard, P.M.; Obbard, D.J. Isolation of a natural DNA virus of Drosophila melanogaster, and characterisation of host resistance and immune responses. bioRxiv 2017, 215111. [Google Scholar] [CrossRef]
- Ramirez, J.L.; Dimopoulos, G. The Toll immune signaling pathway control conserved anti-dengue defenses across diverse Ae. aegypti strains and against multiple dengue virus serotypes. Dev. Comp. Immunol. 2010, 34, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Fragkoudis, R.; Chi, Y.; Siu, R.W.C.; Barry, G.; Attarzadeh-Yazdi, G.; Merits, A.; Nash, A.A.; Fazakerley, J.K.; Kohl, A. Semliki Forest virus strongly reduces mosquito host defence signaling. Insect Mol. Biol. 2008, 17, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Waldock, J.; Olson, K.E.; Christophides, G.K. Anopheles gambiae Antiviral Immune Response to Systemic O’nyong-nyong Infection. PLoS Negl. Trop. Dis. 2012, 6, e1565. [Google Scholar] [CrossRef] [PubMed]
- Carissimo, G.; Pondeville, E.; McFarlane, M.; Dietrich, I.; Mitri, C.; Bischoff, E.; Antoniewski, C.; Bourgouin, C.; Failloux, A.-B.; Kohl, A.; et al. Antiviral immunity of Anopheles gambiae is highly compartmentalized, with distinct roles for RNA interference and gut microbiota. Proc. Natl. Acad. Sci. USA 2015, 112, E176–E185. [Google Scholar] [CrossRef] [PubMed]
- Mondotte, J.A.; Saleh, M.C. Antiviral Immune Response and the Route of Infection in Drosophila melanogaster. In Advances in Virus Research, 1st ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2017. [Google Scholar]
- Prasad, A.N.; Brackney, D.E.; Ebel, G.D. The role of innate immunity in conditioning mosquito susceptibility to West Nile virus. Viruses 2013, 5, 3142–3170. [Google Scholar] [CrossRef] [PubMed]
- Colpitts, T.M.; Cox, J.; Vanlandingham, D.L.; Feitosa, F.M.; Cheng, G.; Kurscheid, S.; Wang, P.; Krishnan, M.N.; Higgs, S.; Fikrig, E. Alterations in the Aedes aegypti transcriptome during infection with west nile, dengue and yellow fever viruses. PLoS Pathog. 2011, 7. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Zhang, R.; Zhang, B.; Zhao, T.; Wang, P.; Liang, G.; Cheng, G. Blood meal acquisition enhances arbovirus replication in mosquitoes through activation of the GABAergic system. Nat. Commun. 2017, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Settles, E.W.; Friesen, P.D. Flock House Virus Induces Apoptosis by Depletion of Drosophila Inhibitor-of-Apoptosis Protein DIAP1. J. Virol. 2008, 82, 1378–1388. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.M.; May, R.M. Coevolution of Hosts and Parasites. Parasitology 1982, 85, 411–426. [Google Scholar] [CrossRef] [PubMed]
- Flor, H.H. The complementary genetic system in flax and flax rust. Adv. Gen. 1956, 8, 29–54. [Google Scholar]
- Grosberg, R.K.; Hart, M.W. Mate selection and the evolution of highly polymorphic self/nonself recognition genes. Science 2000, 289, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Sessions, O.M.; Barrows, N.J.; Souza-Neto, J.A.; Robinson, T.J.; Hershey, C.L.; Rodgers, M.A.; Ramirez, J.L.; Dimopoulos, G.; Yang, P.L.; Pearson, J.L.; et al. Discovery of insect and human dengue virus host factors. Nature 2009, 458, 1047–1050. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Longdon, B.; Jiggins, F.M. Vertically transmitted viral endosymbionts of insects: Do sigma viruses walk alone? Proc. R. Soc. B Biol. Sci. 2012, 279, 3889–3898. [Google Scholar] [CrossRef] [PubMed]
- Fleuriet, A. Presence of the Hereditary Rhabdovirus Sigma and Polymorphism for a Gene for Resistance to This Virus in Natural Populations of Drosophila Melanogaster. Evolution 1976, 30, 735–739. [Google Scholar] [CrossRef] [PubMed]
- Wayne, M.L.; Contamine, D.; Kreitman, M. Molecular population genetics of ref(2)P, a locus which confers viral resistance in Drosophila. Mol. Biol. Evol. 1996, 13, 191–199. [Google Scholar] [CrossRef] [PubMed]
- Avila, A.; Silverman, N.; Diaz-Meco, M.T.; Moscat, J. The Drosophila Atypical Protein Kinase C-Ref(2)P Complex Constitutes a Conserved Module for Signaling in the Toll Pathway. Mol. Cell. Biol. 2002, 22, 8787–8795. [Google Scholar] [CrossRef] [PubMed]
- Carré-Mlouka, A.; Gaumer, S.; Gay, P.; Petitjean, A.M.; Coulondre, C.; Dru, P.; Bras, F.; Dezélée, S.; Contamine, D. Control of sigma virus multiplication by the ref(2)P gene of Drosophila melanogaster: An in vivo study of the PB1 domain of Ref(2)P. Genetics 2007, 176, 409–419. [Google Scholar] [CrossRef] [PubMed]
- Nezis, I.P.; Simonsen, A.; Sagona, A.P.; Finley, K.; Gaumer, S.; Contamine, D.; Rusten, T.E.; Stenmark, H.; Brech, A. Ref(2)P, the Drosophila melanogaster homologue of mammalian p62, is required for the formation of protein aggregates in adult brain. J. Cell Biol. 2008, 180, 1065–1071. [Google Scholar] [CrossRef] [PubMed]
- Contamine, D.; Petitjean, A.M.; Ashburner, M. Genetic resistance to viral infection: The molecular cloning of a Drosophila gene that restricts infection by the rhabdovirus sigma. Genetics 1989, 123, 525–533. [Google Scholar] [PubMed]
- Gay, P. Drosophila genes which intervene in multiplication of sigma virus (author’s transl). Mol. Gen. Genet. 1978, 159, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Wilfert, L.; Jiggins, F.M. Disease association mapping in Drosophila can be replicated in the wild. Biol. Lett. 2010, 6, 666–668. [Google Scholar] [CrossRef] [PubMed]
- Magwire, M.M.; Fabian, D.K.; Schweyen, H.; Cao, C.; Longdon, B.; Bayer, F.; Jiggins, F.M. Genome-Wide Association Studies Reveal a Simple Genetic Basis of Resistance to Naturally Coevolving Viruses in Drosophila melanogaster. PLoS Genet. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Magwire, M.M.; Bayer, F.; Webster, C.L.; Cao, C.; Jiggins, F.M. Successive increases in the resistance of Drosophila to viral infection through a transposon insertion followed by a duplication. PLoS Genet. 2011, 7, e1002337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cogni, R.; Cao, C.; Day, J.P.; Bridson, C.; Jiggins, F.M. The genetic architecture of resistance to virus infection in Drosophila. Mol. Ecol. 2016, 25, 5228–5241. [Google Scholar] [CrossRef] [PubMed]
- Martins, N.E.; Faria, V.G.; Nolte, V.; Schlotterer, C.; Teixeira, L.; Sucena, E.; Magalhaes, S. Host adaptation to viruses relies on few genes with different cross-resistance properties. Proc. Natl. Acad. Sci. USA 2014, 111, 5938–5943. [Google Scholar] [CrossRef] [PubMed]
- Cao, C.; Cogni, R.; Barbier, V.; Jiggins, F.M. Complex coding and regulatory polymorphisms in a restriction factor determine the susceptibility of Drosophila to viral infection. Genetics 2017, 206, 2159–2173. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Quillery, E.; Noël, V.; Richardson, J.H.; Jarman, R.G.; Thomas, W.; Chevillon, C.; Scott, T.W. Specificity of resistance to dengue virus isolates is associated with genotypes of the mosquito antiviral gene Dicer-2 Specificity of resistance to dengue virus isolates is associated with genotypes of the mosquito antiviral gene Dicer-2. Proc. R. Soc. B 2013, 280, 20122437. [Google Scholar] [CrossRef] [PubMed]
- Fansiri, T.; Fontaine, A.; Diancourt, L.; Caro, V.; Thaisomboonsuk, B.; Richardson, J.H.; Jarman, R.G.; Ponlawat, A.; Lambrechts, L. Genetic Mapping of Specific Interactions between Aedes aegypti Mosquitoes and Dengue Viruses. PLoS Genet. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Bosio, C.F.; Fulton, R.E.; Salasek, M.L.; Beaty, B.J.; Black, W.C., IV. Quantitative trait loci that control vector competence for dengue-2 virus in the mosquito Aedes aegypti. Genetics 2000, 156, 687–698. [Google Scholar] [PubMed]
- Graham, D.H.; Holmes, J.L.; Beaty, B.J.; Black, W.C., IV. Quantitative trait loci conditioning transovarial transmission of La Crosse virus in the eastern treehole mosquito, Ochlerotatus triseriatus. Insect Mol. Biol. 2003, 12, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Aminetzach, Y.T.; Macpherson, J.M.; Petrov, D.A. Evolution: Pesticide resistance via transposition-mediated adaptive gene truncation in Drosophila. Science 2005, 309, 764–767. [Google Scholar] [CrossRef] [PubMed]
- Lentz, T.L.; Burrage, T.G.; Smith, L.; Tignor, G.H. The acetylcholine receptor as a cellular receptor for rabies virus. Yale J. Biol. Med. 1983, 56, 315–322. [Google Scholar] [PubMed]
- Yu, J.H.; Yang, W.-H.; Gulick, T.; Bloch, K.D.; Bloch, D.B. Ge-1 is a central component of the mammalian cytoplasmic mRNA processing body. RNA 2005, 11, 1795–1802. [Google Scholar] [CrossRef] [PubMed]
- Jinek, M.; Eulalio, A.; Lingel, A.; Helms, S.; Conti, E.; Izaurralde, E. The C-terminal region of Ge-1 presents conserved structural features required for P-body localization. RNA 2008, 14, 1991–1998. [Google Scholar] [CrossRef] [PubMed]
- Brun, P.; Plus, N. The viruses of Drosophila. In The Genetics and Biology of Drosophila; Academic Press: London, UK, 1980; pp. 625–702. [Google Scholar]
- Bangham, J.; Knott, S.A.; Kim, K.W.; Young, R.S.; Jiggins, F.M. Genetic variation affecting host-parasite interactions: Major-effect quantitative trait loci affect the transmission of sigma virus in Drosophila melanogaster. Mol. Ecol. 2008, 17, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Fricke, R.; Gohl, C.; Dharmalingam, E.; Grevelhörster, A.; Zahedi, B.; Harden, N.; Kessels, M.; Qualmann, B.; Bogdan, S. Drosophila Cip4/Toca-1 Integrates Membrane Trafficking and Actin Dynamics through WASP and SCAR/WAVE. Curr. Biol. 2009, 19, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Duffy, S.; Shackelton, L.A.; Holmes, E.C. Rates of evolutionary change in viruses: Patterns and determinants. Nat. Rev. Genet. 2008, 9, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Ebert, D. Host-parasite coevolution: Insights from the Daphnia-parasite model system. Curr. Opin. Microbiol. 2008, 11, 290–301. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Dudas, G. The genetics of host-virus coevolution in invertebrates. Curr. Opin. Virol. 2014, 8, 73–78. [Google Scholar] [CrossRef] [PubMed]
- Dru, P.; Bras, F.; Dezelee, S.; Gay, P.; Petitjean, A.M.; Pierre-Deneubourg, A.; Teninges, D.; Contamine, D. Unusual variability of the Drosophila melanogaster ref(2)P protein which controls the multiplication of sigma rhabdovirus. Genetics 1993, 133, 943–954. [Google Scholar] [PubMed]
- Fleuriet, A. Perpetuation of the hereditary sigma virus in populations of its host, Drosophila melanogaster. Geographical analysis of correlated polymorphisms. Genetica 1986, 70, 167–177. [Google Scholar] [CrossRef]
- Fleuriet, A.; Sperlich, D. Evolution of the Drosophila melanogaster-sigma virus system in a natural population from Tuebingen. Theor. Appl. Genet. 1992, 85, 186–189. [Google Scholar] [CrossRef] [PubMed]
- Fleuriet, A.; Periquet, G. Evolution of the Drosophila melanogaster-sigma virus system in natural populations from Languedoc (southern France). Arch. Virol. 1993, 129, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Bangham, J.; Obbard, D.J.; Kim, K.-W.; Haddrill, P.R.; Jiggins, F.M. The age and evolution of an antiviral resistance mutation in Drosophila melanogaster. Proc. Biol. Sci. 2007, 274, 2027–2034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wilfert, L.; Jiggins, F.M. The dynamics of reciprocal selective sweeps of host resistance and a parasite counter-adaptation in Drosophila. Evolution 2013, 67, 761–773. [Google Scholar] [CrossRef] [PubMed]
- Unckless, R.L.; Howick, V.M.; Lazzaro, B.P. Convergent Balancing Selection on an Antimicrobial Peptide in Drosophila. Curr. Biol. 2016, 26, 257–262. [Google Scholar] [CrossRef] [PubMed]
- Unckless, R.L.; Lazzaro, B.P. The potential for adaptive maintenance of diversity in insect antimicrobial peptides. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2016, 371, 20150291. [Google Scholar] [CrossRef] [PubMed]
- Hain, D.; Bettencourt, B.R.; Okamura, K.; Csorba, T.; Meyer, W.; Jin, Z.; Biggerstaff, J.; Siomi, H.; Hutvagner, G.; Lai, E.C.; et al. Natural Variation of the Amino-Terminal Glutamine-Rich Domain in Drosophila Argonaute2 Is Not Associated with Developmental Defects. PLoS ONE 2010, 5. [Google Scholar] [CrossRef] [PubMed]
- Palmer, W.H.; Obbard, D.J. Variation and Evolution in the Glutamine-Rich Repeat Region of Drosophila Argonaute-2. G3 Genes Genomes Genet. 2016, 6, 2563–2572. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Jiggins, F.M.; Halligan, D.L.; Little, T.J. Natural selection drives extremely rapid evolution in antiviral RNAi genes. Curr. Biol. 2006, 16, 580–585. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Welch, J.J.; Kim, K.W.; Jiggins, F.M. Quantifying adaptive evolution in the Drosophila immune system. PLoS Genet. 2009, 5, e1000698. [Google Scholar] [CrossRef] [PubMed]
- Palmer, W.H.; Hadfield, J.D.; Obbard, D.J. RNA Interference Pathways Display High Rates of Adaptive Protein Evolution in Multiple Invertebrates. Genetics 2018. [Google Scholar] [CrossRef] [PubMed]
- Kolaczkowski, B.; Hupalo, D.N.; Kern, A.D. Recurrent adaptation in RNA interference genes across the Drosophila phylogeny. Mol. Biol. Evol. 2011, 28, 1033–1042. [Google Scholar] [CrossRef] [PubMed]
- Obbard, D.J.; Jiggins, F.M.; Bradshaw, N.J.; Little, T.J. Recent and recurrent selective sweeps of the antiviral RNAi gene argonaute-2 in three species of Drosophila. Mol. Biol. Evol. 2011, 28, 1043–1056. [Google Scholar] [CrossRef] [PubMed]
- Lewis, S.H.; Webster, C.L.; Salmela, H.; Obbard, D.J. Repeated duplication of Argonaute2 is associated with strong selection and testis specialization in Drosophila. Genetics 2016, 204, 757–769. [Google Scholar] [CrossRef] [PubMed]
- Schlenke, T.A.; Begun, D.J. Natural selection drives Drosophila immune system evolution. Genetics 2003, 164, 1471–1480. [Google Scholar] [PubMed]
- Sackton, T.B.; Lazzaro, B.P.; Schlenke, T.A.; Evans, J.D.; Hultmark, D.; Clark, A.G. Dynamic evolution of the innate immune system in Drosophila. Nat. Genet. 2007, 39, 1461–1468. [Google Scholar] [CrossRef] [PubMed]
- Han, M.; Qin, S.; Song, X.; Li, Y.; Jin, P.; Chen, L.; Ma, F. Evolutionary rate patterns of genes involved in the Drosophila Toll and Imd signaling pathway. BMC Evol. Biol. 2013, 13, 245. [Google Scholar] [CrossRef] [PubMed]
- Juneja, P.; Lazzaro, B.P. Haplotype structure and expression divergence at the Drosophila cellular immune gene eater. Mol. Biol. Evol. 2010, 27, 2284–2299. [Google Scholar] [CrossRef] [PubMed]
- Nene, V.; Wortman, J.R.; Lawson, D.; Haas, B.; Kodira, C.; Tu, Z.; Loftus, B.; Xi, Z.; Megy, K.; Grabherr, M.; et al. Genome sequence of Aedes aegypti, a major arbovirus vector. Science 2007, 316, 1718–1723. [Google Scholar] [CrossRef] [PubMed]
- Bosio, C.; Beaty, B.; Black, W.C. Quantitative genetics of vector competence for dengue-2 virus in Aedes aegypti. Am. J. Trop. Med. Hyg. 1998, 59, 965–970. [Google Scholar] [CrossRef] [PubMed]
- Bennett, K.E.; Beaty, B.J.; Black, W.C. Selection of D2S3, an Aedes aegypti (Diptera: Culicidae) Strain with High Oral Susceptibility to Dengue 2 Virus and D2MEB, a Strain with a Midgut Barrier to Dengue 2 Escape. J. Med. Entomol. 2005, 42, 110–119. [Google Scholar] [CrossRef]
- Gomez-Machorro, C.; Bennett, K.E.; Munoz, M.D.L.; Black, W.C., IV. Quantitative trait loci affecting dengue midgut infection barriers in an advanced intercross line of Aedes aegypti. Insect Mol. Biol. 2004, 13, 637–648. [Google Scholar] [CrossRef] [PubMed]
- Noriega, F.G.; Pennington, J.E.; Barillas-Mury, C.; Wang, X.Y.; Wells, M.A. Aedes aegypti midgut early trypsin is post-transcriptionally regulated by blood feeding. Insect Mol. Biol. 1996, 5, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Gorrochotegui-Escalante, N.; Lozano-Fuentes, S.; Bennett, K.E.; Molina-Cruz, A.; Beaty, B.J.; Black, W.C., IV. Association mapping of segregating sites in the early trypsin gene and susceptibility to dengue-2 virus in the mosquito Aedes aegypti. Insect Biochem. Mol. Biol. 2005, 35, 771–788. [Google Scholar] [CrossRef] [PubMed]
- Molina-Cruz, A.; Gupta, L.; Richardson, J.; Bennett, K.; Black, W.C., IV; Barillas-Mury, C. Effect of mosquito midgut trypsin activity on dengue-2 virus infection and dissemination in Aedes aegypti. Am. J. Trop. Med. Hyg. 2005, 72, 631–637. [Google Scholar] [PubMed]
- Brackney, D.E.; Foy, B.D.; Olson, K.E. The effects of midgut serine proteases on dengue virus type 2 infectivity of Aedes aegypti. Am. J. Trop. Med. Hyg. 2008, 79, 267–274. [Google Scholar] [PubMed]
- Barón, O.L.; Ursic-Bedoya, R.J.; Lowenberger, C.A.; Ocampo, C.B. Differential Gene Expression from Midguts of Refractory and Susceptible Lines of the Mosquito, Aedes aegypti, Infected with Dengue-2 Virus. J. Insect Sci. 2010, 10, 1–23. [Google Scholar] [CrossRef] [PubMed]
- Anderson, J.R.; Schneider, J.R.; Grimstad, P.R.; Severson, D.W. Quantitative genetics of vector competence for La Crosse virus and body size in Ochlerotatus hendersoni and Ochlerotatus triseriatus interspecific hybrids. Genetics 2005, 169, 1529–1539. [Google Scholar] [CrossRef] [PubMed]
- Borucki, M.K.; Kempf, B.J.; Blitvich, B.J.; Blair, C.D.; Beaty, B.J. La Crosse virus: Replication in vertebrate and invertebrate hosts. Microbes Infect. 2002, 4, 341–350. [Google Scholar] [CrossRef]
- Tran, K.T.; Vazeille-Falcoz, M.; Mousson, L.; Tran, H.H.; Rodhain, F.; Ngugen, T.H.; Failloux, A.B. Aedes aegypti in Ho Chi Minh City (Viet Nam): Susceptibility to dengue 2 virus and genetic differentiation. Trans. R. Soc. Trop. Med. Hyg. 1999, 93, 581–586. [Google Scholar] [PubMed]
- Mousson, L.; Vazeille, M.; Chawprom, S.; Prajakwong, S.; Rodhain, F.; Failloux, A.-B. Genetic structure of Aedes aegypti populations in Chiang Mai (Thailand) and relation with dengue transmission. Trop. Med. Int. Health 2002, 7, 865–872. [Google Scholar] [CrossRef] [PubMed]
- Bernhardt, S.A.; Simmons, M.P.; Olson, K.E.; Beaty, B.J.; Blair, C.D.; Black, W.C. Rapid Intraspecific Evolution of miRNA and siRNA Genes in the Mosquito Aedes aegypti. PLoS ONE 2012, 7. [Google Scholar] [CrossRef] [PubMed]
- Halbach, R.; Junglen, S.; van Rij, R.P. Mosquito-specific and mosquito-borne viruses: Evolution, infection, and host defense. Curr. Opin. Insect Sci. 2017, 22, 16–27. [Google Scholar] [CrossRef] [PubMed]
- Behura, S.K.; Gomez-Machorro, C.; Harker, B.W.; deBruyn, B.; Lovin, D.D.; Hemme, R.R.; Mori, A.; Romero-Severson, J.; Severson, D.W. Global cross-talk of genes of the mosquito Aedes aegypti in response to dengue virus infection. PLoS Negl. Trop. Dis. 2011, 5. [Google Scholar] [CrossRef] [PubMed]
- Ocampo, C.B.; Caicedo, P.A.; Jaramillo, G.; Ursic Bedoya, R.; Baron, O.; Serrato, I.M.; Cooper, D.M.; Lowenberger, C. Differential Expression of Apoptosis Related Genes in Selected Strains of Aedes aegypti with Different Susceptibilities to Dengue Virus. PLoS ONE 2013, 8. [Google Scholar] [CrossRef] [PubMed]
- Clem, R.J. Arboviruses and apoptosis: The role of cell death in determining vector competence. J. Gen. Virol. 2016, 97, 1033–1036. [Google Scholar] [CrossRef] [PubMed]
- Vaidyanathan, R.; Scott, T.W. Apoptosis in mosquito midgut epithelia associated with West Nile virus infection. Apoptosis 2006, 11, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Girard, Y.A.; Schneider, B.S.; McGee, C.E.; Wen, J.; Han, V.C.; Popov, V.; Mason, P.W.; Higgs, S. Salivary gland morphology and virus transmission during long-term cytopathologic West Nile virus infection in Culex mosquitoes. Am. J. Trop. Med. Hyg. 2007, 76, 118–128. [Google Scholar] [PubMed]
- Mercado-Curiel, R.F.; Black, W.C., IV; Mũoz, M.D.L. A dengue receptor as possible genetic marker of vector competence in Aedes aegypti. BMC Microbiol. 2008, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Sim, S.; Jupatanakul, N.; Ramirez, J.L.; Kang, S.; Romero-Vivas, C.M.; Mohammed, H.; Dimopoulos, G. Transcriptomic Profiling of Diverse Aedes aegypti Strains Reveals Increased Basal-level Immune Activation in Dengue Virus-refractory Populations and Identifies Novel Virus-vector Molecular Interactions. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Angleró-Rodríguez, Y.I.; Talyuli, O.A.; Blumberg, B.J.; Kang, S.; Demby, C.; Shields, A.; Carlson, J.; Jupatanakul, N.; Dimopoulos, G. An Aedes aegypti-associated fungus increases susceptibility to dengue virus by modulating gut trypsin activity. eLife 2017, 6, e28844. [Google Scholar] [CrossRef] [PubMed]
- Nishi, T.; Forgac, M. The vacuolar (H+)-ATPases—Nature’s most versatile proton pumps. Nat. Rev. Mol. Cell Biol. 2002, 3, 94–103. [Google Scholar] [CrossRef] [PubMed]
- Smit, J.M.; Moesker, B.; Rodenhuis-Zybert, I.; Wilschut, J. Flavivirus cell entry and membrane fusion. Viruses 2011, 3, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Brownlie, J.C.; O’Neill, S.L.; Johnson, K.N. Wolbachia and virus protection in insects. Science 2008, 322, 702. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, L.; Ferreira, Á.; Ashburner, M. The bacterial symbiont Wolbachia induces resistance to RNA viral infections in Drosophila melanogaster. PLoS Biol. 2008, 6, 2753–2763. [Google Scholar] [CrossRef] [PubMed]
- Serbus, L.R.; Casper-Lindley, C.; Landmann, F.; Sullivan, W. The Genetics and Cell Biology of Wolbachia-Host Interactions. Annu. Rev. Genet. 2008, 42, 683–707. [Google Scholar] [CrossRef] [PubMed]
- Osborne, S.E.; Leong, Y.S.; O’Neill, S.L.; Johnson, K.N. Variation in antiviral protection mediated by different Wolbachia strains in Drosophila simulans. PLoS Pathog. 2009, 5, e1000656. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Bian, G.; Pan, X.; Xi, Z. Wolbachia induces density-dependent inhibition to dengue virus in mosquito cells. PLoS Negl. Trop. Dis. 2012, 6, e1754. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Longdon, B.; Bauer, S.; Chan, Y.S.; Miller, W.J.; Bourtzis, K.; Teixeira, L.; Jiggins, F.M. Symbionts Commonly Provide Broad Spectrum Resistance to Viruses in Insects: A Comparative Analysis of Wolbachia Strains. PLoS Pathog. 2014, 10, e1004369. [Google Scholar] [CrossRef] [PubMed]
- Martinez, J.; Tolosana, I.; Ok, S.; Smith, S.; Snoeck, K.; Day, J.P.; Jiggins, F.M. Symbiont strain is the main determinant of variation in Wolbachia-mediated protection against viruses across Drosophila species. Mol. Ecol. 2017, 26, 4072–4084. [Google Scholar] [CrossRef] [PubMed]
- Terradas, G.; McGraw, E.A. Wolbachia-mediated virus blocking in the mosquito vector Aedes aegypti. Curr. Opin. Insect Sci. 2017, 22, 37–44. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rancès, E.; Hedges, L.M.; Gofton, A.W.; Johnson, K.N.; O’Neill, S.L.; McGraw, E.A. Dietary Cholesterol Modulates Pathogen Blocking by Wolbachia. PLoS Pathog. 2013, 9, e1003459. [Google Scholar] [CrossRef] [PubMed]
- Caragata, E.P.; Rezende, F.O.; Simões, T.C.; Moreira, L.A. Diet-Induced Nutritional Stress and Pathogen Interference in Wolbachia-Infected Aedes aegypti. PLoS Negl. Trop. Dis. 2016, 10, e0005158. [Google Scholar] [CrossRef] [PubMed]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia induces reactive oxygen species (ROS)-dependent activation of the Toll pathway to control dengue virus in the mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hussain, M.; O’Neill, S.L.; Asgari, S. Wolbachia uses a host microRNA to regulate transcripts of a methyltransferase, contributing to dengue virus inhibition in Aedes aegypti. Proc. Natl. Acad. Sci. USA 2013, 110, 10276–10281. [Google Scholar] [CrossRef] [PubMed]
- Wong, Z.S.; Brownlie, J.C.; Johnson, K.N. Oxidative stress correlates with Wolbachia-mediated antiviral protection in Wolbachia-Drosophila associations. Appl. Environ. Microbiol. 2015, 81, 3001–3005. [Google Scholar] [CrossRef] [PubMed]
- Bhattacharya, T.; Newton, I.L.G.; Hardy, R.W. Wolbachia elevates host methyltransferase expression to block an RNA virus early during infection. PLoS Pathog. 2017, 13, e1006427. [Google Scholar] [CrossRef] [PubMed]
- Terradas, G.; Joubert, D.A.; McGraw, E.A. The RNAi pathway plays a small part in Wolbachia-mediated blocking of dengue virus in mosquito cells. Sci. Rep. 2017, 7, 43847. [Google Scholar] [CrossRef] [PubMed]
- Hedges, L.M.; Yamada, R.; O’Neill, S.L.; Johnson, K.N. The small interfering RNA pathway is not essential for wolbachia-mediated antiviral protection in Drosophila melanogaster. Appl. Environ. Microbiol. 2012, 78, 6773–6776. [Google Scholar] [CrossRef] [PubMed]
- Rancès, E.; Ye, Y.H.; Woolfit, M.; McGraw, E.A.; O’Neill, S.L. The relative importance of innate immune priming in Wolbachia-mediated dengue interference. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Rances, E.; Johnson, T.K.; Popovici, J.; Iturbe-Ormaetxe, I.; Zakir, T.; Warr, C.G.; O’Neill, S.L. The Toll and Imd Pathways Are Not Required for Wolbachia-Mediated Dengue Virus Interference. J. Virol. 2013, 87, 11945–11949. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Marialva, M.S.P.; Yamada, R.; O’Neill, S.L.; Teixeira, L. High anti-viral protection without immune upregulation after interspecies Wolbachia transfer. PLoS ONE 2014, 9, e99025. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Marialva, M.S.P.; Esteves, S.S.; Weinert, L.A.; Martinez, J.; Jiggins, F.M.; Teixeira, L. Wolbachia Variants Induce Differential Protection to Viruses in Drosophila melanogaster: A Phenotypic and Phylogenomic Analysis. PLoS Genet. 2013, 9, e1003896. [Google Scholar] [CrossRef] [PubMed]
- Min, K.-T.; Benzer, S. Wolbachia, normally a symbiont of Drosophila, can be virulent, causing degeneration and early death. Proc. Natl. Acad. Sci. USA 1997, 94, 10792–10796. [Google Scholar] [CrossRef] [PubMed]
- Woolfit, M.; Iturbe-Ormaetxe, I.; Brownlie, J.C.; Walker, T.; Riegler, M.; Seleznev, A.; Popovici, J.; Rancès, E.; Wee, B.A.; Pavlides, J.; et al. Genomic evolution of the pathogenic Wolbachia strain, wMelPop. Genome Biol. Evol. 2013, 5, 2189–2204. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Teixeira, L. Mutualism Breakdown by Amplification of Wolbachia Genes. PLoS Biol. 2015, 13, e1002065. [Google Scholar] [CrossRef] [PubMed]
- Chrostek, E.; Teixeira, L. Within host selection for faster replicating bacterial symbionts. bioRxiv 2017, 222240. [Google Scholar] [CrossRef]
- Rohrscheib, C.E.; Frentiu, F.D.; Horn, E.; Ritchie, F.K.; van Swinderen, B.; Weible, M.W.; O’Neill, S.L.; Brownlie, J.C. Intensity of Mutualism Breakdown Is Determined by Temperature Not Amplification of Wolbachia Genes. PLoS Pathog. 2016, 12, e1005888. [Google Scholar] [CrossRef] [PubMed]
- McMeniman, C.J.; Lane, A.M.; Fong, A.W.C.; Voronin, D.A.; Iturbe-Ormaetxe, I.; Yamada, R.; McGraw, E.A.; O’Neill, S.L. Host adaptation of a Wolbachia strain after long-term serial passage in mosquito cell lines. Appl. Environ. Microbiol. 2008, 74, 6963–6969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engelstädter, J.; Hurst, G.D.D. The Ecology and Evolution of Microbes that Manipulate Host Reproduction. Annu. Rev. Ecol. Evol. Syst. 2009, 40, 127–149. [Google Scholar] [CrossRef]
- Martinez, J.; Ok, S.; Smith, S.; Snoeck, K.; Day, J.P.; Jiggins, F.M. Should Symbionts Be Nice or Selfish? Antiviral Effects of Wolbachia Are Costly but Reproductive Parasitism Is Not. PLoS Pathog. 2015, 11, e1005021. [Google Scholar] [CrossRef] [PubMed]
- Riegler, M.; Sidhu, M.; Miller, W.J.; O’Neill, S.L. Evidence for a global Wolbachia replacement in Drosophila melanogaster. Curr. Biol. 2005, 15, 1428–1433. [Google Scholar] [CrossRef] [PubMed]
- Nunes, M.D.S.; Nolte, V.; Schlötterer, C. Nonrandom Wolbachia infection status of Drosophila melanogaster strains with different mtDNA haplotypes. Mol. Biol. Evol. 2008, 25, 2493–2498. [Google Scholar] [CrossRef] [PubMed]
- Richardson, M.F.; Weinert, L.A.; Welch, J.J.; Linheiro, R.S.; Magwire, M.M.; Jiggins, F.M.; Bergman, C.M. Population Genomics of the Wolbachia Endosymbiont in Drosophila melanogaster. PLoS Genet. 2012, 8, e1003129. [Google Scholar] [CrossRef] [PubMed]
- Turelli, M.; Hoffmann, A.A. Rapid spread of an inherited incompatibility factor in California Drosophila. Nature 1991, 353, 440–442. [Google Scholar] [CrossRef] [PubMed]
- James, A.C.; Ballard, J.W.O. Expression of cytoplasmic incompatibility in Drosophila simulans and its impact on infection frequencies and distribution of Wolbachia pipientis. Evolution 2000, 54, 1661–1672. [Google Scholar] [CrossRef] [PubMed]
- Kriesner, P.; Hoffmann, A.A.; Lee, S.F.; Turelli, M.; Weeks, A.R. Rapid Sequential Spread of Two Wolbachia Variants in Drosophila simulans. PLoS Pathog. 2013, 9, e1003607. [Google Scholar] [CrossRef] [PubMed]
- Mousson, L.; Zouache, K.; Arias-Goeta, C.; Raquin, V.; Mavingui, P.; Failloux, A.-B. The Native Wolbachia Symbionts Limit Transmission of Dengue Virus in Aedes albopictus. PLoS Negl. Trop. Dis. 2012, 6, e1989. [Google Scholar] [CrossRef] [PubMed]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia Symbiont in Aedes aegypti Limits Infection with Dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M. The spread of Wolbachia through mosquito populations. PLoS Biol. 2017, 15, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Montgomery, B.L.; Popovici, J.; Iturbe-Ormaetxe, I.; Johnson, P.H.; Muzzi, F.; Greenfield, M.; Durkan, M.; Leong, Y.S.; Dong, Y.; et al. Successful establishment of Wolbachia in Aedes populations to suppress dengue transmission. Nature 2011, 476, 454–459. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Iturbe-Ormaetxe, I.; Callahan, A.G.; Phillips, B.L.; Billington, K.; Axford, J.K.; Montgomery, B.; Turley, A.P.; O’Neill, S.L. Stability of the wMel Wolbachia Infection following Invasion into Aedes aegypti Populations. PLoS Negl. Trop. Dis. 2014, 8. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Tran, B.C.N.; Le, N.T.H.; Luong, T.T.H.; Nguyen, T.T.; Nguyen, P.T.; Nguyen, C.V.V.; Nguyen, H.T.C.; Vu, T.T.; Vo, L.T.; et al. Field- and clinically derived estimates of Wolbachia -mediated blocking of dengue virus transmission potential in Aedes aegypti mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, 361–366. [Google Scholar] [CrossRef] [PubMed]
- Robinson, C.M.; Pfeiffer, J.K. Viruses and the Microbiota. Annu. Rev. Virol. 2014, 1, 55–69. [Google Scholar] [CrossRef] [PubMed]
- Jakubowska, A.K.; Vogel, H.; Herrero, S. Increase in Gut Microbiota after Immune Suppression in Baculovirus-infected Larvae. PLoS Pathog. 2013, 9, e1003379. [Google Scholar] [CrossRef] [PubMed]
- Sansone, C.; Cohen, J.; Gold, B.; Zhu, W.; Misic, A.M.; Beiting, D.P.; Winter, S.E.; Cherry, S. Aging-associated dysbiosis increases susceptibility to enteric viral infection in Drosophila. bioRxiv 2017, 156455. [Google Scholar] [CrossRef]
- Buchon, N.; Broderick, N.A.; Chakrabarti, S.; Lemaitre, B. Invasive and indigenous microbiota impact intestinal stem cell activity through multiple pathways in Drosophila. Genes Dev. 2009, 23, 2333–2344. [Google Scholar] [CrossRef] [PubMed]
- Corby-Harris, V.; Pontaroli, A.C.; Shimkets, L.J.; Bennetzen, J.L.; Habel, K.E.; Promislow, D.E.L. Geographical distribution and diversity of bacteria associated with natural populations of Drosophila melanogaster. Appl. Environ. Microbiol. 2007, 73, 3470–3479. [Google Scholar] [CrossRef] [PubMed]
- Chandler, J.A.; Lang, J.; Bhatnagar, S.; Eisen, J.A.; Kopp, A. Bacterial communities of diverse Drosophila species: Ecological context of a host-microbe model system. PLoS Genet. 2011, 7, e1002272. [Google Scholar] [CrossRef] [PubMed]
- Van Tol, S.; Dimopoulos, G. Influences of the Mosquito Microbiota on Vector Competence. Adv. Insect Physiol. 2016, 51, 243–291. [Google Scholar]
- Ramirez, J.L.; Souza-Neto, J.; Cosme, R.T.; Rovira, J.; Ortiz, A.; Pascale, J.M.; Dimopoulos, G. Reciprocal tripartite interactions between the Aedes aegypti midgut microbiota, innate immune system and dengue virus influences vector competence. PLoS Negl. Trop. Dis. 2012, 6, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, J.L.; Short, S.M.; Bahia, A.C.; Saraiva, R.G.; Dong, Y.; Kang, S.; Tripathi, A.; Mlambo, G.; Dimopoulos, G. Chromobacterium Csp_P Reduces Malaria and Dengue Infection in Vector Mosquitoes and Has Entomopathogenic and In Vitro Anti-pathogen Activities. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Apte-Deshpande, A.; Paingankar, M.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera a midgut inhabitant of Aedes aegypti mosquito enhances its susceptibility to dengue-2 virus. PLoS ONE 2012, 7, e40401. [Google Scholar] [CrossRef] [PubMed]
- Apte-Deshpande, A.D.; Paingankar, M.S.; Gokhale, M.D.; Deobagkar, D.N. Serratia odorifera mediated enhancement in susceptibility of Aedes aegypti for chikungunya virus. Indian J. Med. Res. 2014, 139, 762–768. [Google Scholar] [PubMed]
- Muturi, E.J.; Bara, J.J.; Rooney, A.P.; Hansen, A.K. Midgut fungal and bacterial microbiota of Aedes triseriatus and Aedes japonicus shift in response to La Crosse virus infection. Mol. Ecol. 2016, 25, 4075–4090. [Google Scholar] [CrossRef] [PubMed]
- Joyce, J.D.; Nogueira, J.R.; Bales, A.A.; Pittman, K.E.; Anderson, J.R. Interactions Between La Crosse Virus and Bacteria Isolated From the Digestive Tract of Aedes albopictus (Diptera: Culicidae). J. Med. Entomol. 2011, 48, 389–394. [Google Scholar] [CrossRef] [PubMed]
- Shirasu-Hiza, M.M.; Dionne, M.S.; Pham, L.N.; Ayres, J.S.; Schneider, D.S. Interactions between circadian rhythm and immunity in Drosophila melanogaster. Curr. Biol. 2007, 17, R353–R355. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.E.; Edery, I. Circadian Regulation in the Ability of Drosophila to Combat Pathogenic Infections. Curr. Biol. 2008, 18, 195–199. [Google Scholar] [CrossRef] [PubMed]
- Stone, E.F.; Fulton, B.O.; Ayres, J.S.; Pham, L.N.; Ziauddin, J.; Shirasu-Hiza, M.M. The circadian clock protein timeless regulates phagocytosis of bacteria in Drosophila. PLoS Pathog. 2012, 8, e1002445. [Google Scholar] [CrossRef] [PubMed]
- Siva-Jothy, M.T.; Thompson, J.J.W. Short-term nutrient deprivation affects immune function. Physiol. Entomol. 2002, 27, 206–212. [Google Scholar] [CrossRef]
- Fellous, S.; Lazzaro, B.P. Larval food quality affects adult (but not larval) immune gene expression independent of effects on general condition. Mol. Ecol. 2010, 19, 1462–1468. [Google Scholar] [CrossRef] [PubMed]
- Povey, S.; Cotter, S.C.; Simpson, S.J.; Wilson, K. Dynamics of macronutrient self-medication and illness-induced anorexia in virally infected insects. J. Anim. Ecol. 2014, 83, 245–255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Unckless, R.L.; Rottschaefer, S.M.; Lazzaro, B.P. The Complex Contributions of Genetics and Nutrition to Immunity in Drosophila melanogaster. PLoS Genet. 2015, 11, e1005030. [Google Scholar] [CrossRef] [PubMed]
- Muturi, E.J.; Kim, C.H.; Alto, B.W.; Berenbaum, M.R.; Schuler, M.A. Larval environmental stress alters Aedes aegypti competence for Sindbis virus. Trop. Med. Int. Health 2011, 16, 955–964. [Google Scholar] [CrossRef] [PubMed]
- Horne, K.M.E.; Vanlandingham, D.L. Bunyavirus-vector interactions. Viruses 2014, 6, 4373–4397. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Reiskind, M.H.; Lounibos, L.P. Size Alters Susceptibility of Vectors to Dengue Virus Infection and Dissemination. Am. J. Trop. Med. Hyg. 2008, 79, 688–695. [Google Scholar] [PubMed]
- Schneider, J.R.; Mori, A.; Romero-Severson, J.; Chadee, D.D.; Severson, D.W. Investigations of dengue-2 susceptibility and body size among Aedes aegypti populations. Med. Vet. Entomol. 2007, 21, 370–376. [Google Scholar] [CrossRef] [PubMed]
- Bara, J.J.; Parker, A.T.; Muturi, E.J. Comparative Susceptibility of Ochlerotatus japonicus, Ochlerotatus triseriatus, Aedes albopictus, and Aedes aegypti (Diptera: Culicidae) to La crosse virus. J. Med. Entomol. 2016, 53, 1415–1421. [Google Scholar] [CrossRef] [PubMed]
- Alto, B.W.; Lounibos, L.P.; Mores, C.N.; Reiskind, M.H. Larval competition alters susceptibility of adult Aedes mosquitoes to dengue infection. Proc. R. Soc. B Biol. Sci. 2008, 275, 463–471. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.S.; Alcalay, Y.; Lovin, D.D.; Cunningham, J.M.; Eng, M.W.; Chadee, D.D.; Severson, D.W. Larval stress alters dengue virus susceptibility in Aedes aegypti (L.) adult females. Acta Trop. 2017, 174, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Kay, B.H.; Fanning, I.D.; Mottram, P. Rearing temperature influences flavivirus vector competence of mosquitoes. Med. Vet. Entomol. 1989, 3, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Dohm, D.J.; O’Guinn, M.L.; Turell, M.J. Effect of environmental temperature on the ability of Culex pipiens (Diptera: Culicidae) to transmit West Nile virus. J. Med. Entomol. 2002, 39, 221–225. [Google Scholar] [CrossRef] [PubMed]
- Richards, S.L.; Mores, C.N.; Lord, C.C.; Tabachnick, W.J. Impact of Extrinsic Incubation Temperature and Virus Exposure on Vector Competence of Culex pipiens quinquefasciatus Say (Diptera: Culicidae) for West Nile Virus. Vector-Borne Zoonotic Dis. 2007, 7, 629–636. [Google Scholar] [CrossRef] [PubMed]
- Kilpatrick, A.M.; Meola, M.A.; Moudy, R.M.; Kramer, L.D. Temperature, viral genetics, and the transmission of West Nile virus by Culex pipiens mosquitoes. PLoS Pathog. 2008, 4. [Google Scholar] [CrossRef] [PubMed]
- Rohani, A.; Wong, Y.C.; Zamre, I.; Lee, H.L.; Zurainee, M.N. The effect of extrinsic incubation temperature on development of dengue serotype 2 and 4 viruses in Aedes aegypti (L.). Southeast Asian J. Trop. Med. Public Health 2009, 40, 942–950. [Google Scholar] [PubMed]
- Xiao, F.-Z.; Zhang, Y.; Deng, Y.-Q.; He, S.; Xie, H.-G.; Zhou, X.-N.; Yan, Y.-S. The effect of temperature on the extrinsic incubation period and infection rate of dengue virus serotype 2 infection in Aedes albopictus. Arch. Virol. 2014, 159, 3053–3057. [Google Scholar] [CrossRef] [PubMed]
- Kramer, L.D.; Hardy, J.L.; Presser, S.B. Effect of temperature of extrinsic incubation on the vector competence of Culex tarsalis for western equine encephalomyelitis virus. Am. J. Trop. Med. Hyg. 1983, 32, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Westbrook, C.J.; Reiskind, M.H.; Pesko, K.N.; Greene, K.E.; Lounibos, L.P. Larval environmental temperature and the susceptibility of Aedes albopictus Skuse (Diptera: Culicidae) to Chikungunya virus. Vector Borne Zoonotic Dis. 2010, 10, 241–247. [Google Scholar] [CrossRef] [PubMed]
- Adelman, Z.N.; Anderson, M.A.E.; Wiley, M.R.; Murreddu, M.G.; Samuel, G.H.; Morazzani, E.M.; Myles, K.M. Cooler Temperatures Destabilize RNA Interference and Increase Susceptibility of Disease Vector Mosquitoes to Viral Infection. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Samuel, G.H.; Adelman, Z.N.; Myles, K.M. Temperature-dependent effects on the replication and transmission of arthropod-borne viruses in their insect hosts. Curr. Opin. Insect Sci. 2016, 16, 108–113. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Paaijmans, K.P.; Fansiri, T.; Carrington, L.B.; Kramer, L.D.; Thomas, M.B.; Scott, T.W. Impact of daily temperature fluctuations on dengue virus transmission by Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Fluctuations at a Low Mean Temperature Accelerate Dengue Virus Transmission by Aedes aegypti. PLoS Negl. Trop. Dis. 2013, 7. [Google Scholar] [CrossRef] [PubMed]
- Carrington, L.B.; Seifert, S.N.; Armijos, M.V.; Lambrechts, L.; Scott, T.W. Reduction of Aedes aegypti vector competence for dengue virus under large temperature fluctuations. Am. J. Trop. Med. Hyg. 2013, 88, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Zuk, M. The Sicker Sex. PLoS Pathog. 2009, 5, e1000267. [Google Scholar] [CrossRef] [PubMed]
- Duneau, D.F.; Kondolf, H.C.; Im, J.H.; Ortiz, G.A.; Chow, C.; Fox, M.A.; Eugénio, A.T.; Revah, J.; Buchon, N.; Lazzaro, B.P. The Toll pathway underlies host sexual dimorphism in resistance to both Gram-negative and Gram-positive bacteria in mated Drosophila. BMC Biol. 2017, 15, 124. [Google Scholar] [CrossRef] [PubMed]
- Gupta, V.; Stewart, C.O.; Rund, S.S.C.; Monteith, K.; Vale, P.F. Costs and benefits of sublethal Drosophila C virus infection. J. Evol. Biol. 2017, 30, 1325–1335. [Google Scholar] [CrossRef] [PubMed]
- Rolff, J. Bateman’s principle and immunity. Proc. R. Soc. B Biol. Sci. 2002, 269, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Zuk, M.; Stoehr, A.M. Immune Defense and Host Life History. Am. Nat. 2002, 160, S9–S22. [Google Scholar] [CrossRef] [PubMed]
- Stoehr, A.M.; Kokko, H. Sexual dimorphism in immunocompetence: What does life-history theory predict? Behav. Ecol. 2006, 17, 751–756. [Google Scholar] [CrossRef]
- Nunn, C.L.; Lindenfors, P.; Pursall, E.R.; Rolff, J. On sexual dimorphism in immune function. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 61–69. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Regan, J.C.; Khericha, M.; Dobson, A.J.; Bolukbasi, E.; Rattanavirotkul, N.; Partridge, L. Sex difference in pathology of the ageing gut mediates the greater response of female lifespan to dietary restriction. eLife 2016, 5, e10956. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zipperlen, P.; Kubli, E. Drosophila Sex-Peptide Stimulates Female Innate Immune System after Mating via the Toll and Imd Pathways. Curr. Biol. 2005, 15, 1690–1694. [Google Scholar] [CrossRef] [PubMed]
- Fedorka, K.M.; Linder, J.E.; Winterhalter, W.; Promislow, D. Post-mating disparity between potential and realized immune response in Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2007, 274, 1211–1217. [Google Scholar] [CrossRef] [PubMed]
- Short, S.M.; Lazzaro, B.P. Female and male genetic contributions to post-mating immune defence in female Drosophila melanogaster. Proc. R. Soc. B Biol. Sci. 2010, 277, 3649–3657. [Google Scholar] [CrossRef] [PubMed]
- Schwenke, R.A.; Lazzaro, B.P. Juvenile Hormone Suppresses Resistance to Infection in Mated Female Drosophila melanogaster. Curr. Biol. 2017, 27, 596–601. [Google Scholar] [CrossRef] [PubMed]
- Lawniczak, M.K.N.; Barnes, A.I.; Linklater, J.R.; Boone, J.M.; Wigby, S.; Chapman, T. Mating and immunity in invertebrates. Trends Ecol. Evol. 2007, 22, 48–55. [Google Scholar] [CrossRef] [PubMed]
- Rolff, J.; Siva-Jothy, M.T. Copulation corrupts immunity: A mechanism for a cost of mating in insects. Proc. Natl. Acad. Sci. USA 2002, 99, 9916–9918. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Heyland, A.; Rus, F.; Porpiglia, E.; Sherlock, C.; Yamamoto, R.; Garbuzov, A.; Palli, S.R.; Tatar, M.; Silverman, N. Hormonal regulation of the humoral innate immune response in Drosophila melanogaster. J. Exp. Biol. 2008, 211, 2712–2724. [Google Scholar] [CrossRef] [PubMed]
- Flatt, T.; Tu, M.-P.; Tatar, M. Hormonal pleiotropy and the juvenile hormone regulation of Drosophila development and life history. BioEssays 2005, 27, 999–1010. [Google Scholar] [CrossRef] [PubMed]
- Moshitzky, P.; Fleischmann, I.; Chaimov, N.; Saudan, P.; Klauser, S.; Kubli, E.; Applebaum, S.W. Sex-peptide activates juvenile hormone biosynthesis in the Drosophila melanogaster corpus allatum. Arch. Insect Biochem. Physiol. 1996, 32, 363–374. [Google Scholar] [CrossRef]
- Burand, J.P.; Kim, W.; Afonso, C.L.; Tulman, E.R.; Kutish, G.F.; Lu, Z.; Rock, D.L. Analysis of the genome of the sexually transmitted insect virus Helicoverpa zea nudivirus 2. Viruses 2012, 4, 28–61. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, J.; Nakai, M.; Shinoda, T. A virus carries a gene encoding juvenile hormone acid methyltransferase, a key regulatory enzyme in insect metamorphosis. Sci. Rep. 2017, 7, 13522. [Google Scholar] [CrossRef] [PubMed]
- Palmer, W.H.; Medd, N.; Obbard, D.J. University of Edinburgh, Edinburgh, UK. PCR survey for viruses in Drosophila melanogaster. Unpublished work. 2016. [Google Scholar]
- Chen, Y.; Zhao, Y.; Hammond, J.; Hsu, H.T.; Evans, J.; Feldlaufer, M. Multiple virus infections in the honey bee and genome divergence of honey bee viruses. J. Invertebr. Pathol. 2004, 87, 84–93. [Google Scholar] [CrossRef] [PubMed]
- Washburn, J.O.; Haas-Stapleton, E.J.; Tan, F.F.; Beckage, N.E.; Volkman, L.E. Co-infection of Manduca sexta larvae with polydnavirus from Cotesia congregata increases susceptibility to fatal infection by Autographa californica M Nucleopolyhedrovirus. J. Insect Physiol. 2000, 46, 179–190. [Google Scholar] [CrossRef]
- Berry, B.; Deddouche, S.; Kirschner, D.; Imler, J.-L.; Antoniewski, C. Viral Suppressors of RNA Silencing Hinder Exogenous and Endogenous Small RNA Pathways in Drosophila. PLoS ONE 2009, 4, e5866. [Google Scholar] [CrossRef] [PubMed]
- Kuwata, R.; Isawa, H.; Hoshino, K.; Sasaki, T.; Kobayashi, M.; Maeda, K.; Sawabe, K. Analysis of mosquito-borne flavivirus superinfection in culex tritaeniorhynchus (Diptera: Culicidae) cells persistently infected with culex flavivirus (Flaviviridae). J. Med. Entomol. 2015, 52, 222–229. [Google Scholar] [CrossRef] [PubMed]
- Myers, R.M.; Carey, D.E. Concurrent Isolation from Patient of Two Arboviruses, Chikungunya and Dengue Type 2. Science 1967, 157, 1307–1308. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.F.; Su, C.L.; Shu, P.Y.; Yang, C.F.; Liao, T.L.; Cheng, C.H.; Hu, H.C.; Huang, J.H. Concurrent isolation of chikungunya virus and dengue virus from a patient with coinfection resulting from a trip to Singapore. J. Clin. Microbiol. 2010, 48, 4586–4589. [Google Scholar] [CrossRef] [PubMed]
- Parreira, R.; Centeno-Lima, S.; Lopes, A.; Portugal-Calisto, D.; Constantino, A.; Nina, J. Dengue virus serotype 4 and Chikungunya virus coinfection in a traveller returning from Luanda, Angola, January 2014. Eurosurveillance 2014, 19, 6–9. [Google Scholar] [CrossRef]
- Villamil-Gómez, W.E.; González-Camargo, O.; Rodriguez-Ayubi, J.; Zapata-Serpa, D.; Rodriguez-Morales, A.J. Dengue, chikungunya and Zika co-infection in a patient from Colombia. J. Infect. Public Health 2016, 9, 684–686. [Google Scholar] [CrossRef] [PubMed]
- Vazeille, M.; Mousson, L.; Martin, E.; Failloux, A.B. Orally co-infected Aedes albopictus from La Reunion Island, Indian Ocean, can deliver both dengue and chikungunya infectious viral particles in their saliva. PLoS Negl. Trop. Dis. 2010, 4, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Rückert, C.; Weger-Lucarelli, J.; Garcia-Luna, S.M.; Young, M.C.; Byas, A.D.; Murrieta, R.A.; Fauver, J.R.; Ebel, G.D. Impact of simultaneous exposure to arboviruses on infection and transmission by Aedes aegypti mosquitoes. Nat. Commun. 2017, 8, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Göertz, G.P.; Vogels, C.B.F.; Geertsema, C.; Koenraadt, C.J.M.; Pijlman, G.P. Mosquito co-infection with Zika and chikungunya virus allows simultaneous transmission without affecting vector competence of Aedes aegypti. PLoS Negl. Trop. Dis. 2017, 11, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Blitvich, B.J.; Firth, A.E. Insect-specific flaviviruses: A systematic review of their discovery, host range, mode of transmission, superinfection exclusion potential and genomic organization. Viruses 2015, 7, 1927–1959. [Google Scholar] [CrossRef] [PubMed]
- Hobson-Peters, J.; Yam, A.W.Y.; Lu, J.W.F.; Setoh, Y.X.; May, F.J.; Kurucz, N.; Walsh, S.; Prow, N.A.; Davis, S.S.; Weir, R.; et al. A New Insect-Specific Flavivirus from Northern Australia Suppresses Replication of West Nile Virus and Murray Valley Encephalitis Virus in Co-infected Mosquito Cells. PLoS ONE 2013, 8, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Hall-Mendelin, S.; McLean, B.J.; Bielefeldt-Ohmann, H.; Hobson-Peters, J.; Hall, R.A.; Van Den Hurk, A.F. The insect-specific Palm Creek virus modulates West Nile virus infection in and transmission by Australian mosquitoes. Parasites Vectors 2016, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kenney, J.L.; Solberg, O.D.; Langevin, S.A.; Brault, A.C. Characterization of a novel insect-specific flavivirus from Brazil: Potential for inhibition of infection of arthropod cells with medically important flaviviruses. J. Gen. Virol. 2014, 95, 2796–2808. [Google Scholar] [CrossRef] [PubMed]
- Goenaga, S.; Kenney, J.L.; Duggal, N.K.; Delorey, M.; Ebel, G.D.; Zhang, B.; Levis, S.C.; Enria, D.A.; Brault, A.C. Potential for co-infection of a mosquito-specific flavivirus, nhumirim virus, to block west nile virus transmission in mosquitoes. Viruses 2015, 7, 5801–5812. [Google Scholar] [CrossRef] [PubMed]
- Nasar, F.; Erasmus, J.H.; Haddow, A.D.; Tesh, R.B.; Weaver, S.C. Eilat virus induces both homologous and heterologous interference. Virology 2015, 484, 51–58. [Google Scholar] [CrossRef] [PubMed]
- Bolling, B.G.; Olea-Popelka, F.J.; Eisen, L.; Moore, C.G.; Blair, C.D. Transmission dynamics of an insect-specific flavivirus in a naturally infected Culex pipiens laboratory colony and effects of co-infection on vector competence for West Nile virus. Virology 2012, 427, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Longdon, B.; Cao, C.; Martinez, J.; Jiggins, F.M. Previous Exposure to an RNA Virus Does Not Protect against Subsequent Infection in Drosophila melanogaster. PLoS ONE 2013, 8, e73833. [Google Scholar] [CrossRef] [PubMed]
- Tidbury, H.J.; Pedersen, A.B.; Boots, M. Within and transgenerational immune priming in an insect to a DNA virus. Proc. R. Soc. B Biol. Sci. 2011, 278, 871–876. [Google Scholar] [CrossRef] [PubMed]
- Crochu, S.; Cook, S.; Attoui, H.; Charrel, R.N.; De Chesse, R.; Belhouchet, M.; Lemasson, J.J.; de Micco, P.; de Lamballerie, X. Sequences of flavivirus-related RNA viruses persist in DNA form integrated in the genome of Aedes spp. mosquitoes. J. Gen. Virol. 2004, 85, 1971–1980. [Google Scholar] [CrossRef] [PubMed]
- Nag, D.K.; Brecher, M.; Kramer, L.D. DNA forms of arboviral RNA genomes are generated following infection in mosquito cell cultures. Virology 2016, 498, 164–171. [Google Scholar] [CrossRef] [PubMed]
- Katzourakis, A.; Gifford, R.J. Endogenous viral elements in animal genomes. PLoS Genet. 2010, 6. [Google Scholar] [CrossRef]
- Fort, P.; Albertini, A.; Van-Hua, A.; Berthomieu, A.; Roche, S.; Delsuc, F.; Pasteur, N.; Capy, P.; Gaudin, Y.; Weill, M. Fossil rhabdoviral sequences integrated into arthropod genomes: Ontogeny, evolution, and potential functionality. Mol. Biol. Evol. 2012, 29, 381–390. [Google Scholar] [CrossRef] [PubMed]
- Suzuki, Y.; Frangeul, L.; Dickson, L.B.; Blanc, H.; Verdier, Y.; Vinh, J.; Lambrechts, L.; Saleh, M.-C. Uncovering the Repertoire of Endogenous Flaviviral Elements in Aedes Mosquito Genomes. J. Virol. 2017, 91, e00571-17. [Google Scholar] [CrossRef] [PubMed]
- Whitfield, Z.J.; Dolan, P.T.; Kunitomi, M.; Tassetto, M.; Seetin, M.G.; Oh, S.; Heiner, C.; Paxinos, E.; Andino, R. The Diversity, Structure, and Function of Heritable Adaptive Immunity Sequences in the Aedes aegypti Genome. Curr. Biol. 2017, 27, 3511–3519. [Google Scholar] [CrossRef] [PubMed]
- Palatini, U.; Miesen, P.; Carballar-Lejarazu, R.; Ometto, L.; Rizzo, E.; Tu, Z.; van Rij, R.P.; Bonizzoni, M. Comparative genomics shows that viral integrations are abundant and express piRNAs in the arboviral vectors Aedes aegypti and Aedes albopictus. BMC Genom. 2017, 18, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Miesen, P.; Joosten, J.; van Rij, R.P. PIWIs Go Viral: Arbovirus-Derived piRNAs in Vector Mosquitoes. PLoS Pathog. 2016, 12, e1006017. [Google Scholar] [CrossRef] [PubMed]
- Dudchenko, O.; Batra, S.S.; Omer, A.D.; Nyquist, S.K.; Hoeger, M.; Durand, N.C.; Shamim, M.S.; Machol, I.; Lander, E.S.; Aiden, A.P.; et al. De novo assembly of the Aedes aegypti genome using Hi-C yields chromosome-length scaffolds. Science 2017, 356, 92–95. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Antiviral innate immune pathways in dipteran insects. Toll pathway: Detection of pathogen-associated molecular patterns (PAMPs) by pathogen recognition-receptors leads to the proteolytic maturation of Spätzle, which binds to and activates Toll (dotted arrows). Activated Toll recruits adapter proteins MyD88, Tube and Pelle which targets Cactus for proteasomal degradation via phosphorylation. Cactus degradation releases the transcription factor Dorsal (Rel1 in mosquitoes) or Dif (Dorsal-related immune factor) when activated in response to bacterial infection. These translocate to the nucleus and activates the transcription of Toll pathway-regulated genes (e.g., cecropin, defensin and drosomycin). IMD pathway: Gram-negative PAMPs (e.g., peptidoglycan) bind to peptidoglycan recognition protein, PGRP-LC (or PGRP-LE) and signal through the adapter molecules IMD (Immune Deficiency) and dFADD (Drosophila Fas-associated death domain). This stimulates the caspase Dredd (Death-related ced-3/Nedd2-like protein), dTAK1 (Drosophila transforming growth factor β-activated kinase 1), and dTAK1 adapter protein dTAB2 (TAK1-binding protein 2). These proteins signal through the JNK pathway, and activate the IκB kinases kenny and ird-5, which phosphorylate the C-terminal tail of the transcription factor Relish (Rel2 in mosquitoes), leading to its subsequent activation via proteolytic cleavage by Dredd. Activated Relish translocates to the nucleus and drives the expression of genes regulated by the IMD-pathway (e.g., diptericin, attacin and metchnikowin). JAK-STAT pathway: Virus infection, possibly through induction of stress or cellular damage, triggers the activation of the JAK (Janus kinase)-STAT (signal transducers and activators of transcription) pathway, which begins with the binding of a cytokine of the unpaired (upd) family to the dimeric receptor, domeless. Subsequently, the receptor-associated JAK-tyrosine kinase hopscotch phosphorylates the cytoplasmic tail of domeless, leading to the recruitment of Stat92E. After Jak-mediated phosphorylation, Stat92E proteins dimerize and shuttle to the nucleus to activate the transcription of genes such as vir-1, TotM, DVRF1 and DVRF2. RNAi pathway: Double-stranded RNA derived from virus replication intermediates are recognized and processed by Dicer-2 into 21 nt small-interfering RNAs (siRNAs), which are then loaded onto an Argonaute-2 (Ago2)-containing RNA-induced silencing complex (RISC). This complex degrades one of the two strands and uses the other strand as a guide RNA to target complementary viral sequences. Dicer-2 can also activate the expression of the cytokine Vago through an unknown pathway (visualized as ‘?’ in the figure). RNA decay pathways: Single-stranded viral mRNA can be targeted in the 3′–5′ direction through the RNA exosome degradation pathway. 5′–3′ degradation occurs through decapping enzymes Dcp1 and Dcp2 and the RNA exonuclease Xrn1. Autophagy: Some viruses can bind to the transmembrane receptor Toll-7, resulting in the induction of autophagy. This is most likely in an indirect manner by negatively regulating the PI3K (phosphatidylinositol 3-kinase)-Akt pathway.
Figure 1.
Antiviral innate immune pathways in dipteran insects. Toll pathway: Detection of pathogen-associated molecular patterns (PAMPs) by pathogen recognition-receptors leads to the proteolytic maturation of Spätzle, which binds to and activates Toll (dotted arrows). Activated Toll recruits adapter proteins MyD88, Tube and Pelle which targets Cactus for proteasomal degradation via phosphorylation. Cactus degradation releases the transcription factor Dorsal (Rel1 in mosquitoes) or Dif (Dorsal-related immune factor) when activated in response to bacterial infection. These translocate to the nucleus and activates the transcription of Toll pathway-regulated genes (e.g., cecropin, defensin and drosomycin). IMD pathway: Gram-negative PAMPs (e.g., peptidoglycan) bind to peptidoglycan recognition protein, PGRP-LC (or PGRP-LE) and signal through the adapter molecules IMD (Immune Deficiency) and dFADD (Drosophila Fas-associated death domain). This stimulates the caspase Dredd (Death-related ced-3/Nedd2-like protein), dTAK1 (Drosophila transforming growth factor β-activated kinase 1), and dTAK1 adapter protein dTAB2 (TAK1-binding protein 2). These proteins signal through the JNK pathway, and activate the IκB kinases kenny and ird-5, which phosphorylate the C-terminal tail of the transcription factor Relish (Rel2 in mosquitoes), leading to its subsequent activation via proteolytic cleavage by Dredd. Activated Relish translocates to the nucleus and drives the expression of genes regulated by the IMD-pathway (e.g., diptericin, attacin and metchnikowin). JAK-STAT pathway: Virus infection, possibly through induction of stress or cellular damage, triggers the activation of the JAK (Janus kinase)-STAT (signal transducers and activators of transcription) pathway, which begins with the binding of a cytokine of the unpaired (upd) family to the dimeric receptor, domeless. Subsequently, the receptor-associated JAK-tyrosine kinase hopscotch phosphorylates the cytoplasmic tail of domeless, leading to the recruitment of Stat92E. After Jak-mediated phosphorylation, Stat92E proteins dimerize and shuttle to the nucleus to activate the transcription of genes such as vir-1, TotM, DVRF1 and DVRF2. RNAi pathway: Double-stranded RNA derived from virus replication intermediates are recognized and processed by Dicer-2 into 21 nt small-interfering RNAs (siRNAs), which are then loaded onto an Argonaute-2 (Ago2)-containing RNA-induced silencing complex (RISC). This complex degrades one of the two strands and uses the other strand as a guide RNA to target complementary viral sequences. Dicer-2 can also activate the expression of the cytokine Vago through an unknown pathway (visualized as ‘?’ in the figure). RNA decay pathways: Single-stranded viral mRNA can be targeted in the 3′–5′ direction through the RNA exosome degradation pathway. 5′–3′ degradation occurs through decapping enzymes Dcp1 and Dcp2 and the RNA exonuclease Xrn1. Autophagy: Some viruses can bind to the transmembrane receptor Toll-7, resulting in the induction of autophagy. This is most likely in an indirect manner by negatively regulating the PI3K (phosphatidylinositol 3-kinase)-Akt pathway.


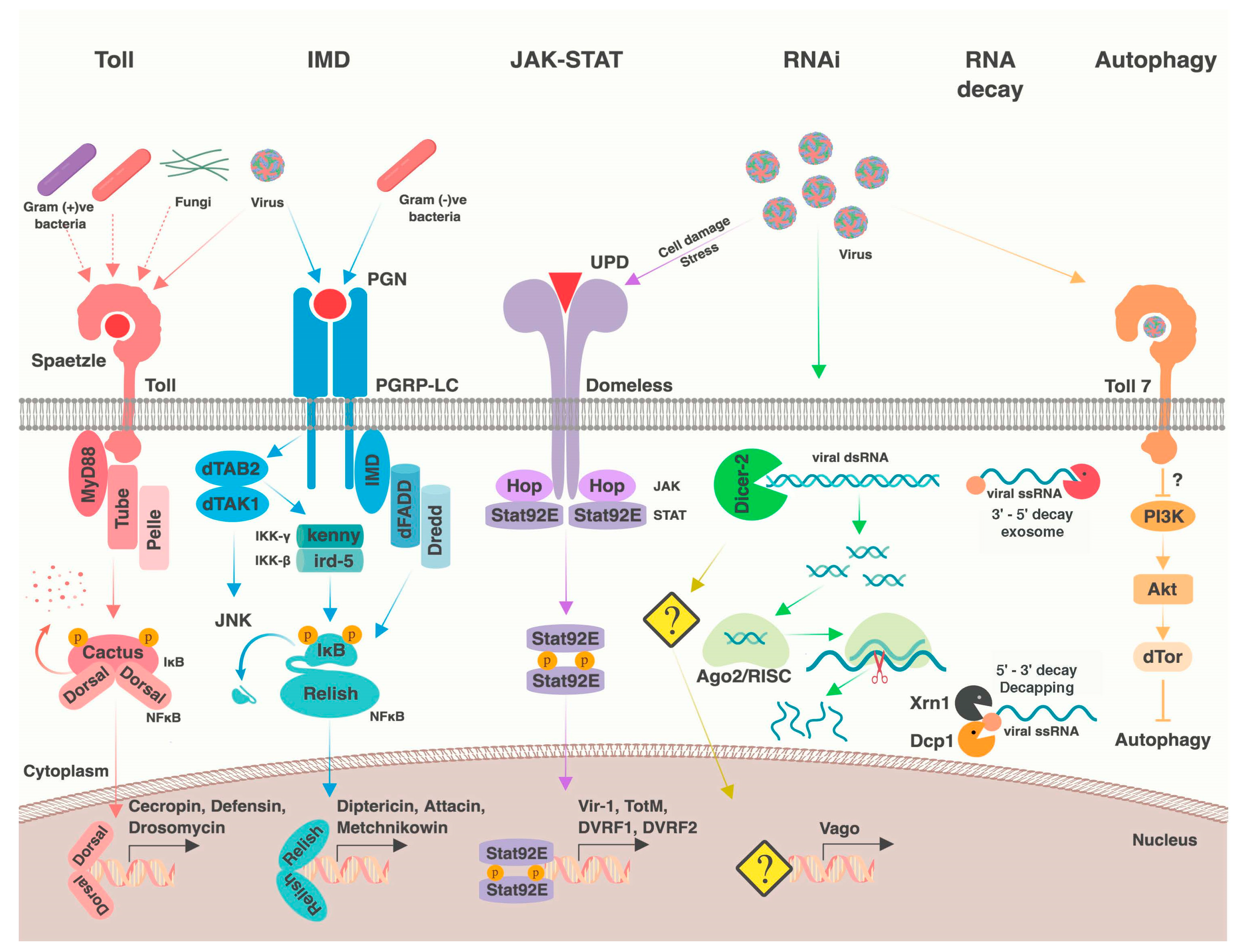{kind=link}
Table 1.
Viruses discussed in this review.
| Virus Name | Abbreviation | Genome | Family | Host Restriction |
|---|---|---|---|---|
| Cricket paralysis virus | CrPV | (+) ssRNA | Dicistroviridae | Insect-specific |
| Drosophila C virus | DCV | (+) ssRNA | Dicistroviridae | Insect-specific |
| Culex flavivirus | CxFV | (+) ssRNA | Flaviviridae | Insect-specific |
| Nhumirim virus | NHUMV | (+) ssRNA | Flaviviridae | Insect-specific |
| Palm Creek virus | PCV | (+) ssRNA | Flaviviridae | Insect-specific |
| Flock House virus | FHV | (+) ssRNA | Nodaviridae | Insect-specific |
| Eilat virus | EILV | (+) ssRNA | Togaviridae | Insect-specific |
| Nora virus | Nora | (+) ssRNA | Unclassified Picornavirales | Insect-specific |
| Drosophila melanogaster sigma virus | DmelSV | (−) ssRNA | Rhabdoviridae | Insect-specific |
| Drosophila X virus | DXV | dsRNA | Birnaviridae | Insect-specific |
| Invertebrate iridescent virus 6 | IIV6 | dsDNA | Iridoviridae | Insect-specific |
| Kallithea virus | Kallithea | dsDNA | Nudiviridae | Insect-specific |
| Dengue virus | DENV | (+) ssRNA | Flaviviridae | Arbovirus |
| Japanese encephalitis virus | JEV | (+) ssRNA | Flaviviridae | Arbovirus |
| Murray Valley encephalitis virus | MVEV | (+) ssRNA | Flaviviridae | Arbovirus |
| St Louis encephalitis virus | SLEV | (+) ssRNA | Flaviviridae | Arbovirus |
| West Nile virus | WNV | (+) ssRNA | Flaviviridae | Arbovirus |
| Yellow fever virus | YFV | (+) ssRNA | Flaviviridae | Arbovirus |
| Zika virus | ZIKV | (+) ssRNA | Flaviviridae | Arbovirus |
| Chikungunya virus | CHIKV | (+) ssRNA | Togaviridae | Arbovirus |
| O’nyong’nyong virus | ONNV | (+) ssRNA | Togaviridae | Arbovirus |
| Semliki Forest virus | SFV | (+) ssRNA | Togaviridae | Arbovirus |
| Sindbis virus | SINV | (+) ssRNA | Togaviridae | Arbovirus |
| Venezuelan equine encephalitis virus | VEEV | (+) ssRNA | Togaviridae | Arbovirus |
| Western equine encephalitis virus | WEEV | (+) ssRNA | Togaviridae | Arbovirus |
| La Crosse encephalitis virus | LACV | (−) ssRNA | Peribunyaviridae | Arbovirus |
| Rift Valley fever virus | RVFV | (−) ssRNA | Phenuiviridae | Arbovirus |
| Vesicular stomatitis virus | VSV | (−) ssRNA | Rhabdoviridae | Arbovirus |
Table 2.
Genetic variants associated with viral resistance.
| Host Factor | Virus | Host | Associated Mutation | Phenotype (Homozygotes) | Population Frequency (Resistant Allele) | Related Processes | Methodology | References |
|---|---|---|---|---|---|---|---|---|
| ref(2)P | DmelSV | Dmel | NS polymorphism | 24% reduction in infection rate | 24% | Autophagy | Genetic mapping, mutagenesis, GWAS | [127,128,129,130] |
| CHKov1/CHKov2 | DmelSV | Dmel | TE insertion, rearrangement | 52% reduction in infection rate | TE: 83%; rearrangement <0.5% | Predicted acetylcholine esterase | Genetic mapping, GWAS | [130,131] |
| Ge-1 | DmelSV | Dmel | 26 aa deletion | 97% reduction in infection rate | 1% | RNA decay | Genetic mapping | [40] |
| ref(1)H QTL | DmelSV | Dmel | Unknown | Unknown | Unknown | Unknown | Genetic mapping | [128] |
| ref(3)O QTL | DmelSV | Dmel | Unknown | Unknown | Unknown | Unknown | Genetic mapping | [128] |
| ref(3)V QTL | DmelSV | Dmel | Unknown | Unknown | Unknown | Unknown | Genetic mapping | [128] |
| X13 QTL | DmelSV | Dmel | Unknown | 38% reduction in infection rate | Unknown | Unknown | Genetic mapping | [132] |
| X65 QTL | DmelSV | Dmel | Unknown | 13% reduction in infection rate | Unknown | Unknown | Genetic mapping | [132] |
| 2R70 QTL | DmelSV | Dmel | Unknown | 11% reduction in infection rate | Unknown | Unknown | Genetic mapping | [132] |
| 3R64 QTL | DmelSV | Dmel | Unknown | 12% reduction in infection rate | Unknown | Unknown | Genetic mapping | [132] |
| pastrel | DCV | Dmel | 7 alleles: cis-regulatory, structural, NS variants | 16%, 57%, or 80% increase in survival | NS: 7–33%, Structural variants: 4–51% | Unknown | GWAS, experimental evolution | [130,133,134] |
| 2R69 QTL | DCV | Dmel | Unknown | 2.5 day increase in survival time | Unknown | Unknown | Recombinant inbred line | [132] |
| 2L18 QTL | DCV | Dmel | Unknown | 0.75 day increase in survival time | Unknown | Unknown | Recombinant inbred line | [132] |
| APC7 | DCV | Dmel | Synonymous polymorphism | 95% increase in survival | 3% | Cell cycle | GWAS | [130] |
| Ubc-E2H | DCV | Dmel | Intronic polymorphism | Not reported | 27% (lab-maintained population) | Predicted ubiquitin ligase | Experimental evolution | [133] |
| Cip4 | Kallithea | Dmel | NS polymorphism | 27% increase in survival | 77% | Membrane trafficking | GWAS | [107] |
| Dicer-2 | DENV-1 | Aaeg | Unknown | Explains 17.8% of viral dissemination | Unknown | RNAi | Genetic mapping | [135] |
| 71CGT1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains up to 7.6% variation in MIB | Unknown | Unknown | Genetic mapping | [136] |
| 335CGA1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 8.1% variation in MIB and ≤21.4% in dissemination | Unknown | Unknown | Genetic mapping | [136] |
| 88CA1/88GAA1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 12% variation in MIB and ≤75.6% in titer | Unknown | Unknown | Genetic mapping | [136] |
| 301CT1/301ACG1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 11.4% variation in MIB | Unknown | Unknown | Genetic mapping | [136] |
| B19 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 6.2% variation in MIB | Unknown | Unknown | Genetic mapping | [136] |
| 69TGA1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 22.6% variation in dissemination and ≤ 8.9% in titer | Unknown | Unknown | Genetic mapping | [136] |
| 201AAT1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 15.2% variation in titer | Unknown | Unknown | Genetic mapping | [136] |
| 470CT2/470AG1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 12.3% variation in titer | Unknown | Unknown | Genetic mapping | [136] |
| 17ATA1 QTL | DENV-1/3 | Aaeg | Unknown | QTL explains ≤ 13.7% variation in titer | Unknown | Unknown | Genetic mapping | [136] |
| early trypsin QTL | DENV-2 | Aaeg | Unknown | 86% decrease in infection dissemination | Unknown | Blood meal digestion | Genetic mapping | [137] |
| B18.621 QTL | DENV-2 | Aaeg | Unknown | 87% decrease in infection dissemination | Unknown | Unknown | Genetic mapping | [137] |
| B20.392 QTL | LACV | Otri | Unknown | Transovarial transmission rate increased from 0 to 60% | Unknown | Unknown | Genetic mapping | [138] |
| C01.385/C13.5573 QTL | LACV | Otri | 2 linked QTL with unknown mutations | Transovarial transmission rate increased from 0 to 60% | Unknown | Unknown | Genetic mapping | [138] |
Dmel, D. melanogaster; Aaeg, Ae. aegypti; Otri, O. triseriatus; aa, amino acid; MIB, midgut infection barrier; NS, nonsynonymous; S, synonymous; TE, transposable element; GWAS, genome-wide association study.
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Palmer, W.H.; Varghese, F.S.; Van Rij, R.P. Natural Variation in Resistance to Virus Infection in Dipteran Insects. Viruses 2018, 10, 118. https://0-doi-org.brum.beds.ac.uk/10.3390/v10030118
AMA Style
Palmer WH, Varghese FS, Van Rij RP. Natural Variation in Resistance to Virus Infection in Dipteran Insects. Viruses. 2018; 10(3):118. https://0-doi-org.brum.beds.ac.uk/10.3390/v10030118
Chicago/Turabian StylePalmer, William H., Finny S. Varghese, and Ronald P. Van Rij. 2018. "Natural Variation in Resistance to Virus Infection in Dipteran Insects" Viruses 10, no. 3: 118. https://0-doi-org.brum.beds.ac.uk/10.3390/v10030118
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.