Synergistic Viral Replication of Marek’s Disease Virus and Avian Leukosis Virus Subgroup J is Responsible for the Enhanced Pathogenicity in the Superinfection of Chickens


Abstract
:1. Introduction
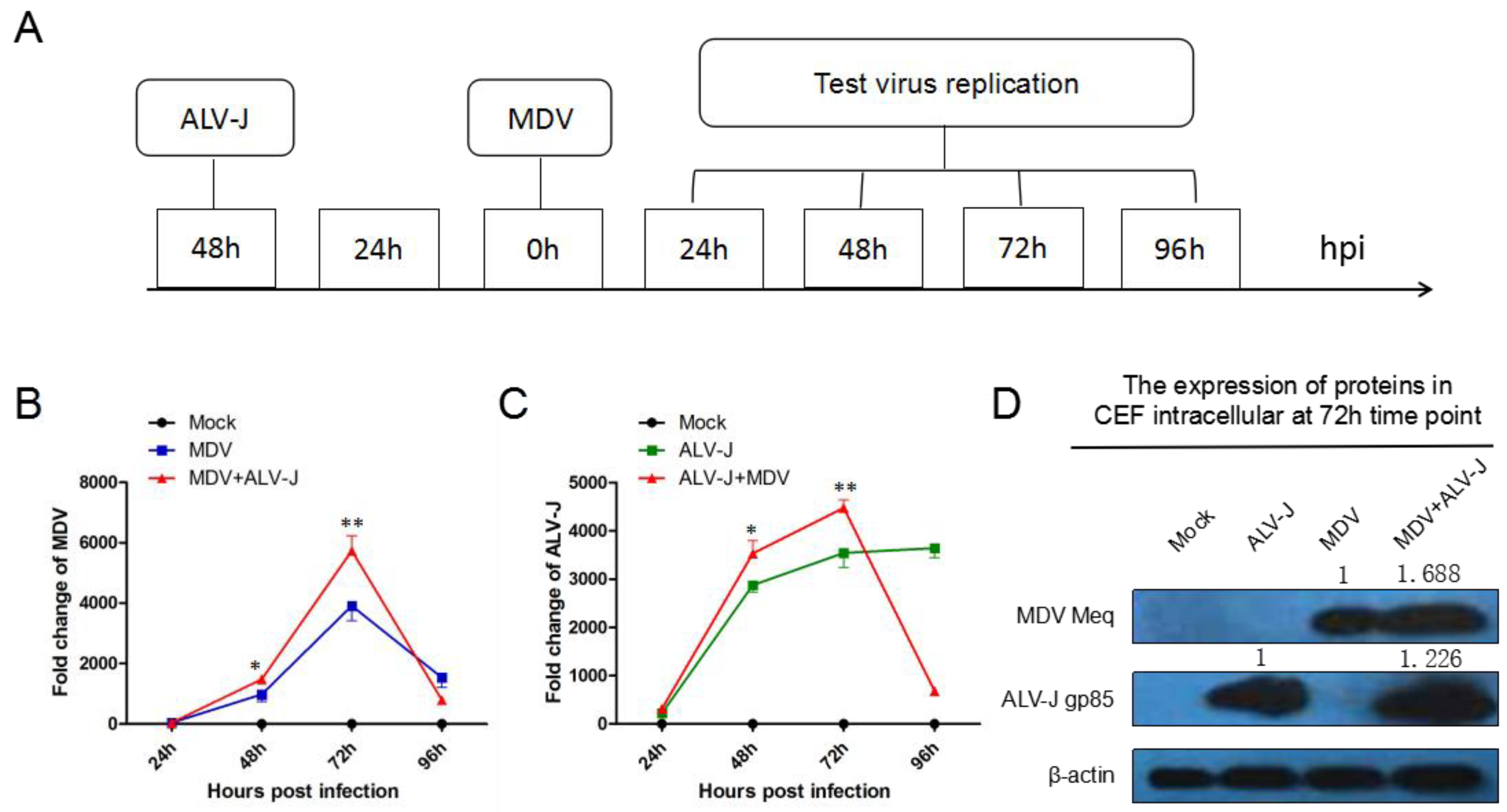
2. Materials and Methods
2.1. Viruses, Cells, and Animals
2.2. Quantitative Reverse Transcription PCR (RT-qPCR)
2.3. Western Blot
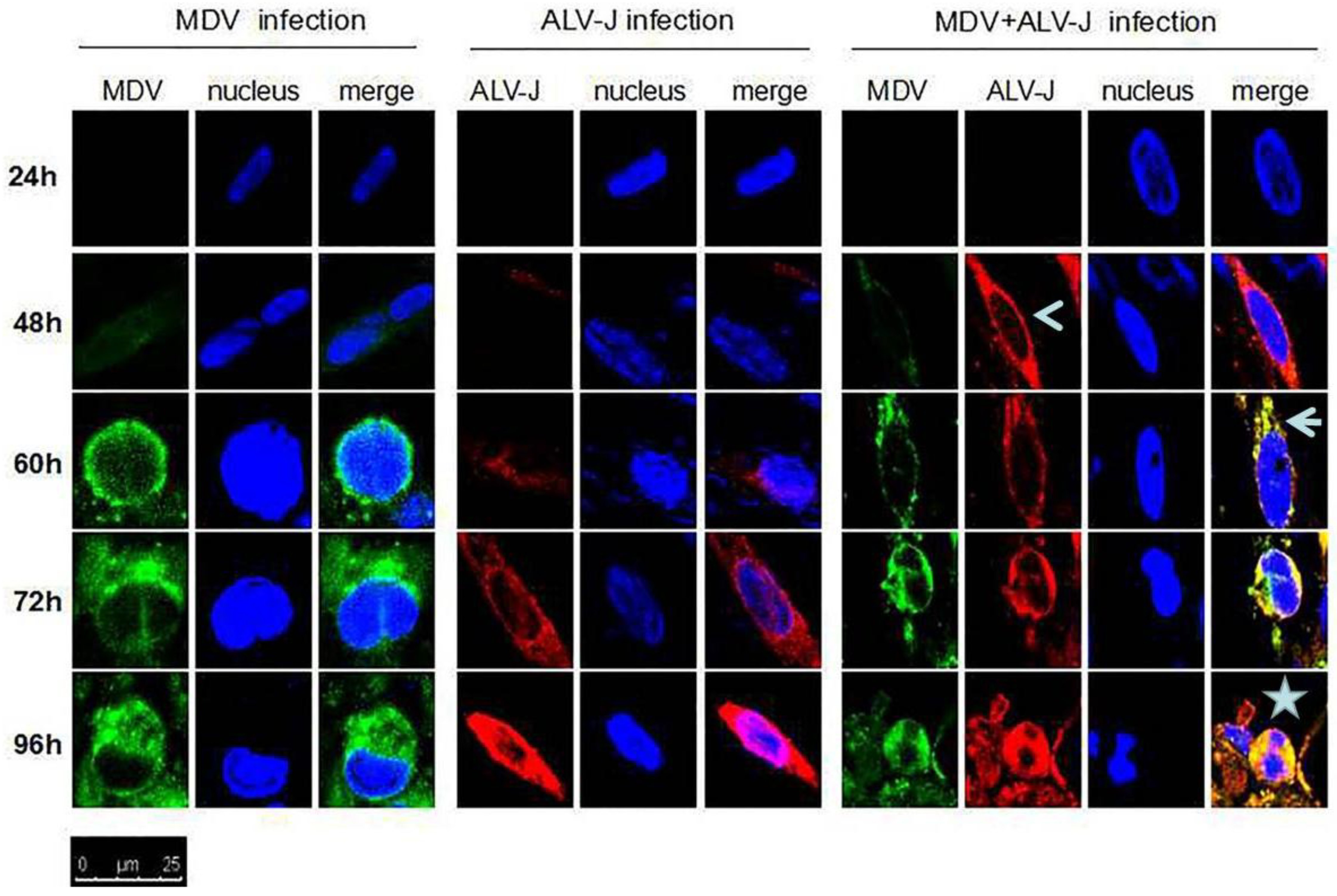
2.4. Confocal Laser Scanning Microscope Assay
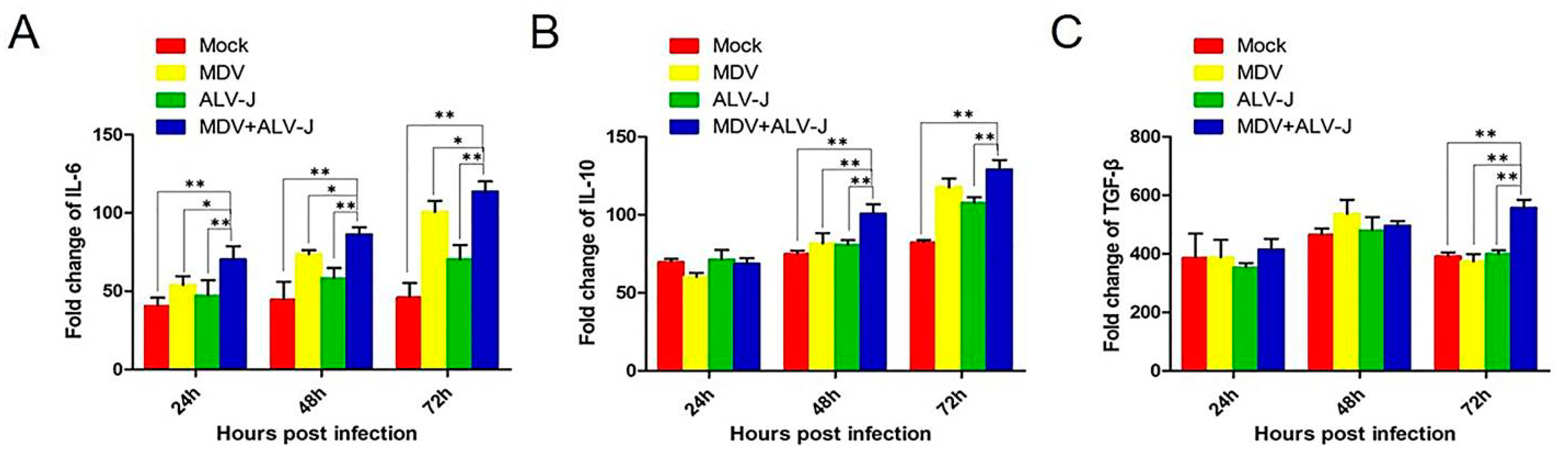
2.5. ELISA for Cytokines
2.6. Pathogenicity Assay in SPF Chickens
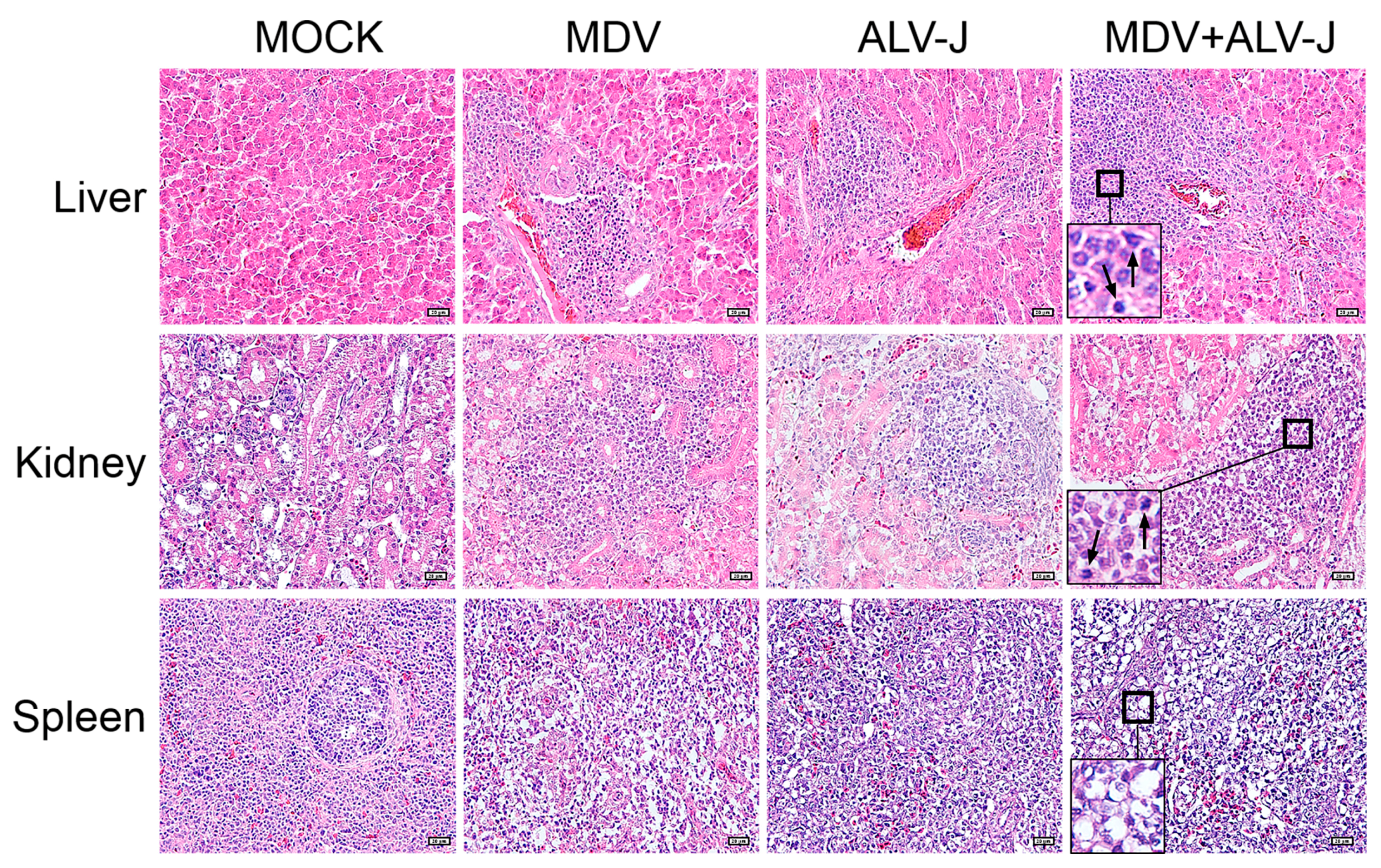
2.7. Histopathological Examination
3. Results
3.1. MDV and ALV-J Synergistically Increase Viral Replication In Vitro
3.2. MDV and ALV-J Synergistically Induce Cytopathy
3.3. Inflammatory Mediator Secretion
3.4. Mortality and Survival Curve
3.5. Tumorigenesis and Pathogenicity In Vivo
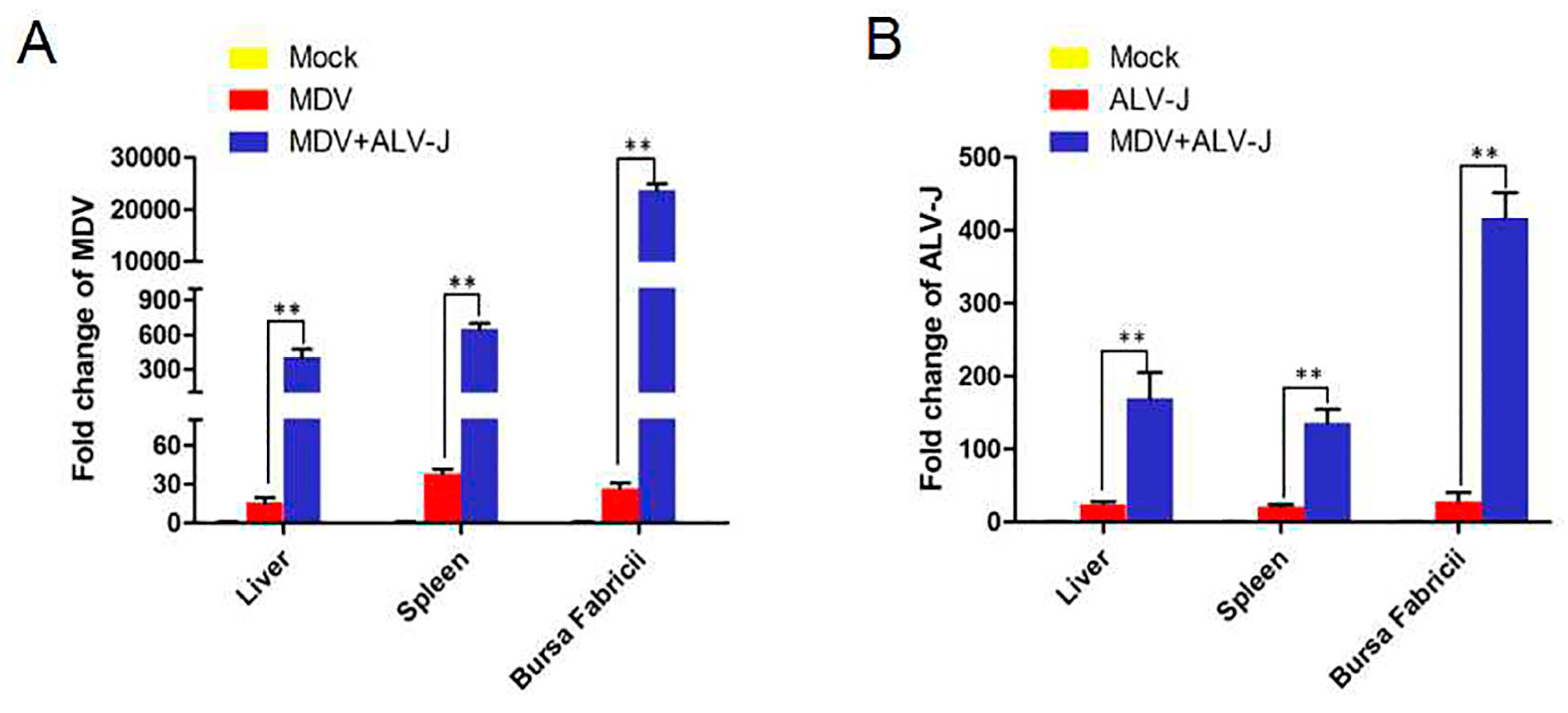
3.6. Virus Load in Tissues
4. Discussions
5. Conclusions
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Davison, A.J.; Eberle, R.; Ehlers, B.; Hayward, G.S.; Mcgeoch, D.J.; Minson, A.C.; Pellett, P.E.; Roizman, B.; Studdert, M.J.; Thiry, E. The order Herpesvirales. Arch. Virol. 2009, 154, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Baigent, S.; Davison, F. Marek’s disease virus: Biology and life cycle, In Marek’s Disease, an Evolving Problem; Davison, F., Nair, V., Eds.; Elsevier Academic Press: Oxford, UK, 2004; pp. 62–77. [Google Scholar]
- Hirai, K.; Sakaguchi, M. Polyvalent Recombinant Marek’s Disease Virus Vaccine against Poultry Diseases. Curr. Top. Microbiol. Immunol. 2001, 255, 261–287. [Google Scholar] [PubMed]
- Morrow, C.; Fehler, F. Marek’s disease: An evolving problem, In Marek’s Disease: A Worldwide Problem; Davison, F., Nair, V., Eds.; Elsevier Academic Press: London, UK, 2004; pp. 49–61. [Google Scholar]
- Boodhoo, N.; Gurung, A.; Sharif, S.; Behboudi, S. Marek’s disease in chickens: A review with focus on immunology. Vet. Res. 2016, 47, 119. [Google Scholar] [CrossRef] [PubMed]
- Witer, R.L. Marek’s disease vaccines—Past, present and future (Chicken vs. virus—A battle of the centuries). In Current Progress on Marek's Disease Research; Schat, K.A., Morgan, R.W., Parcells, M.S., Spencer, J.L., Eds.; American Association of Avian Pathologist: Kennett Square, PA, USA, 2001; pp. 1–9. [Google Scholar]
- Davison, F.; Nair, V. Use of Marek’s disease vaccines: Could they be driving the virus to increasing virulence? Expert Rev. Vaccines 2005, 4, 77–88. [Google Scholar] [CrossRef] [PubMed]
- Gimeno, I.M. Marek’s disease vaccines: A solution for today but a worry for tomorrow? Vaccine 2008, 26, C31–C41. [Google Scholar] [CrossRef] [PubMed]
- Rozins, C.; Day, T. The industrialization of farming may be driving virulence evolution. Evol. Appl. 2017, 10, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Witter, R.L. Increased virulence of Marek’s disease virus field isolates. Avian Dis. 1997, 41, 149–163. [Google Scholar] [CrossRef] [PubMed]
- Read, A.F.; Baigent, S.J.; Powers, C.; Kgosana, L.B.; Blackwell, L.; Smith, L.P.; Kennedy, D.A.; Walkdenbrown, S.W.; Nair, V.K. Imperfect Vaccination Can Enhance the Transmission of Highly Virulent Pathogens. PLoS Biol. 2015, 13, e1002198. [Google Scholar] [CrossRef] [PubMed]
- Wen, Y.; Huang, Q.; Yang, C.; Pan, L.; Wang, G.; Qi, K.; Liu, H. Characterizing the histopathology of natural co-infection with Marek’s disease virus and subgroup J avian leukosis virus in egg-laying hens. Avian Pathol. 2018, 47, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.; Borenshtain, R.; Kung, H.J.; Witter, R.L. Molecular indications for in vivo integration of the avian leukosis virus, subgroup J-long terminal repeat into the Marek’s disease virus in experimentally dually-infected chickens. Virus Genes 2002, 24, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Sun, G.R.; Zhang, Y.P.; Zhou, L.Y.; Lv, H.C.; Zhang, F.; Li, K.; Gao, Y.L.; Qi, X.L.; Cui, H.Y.; Wang, Y.Q. Co-Infection with Marek’s Disease Virus and Reticuloendotheliosis Virus Increases Illness Severity and Reduces Marek’s Disease Vaccine Efficacy. Viruses 2017, 21, 158. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Wang, L.; Sun, S. Natural Infection with Avian Hepatitis E Virus and Marek’s Disease Virus in Brown Layer Chickens in China. Avian Dis. 2016, 60, 698–704. [Google Scholar] [CrossRef] [PubMed]
- Haridy, M.; Sasaki, J.; Okada, K.; Goryo, M. Persistence of inclusions and antigens of chicken anemia virus in Marek’s disease lymphoma. Res. Vet. Sci. 2012, 93, 1353–1360. [Google Scholar] [CrossRef] [PubMed]
- Payne, L.N.; Brown, S.R.; Bumstead, N.; Howes, K.; Frazier, J.A.; Thouless, M.E. A novel subgroup of exogenous avian leukosis virus in chickens. J. Gen. Virol. 1991, 72, 801–807. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Z.Q.; Liu, J.Z.; Cui, Z.Z.; Li, Z. Tumors associated with avian leukosis virus subgroup J in layer hens during 2007 to 2009 in China. J. Vet. Med. Sci. 2010, 72, 1027–1033. [Google Scholar] [CrossRef] [PubMed]
- Fadly, A.M.; Smith, E.J. Isolation and some characteristics of a subgroup J-like avian leukosis virus associated with myeloid leukosis in meat-type chickens in the United States. Avian Dis. 1999, 43, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Wei, P.; Gao, Y.; Sun, F.; Qin, L.; Liu, Z.; Yun, B.; Wang, Y.; Qi, X.; Gao, H.; Wang, X. Novel sequences of subgroup J avian leukosis viruses associated with hemangioma in Chinese layer hens. Virol. J. 2011, 8, 552. [Google Scholar]
- Venugopal, K. Avian leukosis virus subgroup J: A rapidly evolving group of oncogenic retroviruses. Res. Vet. Sci. 1999, 67, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Davidson, I.; Borenshtain, R. The feather tips of commercial chickens are a favorable source of DNA for the amplification of Marek’s disease virus and avian leukosis virus, subgroup J. Avian Pathol. 2002, 31, 237–240. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.C.; Huang, D.; Pu, C.E.; Wang, C.H. Avian oncogenic virus differential diagnosis in chickens using oligonucleotide microarray. J. Virol. Methods 2014, 210, 45–50. [Google Scholar] [CrossRef] [PubMed]
- Giansanti, F.; Giardi, M.F.; Massucci, M.T.; Botti, D.; Antonini, G. Ovotransferrin expression and release by chicken cell lines infected with Marek’s disease virus. Biochem. Cell Biol. 2007, 85, 150–155. [Google Scholar] [CrossRef] [PubMed]
- Baigent, S.J.; Smith, L.P.; Nair, V.K.; Currie, R.J. Vaccinal control of Marek’s disease: Current challenges, and future strategies to maximize protection. Vet. Immunol. Immunopathol. 2006, 112, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Mascia, T.; Gallitelli, D. Synergies and antagonisms in virus interactions. Plant Sci. 2016, 252, 176–192. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.J.; Zhang, X.Y.; Liu, S.Y.; Wang, Y.; Li, D.W.; Yu, J.L.; Han, C.G. Synergistic infection of BrYV and PEMV 2 increases the accumulations of both BrYV and BrYV-derived siRNAs in Nicotiana benthamiana. Sci. Rep. 2017, 7, 45132. [Google Scholar] [CrossRef] [PubMed]
- Liberto, M.C.; Zicca, E.; Pavia, G.; Quirino, A.; Marascio, N.; Torti, C.; Focà, A. Virological Mechanisms in the Coinfection between HIV and HCV. Mediat. Inflamm. 2015, 2015, 320532. [Google Scholar] [CrossRef] [PubMed]
- Dittmer, D.P.; Damania, B. Kaposi sarcoma-associated herpesvirus: Immunobiology, oncogenesis, and therapy. J. Clin. Investig. 2016, 126, 3165–3175. [Google Scholar] [CrossRef] [PubMed]
- Diogo, P.; Miguel, A.P.J. The Role of microRNAs in the Pathogenesis of Herpesvirus Infection. Viruses 2016, 8, 156. [Google Scholar]
- Paoli, P.D.; Carbone, A. Microenvironmental abnormalities induced by viral cooperation: Impact on lymphomagenesis. Semin. Cancer Biol. 2015, 34, 70–80. [Google Scholar] [CrossRef] [PubMed]
- Abraham, A.G.; D’Souza, G.; Jing, Y.; Gange, S.J.; Sterling, T.R.; Silverberg, M.J.; Saag, M.S.; Rourke, S.B.; Rachlis, A.; Napravnik, S. Invasive cervical cancer risk among HIV-infected women: A North American multicohort collaboration prospective study. J. Acquir. Immune Defic. Syndr. 2013, 62, 405–431. [Google Scholar] [CrossRef] [PubMed]
- Guidry, J.T.; Scott, R.S. The Interaction between Human Papillomavirus and Other Viruses. Virus Res. 2017, 231, 139–147. [Google Scholar] [CrossRef] [PubMed]
- Tallarida, R.J. Quantitative methods for assessing drug synergism. Genes Cancer 2011, 2, 1003–1008. [Google Scholar] [CrossRef] [PubMed]
- Mercader, M.; Taddeo, B.; Panella, J.R.; Chandran, B.; Nickoloff, B.J.; Foreman, K.E. Induction of HHV-8 lytic cycle replication by inflammatory cytokines produced by HIV-1-infected T cells. Am. J. Pathol. 2000, 156, 1961–1971. [Google Scholar] [CrossRef]
- Barillari, G.; Ensoli, B. Angiogenic Effects of Extracellular Human Immunodeficiency Virus Type 1 Tat Protein and Its Role in the Pathogenesis of AIDS-Associated Kaposi’s Sarcoma. Clin. Microbiol. Rev. 2002, 15, 310–326. [Google Scholar] [CrossRef] [PubMed]
- Yao, S.; Hu, M.; Hao, T.; Li, W.; Xue, X.; Xue, M.; Zhu, X.; Zhou, F.; Qin, D.; Yan, Q. MiRNA-891a-5p mediates HIV-1 Tat and KSHV Orf-K1 synergistic induction of angiogenesis by activating NF-κB signaling. Nucleic Acids Res. 2015, 43, 9362–9378. [Google Scholar] [CrossRef] [PubMed]
- Khan, K.A.; Abbas, W.; Varin, A.; Kumar, A.; Di Martino, V.; Dichamp, I.; Herbein, G. HIV-1 Nef interacts with HCV Core, recruits TRAF2, TRAF5 and TRAF6, and stimulates HIV-1 replication in macrophages. J. Innate Immun. 2013, 5, 639–656. [Google Scholar] [CrossRef] [PubMed]
- Birdwell, C.E.; Queen, K.J.; Kilgore, P.C.; Rollyson, P.; Trutschl, M.; Cvek, U.; Scott, R.S. Genome-wide DNA methylation as an epigenetic consequence of Epstein-Barr virus infection of immortalized keratinocytes. J. Virol. 2014, 88, 11442–11458. [Google Scholar] [CrossRef] [PubMed]
- Queen, K.J.; Shi, M.; Zhang, F.; Cvek, U.; Scott, R.S. Epstein-Barr Virus Induced Epigenetic Alterations Following Transient Infection. Int. J. Cancer 2013, 132, 2076–2086. [Google Scholar] [CrossRef] [PubMed]
- Schlums, H.; Cichocki, F.; Tesi, B.; Theorell, J.; Beziat, V.; Holmes, T.D.; Han, H.; Chiang, S.C.; Foley, B.; Mattsson, K. Cytomegalovirus infection drives adaptive epigenetic diversification of NK cells with altered signaling and effector function. Immunity 2015, 42, 443–456. [Google Scholar] [CrossRef] [PubMed]
- Minarovits, J.; Demcsák, A.; Banati, F.; Niller, H.H. Epigenetic Dysregulation in Virus-Associated Neoplasms. Adv. Exp. Med. Biol. 2016, 879, 71–90. [Google Scholar] [PubMed]
- Cavignac, Y.; Esclatine, A. Herpesviruses and Autophagy: Catch Me If You Can! Viruses 2010, 2, 314–333. [Google Scholar] [CrossRef] [PubMed]
- Castanier, C.; Arnoult, D. Mitochondrial localization of viral proteins as a means to subvert host defense. Biochim. Biophys. Acta 2011, 1813, 575–583. [Google Scholar] [CrossRef] [PubMed]
- Morgan, R.W.; Sofer, L.; Anderson, A.S.; Bernberg, E.L.; Cui, J.; Burnside, J. Induction of Host Gene Expression following Infection of Chicken Embryo Fibroblasts with Oncogenic Marek’s Disease Virus. J. Virol. 2001, 75, 533–539. [Google Scholar] [CrossRef] [PubMed]
- Parvizi, P.; Read, L.; Abdul-Careem, M.F.; Lusty, C.; Sharif, S. Cytokine gene expression in splenic CD4+ and CD8+ T-cell subsets of chickens infected with Marek’s disease virus. Viral Immunol. 2009, 22, 31–38. [Google Scholar] [CrossRef] [PubMed]
- Heidari, M.; Zhang, H.M.; Sharif, S. Marek’s disease virus induces Th-2 activity during cytolytic infection. Viral Immunol. 2008, 21, 203–214. [Google Scholar] [CrossRef] [PubMed]
- Xu, M.; Fitzgerald, S.D.; Zhang, H.; Karcher, D.M.; Heidari, M. Very virulent plus strains of MDV induce an acute form of transient paralysis in both susceptible and resistant chicken lines. Viral Immunol. 2012, 25, 306–323. [Google Scholar] [CrossRef] [PubMed]
- Gurung, A.; Kamble, N.; Kaufer, B.B.; Pathan, A.; Behboudi, S. Association of Marek’s Disease induced immunosuppression with activation of a novel regulatory T cells in chickens. PLoS Pathog. 2017, 13, e1006745. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Target Gene | Orientation | Sequence | Size (bp) |
|---|---|---|---|
| Meq | Forward | 5′-GTTTCTCCAGATTCCACCTC-3′ | 231 |
| Reverse | 5′-TGCAACAATGCGTTCTTAT-3′ | ||
| Env | Forward | 5′-TGCGTGCGTGGTTATTATTTC-3′ | 144 |
| Reverse | 5′-AATGGTGAGGTCGCTGACTGT-3′ | ||
| GADPH | Forward | 5′-GAACATCATCCCAGCGTCCA-3′ | 132 |
| Reverse | 5′-CGGCAGGTCAGGTCAACAAC-3′ |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, J.; Zhao, G.-L.; Wang, X.-M.; Du, X.-S.; Su, S.; Li, C.-G.; Nair, V.; Yao, Y.-X.; Cheng, Z.-Q. Synergistic Viral Replication of Marek’s Disease Virus and Avian Leukosis Virus Subgroup J is Responsible for the Enhanced Pathogenicity in the Superinfection of Chickens. Viruses 2018, 10, 271. https://0-doi-org.brum.beds.ac.uk/10.3390/v10050271
Zhou J, Zhao G-L, Wang X-M, Du X-S, Su S, Li C-G, Nair V, Yao Y-X, Cheng Z-Q. Synergistic Viral Replication of Marek’s Disease Virus and Avian Leukosis Virus Subgroup J is Responsible for the Enhanced Pathogenicity in the Superinfection of Chickens. Viruses. 2018; 10(5):271. https://0-doi-org.brum.beds.ac.uk/10.3390/v10050271
Chicago/Turabian StyleZhou, Jing, Guo-Liang Zhao, Xiao-Man Wang, Xu-Sheng Du, Shuai Su, Chen-Gui Li, Venugopal Nair, Yong-Xiu Yao, and Zi-Qiang Cheng. 2018. "Synergistic Viral Replication of Marek’s Disease Virus and Avian Leukosis Virus Subgroup J is Responsible for the Enhanced Pathogenicity in the Superinfection of Chickens" Viruses 10, no. 5: 271. https://0-doi-org.brum.beds.ac.uk/10.3390/v10050271