1. Introduction
The
Papaver genus in the
Papaveraceae family comprises up to 100 plant species distributed in various areas of the world. Among them, viral infection of two
Papaver species was investigated in the present study, i.e., oilseed poppy (
P. somniferum L.) and common poppy (
P. rhoeas L.). Originating from Asia Minor or from the Western Mediterranean region, oilseed poppy cultivation covers a wide geographic area around the world due to the adaption of this species to various climatic conditions as a result of its long history of domestication and breeding. The oilseed poppy is legally grown in Slovakia for the seeds used as an ingredient in some foods and for the seed capsules containing several alkaloids, including morphine, codeine, papaverine, and thebaine, valuable in the pharmaceutical industry [
1,
2,
3]. On the contrary, common poppy is a red-flowering agricultural weed abundantly spread in Europe [
4].
Few viruses infecting these
Papaver spp. have been reported to date [
5]. Viruses known from older records to infect these species include
Bean yellow mosaic virus (BYMV),
Beet mosaic virus (BtMV) and
Turnip mosaic virus (TuMV) from the genus
Potyvirus, and a polerovirus,
Beet western yellows virus (BWYV) [
5,
6]. More recently, a new umbravirus,
Opium poppy mosaic virus, was identified as causing leaf mosaic and mottling in opium poppy in New Zealand [
7] and
Tomato leaf curl New Delhi virus was detected from poppies showing curling of leaves in India [
8]. Despite these reports, knowledge of viral pathogens naturally infecting poppy remains limited.
TuMV is noted as a particularly damaging pathogen worldwide, causing diseases in more than 300 mono- and dicotyledonous plant species, including not only economically important vegetable, oilseed, forage, and ornamental crops, but also a wide range of weed species [
9,
10]. TuMV is transmitted in a non-persistent manner by aphids. Similar to other potyviruses, its genome consists of a single-stranded positive sense RNA molecule of ca. 9800 nucleotides. It contains one open reading frame (ORF) encoding a large polyprotein that is processed to at least 10 final gene products [
11]. An additional product P3N-PIPO produced from an alternative ORF, probably by a polymerase slippage mechanism, has been detected, as in other potyviruses [
12,
13].
The currently known TuMV diversity includes six TuMV genogroups revealed by phylogenetic analysis of numerous TuMV isolates collected around the world: basal-B (
Brassica), basal-BR (
Brassica/
Raphanus), Asian-BR, world-B, Iranian, and Orchis [
14,
15]. Despite the huge number of sequences accumulated during the recent years, the only genome sequence from oilseed poppy, originating from Poland, was reported as belonging to the world-B group [
14].
In the present work, 4 complete TuMV genomes from a cultivated and a wild-growing Papaver sp. were characterised, showing their respective affiliation to the world-B and basal-B groups. Moreover, biological assays indicated that intensity of symptoms induced in experimentally inoculated oilseed poppy were correlated with the virus accumulation levels in leaves.
2. Material and Methods
2.1. Samples
An oilseed poppy plant (
P. somniferum, labelled as HC9) showing suspicious symptoms (chlorosis, slight deformation of leaf lamina,
Supplemental Figure S1) and growing in a commercial field near Hlohovec, western Slovakia, was sampled in May 2016 and subjected to the high throughput sequencing (HTS).
Additionally, a total of 56 oil poppy plants and 10 samples from common poppy (P. rhoeas), growing as weeds near cultivated oilseed poppy plants in 5 different locations in Slovakia in early June 2017 (Pezinok, Hlohovec, Malý Šariš, Habovka, Martin) were tested for the presence of TuMV using double-antibody sandwich enzyme-linked immunosorbent assay (DAS-ELISA, Bioreba, Reinach, Switzerland). Two samples from oilseed poppy (MS4 and MS15, both from Malý Šariš) and 1 sample from common poppy (PK1, from Pezinok) testing positive for TuMV were further processed by HTS.
2.2. Analysis of the Virome and Determination of the TuMV Full Length Genome Sequences by HTS
Total RNAs from infected poppy leaves were extracted using the NucleoSpin RNA Plant kit (Macherey-Nagel, Duren, Germany) and ribosomal RNA was removed using the Ribo-Zero rRNA Removal Kit (Illumina, San Diego, CA, USA). The rRNA-depleted total RNAs sample was used for double stranded cDNA synthesis using the SuperScript II (Thermo Fisher Scientific, Waltham, MA, USA) kit. The cDNA was then column-purified with the DNA Clean & Concentrator™-5—DNA kit (Zymo Research, Irvine, CA, USA) and quantified with the Qubit 2.0 Fluorometer (Thermo Fisher Scientific, Waltham, MA, USA). The sample was then processed with the transposon-based chemistry library preparation kit (Nextera XT, Illumina, San Diego, CA, USA). Low-cycle PCR and mutual indexing of the fragments was carried out. Fragments were purified with 1.8× AMPure XP beads (BeckmanCoulter, Brea, CA, USA) without size selection. The fragment size structure of the DNA library was assessed using the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA). The equimolar pool of 4 nM DNA libraries was denatured, diluted to 13 pM and sequenced (200-bp paired-end sequencing) on the Illumina MiSeq platform (Illumina, San Diego, CA, USA).
High-quality trimmed reads were used for de novo assembly and contigs aligned to the viral genomes database (
ftp://ftp.ncbi.nih.gov/genomes/Viruses/all.fna.tar.gz) using CLC Genomics Workbench 7.5 (Qiagen, Hilden, Germany) and Geneious v.8.1.9. (Biomatters, Auckland, New Zealand) Subsequently, the reads were mapped against the reference TuMV sequence (NC_002509). When necessary, the accuracy of the obtained HTS-based sequences was verified by direct Sanger sequencing of PCR products using primers designed from the HTS-based sequences. In parallel, to assess the intra-isolate sequence heterogeneity, PCR products were cloned into the pGEM-T Easy cloning vector (Promega, Madison, WI, USA) and 6–8 randomly chosen clones were sequenced using universal pUC primers.
The nearly complete nucleotide TuMV sequences reported in this paper (missing 37 nt at the 5′ extremity as compared to the reference NC_002509 genome) have been deposited in the GenBank database under accession numbers MH469724 (PK1), MH469725 (HC9), MH469726 (MS15), and MH469727 (MS04).
Phylogenetic analyses and comparisons with available TuMV sequences (
www.ncbi.nlm.nih.gov) were performed using the MEGA v.7 [
16] and DnaSP v.5 [
17] programs.
Searches for potential recombination events and identification of recombination breakpoints employed seven methods, including RDP, GENECONV, Bootscan, Maxchi, Chimaera, SiScan, and 3SEQ, implemented in the RDP4 v.4.1 software [
18] using default settings. Recombination events were noted if supported by at least four different methods (
p-values < 1.0 × 10
−6).
2.3. Inoculation of Oilseed Poppy and Experimental Plants
The oilseed poppy genotypes ZB24 and ZB25, new-breeding lines of the Slovak origin (National Agriculture and Food Centre, Malý Šariš, Slovakia), were tested for their susceptibility to the TuMV isolates HC9 and PK1 after experimental mechanical inoculation. For the inoculation experiments, homogenous lots of 12 plants were mechanically inoculated at the 3–4 true leaves stage using an inoculum prepared by grinding TuMV-infected leaves 1:10 (w/v) in 0.1 M sodium phosphate buffer, pH 7.0. Nicotiana benthamiana, Raphanus sativus cv. Slovana and R. sativus subs. Niger cv. Kulatá černá were simultaneously inoculated as controls. Inoculated plants were grown in an insect-proof cultivation room under controlled conditions (14 h light/10 h dark photoperiod, 55 μmol·m−2·s−1 photon flux density, day/night temperature: 25/18 °C). Development of symptoms was monitored visually and TuMV presence was verified by DAS-ELISA 14 days post inoculation. The experiment was repeated 2 times.
2.4. Estimation of Viral Accumulation in Plants
The estimation of viral accumulation in oilseed poppy leaves were performed 14 days post inoculation, using plant issues from second inoculation experiment. The uppermost fully developed leaves were sampled from the plants and equal leaf weight was used for sample preparation. Semiquantitative DAS-ELISA was performed following a previously described protocol [
19]. All the samples were homogenised in PBS buffer (1/25
w/
v) containing 0.05% Tween-20 and 2% polyvinylpyrrolidone and subsequently applied to the same ELISA plate, so that absorbance values measured at 405 nm for different genotypes could be compared.
For western blot analysis [
20], the crude sap from plant leaves (ground 1:5 in PBS and clarified by 5 min centrifugation at 13,000 g) was mixed 1:1 with Laemmli sample buffer, subjected to SDS-PAGE in 12% gel and stained by Coomassie Brilliant Blue (CBB). Alternatively, the proteins were transferred to a PVDF membrane by a semidry blotting apparatus and detected using polyclonal anti-TuMV antibody (Bioreba, cat. No. 161015)/goat anti-rabbit IgG antibody conjugated with alkaline phosphatase (Sigma, St. Louis, MO, USA) and NBT/BCIP as substrate.
3. Results and Discussion
In an effort to assess the presence of viral pathogens naturally infecting poppy, total RNAs isolated from leaves of an oilseed poppy plant showing virus-like symptoms were subjected to HTS. The analysis of the sequence data obtained from this sample (HC9, ca. 2.1 millions of reads of mean length 89.7 nucleotides) enabled us to obtain a large contig of ca. 9 kb, corresponding to TuMV. This was further confirmed by the mapping of individual reads against the NC_002509 reference TuMV sequence, allowing the reconstruction of the nearly complete genome (missing 37 nt at the 5′ end as compared to NC_002509) with a mean coverage of 483×. No other viral agent was identified in the HC9 sample.
This discovery prompted us to perform a screening for TuMV presence in a larger set of 66 samples of Papaver somniferum and P. rhoeas from 5 different locations in Slovakia. DAS-ELISA revealed TuMV infection in 2 out of 56 oilseed poppy plants and in 1 out of 10 common poppy plants. These samples, labelled as MS4 and MS15 (both from P. somniferum grown in Malý Šariš) and PK1 (from P. rhoeas found in Pezinok), were further analysed using HTS.
De novo assembly of high-quality reads (between 2.1 and 3.1 million depending on the sample) from MS4, MS15, and PK1 resulted in the assembly of nearly complete TuMV genomes (mean coverage 210–709×). While only TuMV was identified in the MS4 and MS15 samples, the PK1 sample was found to be co-infected with Cucumber mosaic virus (CMV).
In accordance with typical potyvirus genomic organization [
11,
21], a large ORF (9489–9495 nts in length) was identified in all 4 isolates, encoding a polyprotein of 3162 aa (PK1) or 3164 aa (HC9, MS4, MS15) with an estimated MW of 357.30–357.45 kDa, respectively (
http://web.expasy.org/protparam/). A two-amino acid deletion at position 39–40 of the capsid protein (aa 2915–2916 based on the reference NC_002509 polyprotein) is responsible for a shorter polyprotein of PK1. Nine putative cleavage sites were identified in all polyproteins. The PIPO ORF [
12] was identified in silico downstream of the highly conserved motif GA6 (nt position 3041–3047). While PIPO of the PK1 isolate encodes deduced product of 60 aa, that identified in HC9, MS04, and MS15 encodes a longer product of 69 aa.
All potyvirus motifs involved in the virus infectious cycle were perfectly conserved among the four Slovak TuMV poppy isolates at the position previously indicated [
22]. The conservative motifs essential for aphid transmission of potyviruses (KITC
414–417, PTK
672–674, DAG
2882–2884) were found in their polyprotein sequences at the expected positions.
Based on a Blast search, the closest relatives of the PK1 isolate are basal-B group isolates from Greece (GRC45, GRC70) and Turkey (TUR59, TUR60, TUR61), but PK1 divergence is significant, as it shows only ca. 89% nt identity with them. PK1 thus appears as a divergent isolate of the basal-B genogroup. On the contrary, MS04 and MS15 (having 99.6% mutual nt identity) were 99% identical to the Croatian CRO184A isolate while HC9 was ca. 97% identical to the CZE5, POL1, and POL2 isolates from the Czech Republic and Poland, respectively, all of which belong to the word-B group (subgroup 2). The affiliation of Slovak poppy isolates was further confirmed by a phylogenetic analysis performed using a complete database of non-recombinant sequences belonging to the 6 recognised TuMV genogroups [
14,
15] (
Figure 1).
RNA recombination is one of the driving forces shaping the genetic variability in TuMV, with various recombination events described, including both intra- and inter-lineage recombinations [
23,
24,
25,
26,
27]. The four Slovak isolates were assessed for possible evidence of recombination using a set of 33 non-recombinant TuMV isolates [
14,
15] (accession numbers listed in
Figure 1). The RDP4 analysis [
18] revealed a recombination event in the genome of the PK1 isolate, with breakpoints in the HC-Pro gene (nt positions 1195–2317). This event was supported by 5 programs of the RDP4 package (for details see
Supplemental Table S1). Using a Blast search, the closest potential donor sequence (indicating the lineage that was likely to have provided this region of the recombinant genome) was found to be the Turkish AP017736 isolate from
Raphanus raphanistrum with 91% nucleotide identity. Although a number of recombination events affecting TuMV have been described [
14,
15,
23,
24,
25,
26,
27], the recombination pattern and breakpoints identified in the PK1 genome had not been previously reported.
The genome of the other three Slovak isolates (HC9, MS4, and MS15) did not reveal statistically supported evidence for recombination. However, HC9 was tentatively identified as a minor parent of the Czech AB252107 isolate and MS15 as a tentative minor parent involved in a recombination event affecting the 3′ region of the genome of several Vietnamese and Chinese isolates from the world-B group (AB747293, AB747297, AB747307, AB747312-15, HQ446216-17,
Figure 1), all undergoing a previously overlooked intra-lineage recombination (supported by 6 algorithms,
Supplementary Table S1). Our results thus further highlight the important role of recombination in the evolutionary history of TuMV.
Despite the importance of TUMV and its negative effect to the production of many crops, mainly the studies focusing on
Brassicaceae were reported [
28,
29,
30,
31,
32]. On the contrary, there is no data about the TuMV/oilseed poppy relationship [
6]. The susceptibility of two oilseed poppy genotypes, ZB24 and ZB25, was therefore evaluated by experimental mechanical inoculation with TuMV isolates PK1 and HC-9, belonging respectively to the world-B and basal-B molecular clades.
As only TuMV infection was revealed by HTS analysis of the HC9 sample, the original leaf sample, stored at −80 °C, was used for these experimental inoculations, providing reassurance on the purity of the inoculum. In parallel, since the original PK1 sample was found coinfected with CMV, to ensure a single TuMV infection, the plant sap from original host plant was highly diluted (1/150, w/v) and a set of Nicotiana benthamiana plants were mechanically inoculated. These tobacco plants were then individually tested for TuMV and CMV presence. An unambiguously TuMV singly-infected plant, repeatedly tested CMV-negative using ELISA, RT-PCR, and back inoculation to Cucumis sativus was used as the source of PK1 TuMV isolate (details of CMV detection methods used available upon request).
The natural viral isolates consist of a cloud of closely related viral genomes, called quasispecies, and this mutant spectra and not individual genomes are the target of evolutionary events [
33]. This gentle sequence heterogeneity is usually overlooked in standard sequencing efforts. In this work, an “isolate” refers to the determined single consensus dominant sequence, further used for comparisons of isolates and phylogenetic purposes.
The intra-isolate sequence homogeneity estimated from HTS reads and from sequencing of individual PCR-derived clones suggested the presence of unique TuMV population in each sample. Therefore, to not impose additional selection pressure to TuMV population, a passage through a local lesion host was omitted. However, we cannot exclude the possibility of a mixed infection of TuMV population by a contaminant TuMV variant. If any, such a low-frequency heterogeneity would not have an impact on the biological or molecular data obtained.
Both TuMV isolates could be mechanically transmitted to oilseed poppy plants with 40–66% efficacy (
Table 1) as proved by DAS-ELISA. None of the symptomless plants tested TuMV-positive, showing a clear correlation between symptoms expression and virus presence as detected by DAS-ELISA. Different inoculum sources were used for these biological assays, likely resulting in different virus titers (original infected oilseed poppy leaves for HC9 or leaves of experimentally infected
N. benthamiana for PK1, respectively), therefore no effort was made to compare the intensity of the symptoms induced by the two isolates. Notwithstanding, in all cases, two types of symptoms were observed (
Table 1,
Figure 2). One type of symptom consisted of chlorotic patterns and dark green spots on the leaf lamina (termed as “moderate”). The second type of symptom (“severe”) consisted of shoestringing, an extreme leaf deformation with loss of the limb part of the leaves (
Figure 2). Irrespective of the symptomatology type, subsequent systemic necrosis was observed, leading to the stunting, wilting, and finally dying of positive plants 35–45 days post-inoculation. The experimental inoculation of
N. benthamiana plants lead to 100% infection, with both TuMV isolates inducing leaf deformation, mosaics, and plant stunting. On the contrary, only one out of 40
R. sativus inoculated plants ended up infected, suggesting that this species may not be adapted to the study of Slovak TuMV isolates from poppy.
The symptoms and their development are the result of a complex plant–virus interplay [
34]. Moderate and severe symptoms were observed in both oilseed poppy genotypes, irrespective of the TuMV isolate used, suggesting that this is apparently not an isolate- or host genotype-specific phenomenon. To investigate it further, the accumulation of viral CP was evaluated in symptomatic leaves showing either the moderate or the severe symptoms. Semi-quantitative DAS-ELISA provided approximately 2.2–3.1 times higher OD values for samples prepared from leaves showing severe symptoms (
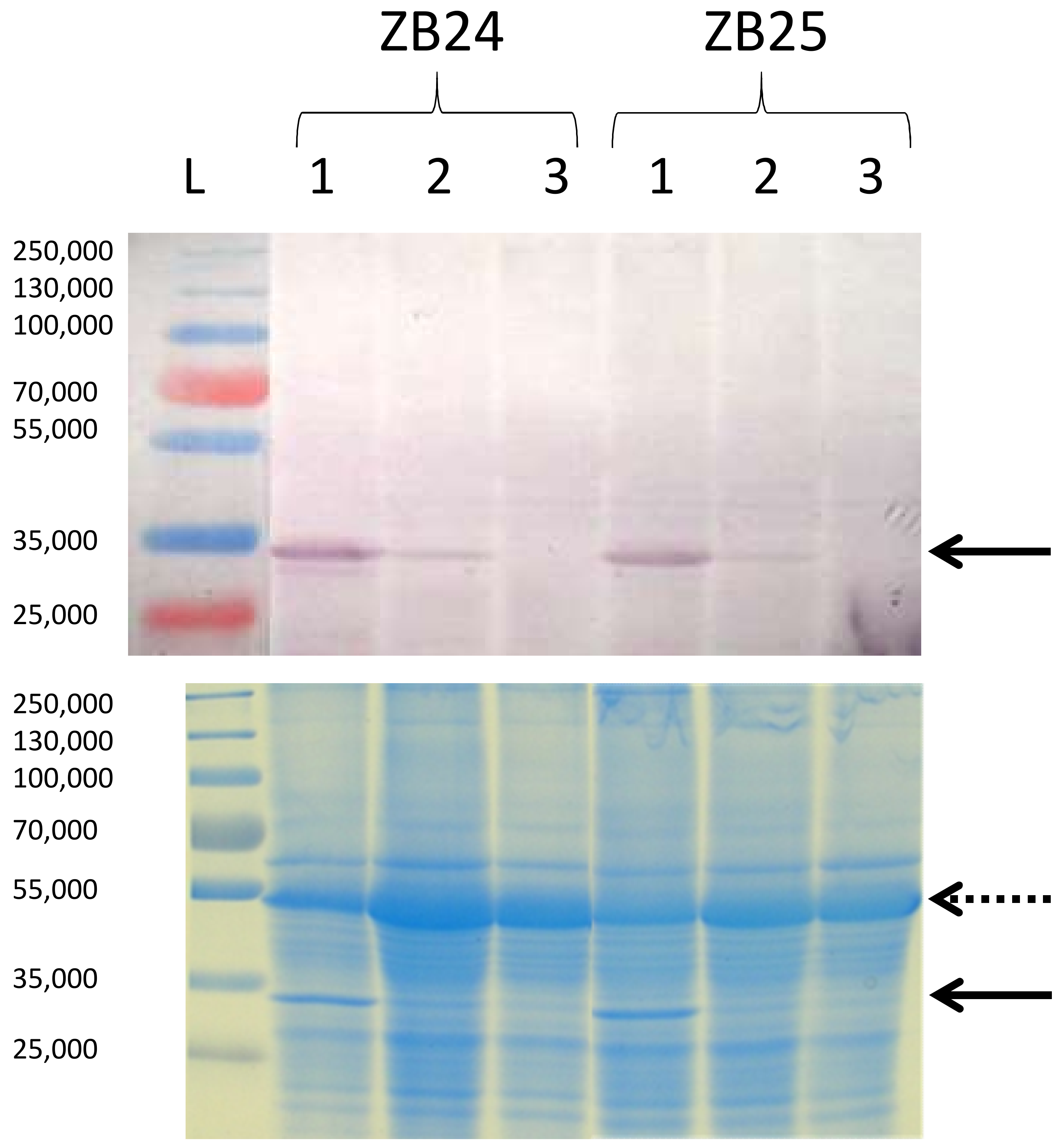
Table 1). This correlation of TuMV CP accumulation with the symptoms type was confirmed by immunoblotting (
Figure 3). These results indicate that the intensity of symptoms was positively correlated with virus accumulation in the leaf.
Possible hypotheses to explain the observed differences in symptomatology of oilseed poppy plants of the same genotype and inoculated with the same TuMV isolate could be a slightly uneven initial inoculum load [
35] and/or differences in the physiological states of the young inoculated plants, despite the precautions taken to standardise the experiment. Although the experimental mechanical transmission of TuMV did not mimic the natural inoculation of the virus by its aphid vectors, our results suggest that natural TuMV infection of young oilseed poppy plants could also result in their death by systemic necrosis under field conditions.
In conclusion, the present work extends our knowledge on the host range of TuMV, showing world-B and basal-B TuMV isolates are able to naturally infect both cultivated and weed Papaver sp. and to cause significant symptoms in these hosts.

 ,
, 



{kind=link}
{kind=link}
{kind=link}