Maize Dwarf Mosaic Virus: From Genome to Disease Management


1
Institute of Systems Biology, Universiti Kebangsaan Malaysia, 43600 Bangi, Malaysia
2
School of Bioscience and Biotechnology, Faculty of Science and Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Malaysia
*
Author to whom correspondence should be addressed.
Viruses 2018, 10(9), 492; https://0-doi-org.brum.beds.ac.uk/10.3390/v10090492
Submission received: 24 July 2018
/
Revised: 22 August 2018
/
Accepted: 28 August 2018
/
Published: 13 September 2018
(This article belongs to the Section Viruses of Plants, Fungi and Protozoa)
Abstract
:Maize dwarf mosaic virus (MDMV) is a serious maize pathogen, epidemic worldwide, and one of the most common virus diseases for monocotyledonous plants, causing up to 70% loss in corn yield globally since 1960. MDMV belongs to the genus Potyvirus (Potyviridae) and was first identified in 1964 in Illinois in corn and Johnsongrass. MDMV is a single stranded positive sense RNA virus and is transmitted in a non-persistent manner by several aphid species. MDMV is amongst the most important virus diseases in maize worldwide. This review will discuss its genome, transmission, symptomatology, diagnosis and management. Particular emphasis will be given to the current state of knowledge on the diagnosis and control of MDMV, due to its importance in reducing the impact of maize dwarf mosaic disease, to produce an enhanced quality and quantity of maize.
1. Introduction
Maize (Zea mays L.) is one of the most highly cultivated crops worldwide. It is the third largest crop grown in the developing world and it has been identified as a major staple food in Africa [1]. In south and eastern Africa, maize serves as a primary food for more than 24 million families and also as a subsistence food source for poor populations outside Asia [2]. Approximately 177 million hectares of tropical, semitropical and temperate zones are used to cultivate maize [3] and 875 million metric tons of maize are produced every year [4].
More than 50 viruses have been shown to infect maize [5]. These include Sorghum mosaic virus (SrMV) [6,7], Johnsongrass mosaic virus (JGMV) [8], Maize chlorotic mottle virus (MCMV), and Maize chlorotic dwarf virus (MCDV) [9]. Among all the maize-infecting viruses, Maize dwarf mosaic virus (MDMV) is the most common disease agent in this monocotyledonous crop globally [10], with incidences of maize dwarf mosaic (MDM) disease reported in Africa, United States, Asia and Europe [11].
MDMV strains’ nomenclature was in a state of confusion, since MDMV is intimately correlated with the Sugarcane mosaic virus (SCMV) [12]. The organization of inclusions and alterations produced by four isolates representing different virus strains from the subgroup of SCMV in infected cells were studied and differentiated by cellular pathology [13]. The results obtained demonstrated the presence of typical inclusions only in MDMV infected cells, but laminar aggregations were also exist in SCMV infected cells [14].
MDMV is classified in Potyviridae, the most numerous group of plant viruses [15]. There are 8 genera under the family; Potyvirus, Tritimovirus, Brambyvirus, Rymovirus, Ipomovirus, Bymovirus, Macluravirus, and Poacevirus [16]. MDMV belongs to the genus Potyvirus. In 1964, flexuous rod-shaped MDMV with a length of 750 nm and diameter of 12–15 nm was identified for the first time in Illinois [17]. MDMV is a holoparasite that requires a vector or host to reproduce and survive [18]. The infected plants take up to 15 days to exhibit the symptoms [19]. In this case, both serological and molecular methods have been developed to diagnose the MDM diseased plants more efficiently, eliminating the dependence on symptoms observation for identification purposes. The viral particles were observed to be distributed generally in cytoplasm and infrequently in plasmodesmata [20]. The lowest temperature needed to inactivate MDMV completely is 54–58 °C. At room temperature, survival in vitro for MDMV lasts 1–2 days while at 0–4 °C, it lasts 3–5 days [21,22].
In general, the loss in maize yield caused by MDMV can be up to 70% [23,24], mainly due to a reduction in the rate of photosynthesis and elevation in the rate of respiration [25]. Taking into account the economic significance of the disease, our current review has discussed available information from genome to global management of MDMV, so that greater effort would be put towards further research work in controlling MDM disease for the maintenance of profitable production and the good health of maize crops in future.
2. Genome
Like other potyviruses, MDMV is a positive-stranded RNA [18,26]. The MDMV genome is ~9500 base pairs in length with a covalently bounded viral-genome-linked protein, VPg at its 5′ end as well as a poly (A) tail attached to its 3′ end [10]. A large 338 kDa polyprotein is translated from a single open reading frame (ORF) [27], which is subsequently proteolytically cleaved by three self-coded proteinases to yield 10 final proteins (P1, HC-Pro, P3, 6K1, CI, 6K2, NIa-VPg, NIa-Pro, NIb, and CP) with multiple functions [28,29]. The last cistron that encodes the capsid protein (CP) [30] is greatly conserved among numerous potyvirus species [31]. This protein subunit of MDMV has a molecular weight of 28.5 × 103 according to amino acid analyses [32]. The C-terminal regions of the coat protein primarily plays a role in the process of encapsidation and cell-to-cell transport and the flexible N-terminus is involved in long distance and systemic transport and contains the DAG motif essential for aphid transmission competence [33].
Several complete sequences of MDMV isolates are currently available (Table 1). The viral genome of MDMV-BG is composed of 9515 nucleotides and contains an open reading frame encoding 3042 amino acids, flanked by 30 and 50 untranslated regions (UTRs) of 139 and 250 nucleotides, respectively. Meanwhile, the genome of MDMV-Sp is made up of 9414 nt and contains one ORF encoding 3042 amino acids, flanked by 50 and 30 UTRs of 138 and 234 nucleotides, respectively. The similarities in nucleotide and amino acid sequence between MDMV-BG and MDMV-Sp are 85% and 93%, respectively [27]. MDMV-OH1 encoded P1 protease (nt 1400–838), HC-Pro (nt 839–2218), P3 protease (nt 2219–3259), 6K1 (nt 3260–3460), CI (nt 3461–5350), 6K2 (nt 5351–5509), NIa-VPg (nt 5510–6076), NIa-Pro (nt 6077–6802), NIb-replicase (nt 6803–8365), CP (nt 8366–9180). It also encodes the “PIPO” (nt 2678-2922) that overlaps the P3 gene. PIPO is a new ORF recently described to be expressed as a result from the transcriptional slippage specific to the viral RNA polymerase [34], it well conserved and has a strong bioinformatics coding signature throughout the Potyviridae family members [35]. It is translated in the +2 reading frame relative to the potyviral long ORF as the P3N-PIPO fusion protein [36]. Potyviral P3N-PIPO interacts with the PCaP1 host plasma membrane protein to function in cell-to-cell movement [37]. MDMV-OH1 has an identity of about 97% and 98% in nucleotide sequences and polyprotein amino acid sequences respectively with MDMV-OH2. Both MDMV-OH isolates are more related to MDMV-It isolate rather to other European isolates based on the phylogenetic tree constructed.
The construction of the full-length infectious cDNA clones has been an important technique in the understanding of virus fundamental mechanisms studies [42]. Moreover, generation of infectious clone allows mutagenesis studies, insertion, substitution and deletions, to be conducted at any particular region of the viral genome [43]. Gell [44] initiated the attempt to develop the full-length virulent MDMV clones. Unfortunately, no typical symptoms of MDMV infection were observed in the inoculated plants and no viral particles were detected using the polymerase chain reaction (PCR) approach. The next attempt was made in 2012, Stewart et al. [40] managed to produce the full-length infectious MDMV cDNA clone using MDMV-OH1 isolate. The study managed to overcome the toxicity exhibited by plasmids containing full-length MDMV in Escherichia coli. According to Ali et al. [45], cDNA clones are often unstable due to the toxicity of the viral proteins such as P3 and CI for bacteria.
In earlier literature, there was a considerable confusion in the taxonomy of the potyvirus, due to huge size of the group and wide diversification among the species [6]. MDMV strains have been differentiated serologically [46,47] and by host range [48,49]. Serological relationship investigation of 13 American and 4 Australian strains of MDMV and SCMV using virus-specific polyclonal antisera directed against N-terminal regions of their coat proteins led to the establishment of four groups of viruses: (1) MDMV (MDMV-A, MDMV-D, MDMV-E, MDMV-F); (2) SCMV (MDMV-B); (3) JGMV (MDMV-O) and (4) SrMV forming a subgroup of SCMV [50]. This shows that a serological approach has the potential to be utilized as a taxonomic parameter.
In the context of host range, of the two principal strains, MDMV-A was found to be a johnsongrass-infecting strain, while MDMV-B is a non-johnsongrass-infecting strain [51]. MDMV-A was further differentiated from MDMV-B by the means of relative molecular mass (Mr) of RNA. Mr of MDMV-A RNA derived from three methods showed a value of 3.3 × 106 and for MDMV-B, it was reported as 3.0 × 106 [52]. Comparison in RNA nucleotide composition showed that both MDMV-A and MDMV-B strains consist of adenylic and guanylic acids in same quantity. Nevertheless, their high adenylic content in general adds credence to the suggestion that high content of adenylic acid might be a characteristic of potyviruses [53]. Furthermore, MDMV-A has a higher cytidylic acid content but lower uridylic acid content than MDMV-B [32].
3. Transmission
Aphids are the most widely spread vectors of plant viruses and have the potential to transmit both persistent and non-persistent viruses. Aphids transmit more than 200 species of plant viruses in a non-persistent way [54]. Among them, insects of the order Homoptera, with their piercing-sucking mouthparts, are highly efficient plant viral vectors. Within this order, the softbodied aphids (Family Aphididae) account for more than 50% of the vector species that transmit over 60% of the viruses [55]. Over 20 different aphid species are able to transmit MDMV non-persistently [56]. The broad range includes Rhopalosiphum maidis (Fitch) (Figure 1), Myzus persicae (Sulzer), Rhopalosiphum padi (Linnaeus), Rhopalosiphum poae (Gill.), Brevicoryne brassicae (Linnaeus) and Rhopalosiphum fitchii (Sand.) [57].
MDMV has a short acquisition access period (AAP) of 10–30 s [58]. Aphid transmission of the virus is closely correlated to the retention of the virus in stylets [59]. Berger et al. [60] also discovered that retention of MDMV was much longer when increasing the time of acquisition time. Previously, periods of retention around 15–20 min have been recorded [58,61]. However, longer periods of up to 240 min have also been recorded by for the retention of MDMV-A by M. persicae [62].
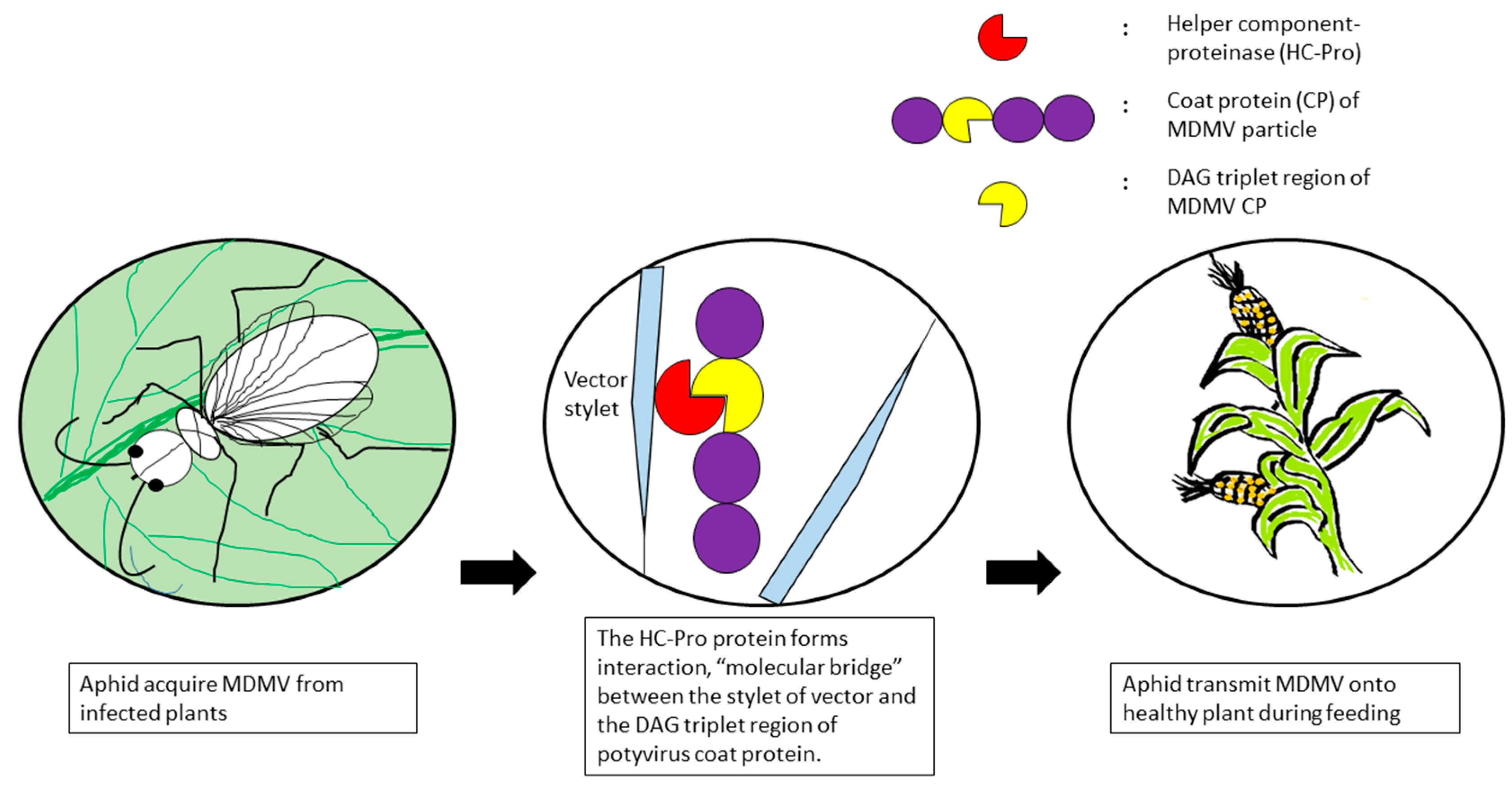
Appendages of aphids are where the virus particles have been retained either directly or indirectly before being inoculated into plants. There are two viral encoded factors that mediated the attachment of viruses to the appendages of aphids in indirect binding, the CP as the component of virion and the helper component-proteinase (HC-Pro) [63]. Helper component is a non-structural protein found in diseased plants but not in healthy tissues. The HC-Pro protein forms interactions between the stylet of vector and the virus coat protein, thus performs its function as a “molecular bridge” (Figure 2) [64,65]. Transmission of potyviruses is linked to a triplet 3-amino acid sequence, DAG (Asp–Ala–Gly), near the N-terminal region of the coat protein [66]. The simplest hypothesis to explain the behavioural difference of the N-terminus of the CP when it acts as a part of the virus particle or as free protein is that in the entire particle, the N-terminus is not available to interact with the aphid’s stylet and so the presence of HC-Pro is needed to cause a structural change, unfolding the N-terminus of the coat protein. Since the DAG triplet in the N-terminal region of the coat protein is crucial for aphid transmission [67], substitution of any of the 3 amino acid residues or the residue after the DAG motif will minimise aphid transmission drastically, but not mechanical transmission of the virus. Although the viral factors involved in transmission are quite clear, the receptors in aphids that allow retention as well as inoculation of non-persistent viruses remain unidentified but are believed to be localised at the distal edge of the stylet bundle [68,69].
Transmission occurs at different rates according to the influence of several factors. Firstly, fasted aphids transmit potyviruses more effectively (15%) compared to non-fasted aphids (5%) [70]. Fasting allows the interfering substances, plant components, to be egested or swallowed, clearing the alimentary canal, and hence enhances the virions retention in aphid’s stylet [71]. Furthermore, transmission of MDMV was positively related to leaf age, as the MDMV concentration was lower in older leaves, resulting in decreased aphid transmission [70]. It was found that a group of aphids improved MDMV transmission compared to single aphid transfer. This is probably due to the relatively short retention period and low transmission rate of MDMV. In addition, the efficiency of aphid transmission of MDMV is generally related to virus concentration in vivo in corn leaves. Therefore, any factors such as nutrition, host species, temperature, age of infection that could affect MDMV concentration in vivo in corn most likely would affect the efficiency of aphid transmission directly or indirectly [70]. It was also reported that the aphid species and virus strains greatly affect the efficiency of transmission [72]. Last but not least, both temperature and humidity perform an important function in the epidemiological study of MDMV. These factors greatly affect the efficiency and quantity of aphid vectors. For an example, S. graminum developed high populations under low temperature and low humidity early in the season, resulting in massive dispersal of viruliferous adults during the corn growing season [73].
Besides aphid transmission, MDMV can also be seed transmitted [20]. Seed transmission rates of up to 0.5% have been reported for the incidence of maize dwarf mosaic disease by Boothroyd et al. [74], and its expression appeared to be in two ways: as mosaic symptoms in certain plants and no symptoms in other infected plants. MDMV was present in 0.4% of field corn and sweet corn seeds [46] as well as in 0.2% of the tested maize inbred lines [75], both figures are based on symptom expression. Further reported rates for seed transmission in corn are 0.007% [76], 0.006% [77], 0.005% and non-existent [78]. For example, seeds dissected at different maturity stages and analysed for the presence of MDMV viruses through enzyme-linked immunosorbent assay (ELISA) and infectivity tests showed that at 21 days after pollination, MDMV was always discovered in the pericarp, but unfrequently in the endosperm or embryo [78]. The absence of virus infection in the embryo indicated limited or no seed transmission. The proportion of MDMV in seed parts declined from 100% at 21 days after pollination to rare at maturity in the pericarp. Upon reaching maturity, the pericarp transformed from a very moist nature to a hard, dry membrane. Thus, it is not unexpected since the virus within might have been inactivated. Infecting female gametophytes appeared to be the major MDMV seed transmission mechanism in sweet corn. MDMV was not detected in the male gametophytes, but was present in the unfertilized kernels, silk, glumes and whole anthers [77].
4. Symptom
Symptomatology of Maize dwarf mosaic virus has been described in several studies. In general, field plants infected by MDMV exhibit mosaic patterns [79], which normally initiate near the lower part of the youngest leaves and appear uneven and diffuse [18]. Mosaics formed by MDMV-A usually occur between leaf veins and so develop stripes [72]. Mosaics might develop as yellowish streaks that run throughout the edge of the leaf (Figure 1), and may disappear during hot weather and be replaced with common chlorosis in subsequent growth [18]. Chlorotic bands, or an “A” shape, occur when the chlorotic regions combine forming continuous streaks along the veins. Older plants develop only chlorotic indications on upper leaves and red streaks at times on mature leaves in late infections [73]. Other symptoms include mottling spots and irregular necrotic lesions [80]. During progression of earlier symptoms, dark and light green mottles appear on leaves. Formation of mosaics, flecks and rings on leaves is a consequence of an increase in intensity of dark and light green mottles as the disease develops [72]. Infected plant cells have been observed to consist of characteristic inclusion bodies, also known as cytoplasmic inclusions (CI), with closely associated cytoplasmic vesicles [81,82]. The CIs are composed of a virus encoded protein subunit with a relative molecular mass of 68 kDa [83].
Overall, infection at juvenile growth stages delays maturity and causes the loss of a large number of kernels at the basal end of the ear, which is usually known as butt blanking [23,24,84]. Occurrence of butt blanking is due to growth retardation of pollen germ tubes on diseased plants’ silks [85]. Stunting, reduction in plant weight [86], delay in silking up to four days [87], decrease in ear weight, ear diameter [88], head size and head numbers are also a part of MDMV symptomology. At times, corn plants infected with MDMV might show a delay in flowering as well as a poor grain set and fill [89].
5. Diagnostic Method
It is quite challenging to detect virus disease in corn by observing the symptoms alone, since symptoms vary according to plant genotype, infection time, condition of environment, and the potentiality for various infections. In consequence, observable diagnosis is validated using serological and molecular tests (Table 2). Available diagnostics for MDMV include commercialised enzyme-linked immunosorbent assays (ELISA) using antisera and reverse transcription-polymerase chain reaction (RT-PCR) involving virus genome sequence [3].
It is vital for the identification of MDMV to be confirmed using RT-PCR or ELISA. The advantage of RT-PCR is that it is much easier to acquire sequences from the database and thus information about the virus strain and basis of the viral isolates can be gathered [18]. RT-PCR procedure for MDMV strains begins with the grinding of maize infected sample in suitable extraction buffer, such as basic phenol or detergent mixture, followed by addition of ammonium acetate and chloroform-isoamyl alcohol and then centrifugation at low speed to obtain the nucleic acid containing supernatant [90]. Precipitate of the supernatant will be washed using 70% ethanol and resuspended, from which one microlitre of total RNA will be proceeded to the first cDNA synthesis [30]. This will be carried forward in order to perform RT-PCR using primers specifically designed for amplification of the available MDMV genome sequence. RT-PCR product will be sequenced and compared with known MDMV sequences. This will be carried out by conducting multiple sequence alignment (MSA). A previous study by Giolitti et al. [19] reported that the virus infecting maize fields in Chile is closely related to MDMV isolate from Argentina (MDMV-Arg), since RT-PCR conducted on those infected maize sample from Chile using primers designated for the gene MDMV-Arg capsid protein yielded only one band of the size, as expected.
For the serology part, the principle involved is simple. When an antigen is injected parenterally into the body of a rabbit followed by booster injections, it will trigger the release of particular antibodies. Rabbit will be then bled to obtain the blood serum containing immunoglobulins [88]. The antibodies respond to the antigen that activated their formation specifically in some observable ways. Plant viruses could be used as antigens, and in any plant suspected to have infection the presence of that particular antigen can be determined by using homologous antisera [98]. The ELISA method of serology is a hypersensitive technique for diagnosis of lower antigen concentrations in both crude and purified extract of viruses [99]. Its specificity is also useful for the differentiation of very closely related viral isolates [100]. ELISA tests are extremely cost effective as well as being relatively to use on an easy routine basis.
Although many variations have been developed, most investigators favour the ELISA type called double-antibody sandwich (DAS). A capture antibody is employed to coat a solid phase and is used to immobilize the virus in this assay. A second antibody, conjugated with an enzyme generally alkaline phosphatase is used to detect the immobilized virus by virtue of reaction with a substrate appropriate to the enzyme. The polyclonal antisera utilized in this assay is also often able to respond to antigens from uninfected plant extracts, since the avidity and affinity of the antibody molecule for the virus has been altered, which induced modification in their strain specificity [96]. This is due to the conformational changes in the polyclonal antibody molecule caused by conjugation of the enzyme to the second antibody [101,102]. This problem can often be circumvented by using capture and second antibody prepared in different animal species and detecting the second antibody with an enzyme-conjugated anti-species specific antibody [103]. However, specific antisera from different animal species are often unavailable. The unique characteristics of monoclonal antibodies offer the potential to develop a DAS-ELISA that avoids modifications of second antibody reactivity by molecular conjugation and does not require use of antisera raised in different animal species [92]. Almost all the monoclonal antibodies are specific to the homologous antigen. Utilization of monoclonal antisera for the detection of plant virus in diagnosis and epidemiology is a possible replacement for the use of polyclonal antibodies [96].
For this DAS-ELISA assay, results will be interpreted from the absorbance readings at 405 nm apart from examining the wells visually or with the aid of a plate reader for any colour change. Development of yellow colour in wells shows positive reactions, whilst negative reactions are demonstrated by no critical development of colour in the wells. Test values are considered to be valid if, and only if, wells of positive control result in a positive reaction while the healthy control and buffer containing wells stay clear. Samples of extract that give absorbance readings greater than two times the average of healthy controls could be read as a positive result [18]. Generally, for the positive results, similar values should be read in comparison of A405 nm values of the infected leaves sap to that with purified MDMV.
ELISA was carried out on hundreds of MDMV infected and non-infected corn plant saps, with a 100% correct result of the positive and negative controls. The sensitivity of ELISA test is 100 times higher compared to the regular tests of infectivity. Hence, application of ELISA for the diagnosis of plant viruses is much encouraged. This is also because the visual evaluation of the assay on the presence of viruses in plant tissue extracts is found to be reliable [92].
6. Control
6.1. Use of Insecticide and Johnsongrass Eradication
Once the presence of MDMV in plant has been detected, it is important to implement control strategies to reduce further yield loss. Arthropods play a role as a transmitting agent for all economically vital virus-induced diseases, including MDM disease in corn. Hence, occurrence of the disease requires three elements to be present simultaneously in a relevant environment: the virus, the transmitting vector and a vulnerable host [3]. One of the common approaches to control the spread of virus diseases in corn is by interrupting vector-maize interaction by reducing vector numbers on susceptible maize. This is possible by applying chemical insecticides or aphicides [104]. However, this method only restricts internal spread of the virus within a site and unfortunately affect the soil fertility [105]. Furthermore, previously Toler [106] demonstrated that MDM disease is relatively unaffected by the use of insecticides. Another common measure to control maize dwarf mosaic is breaking pathogen-vector and pathogen-maize interrelations by eliminating virus sources [3]. Johnsongrass, Sorghum halepense, is the main host of MDMV [107,108]. Yossen et al. [109] identified an MDMV isolate from johnsongrass and the plant can act as an overwintering host providing a shield for the virus [19]. Similarly, infection of MDMV in Spain is reported to be correlated with the abundance of johnsongrass [110]. Therefore, eradication of johnsongrass could effectively limit virus spread [111]. However, it is extremely difficult to do this in fields continuously cropped with maize. Thus, it is advisable to carry out crop rotation with a non-gramineous crop like soy bean, Glycine max, in fields with a perennial johnsongrass problem or a history of maize virus diseases. This has special merit with the advent of new “over-the-top” graminicides that can be used throughout the season in soybean fields to eradicate johnsongrass, with no residue carryover the following year. In addition, troublesome fields could be left fallow for half a season and an intensive effort at johnsongrass eradication could be attempted [73]. Apart from johnsongrass, the mature maize crop has the potential to act as a reservoir of virus for a newly cultivated crop, again serving as a transmitting agent source for viruses [112,113]. In this case, postponing the date of planting maize to prevent the abundancy of vector populations could allow some control over the disease.
6.2. S-Methylmethionine Pre-Treatment
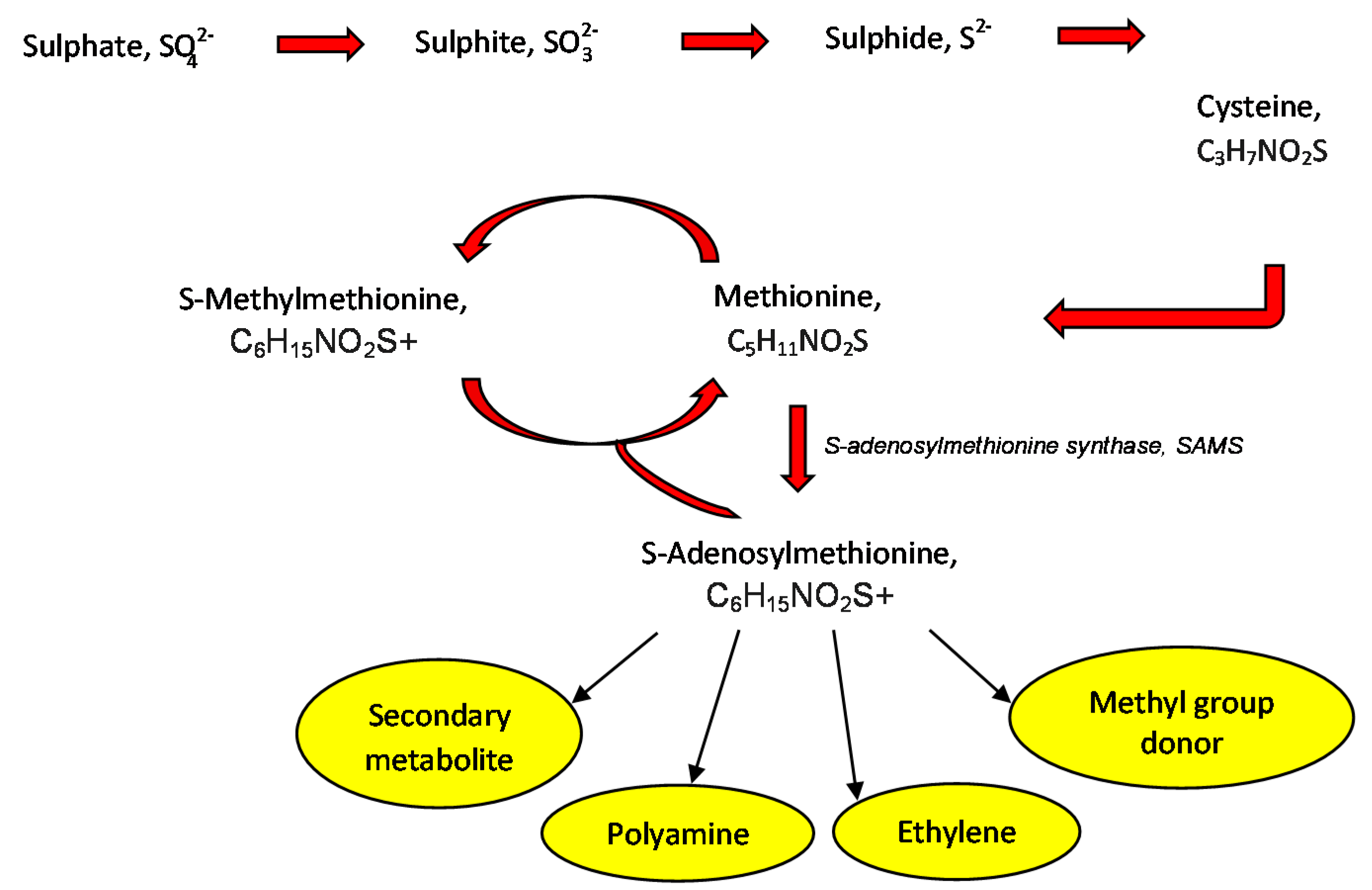
Plants are specified with sulphur autotrophy, a series of actions whereby sulphur is absorbed in the oxidised forms from the land, then undergo reduction before being added to cysteine and methionine. The final compound could be converted into S-Adenosylmethionine (SAM), a substrate that takes part as a methyl donor in the synthesis of S-Methylmethionine (SMM) derived from methionine [114,115]. SMM [(CH3)2-S-(CH2)2-CH(NH2)-COOH] naturally exists in the kingdom plantae as a non-coded and sulphur consisting amino acid [1]. SMM could be converted back into methionine through in a cyclic pathway, called the SMM cycle [116] (Figure 3).
SMM confers resistance in plants, being a direct precursor of osmoprotectant sulfopropionates, and it also stimulates the anabolism of other regulatory and protective compounds such as polyamine and ethylene [117,118]. When stress factors are present, the up-regulation of the phenylpropanoid biosynthetic pathway is detected, and that contributes to the production of certain fenoloids, flavonoids and anthocyanins, which are compounds characterised by antioxidant quality and show higher absorbance in the UV spectrum [119]. The potential of SMM has been proven not only against abiotic, but also against biotic stresses, since SMM-treated plants demonstrated higher resistance against Maize dwarf mosaic virus infection. The expression changes of 14-3-3-like protein gene G-box factor 14-6, GF14-6 and S-adenosylmethionine synthase (SAMS) were studied in Ludsmerszki et al. [1] during MDMV infection. The product of GF14-6 recognises and degrades the viral coat protein, contributes to RNA silencing. In the SMM pre-treated and afterwards infected plants, a decline in the GF14-6 expression indicates the improved plant defence due to SMM pre-treatment. Additionally, the more prolonged and long-lasting increment is measured in SAMS expression resulting from SMM pre-treatment followed by the infection indicates the importance of the gene product in upregulating SAM formation for methylation processes, thus raising the SMM circular pathway, and further contributing to defence compounds production [1] (Figure 4). The production of defence compounds due to SMM pre-treatment is further supported in the study by [119]; the study showed an incremental increase in the intensity of emitted fluorescence at 440 and 520 nm after plants were treated by SMM, which indicates that the amount of phenoloids increased in these plants. Kocsis et al. [120] demonstrated that during stress conditions SMM prevents chlorophyll loss. The value of the F690/F740 ratio is inversely proportional with a decrease in the amount of chlorophyll localised in the leaves, indicating the increase in chlorophyll content when the plants were treated by SMM. Similar results were achieved with chlorophyll-a fluorescence induction measurements. The Fv/Fm value, that indicates the physiological status of the plant, more specifically the maximal quantum efficiency of PSII decreased in the case of infected plants, was analysed but no significant change was detected when the infected plants also got SMM treatment prior to infection [119].
Since MDMV disrupts the thylakoid [121] and any damage in thylakoid membranes causes the development of highly reactive oxygen species (ROS), this is an area of research focus. Compared to other species of plants, maize vascular bundle sheath cells are unusually sensitive to oxidative stress [122,123]. This is solved by the existence of antioxidant enzymes, primarily ascorbate peroxidase (APX) and guaiacol peroxidase (GPX) in plant cells, which protect the cell through direct scavenging of those reactive molecules [124]. When the combined molecule of SMM and salicyclic acid (SA), known as S-Methylmethionine salicylate (MMS), was applied before infection, lower levels of enzyme activity were recorded, indicating a decreased resistance level, which can be explained by the reduction in the quantity of virus particles due to MMS pre-treatment [125] (Figure 5).
During MDMV infection, the viral particles accumulate in the cytoplasm of the leaf mesophyll cells and may use chloroplasts for their replication [126,127]. The associations between virus coat protein with PSII reaction centres in thylakoid membranes lead to formation of non-fluorescent trap for excitational energy which subsequently will cause an increase in non-photochemical quenching [128]. Accordingly, Ludmerszki et al. [125] reported greater ΔpH dependent process high energy state, qE values for MDMV-infected plants which indicated the appearance of additional quenches of fluorescence in those plants. However, when infected plants were pre-treated with MMS, low qE values were measured, indicating that virus coat protein-thylakoid membrane associations were not formed. This is supported by the ELISA results, showing smaller concentration of MDMV coat protein and viral RNA in the MMS + MDMV plants, giving a clarification for the lower qE values. In general, the protecting effect of SMM treatment for maize plants prior to stress of MDMV infection is being demonstrated in clear entirely.
6.3. Conventional Resistance Breeding
The most effective way to protect corn crops against MDMV is through the breeding of maize lines for resistance [129,130]. Having a limited knowledge of resistant genes and the pathogens, corn breeders and co-operating pathologists with entomologists suggested that the initial step for this method is the identification of resistance sources through screening of the collected germplasm [3]. The resistance screening methods are comprised of natural infection under field conditions, vector inoculation, mechanical inoculation and growing under greenhouse conditions.
Initially, susceptible inbred lines are crossed with resistant ones, and the F1 generation is produced. Some F1 generations are selfed to result in F2 generations, others are back-crossed with susceptible parents to produce BC1 generations [131]. Backcrosses have usually been utilized to transfer virus resistance from resistant, but agronomically undesirable, lines to well-adapted elite lines. The recurrent parent is commonly selected due to its ability to combine well, produce high yield and adapt to wider changes [132]. Selection in F2 and BC1 generations seems to be a favourable method to breed new virus-resistant inbred lines. Such selection for resistance under higher temperatures in the greenhouse normally results in the lines with higher resistance level. Accordingly, three dent inbred lines (D21, D32, FAP136OA) with complete resistance, and four dent inbreds (D06, D09, R2306, FAP1396A) with partial resistance against MDMV under both field and greenhouse conditions were determined. Apart from these, the inbred line Pa405 has been found to be the best known source of resistance to MDMV to date [133,134].
Successful screening is continued with identification of markers linked to genes or QTLs exerting resistance, which can be done using reciprocal translocations [135,136], morphological markers [137] and restriction fragment length polymorphism (RFLP) analysis [138,139]. RFLP analysis of individual back cross plants of the genotypes (Pa405 × yM14 and (Pa405 × K55) × K55, inoculated with MDMV, mapped this resistance gene to a region close to the centromere of chromosome 6. This gene is linked tightly to and located between RFLP marker loci, UMC85 and BNL6.29, and designated as gene Mdm1, gene 1 [140]. Mdm1 is crucial for any resistance reaction because all plants with missing Mdm1 gene rapidly developed symptoms of generalised mosaics. The resistance dominance of Mdm1 was further confirmed in 42 of the 43 inbred lines analysed in the study by [141].
The inheritance of resistance to maize dwarf mosaic disease is also controlled by modifiers or minor genes [142]. For an example, when F1 hybrids, F2 progeny and a recombinant inbred line (RIL) population obtained from a crossing of Oh1V1 to the virus susceptible line Oh28 were examined for their responses to six viruses including MDMV, a dominant QTL responsible for 79% of total variance and several minor QTLs each contributing to 1% of the variance mapped to chromosome 3, and 10 were identified [143]. Moreover, the data representing the percentage of infected plants denoting critical differences both within F1 generations (from 18 to 34%) and within BC1 generations (from 26 to 53%) showed that resistance to MDMV is controlled by major genes, yet the minor genes also involved in modification of the resistance [131]. These hybrids and inbred lines with genes for resistance to MDM do not show asymptomatic reactions after infections constantly [133,144]. Scott and Rosenkranz [145] mentioned that symptomatic responses maybe resulted from micro environmental influences, expression of resistance at late growth stages, concentrations of inoculum above a specific threshold. Both symptomatic tissue with high virus concentration and asymptomatic tissue with no virus detected formed distinct sectors in resistant hybrids [146]. Similarly, ELISA confirmed existence of both inoculated leaves with a positive response for virus and freshly developed leaves with no virion assembly in the same resistant inbred [147].
This indicates that MDMV can undergo replication and spread from cell to cell in leaves of infected plants, but that a barrier to systemic viral propagation is present in resistant plants. Thus, in resistant plants, the virus is restrained in its movement through the leaf vascular system [147]. The transfer of the resistance from resistant hybrids such as Pioneer Brand (PB) 3187 [148] into selected breeding lines will lead to similar resistance mechanisms and result in successful control over maize dwarf mosaic disease.
6.4. Genetically Engineered Resistance
Pathogen-derived resistance, which can be achieved through expression of resistant genes (Table 3), viral proteins or RNAs in transgenic plants through genetic engineering, is another strategy to curb maize dwarf mosaic disease in maize crops [149,150]. For this way of controlling, it is important to acquire knowledge on the concept of resistance expression genetically. As explained earlier, the virus resistance in maize was due to an obstruction to systemic movement of virus in resistant plants [151] where the movement could be prevented by inhibition of the loading of virions, ribonucleoprotein [152], or RNA into phloem or unloading from phloem in either leaves or roots [148]. However, for a better understanding on the mechanism for genetic resistance, global transcriptional response of resistant (Pa405) and susceptible (Oh28) inbred lines was analysed in germinating maize embryos at 4 days post inoculation (dpi). There were no groupings of differentially expressed transcripts found in the previously recognised QTLs crucial for MDMW resistance, on chromosome 6, 3 and 10 [153]. Almost 15% of transcripts were differently expressed between the resistant and susceptible lines. Of transcripts with more than 10-fold greater expression in one line, more than 70% were upregulated in the resistant line. Hsp20/alpha crystalline-like gene and cytochrome P450 are among the differentially regulated transcripts that were explained to be involved in virus resistance. The obtained responsive transcripts required to be further studied on in order to figure out their contribution for resistance [154].
Another effort of cloning Mdm1 allele by chromosome walking was taken to explain the biological basis of Mdm1 mediated resistance in resistant maize [156,157]. Although co-segregation of Mdm1 with the nucleolus organizer region (nor) in maize caused a difficult obstacle to clone Mdm1, yet this initiative still led to the achievement of a high-resolution genetic map in the region of Mdm1 that has raised tools for physical mapping of Mdm1, which is very useful for subsequent genetic engineering procedures [158].
H9-21 is one of the incorporated resistant germplasms against MDMV previously. However, it does not achieve the rate of maize production required due to its inefficient agronomic characteristics [159]. The attempts to genetically improve resistance against MDMV in maize crops were then continued with the development of engineered lines in order to generate cross protection. Cross protection can be triggered by introducing an antisense sequence homologous to MDMV genes of coat protein (CP), replicase and movement proteins that play a role in viral replication, spreading, and particle coating in the single-standard RNA viral genome [160,161]. Among these genes, maize plants transformed with MDMV protease gene (P1), a replication associated protein gene, proved to be resistant to MDMV under controlled conditions [162]. This was possible by the means of RNA interference (RNAi), mediated by self-complementary hairpin RNA (hpRNA), transcribed from a transgenic inverted-repeat sequence. RNAi has been proved to be more useful in conferring virus resistance through gene silencing due to its straight-forward natural defense mechanism [163,164,165]. MDMV resistance generated by RNA interference depends on the length of inverted-repeat sequence, the copy number of T-DNA integration and the repeatability of integration sites [166]. A longer hpRNA is more successful in the virus gene silencing rather than a shorter one [167,168]. Previously, the transfer of 150bp inverted-repeat sequence homologous to MDMV P1 enhanced maize virus resistance yet resulted in T2 lines with intermediate resistance or lower compared to the resistant control. Then, MDMV resistance was further improved through development of an hpRNA expression vector consisting of 451bp inverted-repeat sequences of MDMV protease gene (P1) [169]. Generally, genetic improvement of maize is considered to be a more efficient and more environmentally sustainable measure for virus management [87,170].
7. Concluding Remarks
Detailed studies conducted on the genome and symptomatology of MDMV have resulted in successful visual, serological, as well as molecular, tests of diagnosis. This contributes to efforts of controlling MDM disease in order to reduce the yield loss caused in corn crops. The methods that were utilized in circumventing this problem include eradication of johnsongrass, SMM pre-treatment, resistance breeding and genetic engineering. The efficiency of currently available measures for curbing the spread of MDM disease has been proved, shown by the now rare infection of MDMV being observed in field corn. However, further initiatives can be focused on understanding of maize resistant genes using omics technology approach against MDMV for a better management of MDM disease of maize worldwide.
Author Contributions
Writing—Original Draft Preparation, M.K.; Writing—Review & Editing, M.K., H.B. and I.I. Visualization, M.K. and H.B.; Project Administration, H.B. and I.I.; Funding Acquisition, H.B. and I.I.
Funding
This research was funded by Universiti Kebangsaan Malaysia grant numbers GGPM-2017-072 and DPP-2018-010. The APC was funded by Universiti Kebangsaan Malaysia.
Acknowledgments
The authors would like to thank Kathryn Ford from Bristol University for her editorial comments to improve the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ludmerszki, E.; Rácz, I.; Rudnóy, S. S-methylmethionine alters gene expression of candidate genes in Maize dwarf mosaic virus infected and drought stressed maize plants. Acta Biol. Szeged. 2014, 58, 1–5. [Google Scholar]
- Lukuyu, B.; Murdoch, A.; Mcleod, A.; Doward, P. The impact of Maize streak virus disease on quality and yield of maize forage outputs of the maize-dairy production systems in central highlands of Kenya. In Proceedings of the Livestock Development Group Student Conference, Focus on Livelihoods, University of Reading, Reading, UK, 16 October 2004; p. 16. [Google Scholar]
- Redinbaugh, M.G.; Zambrano, J.L. Control of Virus Diseases in Maize. Adv. Virus Res. 2014, 90, 391–429. [Google Scholar] [CrossRef] [PubMed]
- Food and Agriculture Organizations. FAO Statistical Yearbook; Food and Agriculture Organization of the United Nations: Rome, Italy, 2013; ISBN 9788578110796. [Google Scholar]
- Lapierre, H.; Signoret, P.A. Viruses and Virus Diseases of Poaceae (Gramineae); Institut National de la Recherche Agronomique: Paris, France, 2004; ISBN 2738010881. [Google Scholar]
- Shukla, D.D.; Tosic, M.; Jilka, J.; Ford, R.E.; Toler, R.W.; Langham, M.A.C. Taxonomy of Potyviruses Infecting Maize, Sorghum, and Sugarcane in Australia and the United States as Determined by Reactivities of Polyclonal Antibodies Directed towards Virus-Specific N-Termini of Coat Proteins. Phytopathology 1989, 79, 223. [Google Scholar] [CrossRef]
- Shukla, D.; Ward, C. Structure of Potyvirus Coat Proteins and Its Application in the Taxonomy of the Potyvirus Group. Adv. Virus Res. 1989, 36, 273–314. [Google Scholar] [CrossRef] [PubMed]
- Seifers, D.L.; Salomon, R.; Marie-Jeanne, V.; Alliot, B.; Signoret, P.; Haber, S.; Loboda, A.; Ens, W.; She, Y.M.; Standing, K.G. Characterization of a novel potyvirus isolated from maize in Israel. Phytopathology 2000, 90, 505–513. [Google Scholar] [CrossRef] [PubMed]
- Schurtleff, M.C. Compendium of Corn Diseases; APS Press: St. Paul, MN, USA, 1999; ISBN 0890540292. [Google Scholar]
- Gell, G.; Balázs, E.; Petrik, K. Genetic diversity of Hungarian Maize dwarf mosaic virus isolates. Virus Genes 2010, 40, 277–281. [Google Scholar] [CrossRef] [PubMed]
- Gordon, D.T.; Bradfute, O.E.; Gingery, R.E.; Knoke, J.K.; Louie, R.; Nault, L.R.; Scott, G.E. Introduction: History, geographical distribution, pathogen characteristics, and economic importance. In Viruses and Viruselike Disease of Maize in the United States; Ohio State University: Wooster, OH, USA, 1981; pp. 1–12. [Google Scholar]
- Lübberstedt, T.; Ingvardsen, C.; Melchinger, A.E.; Xing, Y.; Salomon, R.; Redinbaugh, M.G. Two chromosome segments confer multiple potyvirus resistance in maize. Plant Breed. 2006, 125, 352–356. [Google Scholar] [CrossRef]
- Lesemann, D.E.; Shukla, D.D.; Tosic, M.; Huth, W. Differentiation of the four viruses of the Sugarcane mosaic virus subgroup based on cytopathology. Potyvirus Taxon. 1992, 5, 353–361. [Google Scholar] [CrossRef]
- Zielińska, L.; Trzmiel, K.; Jezewska, M. Ultrastructural changes in maize leaf cells infected with Maize dwarf mosaic virus and Sugarcane mosaic virus. Acta Biol. Crac. Ser. Bot. 2012, 54, 97–104. [Google Scholar] [CrossRef]
- Matthews, R. The classification and nomenclature of viruses: Summary of results of meetings of the International Committee on Taxonomy of Viruses in Strasbourg. Intervirology 1982, 16, 53–60. [Google Scholar] [CrossRef]
- Revers, F.; García, J.A. Molecular biology of potyviruses. Adv. Virus Res. 2015, 92, 101–199. [Google Scholar] [CrossRef] [PubMed]
- Tosic, M.; Ford, R.E.; Shukla, D.D.; Jilka, J. Differentiation of Sugarcane, Maize dwarf, Johnsongrass, and Sorghum mosaic viruses based on reactions of oat and some sorghum cultivars. Plant Dis. 1990, 74, 549–552. [Google Scholar] [CrossRef]
- Cao, P.; Eckstein, H.H.; De Rango, P.; Setacci, C.; Ricco, J.B.; de Donato, G.; Becker, F.; Robert-Ebadi, H.; Diehm, N.; Schmidli, J.; et al. Diagnostic Methods. Eur. J. Vasc. Endovasc. Surg. 2011, 42, S13–S32. [Google Scholar] [CrossRef]
- Giolitti, F.; Herrera, M.G.; Madariaga, M.; Lenardon, S.L. Detection of Maize dwarf mosaic virus (MDMV) on maize in Chile. Maydica 2005, 50, 101–104. [Google Scholar]
- Chauhan, R.A. Study of Filamentous Viruses in Maize and Smallgrains. Master’s Thesis, University of Cape Town, Caoe Town, South Africa, 1985. [Google Scholar]
- Gingery, R.E. Chemical and physical properties of maize viruses. South. Coop. Ser. Bull. 1981, 247, 38–39. [Google Scholar]
- Shi, C.L.; Hsu, S.H. Maize dwarf mosaic virus of corn and sorghum in Beijing. Acta Phytopathol. Sin. 1979, 9, 35–40. [Google Scholar]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Yield response of sweet corn to Maize dwarf mosaic virus. Plant Dis. 1981, 65, 900–901. [Google Scholar] [CrossRef]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Yield loss in sweet corn correlated with time of inoculation with Maize dwarf mosaic virus. Plant Dis. 1981, 65, 902–904. [Google Scholar] [CrossRef]
- Gates, D.W.; Gudauskas, R.T. Photosynthesis, respiration and evidence of a metabolic inhibitor in corn infected with Maize dwarf mosaic virus. Phytopathology 1969, 59, 575–580. [Google Scholar]
- Achon, M.A.; Larrañaga, A.; Alonso-Dueñas, N. The population genetics of Maize dwarf mosaic virus in Spain. Arch. Virol. 2012, 157, 2377–2382. [Google Scholar] [CrossRef] [PubMed]
- Kong, P.; Steinbiss, H.H. Complete nucleotide sequence and analysis of the putative poliprotein of Maize dwarf mosaic virus genomic RNA (Bulgarian isolate). Arch. Virol. 1998, 143, 1791–1799. [Google Scholar] [CrossRef] [PubMed]
- Urcuqui-Inchima, S.; Haenni, A.L.; Bernardi, F. Potyvirus proteins: A wealth of functions. Virus Res. 2001, 74, 1–2. [Google Scholar] [CrossRef]
- Tuo, D.; Shen, W.; Yan, P.; Li, X.; Zhou, P. Rapid Construction of Stable Infectious Full-Length cDNA Clone of Papaya leaf distortion mosaic virus Using In-Fusion Cloning. Viruses 2015, 7, 6241–6250. [Google Scholar] [CrossRef] [PubMed]
- Marie-Jeanne, V.; Ioos, R.; Peyre, J.; Alliot, B.; Signoret, P. Differentiation of Poaceae potyviruses by reverse transcription-polymerase chain reaction and restriction analysis. J. Phytopathol. 2000, 148, 141–151. [Google Scholar] [CrossRef]
- Shukla, D.D.; Strike, P.M.; Tracy, S.L.; Gough, K.H.; Ward, C.W. The N and C termini of the coat proteins of potyviruses are surface-located and the N terminus contains the major virus-specific epitopes. J. Gen. Virol. 1988, 69, 1497–1508. [Google Scholar] [CrossRef]
- Hill, J.H.; Ford, R.E.; Benner, H.I. Purification and partial characterization of Maize dwarf mosaic virus strain B (Sugarcane mosaic virus). J. Gen. Virol. 1973, 20, 327–339. [Google Scholar] [CrossRef]
- Petrik, K.; Sebestyén, E.; Gell, G.; Balázs, E. Natural insertions within the N-terminal region of the coat protein of Maize dwarf mosaic potyvirus (MDMV) have an effect on the RNA stability. Virus Genes 2010, 40, 135–139. [Google Scholar] [CrossRef] [PubMed]
- Olspert, A.; Chung, B.Y.W.; Atkins, J.F.; Carr, J.P.; Firth, A.E. Transcriptional slippage in the positive-sense RNA virus family Potyviridae. EMBO Rep. 2015, 16, 995–1004. [Google Scholar] [CrossRef] [PubMed]
- Chung, B.Y.W.; Miller, W.A.; Atkins, J.F.; Firth, A.E. An overlapping essential gene in the Potyviridae. Proc. Natl. Acad. Sci. USA 2008, 105, 5897–5902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillung, J.; Elena, S.F.; Cuevas, J.M. Intra-specific variability and biological relevance of P3N-PIPO protein length in potyviruses. BMC Evol. Biol. 2013, 13, 249. [Google Scholar] [CrossRef] [PubMed]
- Vijayapalani, P.; Maeshima, M.; Nagasaki-Takekuchi, N.; Miller, W.A. Interaction of the trans-frame potyvirus protein P3N-PIPO with host protein PCaP1 facilitates potyvirus movement. PLoS Pathog. 2012, 8, e1002639. [Google Scholar] [CrossRef] [PubMed]
- Masumi, M.; Mostafavi Neishabouri, F.; Nasrollahnejad, S.; Ghahramani, T.; Izadpanah, K. Determination of the Complete Sequence of the Genome of Maize dwarf mosaic virus; Plant Virology Research Center, College of Agriculture, Shiraz University: Shiraz, Iran, 2011. [Google Scholar]
- Achon, M.A.; Serrano, L.; Alonso-Dueñas, N.; Porta, C. Complete genome sequences of Maize dwarf mosaic and Sugarcane mosaic virus isolates coinfecting maize in Spain. Arch. Virol. 2007, 152, 2073–2078. [Google Scholar] [CrossRef] [PubMed]
- Stewart, L.R.; Bouchard, R.; Redinbaugh, M.G.; Meulia, T. Complete sequence and development of a full-length infectious clone of an Ohio isolate of Maize dwarf mosaic virus (MDMV). Virus Res. 2012, 165, 219–224. [Google Scholar] [CrossRef] [PubMed]
- Maize Dwarf Mosaic Virus Isolate Italy, Complete Genome. Available online: https://0-www-ncbi-nlm-nih-gov.brum.beds.ac.uk/nuccore/402216197 (accessed on 16 August 2018).
- Boyer, J.C.; Haenni, A.L. Infectious Transcripts and cDNA Clones of RNA Viruses. Virology 1994, 198, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Duff-farrier, C.R.A.; Candresse, T.; Bailey, A.M.; Boonham, N.; Foster, G.D. Short Communication Evidence for different, host-dependent functioning of Rx against both wild-type and recombinant Pepino mosaic virus. Mol. Plant Pathol. 2016, 17, 120–126. [Google Scholar] [CrossRef] [PubMed]
- Gell, G.M. Molecular Analysis of the Coat Protein Genes of Maize dwarf mosaic virus (MDMV) Populations. Ph.D. Thesis, University of Szent Istvan, Godollo, Hungary, 2011. [Google Scholar]
- Ali, M.C.; Omar, A.S.; Natsuaki, T. An infectious full-length cDNA clone of Potato virus YNTN-NW, a recently reported strain of PVY that causes potato tuber necrotic ringspot disease. Arch. Virol. 2011, 156, 2039–2043. [Google Scholar] [CrossRef]
- Shepherd, R.J.; Holdeman, Q.L. Seed transmission of the Johnsongrass strain of the Sugarcane mosaic virus in Corn. Plant Dis. Report. 1965, 49, 468–469. [Google Scholar]
- Snazelle, T.E.; Bancroft, J.B.; Ullstrup, A.J. Purification and serology of Maize dwarf mosaic and Sugarcane mosaic viruses. Phytopathology 1971, 61, 1059–1063. [Google Scholar] [CrossRef]
- Langenberg, W.G. Leaf-dip serology for the determination of strain relationships of elongated plant viruses. Phytopathology 1974, 64, 128–131. [Google Scholar] [CrossRef]
- Szirmai, J. The occurrence of stripe mosaic disease of maize in Hungary and possibilities of breeding for virus resistance. Acta Phytopathol. Hung. 1968, 3, 189–198. [Google Scholar]
- Shukla, D.D.; Jilka, J.; Tosic, M.; Ford, R.E. A novel approach to the serology of potyviruses involving affinity-purified polyclonal antibodies directed towards virus-specific N termini of coat proteins. J. Gen. Virol. 1989, 70, 13–23. [Google Scholar] [CrossRef]
- Seifers, D.L.; Hackerott, H.L. Estimates of yield loss and virus titre in sorghum hybrids infected with Maize dwarf mosaic virus strain B. Agric. Ecosyst. Environ. 1987, 19, 81–86. [Google Scholar] [CrossRef]
- Hill, J.H.; Benner, H.I. Properties of potyvirus RNAs: Turnip mosaic, Tobacco etch, and Maize dwarf mosaic viruses. Virology 1976, 75, 419–432. [Google Scholar] [CrossRef]
- Hill, J.H.; Shepherd, R.J. Biochemical properties of Turnip mosaic virus. Virology 1972, 47, 807–816. [Google Scholar] [CrossRef]
- Moury, B.; Fabre, F.; Senoussi, R. Estimation of the number of virus particles transmitted by an insect vector. Proc. Natl. Acad. Sci. USA 2007, 104, 17891–17896. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- D’Arcy, C.J.; Nault, L.R. Insect transmission of plant viruses and mycoplasmalike and rickettsialike organisms. Plant Dis. 1982, 66, 99–104. [Google Scholar] [CrossRef]
- Knoke, J.K.; Louie, R. Epiphytology of maize virus diseases. South. Coop. Ser. Bull. 1981, 247, 92–102. [Google Scholar]
- Diagnostis Methods for Maize dwarf mosaic virus. Available online: http://www.padil.gov.au/pbt (accessed on 4 January 2018).
- Bancroft, J.B.; Ullstrup, A.J.; Messieha, M.; Bracker, C.E.; Snazelle, T.E. Some biological and physical properties of a midwestern isolate of Maize dwarf mosaic virus. Phytopathology 1966, 56, 474–478. [Google Scholar]
- Wang, R.Y.; Pirone, T.P. Potyvirus transmission is not increased by pre-acquisition fasting of aphids reared on artificial diet. J. Gen. Virol. 1996, 77, 3145–3148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger, P.; Toler, R.; Harris, K. Evidence for 2 modes of retention of Maize dwarf mosaic virus by Schizaphis graminum. Phytopatholog 1983, 73, 787. [Google Scholar]
- Messieha, M. Studies on aphid transmission of Maize dwarf mosaic virus. Phytopathology 1967, 57, 956–959. [Google Scholar]
- Thongmeearkom, P.; Ford, R.E.; Jedlinski, H. Aphid transmission of Maize dwarf mosaic virus strains. Phytopathology 1976, 66, 332–335. [Google Scholar] [CrossRef]
- Salomon, R.; Bernardi, F. Inhibition of viral aphid transmission by the N-terminus of the Maize dwarf mosaic virus coat protein. Virology 1995, 213, 676–679. [Google Scholar] [CrossRef] [PubMed]
- Pirone, T.P.; Blanc, S. Helper-dependent vector transmission of plant viruses. Annu. Rev. Phytopathol. 1996, 34, 227–247. [Google Scholar] [CrossRef] [PubMed]
- Lacomme, C.; Glais, L.; Bellstedt, D.U.; Dupuis, B.; Karasev, A.V.; Jacquot, E. Virus Y: Biodiversity, Pathogenicity, Epidemiology and Management; Springer International Publishing: Cham, Switzerland, 2017; pp. 1–261. [Google Scholar]
- Nault, L.R. Arthropod Transmission of plant viruses: A new synthesis. Ann. Entomol. Soc. Am. 1997, 90, 521–541. [Google Scholar] [CrossRef]
- Atreya, C.D.; Raccah, B.; Pirone, T.P. A point mutation in the coat protein abolishes aphid transmissibility of a potyvirus. Virology 1990, 178, 161–165. [Google Scholar] [CrossRef]
- Jiang, Y.X.; Lei, H.; Collar, J.L.; Martin, B.; Muniz, M.; Fereres, A. Probing and feeding behavior of two distinct biotypes of Bemisia tabaci (Homoptera: Aleyrodidae) on tomato plants. J. Econ. Entomol. 1999, 92, 357–366. [Google Scholar] [CrossRef]
- Wang, R.Y.; Ammar, E.D.; Thornbury, D.W.; Lopez-Moya, J.J.; Pirone, T.P. Loss of potyvirus transmissibility and helper-component activity correlate with non-retention of virions in aphid stylets. J. Gen. Virol. 1996, 77, 861–867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tu, J.C.; Ford, R.E. Factors affecting aphid transmission of Maize dwarf mosaic virus to corn. Phytopathology 1971, 61, 1516–1521. [Google Scholar] [CrossRef]
- Watson, M.A.; Roberts, F.M. A comparative study of the transmission of Hyoscyamus virus 3, Potato virus Y and Cucumber virus 1 by the vectors Myzus persicae (Sulz), M. circumflexus (Buckton), and Macrosiphum gei (Koch). Proc. R. Soc. Lond. B 1939, 127, 543–576. [Google Scholar] [CrossRef]
- Tsai, J.H.; Brown, L.G. Maize Dwarf Mosaic Virus; Plant Pathology Circular No. 320; Division of Plant Industry: Gainesville, FL, USA, 1989. [Google Scholar]
- Teyssandier, E.E.; Nome, S.F.; Bo, E. Maize virus diseases in Argentina. In Proceedings of the International Maize Virus Disease Colloquium and Workshop, Wooster, OH, USA, 2–6 August 1982; The Ohio State University: Wooster, OH, USA, 1982; pp. 93–99. [Google Scholar]
- Boothroyd, C.W. Seed transmission of Maize dwarf mosaic virus in sweet corn and yield reduction in plants from an infected seed lot. Am. Phytopathol. Soc. 1977, 4, 184. [Google Scholar] [CrossRef]
- Hill, J.H.; Martinson, C.A.; Russell, W.A. Seed Transmission of Maize dwarf mosaic and Wheat streak mosaic viruses in Maize and Response of Inbred Lines. Crop Sci. 1974, 14, 232–235. [Google Scholar] [CrossRef]
- Williams, L.E.; Findley, W.R.; Dollinger, E.J.; Ritter, R.M. Seed transmission studies of Maize dwarf mosaic virus in corn. Plant Dis. Report. 1968, 52, 863–864. [Google Scholar]
- Mikel, M.A.; D’Arcy, C.J.; Ford, R.E. Seed Transmission of Maize dwarf mosaic virus in Sweet Corn. J. Phytopathol. 1984, 110, 185–191. [Google Scholar] [CrossRef]
- Panayotou, P.C. Investigations on seed transmission of Maize dwarf mosaic virus and its effect on the establishment of seedlings. J. Plant Dis. Prot. 1981, 88, 621–625. [Google Scholar]
- Nault, L.R.; Gordon, D.T.; Loayza, J.C. Maize virus and mycoplasma diseases in Peru. Trop. Pest Manag. 1981, 27, 363–369. [Google Scholar] [CrossRef]
- Jones, M.W.; Boyd, E.C.; Redinbaugh, M.G. Responses of maize (Zea mays L.) near isogenic lines carrying Wsm1, Wsm2, and Wsm3 to three viruses in the potyviridae. Theor. Appl. Genet. 2011, 123, 729–740. [Google Scholar] [CrossRef] [PubMed]
- Ford, R.E.; Tosic, M.; Shukla, D.D. Maize Dwarf Mosaic Virus: AAB Descriptions of Plant Viruses Online. 1989. Available online: http://sdb.im.ac.cn/vide/descr465.htm (accessed on 28 January 2018).
- Krass, C.J.; Ford, R.E. Ultrastructure of Corn systemically infected with Maize dwarf mosaic virus. Phytopathology 1969, 59, 431–439. [Google Scholar]
- Chen, S.; Das, P.; Hari, V. In situ localization of ATPase activity in cells of plants infected by Maize dwarf mosaic potyvirus. Arch. Virol. 1994, 134, 433–439. [Google Scholar] [CrossRef] [PubMed]
- Gregory, L.V.; Ayers, J.E. Effect of inoculation with Maize dwarf mosaic virus at several growth stages on yield of sweet corn. Plant Dis. 1982, 66, 801–804. [Google Scholar] [CrossRef]
- Mikel, M.A.; D’Arcy, C.J.; Rhodes, A.M.; Ford, R.E. Effect of Maize dwarf mosaic virus infection on sweet corn pollen and silk. Phytopathology 1982, 72, 428–431. [Google Scholar] [CrossRef]
- Fuchs, E.; Grüntzig, M. Influence of Sugarcane mosaic virus (SCMV) and Maize dwarf mosaic virus (MDMV) on the growth and yield of two maize varieties. J. Plant Dis. Prot. 1995, 102, 44–50. [Google Scholar]
- Williams, M.M.; Pataky, J.K. Interactions between maize dwarf mosaic and weed interference on sweet corn. Field Crop. Res. 2012, 128, 48–54. [Google Scholar] [CrossRef]
- Antignus, Y. Comparative study of two Maize dwarf mosaic virus strains infecting corn and johnsongrass in Israel. Plant Dis. 1987, 71, 687–691. [Google Scholar] [CrossRef]
- Tóbiás, I.; Palkovics, L. An unusual feature at the N-terminal end of the coat protein of Maize dwarf mosaic virus isolated in Hungary. J. Phytopathol. 2004, 152, 445–457. [Google Scholar] [CrossRef]
- Robaglia, C.; Bruening, G.; Haseloff, J.; Gerlach, W.L. Evolution and replication of Tobacco ringspot virus satellite RNA mutants. EMBO J. 1993, 12, 2969–2976. [Google Scholar] [PubMed]
- Jones, F.E.; Hill, J.H.; Durand, D.P. Detection and differentiation of Maize dwarf mosaic virus, strains A and B, by use of different class immunoglobulins in a double-antibody sandwich enzyme-linked immunosorbent assay. Phytopathology 1988, 78, 1118–1124. [Google Scholar] [CrossRef]
- Sum, I.; Németh, M.; Pacsa, A.S. Detection of Maize dwarf mosaic virus with Enzyme-Linked Immunosorbent Assay (ELISA). J. Phytopathol. 1979, 95, 274–279. [Google Scholar] [CrossRef]
- Lenardon, S.L.; Giolitti, F.J. Maize Dwarf Mosaic Potyvirus on Maize in Argentina; IFFIVE, INTA: Cordoba, Argentina, 2006. [Google Scholar]
- Chen, J.; Chen, J.; Adams, M.J. A universal PCR primer to detect members of the Potyviridae and its use to examine the taxonomic status of several members of the family. Arch. Virol. 2001, 146, 757–766. [Google Scholar] [CrossRef] [PubMed]
- Tang, Y.B.; Xing, D.; Zhu, D.B.; Zhou, X.M. High sensitive method detection of plant RNA viruses by Electrochemiluminescence reverse transcription PCR. Prog. Biomed. Opt. Imaging- Proc. SPIE 2007, 6534, 1–8. [Google Scholar] [CrossRef]
- Hill, E.K.; Hill, J.H.; Durand, D.P. Production of monoclonal antibodies to viruses in the potyvirus group: Use in radioimmunoassay. J. Gen. Virol. 1984, 65, 525–532. [Google Scholar] [CrossRef] [PubMed]
- Krawczyk, K.; Uszczyńska-Ratajczak, B.; Majewska, A.; Borodynko-Filas, N. DNA microarray-based detection and identification of bacterial and viral pathogens of maize. J. Plant Dis. Prot. 2017, 124, 577–583. [Google Scholar] [CrossRef]
- Van Slogteren, E.; Van Slogteren, D.H. Serological identification of plant viruses and serological diagnosis of virus diseases of plants. Annu. Rev. Microbiol. 1957, 11, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Siti-Rohana, B.O.; Ahmad, I.B.; Nasuruddin, B.A. Application of protein-A indirect ELISA (PAI-ELISA) for the detection of anti-Smith antibodies in systemic lupus erythematosus patients. World J. Microbiol. Biotechnol. 1995, 11, 206–208. [Google Scholar] [CrossRef] [PubMed]
- Rybicki, E.P.; Von Wechmar, M.B. The serology of the Bromoviruses I. Serological interrelationships of the Bromoviruses. Virology 1981, 109, 391–402. [Google Scholar] [CrossRef]
- Diaco, R.; Lister, R.M.; Hill, J.H.; Durand, D.P. Demonstration of serological relationships among isolates of barley yellow dwarf virus by using polyclonal and monoclonal antibodies. J. Gen. Virol. 1986, 67, 353–362. [Google Scholar] [CrossRef]
- Koenig, R. ELISA in the study of homologous and heterologous reactions of plant viruses. J. Gen. Virol. 1978, 40, 309–318. [Google Scholar] [CrossRef]
- Van Regenmortel, M.H.V. Serology and immunochemistry of plant viruses. Immunol. Commun. 1982, 11, 509–525. [Google Scholar] [CrossRef]
- Ferro, D.N.; Mackenzie, J.D.; Margolies, D.C. Effect of mineral oil and a systemic insecticide on field spread of aphid-borne maize dwarf mosaic virus in sweet corn. J. Econ. Entomol. 1980, 73, 730–735. [Google Scholar] [CrossRef]
- Ismail, B.S.; Ingon, D.; Omar, O. Effects of metolachlor on activities of enzymes in a Malaysian soil. J. Environ. Sci. Health Part B 1996, 31, 1267–1278. [Google Scholar] [CrossRef]
- Toler, R.W. Maize dwarf mosaic, the most important virus disease of sorghum. Plant Dis. 1985, 69, 1011–1015. [Google Scholar] [CrossRef]
- Gordon, D.T.; Bradfute, O.E.; Gingery, R.E.; Knoke, J.K.; Nault, L.R.; Loden, H.D.; Wilkinson, D. Maize virus disease complexes in the United States: Real and potential disease problems. In 33rd Annual Corn and Sorghum Research Conference; American Seed Trade Association: Washington, DC, USA, 1978; pp. 102–133. [Google Scholar]
- Knoke, J.K.; Louie, R.; Madden, L.V.; Gordon, D.T. Spread of Maize dwarf mosaic virus from johnsongrass to corn. Plant Dis. 1983, 67, 367–370. [Google Scholar] [CrossRef]
- Yossen, V.; Nome, S.F.; Latorre, B.A.; Conci, L.R. Occurrence of Maize dwarf mosaic virus (MDMV) in Johnson grass (Sorghum halepense (L.) Pers.) in Chile. Cienc. Investig. Agrar. 1988, 15, 39–45. [Google Scholar]
- Angeles Achon, M.; Sobrepere, M. Incidence of potyviruses in commercial maize fields and their seasonal cycles in Spain. Zeitschrift für Pflanzenkrankheiten und Pflanzenschutz 2001, 108, 399–406. [Google Scholar]
- Stewart, L.R.; Teplier, R.; Todd, J.C.; Jones, M.W.; Cassone, B.J.; Wijeratne, S.; Wijeratne, A.; Redinbaugh, M.G. Viruses in Maize and Johnsongrass in Southern Ohio. Phytopathology 2014, 104, 1360–1369. [Google Scholar] [CrossRef] [PubMed]
- Hruska, A.J.; Gladstone, S.M.; Obando, R. Epidemic roller coaster: Maize stunt disease in Nicaragua. Am. Entomol. 1996, 42, 248–252. [Google Scholar] [CrossRef]
- Shepherd, D.N.; Martin, D.P.; Van der Walt, E.; Dent, K.; Varsani, A.; Rybicki, E.P. Maize streak virus: An old and complex ‘emerging’ pathogen. Mol. Plant Pathol. 2010, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Leustek, T. Repression of cystathionine γ-synthase in Arabidopsis thaliana produces partial methionine auxotrophy and developmental abnormalities. Plant Sci. 2000, 151, 9–18. [Google Scholar] [CrossRef]
- Hacham, Y.; Avraham, T.; Amir, R. The N-terminal region of Arabidopsis cystathionine γ-synthase plays an important regulatory role in methionine metabolism. Plant Physiol. 2002, 128, 454–462. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Paldi, K.; Racz, I.; Szigeti, Z.; Rudnoy, S.Z. The promising role of exogenous s-methylmethionine in agriculture, in the case of maize cultivation. Appl. Ecol. Environ. Res. 2014, 12, 777–785. [Google Scholar] [CrossRef]
- Ranocha, P.; McNeil, S.D.; Ziemak, M.J.; Li, C.; Tarczynski, M.C.; Hanson, A.D. The S-methylmethionine cycle in angiosperms: Ubiquity, antiquity and activity. Plant J. 2001, 25, 575–584. [Google Scholar] [CrossRef] [PubMed]
- Ko, S.; Eliot, A.C.; Kirsch, J.F. S-methylmethionine is both a substrate and an inactivator of 1-aminocyclopropane-1-carboxylate synthase. Arch. Biochem. Biophys. 2004, 421, 85–90. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Rudnóy, S.; Almási, A.; Szigeti, Z.; Rácz, I. The beneficial effects of S-methyl-methionine in maize in the case of Maize dwarf mosaic virus infection. Acta Biol. Szeged. 2011, 55, 109–112. [Google Scholar]
- Kocsis, M.G.; Nolte, K.D.; Rhodes, D.; Shen, T.L.; Gage, D.A.; Hanson, A.D. Dimethylsulfoniopropionate Biosynthesis inSpartina alterniflora 1: Evidence That S-Methylmethionine and Dimethylsulfoniopropylamine Are Intermediates. Plant Physiol. 1998, 117, 273–281. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Almasi, A.; Racz, I.; Szigeti, Z.; Solti, Á.; Olah, C.; Rudnoy, S. S-methylmethionine contributes to enhanced defense against Maize dwarf mosaic virus infection in maize. Braz. J. Bot. 2015, 38, 771–782. [Google Scholar] [CrossRef]
- Asada, K. Radical production and scavenging in the chloroplasts. Photosynth. Environ. 1996, 123–150. [Google Scholar] [CrossRef]
- Kingston-Smith, A.H.; Foyer, C.H. Bundle sheath proteins are more sensitive to oxidative damage than those of the mesophyll in maize leaves exposed to paraquat or low temperatures. J. Exp. Bot. 2000, 51, 123–130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fryer, M.J.; Andrews, J.R.; Oxborough, K.; Blowers, D.A.; Baker, N.R. Relationship between CO2 assimilation, photosynthetic electron transport, and active O2 metabolism in leaves of maize in the field during periods of low temperature. Plant Physiol. 1998, 116, 571–580. [Google Scholar] [CrossRef] [PubMed]
- Ludmerszki, E.; Chounramany, S.; Oláh, C.; Kátay, G.; Rácz, I.; Almási, A.; Solti, Á.; Bélai, I.; Rudnóy, S. Protective role of S-methylmethionine-salicylate in maize plants infected with Maize dwarf mosaic virus. Eur. J. Plant Pathol. 2017, 149, 145–156. [Google Scholar] [CrossRef]
- Mayhew, D.E.; Ford, R.E. Detection of ribonuclease-resistant RNA in chloroplasts of corn leaf tissue infected with Maize dwarf mosaic virus. Virology 1974, 57, 503–509. [Google Scholar] [CrossRef]
- Wei, T.; Huang, T.S.; McNeil, J.; Laliberté, J.F.; Hong, J.; Nelson, R.S.; Wang, A. Sequential recruitment of the endoplasmic reticulum and chloroplasts for plant potyvirus replication. J. Virol. 2010, 84, 799–809. [Google Scholar] [CrossRef] [PubMed]
- Beddard, G.S.; Porter, G. Concentration quenching in chlorophyll. Nature 1976, 260, 366–367. [Google Scholar] [CrossRef]
- Scott, G.E.; Findley, W.R. Breeding for resistance to maize virus diseases: A challenge. Maize Virus Dis. Newsl. 1984, 1984, 12–14. [Google Scholar]
- Martin, I.F.; McCarthy, T.E.; Persely, D.M.; Greber, R.S. Breeding sweet corn for resistance to Sugarcane mosaic virus—Johnsongrass strain in Australia. Maize Virus Dis. Newsl. 1984, 1, 15–16. [Google Scholar]
- Kovács, G.; Milinkó, I.; Gyulavári, O.; Farády, L. Resistance of F1, F2 and BC1generations to Maize dwarf mosaic virus in maize. Arch. Phytopathol. Plant Prot. 1990, 26, 363–368. [Google Scholar] [CrossRef]
- Chahal, G.S.; Gosal, S.S. Principles and Procedures of Plant Breeding: Biotechnological and Conventional Approaches; CRC Press: Boca Raton, FL, USA, 2002; ISBN 084931321X. [Google Scholar]
- Lei, J.D.; Agrios, G.N. Mechanisms of resistance in corn to Maize dwarf mosaic virus. Phytopathology 1986, 76, 1034–1040. [Google Scholar] [CrossRef]
- Louie, R. Effects of genotype and inoculation protocols on resistance evaluation of maize to Maize dwarf mosaic virus strains. Phytopathology 1986, 76, 769–773. [Google Scholar] [CrossRef]
- Findley, W.R.; Dollinger, E.J.; Louie, R.; Knoke, J.K. Locating Genes for Maize Dwarf Mosaic Resistance by Means of Chromosomal Translocations in Corn (Zea mays L.) 1. Crop Sci. 1973, 13, 608–611. [Google Scholar] [CrossRef]
- Scott, G.E.; Rosenkranz, E. Effectiveness of resistance to Maize dwarf mosaic and Maize chlorotic dwarf viruses in maize. Phytopathology 1981, 71, 937–941. [Google Scholar] [CrossRef]
- Scott, G.E. Linkage between Maize dwarf mosaic virus resistance and endosperm color in maize. Crop Sci. 1989, 29, 1478–1480. [Google Scholar] [CrossRef]
- McMullen, M.D.; Louie, R. Identification of a gene for resistance to Wheat streak mosaic virus in maize. Phytopathology 1991, 81, 624–627. [Google Scholar] [CrossRef]
- Ng, C.H.; Wickneswari, R.; Salmijah, S.; Teng, Y.T.; Ismail, B.S. Gene polymorphisms in glyphosate-resistant and -susceptible biotypes of Eleusine indica from Malaysia. Weed Res. 2003, 43, 108–115. [Google Scholar] [CrossRef]
- McMullen, M.D.; Louie, R. The linkage of molecular markers to a gene controlling the symptom response in maize to Maize dwarf mosaic virus. Mol. Plant-Microbe Interact. 1989, 2, 309–311. [Google Scholar] [CrossRef]
- Jones, M.W.; Redinbaugh, M.G.; Louie, R. The Mdm1 locus and maize resistance to Maize dwarf mosaic virus. Plant Dis. 2007, 91, 185–190. [Google Scholar] [CrossRef]
- Louie, R.; Findley, W.R.; Knoke, J.K.; McMullen, M.D. Genetics basis of resistance in maize to five Maize dwarf mosaic virus strains. Crop Sci. 1991, 31, 14–18. [Google Scholar] [CrossRef]
- Zambrano, J.L.; Jones, M.W.; Brenner, E.; Francis, D.M.; Tomas, A.; Redinbaugh, M.G. Genetic analysis of resistance to six virus diseases in a multiple virus-resistant maize inbred line. Theor. Appl. Genet. 2014, 127, 867–880. [Google Scholar] [CrossRef] [PubMed]
- Pataky, J.K.; Murphy, J.F.; D’arcy, C.J. Resistance to Maize dwarf mosaic virus, severity of symptoms, titer of virus, and yield of sweet corn. Plant Dis. 1990, 74, 359–364. [Google Scholar] [CrossRef]
- Scott, G.E.; Rosenkranz, E. Variable Reaction within Corn Inbreds to Maize dwarf mosaic virus not Genetically Controlled1. Crop Sci. 1987, 27, 78–79. [Google Scholar] [CrossRef]
- Jones, R.K.; Tolin, S.A. Concentration of Maize dwarf mosaic virus in susceptible and resistant corn hybrids. Phytopathology 1972, 62, 640–644. [Google Scholar] [CrossRef]
- Kuntze, L.; Fuchs, E.; Grüntzig, M.; Schulz, B.; Klein, D.; Melchinger, A.E. Resistance of early-maturing European maize germplasm to Sugarcane mosaic virus (SCMV) and Maize dwarf mosaic virus (MDMV). Plant Breed. 1997, 116, 499–501. [Google Scholar] [CrossRef]
- Law, M.D.; Moyer, J.W.; Payne, G.A. The effect of host resistance on pathogenesis of Maize dwarf mosaic virus. Phytopathology 1989, 79, 757–761. [Google Scholar] [CrossRef]
- Azad, M.; Kalam, A.; Amin, L.; Sidik, N.M. Gene technology for Papaya ringspot virus disease management. Sci. World J. 2014, 2014, 768038. [Google Scholar] [CrossRef] [PubMed]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for antiviral resistance in transgenic plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar] [CrossRef] [PubMed]
- Louie, R. Vascular puncture of maize kernels for the mechanical transmission of Maize white line mosaic virus and other viruses of maize. Phytopathology 1995, 85, 139–143. [Google Scholar] [CrossRef]
- Atabekov, J.G.; Dorokhov, Y.L. Plant virus-specific transport function and resistance of plants to viruses. Adv. Virus Res. 1984, 29, 313–364. [Google Scholar] [CrossRef] [PubMed]
- Bennetzen, J.L.; Hake, S.C. Handbook of Maize: Its Biology; Springer: New York, NY, USA, 2009; pp. 121–126. ISBN 978-0-387-79417-4e. [Google Scholar]
- Cassone, B.J.; Chen, Z.; Chiera, J.; Stewart, L.R.; Redinbaugh, M.G. Responses of highly resistant and susceptible maize to vascular puncture inoculation with Maize dwarf mosaic virus. Physiol. Mol. Plant Pathol. 2014, 86, 19–27. [Google Scholar] [CrossRef]
- Xing, Y.; Ingvardsen, C.; Salomon, R.; Lübberstedt, T. Analysis of Sugarcane mosaic virus resistance in maize in an isogenic dihybrid crossing scheme and implications for breeding potyvirus-resistant maize hybrids. Genome 2006, 49, 1274–1282. [Google Scholar] [CrossRef] [PubMed]
- Johal, G.S.; Briggs, S.P. Reductase activity encoded by the HM1 disease resistance gene in maize. Science 1992, 258, 985–987. [Google Scholar] [CrossRef] [PubMed]
- Martin, G.B.; Brommonschenkel, S.H.; Chunwongse, J.; Frary, A.; Ganal, M.W.; Spivey, R.; Tanksley, S.D. Map-based cloning of a protein kinase gene conferring disease resistance in tomato. Science 1993, 262, 1432–1436. [Google Scholar] [CrossRef] [PubMed]
- Simcox, K.D.; McMullen, M.D.; Louie, R. Co-segregation of the Maize dwarf mosaic virus resistance gene, Mdm1, with the nucleolus organizer region in maize. Theor. Appl. Genet. 1995, 90, 341–346. [Google Scholar] [CrossRef] [PubMed]
- Xi, Z.Y.; Zhang, S.H.; Li, X.H.; Xie, C.X.; Li, M.S.; Hao, Z.F.; Zhang, D.G.; Liang, Y.H.; Bai, L.; Zhang, S.H. Identification and mapping of a novel Sugarcane mosaic virus resistance gene in maize. Acta Agron. Sin. 2008, 34, 1494–1499. [Google Scholar] [CrossRef]
- Liu, X.; Tan, Z.; Li, W.; Zhang, H.; He, D. Cloning and transformation of SCMV CP gene and regeneration of transgenic maize plants showing resistance to SCMV strain MDB. Afr. J. Biotechnol. 2009, 8, 3747–3753. [Google Scholar]
- Murry, L.E.; Elliott, L.G.; Capitant, S.A.; West, J.A.; Hanson, K.K.; Scarafia, L.; Johnston, S.; DeLuca-Flaherty, C.; Nichols, S.; Cunanan, D.; et al. Transgenic corn plants expressing MDMV strain B coat protein are resistant to mixed infections of Maize dwarf mosaic virus and Maize chlorotic mottle virus. Nat. Biotechnol. 1993, 11, 1559–1564. [Google Scholar] [CrossRef]
- Zhang, Z.Y.; Fu, F.L.; Gou, L.; Wang, H.G.; Li, W.C. RNA interference-based transgenic maize resistant to Maize dwarf mosaic virus. J. Plant Biol. 2010, 53, 297–305. [Google Scholar] [CrossRef]
- Behboodian, B.; Ali, Z.M.; Ismail, I.; Zainal, Z. Postharvest analysis of lowland transgenic tomato fruits harboring hpRNAi-ACO1 construct. Sci. World J. 2012, 2012, 439870. [Google Scholar] [CrossRef] [PubMed]
- Fahim, M.; Ayala-Navarrete, L.; Millar, A.A.; Larkin, P.J. Hairpin RNA derived from viral NIa gene confers immunity to Wheat streak mosaic virus infection in transgenic wheat plants. Plant Biotechnol. J. 2010, 8, 821–834. [Google Scholar] [CrossRef] [PubMed]
- Praveen, S.; Ramesh, S.V.; Mishra, A.K.; Koundal, V.; Palukaitis, P. Silencing potential of viral derived RNAi constructs in Tomato leaf curl virus-AC4 gene suppression in tomato. Transgenic Res. 2010, 19, 45–55. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.Y.; Yang, L.; Zhou, S.F.; Wang, H.G.; Li, W.C.; Fu, F.L. Improvement of resistance to maize dwarf mosaic virus mediated by transgenic RNA interference. J. Biotechnol. 2011, 153, 181–187. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.K.; Lohuis, D.; Goldbach, R.; Prins, M. High frequency induction of RNA-mediated resistance against Cucumber mosaic virus using inverted repeat constructs. Mol. Breed. 2004, 14, 215–226. [Google Scholar] [CrossRef]
- Hily, J.M.; Ravelonandro, M.; Damsteegt, V.; Bassett, C.; Petri, C.; Liu, Z.; Scorza, R. Plum pox virus coat protein gene Intron-hairpin-RNA (ihpRNA) constructs provide resistance to Plum pox virus in Nicotiana benthamiana and Prunus domestica. J. Am. Soc. Hortic. Sci. 2007, 132, 850–858. [Google Scholar]
- Zhang, Z.Y.; Wang, Y.G.; Shen, X.J.; Li, L.; Zhou, S.F.; Li, W.C.; Fu, F.L. RNA interference-mediated resistance to Maize dwarf mosaic virus. Plant Cell Tissue Organ Cult. 2013, 113, 571–578. [Google Scholar] [CrossRef]
- Redinbaugh, M.G.; Jones, M.W.; Gingery, R.E. The genetics of virus resistance in maize (Zea mays L.). Maydica 2004, 49, 183–190. [Google Scholar]
Figure 1.
Mosaic symptoms on the lower leaves and yellow streaks along veins in corn plant as it approaches maturity. Also the fitch, Rhopalosiphum maidis, nymphs on the maize leaves.
Figure 1.
Mosaic symptoms on the lower leaves and yellow streaks along veins in corn plant as it approaches maturity. Also the fitch, Rhopalosiphum maidis, nymphs on the maize leaves.

Figure 2.
Interaction between MDMV and the vector appendage.

Figure 3.
Outline of regulatory and protective compounds anabolism stimulated by S-Methylmethonine.

Figure 4.
Effects of S-methylmethionine application on GF14-6 and S-adenosylmethionine synthase (SAMS) expression level in MDMV infected maize plants.
Figure 4.
Effects of S-methylmethionine application on GF14-6 and S-adenosylmethionine synthase (SAMS) expression level in MDMV infected maize plants.

Figure 5.
Role of S-Methylmethionine salicylate in the prevention of reactive oxygen species (ROS) development in corn plants.
Figure 5.
Role of S-Methylmethionine salicylate in the prevention of reactive oxygen species (ROS) development in corn plants.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Available complete sequences of maize dwarf mosaic virus (MDMV) isolates.
| MDMV Isolates | Geographical Origin | Genbank Accession | References |
|---|---|---|---|
| MDMV | Golestan | JQ280313 | [38] |
| MDMV-Bg | Bulgaria | NC003377 | [27] |
| MDMV-Sp | Spain | AJ416645 | [39] |
| MDMV-OH1 | Ohia | JQ403608 | [40] |
| MDMV-OH2 | Ohia | JQ403609 | [40] |
| MDMV-It | Italy | JX185302 | [41] |
Table 2.
Methods for Maize dwarf mosaic virus (MDMV) detection.
| Method | Platform | Effect on MDMV Variation | Reliability | Efficiency | References |
|---|---|---|---|---|---|
| Indirect Enzyme-Linked Immunosorbent Assay (ELISA) |
| MDMV-A and MDMV-B | Allow detection of MDMV-A and MDMV-B bound IgG among tested leaves sap infected with several different strains of MDMV and Sugarcane mosaic virus (SCMV). | MDMV-A-specific ELISA = antigen detected in sap at a dilution of 1:60 with optimal sample pH (7.5–8.5). MDMV-B-specific ELISA = detect antigen in sap at dilution end point of 1:2560 | [91] |
| Indirect Enzyme-Linked Immunosorbent Assay (ELISA) |
| M-C and M-D | M-C antigen reacts strongly with M-A, M-D, Sorghum red stripe virus (SRV) antisera. M-D antigen reacts strongly with M-D, M-C antisera. | M-C particles react with M-A and SRV antisera in the range of 1:128 to 1:512 dilution end points, with M-D antisera at dilution end point of 1:4. M-D antigen reacted with homologous antisera, M-D up to a dilution of 1:512, with M-C antisera at 1:16 dilution. | [88] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV-A | Visual evaluation at absorbance of 405 nm gives reliable information on MDMV-A presence in leaves extracts tested. | Visible yellow colour formed in wells with antigen diluted up to 10−4. Sensitivity is 100 times better than conventional infectivity test. | [92] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV-A, MVMV-J, MDMV-L, MDMV-SP, MDMV-YU | Among all reference strains, only MDMV strains react positively in DAS-ELISA (recorded OD 405 nm values at least twice the healthy sap OD 405 nm values) with anti-MDMV IgG. | IgG antibody dilution to 1 μg/mL able to detect MDMV antigen dilution to 1/100. | [30] |
| Double-Antibody Sandwich Enzyme-Linked Immunosorbent Assay (DAS ELISA) |
| MDMV | DAS-ELISA absorbance values of infected maize leaf samples from field grown plants and maize leaf sample with MDMV-Arg isolate were highly significant while the values from healthy control and buffer were very low. | MDMV-Arg specific polyclonal IgG strongly reactive up to 1:2000 dilution with MDMV antigen from infected field, Chile. | [19] |
| Capture Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | Single band of expected size (1104 bp) obtained. | Detect MDMV samples from Chile which closely related to MDMV-Arg (Argentina strain) efficiently. | [19] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV-SP, MDMV-Bu, MDMVJIL | Single product of expected size (327 nts) obtained for all tested MDMV isolates. | Efficient for dealing with well-characterized strains, field collected isolates. | [30] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | cDNA fragments of expected size were amplified from the 3′ terminus of RNA genomes of 21 different viruses under family potyviridae including MDMV. | Universal primer designed based on an alignment of the amino acid sequences around the conserved GNNSGQP motif in nuclear inclusion body b (NIb) gene of family potyviridae members. Hence, it is proved useful for detection of potyviridae members. | [94] |
| Reverse Transcription-Polymerase Chain Reaction (RT-PCR) |
| MDMV | Supposed to yield a PCR product of expected size (336 bp) | MDMV can be distinguishable from Sugarcane mosaic virus, SCMV and Johnsongrass mosaic virus, JGMV through RT-PCR | [57] |
| Combined Reverse Transcription Polymerase Chain Reaction (RT-PCR) with Electrochemiluminescence method |
| MDMV | PCR yielded a product with a single band of expected size (643 bp) for all 4 tested viruses including MDMV. | This method has higher sensitivity and lower cost than others. It can effectively detect the MDMV with simplicity and stability. | [95] |
| Competitive Radioimmunoassay (RIA) |
| MDMV-A (referred here as MDMV-AP) and MDMV-B | Feasible alternative to the use of polyclonal antisera in detecting homologous viruses (MDMV, Soybean mosaic, SMV, Lettuce mosaic virus, LMV). | Antigen (purified virus) detected at dilution of 10–50 ng/mL. | [96] |
| Dot Blot Immunoassay |
| MDMV | MDMV symptomatic field grown plants had strong reaction with the polyclonal antiserum against MDMV-Arg isolate while healthy plants were negative. | MDMV samples reacted with MDMV-Arg polyclonal antiserum of dilution up to 1:5000 | [19] |
| Sodium dodecyl sulfate (SDS) immunodiffusion test |
| MDMV-C and MDMV-D | M-C antiserum reacted with both M-C and M-D antigens forming a spur, which indicate partial serological relatedness. M-D antiserum reacted with its homologous viral antigen, M-D. | No precipitin lines were obtained when antisera reacted against healthy crude sap | [88] |
| DNA Microarray (Maizepath)-based detection | Microarray with 60-mer oligonucleotide probes complementary to genomes of 5 maize pathogens including MDMV | MDMV | Obtained results indicate that the fluorescence signals from MDMV, other pathogens and control probes are well distinguished in all performed experiments. | Gives more than 180 K probes in total, thereby classified as high-density microarray that able to investigate thousands of genomic loci in a high-resolution manner. | [97] |
Table 3.
Summary on genes conferring resistance against maize dwarf mosaic virus (MDMV).
| Chromosome | Locus | Resistance Source | Screening Method | Level of Resistance | References |
|---|---|---|---|---|---|
| 6 (short arm) | Mdm1 | Pa405 Oh1V1 | Mechanical inoculation/Greenhouse Mechanical inoculation/Field | High resistance (Dominant gene) | [140,143] |
| 6 | Mdm1 co-localizes with Wsm1 | Wsm1 NIL (near isogenic lines) Oh28SS/RR/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 (Two Mdm1/Wsm1 allele) | Mechanical inoculation/Greenhouse/Field condition | High resistance | [80] |
| 6 | Mdm1 co-localizes with Wsm1 | Wsm1 × Oh28 F1 Oh28SS/RS/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 (One Mdm1/Wsm1 allele) | Mechanical inoculation/Greenhouse/Field condition | Intermediate resistance | [80] |
| 3 & 6 | Wsm2 combine with Mdm1/Wsm1 | Wsm1 Wsm2 NIL Oh28RR/RR/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers symptom incidence | [80,142] |
| 3 & 6 | Wsm2 combine with Mdm1/Wsm1 | Wsm1 Wsm2 × Oh28 F1 Oh28RS/RS/SS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers incidence and severity of disease | [80,142] |
| 6 & 10 | Wsm3 combine with Mdm1/Wsm1 | Wsm1 Wsm3 × Oh28 F1 Oh28SS/RS/RS Left: Wsm2 alleles Middle: Mdm1/Wsm1 alleles Right: Wsm3 | Mechanical inoculation/Greenhouse/Field condition | Lowers disease incidence and severity | [80,141] |
| 3 & 6 | Scmv2 combined with Scmv1 | F7 RR/RR Left: Scmv2 alleles Right: Scmv1 alleles R: Susceptible parent F7 R: Resistant parent FAP1360A | Mechanical inoculation by rubbing infected leaves | Complete resistance | [155] |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Kannan, M.; Ismail, I.; Bunawan, H. Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses 2018, 10, 492. https://0-doi-org.brum.beds.ac.uk/10.3390/v10090492
AMA Style
Kannan M, Ismail I, Bunawan H. Maize Dwarf Mosaic Virus: From Genome to Disease Management. Viruses. 2018; 10(9):492. https://0-doi-org.brum.beds.ac.uk/10.3390/v10090492
Chicago/Turabian StyleKannan, Maathavi, Ismanizan Ismail, and Hamidun Bunawan. 2018. "Maize Dwarf Mosaic Virus: From Genome to Disease Management" Viruses 10, no. 9: 492. https://0-doi-org.brum.beds.ac.uk/10.3390/v10090492
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.