Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica)


 ,
,  , ,
, , {kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
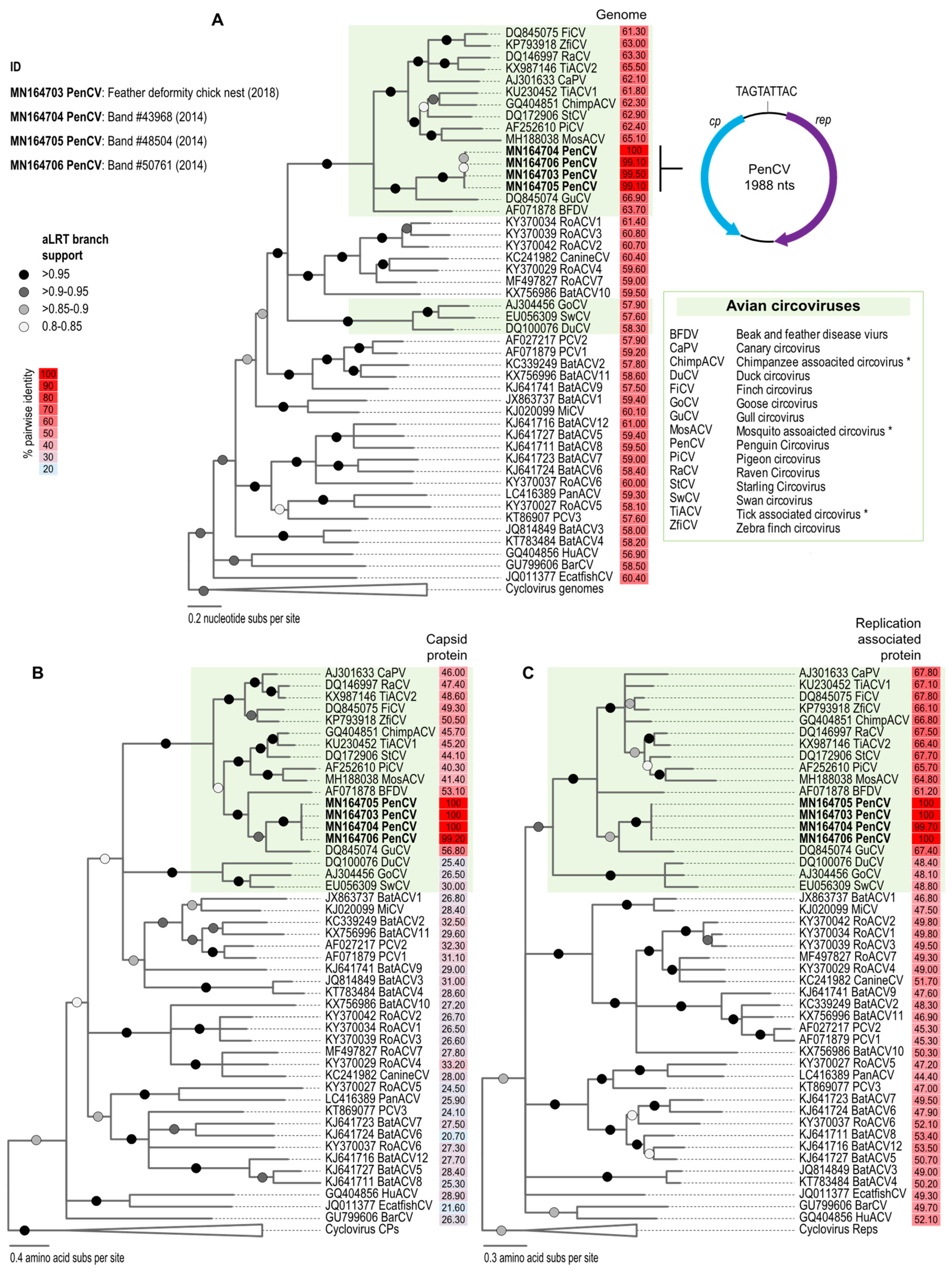
3. Results and Discussion
4. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kane, O.J.; Smith, J.R.; Boersma, P.D.; Parsons, N.J.; Strauss, V.; Garcia-Borboroglu, P.; Villanueva, C. Feather-Loss Disorder in African and Magellanic Penguins. Waterbirds 2010, 33, 415–421. [Google Scholar] [CrossRef]
- Barbosa, A.; Colominas-Ciuró, R.; Coria, N.; Centurión, M.; Sandler, R.; Negri, A.; Santos, M. First record of feather-loss disorder in Antarctic penguins. Antarct. Sci. 2014, 27, 69–70. [Google Scholar] [CrossRef]
- Lyver, P.O.; Barron, M.; Barton, K.J.; Ainley, D.G.; Pollard, A.; Gordon, S.; McNeill, S.; Ballard, G.; Wilson, P.R. Trends in the breeding population of Adelie penguins in the Ross Sea, 1981–2012: A coincidence of climate and resource extraction effects. PLoS ONE 2014, 9, e91188. [Google Scholar] [CrossRef] [PubMed]
- Lynch, H.J.; LaRue, M.A. First global census of the Adélie Penguin. Auk 2014, 131, 457–466. [Google Scholar] [CrossRef]
- Grimaldi, W.W.; Hall, R.J.; White, D.D.; Wang, J.; Massaro, M.; Tompkins, D.M. First report of a feather loss condition in Adelie penguins (Pygoscelis adeliae) on Ross Island, Antarctica, and a preliminary investigation of its cause. Emu 2016, 115, 185–189. [Google Scholar] [CrossRef]
- Smeele, Z.E.; Ainley, D.G.; Varsani, A. Viruses associated with Antarctic wildlife: From serology based detection to identification of genomes using high throughput sequencing. Virus Res. 2018, 243, 91–105. [Google Scholar] [CrossRef]
- Phalen, D.N. Papillomaviruses and Polyomaviruses. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Atkinson, C.T., Eds.; Blackwell Publishing: Melbourne, VIC, Australia, 2007. [Google Scholar]
- Paré, J.A.; Robert, N. Circovirus. In Infectious Diseases of Wild Birds; Thomas, N.J., Hunter, D.B., Atkinson, C.T., Eds.; Blackwell Publishing: Melbourne, VIC, Australia, 2007. [Google Scholar]
- Varsani, A.; Porzig, E.L.; Jennings, S.; Kraberger, S.; Farkas, K.; Julian, L.; Massaro, M.; Ballard, G.; Ainley, D.G. Identification of an avian polyomavirus associated with Adelie penguins (Pygoscelis adeliae). J. Gen. Virol. 2015, 96, 851–857. [Google Scholar] [CrossRef]
- Rosario, K.; Breitbart, M.; Harrach, B.; Segales, J.; Delwart, E.; Biagini, P.; Varsani, A. Revisiting the taxonomy of the family Circoviridae: Establishment of the genus Cyclovirus and removal of the genus Gyrovirus. Arch. Virol. 2017, 162, 1447–1463. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Steffens, W.L.; Latimer, K.S. Characterization of a New Virus from Cockatoos with Psittacine Beak and Feather Disease. Virology 1989, 171, 83–88. [Google Scholar] [CrossRef]
- Morozov, I.; Sirinarumitr, T.; Sorden, S.D.; Halbur, P.G.; Morgan, M.K.; Yoon, K.J.; Paul, P.S. Detection of a novel strain of porcine circovirus in pigs with postweaning multisystemic wasting syndrome. J. Clin. Microbiol. 1998, 36, 2535–2541. [Google Scholar]
- Hamel, A.L.; Lin, L.L.; Nayar, G.P.S. Nucleotide sequence of porcine circovirus associated with postweaning multisystemic wasting syndrome in pigs. J. Virol. 1998, 72, 5262–5267. [Google Scholar] [PubMed]
- Grau-Roma, L.; Fraile, L.; Segalés, J. Recent advances in the epidemiology, diagnosis and control of diseases caused by porcine circovirus type 2. Vet. J. 2011, 187, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Doneley, R.J. Acute beak and feather disease in juvenile African Grey parrots—An uncommon presentation of a common disease. Aust. Vet. J. 2003, 81, 206–207. [Google Scholar] [CrossRef] [PubMed]
- Ritchie, B.W.; Niagro, F.D.; Lukert, P.D.; Latimer, K.S.; Steffens III, W.L.; Pritchard, N. A Review of Psittacine Beak and Feather Disease: Characteristics of the PBFD Virus. J. Assoc. Avian Vet. 1989, 3, 143–149. [Google Scholar] [CrossRef]
- McOrist, S.; Black, D.G.; Pass, D.A.; Scott, P.C.; Marshall, J. Beak and feather dystrophy in wild sulphur-crested cockatoos (Cacatua galerita). J. Wildl. Dis 1984, 20, 120–124. [Google Scholar] [CrossRef]
- Huff, D.G.; Schmidt, R.E.; Fudge, A.M. Psittacine beak and feather syndrome in a blue-fronted Amazon (Amazona aestiva). AAV Today 1988, 2, 84–86. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Latimer, K.S.; Lukert, P.D.; Steffens, W.; Rakich, P.M.; Pritchard, N. Ultrastructural, protein composition, and antigenic comparison of psittacine beak and feather disease virus purified from four genera of psittacine birds. J. Wildl. Dis. 1990, 26, 196–203. [Google Scholar] [CrossRef]
- Pass, D.A.; Perry, R.A. The pathology of psittacine beak and feather disease. Aust. Vet. J. 1984, 61, 69–74. [Google Scholar] [CrossRef]
- Raidal, S.R. Viral skin diseases of birds. Semin. Avian Exot. Pet Med. 1995, 4, 72–82. [Google Scholar] [CrossRef]
- Ritchie, B.W.; Niagro, F.D.; Latimer, K.S.; Steffens, W.L.; Pesti, D.; Ancona, J.; Lukert, P.D. Routes and Prevalence of Shedding of Psittacine Beak and Feather Disease Virus. Am. J. Vet. Res. 1991, 52, 1804–1809. [Google Scholar]
- Rahaus, M.; Desloges, N.; Probst, S.; Loebbert, B.; Lantermann, W.; Wolff, M.H. Detection of beak and feather disease virus DNA in embryonated eggs of psittacine birds. Vet. Med. Czech. 2008, 53, 53–58. [Google Scholar] [CrossRef]
- Soike, D.; Kohler, B.; Albrecht, K. A circovirus-like infection in geese related to a runting syndrome. Avian Pathol. 1999, 28, 199–202. [Google Scholar] [CrossRef]
- Soike, D.; Albrecht, K.; Hattermann, K.; Schmitt, C.; Mankertz, A. Novel circovirus in mulard ducks with developmental and feathering disorders. Vet. Rec. 2004, 154, 792–793. [Google Scholar] [CrossRef] [PubMed]
- Todd, D.; Weston, J.; Ball, N.W.; Borghmans, B.J.; Smyth, J.A.; Gelmini, L.; Lavazza, A. Nucleotide sequence-based identification of a novel circovirus of canaries. Avian Pathol. 2001, 30, 321–325. [Google Scholar] [CrossRef] [PubMed]
- Shivaprasad, H.L.; Hill, D.; Todd, D.; Smyth, J.A. Circovirus infection in a Gouldian finch (Chloebia gouldiae). Avian Pathol. 2004, 33, 525–529. [Google Scholar] [CrossRef]
- Mysore, J.; Read, D.; Daft, B. Circovirus-like particles in finches. In Proceedings of the Annual Conference of the American Association of Veterinary Laboratory Diagnosis, Histopathology Section. Reno, NV, USA, 18–23 October 2007. [Google Scholar]
- Circella, E.; Legretto, M.; Pugliese, N.; Caroli, A.; Bozzo, G.; Accogli, G.; Lavazza, A.; Camarda, A. Psittacine beak and feather disease-like illness in Gouldian finches (Chloebia gouldiae). Avian Dis. 2014, 58, 482–487. [Google Scholar] [CrossRef]
- Raidal, S.R.; Riddoch, P.A. A feather disease in Senegal doves (Streptopelia senegalensis) morphologically similar to psittacine beak and feather disease. Avian Pathol. 1997, 26, 829–836. [Google Scholar] [CrossRef]
- Pass, D.A.; Plant, S.L.; Sexton, N. Natural infection of wild doves (Streptopelia senegalensis) with the virus of psittacine beak and feather disease. Aust. Vet. J. 1994, 71, 307–308. [Google Scholar] [CrossRef]
- Woods, L.W.; Latimer, K.S. Circovirus infection of nonpsittacine birds. J. Avian Med. Surg. 2000, 14, 154–163. [Google Scholar] [CrossRef]
- Fahsbender, E.; Burns, J.M.; Kim, S.; Kraberger, S.; Frankfurter, G.; Eilers, A.A.; Shero, M.R.; Beltran, R.; Kirkham, A.; McCorkell, R.; et al. Diverse and highly recombinant anelloviruses associated with Weddell seals in Antarctica. Virus Evol. 2017, 3. [Google Scholar] [CrossRef]
- Varsani, A.; Kraberger, S.; Jennings, S.; Porzig, E.L.; Julian, L.; Massaro, M.; Pollard, A.; Ballard, G.; Ainley, D.G. A novel papillomavirus in Adelie penguin (Pygoscelis adeliae) faeces sampled at the Cape Crozier colony, Antarctica. J. Gen. Virol. 2014, 95, 1352–1365. [Google Scholar] [CrossRef] [PubMed]
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Kearse, M.; Moir, R.; Wilson, A.; Stones-Havas, S.; Cheung, M.; Sturrock, S.; Buxton, S.; Cooper, A.; Markowitz, S.; Duran, C.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [PubMed]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef] [Green Version]
- Guindon, S.; Dufayard, J.F.; Lefort, V.; Anisimova, M.; Hordijk, W.; Gascuel, O. New algorithms and methods to estimate maximum-likelihood phylogenies: Assessing the performance of PhyML 3.0. Syst. Biol. 2010, 59, 307–321. [Google Scholar] [CrossRef] [Green Version]
- Posada, D. Selection of models of DNA evolution with jModelTest. Methods Mol. Biol. 2009, 537, 93–112. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef] [Green Version]
- Anisimova, M.; Gascuel, O. Approximate likelihood-ratio test for branches: A fast, accurate, and powerful alternative. Syst. Biol. 2006, 55, 539–552. [Google Scholar] [CrossRef]
- Stover, B.C.; Muller, K.F. TreeGraph 2: Combining and visualizing evidence from different phylogenetic analyses. BMC Bioinform. 2010, 11, 7. [Google Scholar] [CrossRef] [Green Version]
- Muhire, B.M.; Varsani, A.; Martin, D.P. SDT: A virus classification tool based on pairwise sequence alignment and identity calculation. PLoS ONE 2014, 9, e108277. [Google Scholar] [CrossRef] [PubMed]
- MacLachlan, N.J.; Dubovi, E.J. Circoviridae and Anelloviridae. In Fenner’s Veterinary Virology; MacLachlan, N.J., Dubovi, E.J., Eds.; Academic Press: Boston, MA, USA, 2017; pp. 259–268. [Google Scholar]
- Fogell, D.J.; Martin, R.O.; Bunbury, N.; Lawson, B.; Sells, J.; McKeand, A.M.; Tatayah, V.; Trung, C.T.; Groombridge, J.J. Trade and conservation implications of new beak and feather disease virus detection in native and introduced parrots. Conserv. Biol. 2018, 32, 1325–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ha, H.J.; Anderson, I.L.; Alley, M.R.; Springett, B.P.; Gartrell, B.D. The prevalence of beak and feather disease virus infection in wild populations of parrots and cockatoos in New Zealand. N. Z. Vet. J. 2007, 55, 235–238. [Google Scholar] [CrossRef] [PubMed]
- Jackson, B.; Lorenzo, A.; Theuerkauf, J.; Barnaud, A.; Duval, T.; Guichard, P.; Bloc, H.; Baouma, A.; Stainton, D.; Kraberge, S.; et al. Preliminary surveillance for beak and feather disease virus in wild parrots of New Caledonia: Implications of a reservoir species for Ouvea Parakeets. Emu 2014, 114, 283–289. [Google Scholar] [CrossRef]
- Massaro, M.; Ortiz-Catedral, L.; Julian, L.; Galbraith, J.A.; Kurenbach, B.; Kearvell, J.; Kemp, J.; van Hal, J.; Elkington, S.; Taylor, G.; et al. Molecular characterisation of beak and feather disease virus (BFDV) in New Zealand and its implications for managing an infectious disease. Arch. Virol. 2012, 157, 1651–1663. [Google Scholar] [CrossRef]
- Ortiz-Catedral, L.; McInnes, K.; Hauber, M.E.; Brunton, D.H. First report of beak and feather disease virus (BFDV) in wild Red-fronted Parakeets (Cyanoramphus novaezelandiae) in New Zealand. Emu 2009, 109, 244–247. [Google Scholar] [CrossRef]
- Dugger, K.M.; Ainley, D.G.; Lyver, P.O.; Barton, K.; Ballard, G. Survival differences and the effect of environmental instability on breeding dispersal in an Adelie penguin meta-population. Proc. Natl. Acad. Sci. USA 2010, 107, 12375–12380. [Google Scholar] [CrossRef] [Green Version]
- Dugger, K.M.; Ballard, G.; Ainley, D.G.; Lyver, P.O.B.; Schine, C. Adelie penguins coping with environmental change: Results from a natural experiment at the edge of their breeding range. Front. Ecol. Evol. 2014, 2. [Google Scholar] [CrossRef] [Green Version]
- Ainley, D. The Adélie Penguin: Bellwether of Climate Change; Columbia University Press: New York, NY, USA, 2002. [Google Scholar]


© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Morandini, V.; Dugger, K.M.; Ballard, G.; Elrod, M.; Schmidt, A.; Ruoppolo, V.; Lescroël, A.; Jongsomjit, D.; Massaro, M.; Pennycook, J.; et al. Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica). Viruses 2019, 11, 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121088
Morandini V, Dugger KM, Ballard G, Elrod M, Schmidt A, Ruoppolo V, Lescroël A, Jongsomjit D, Massaro M, Pennycook J, et al. Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica). Viruses. 2019; 11(12):1088. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121088
Chicago/Turabian StyleMorandini, Virginia, Katie M. Dugger, Grant Ballard, Megan Elrod, Annie Schmidt, Valeria Ruoppolo, Amélie Lescroël, Dennis Jongsomjit, Melanie Massaro, Jean Pennycook, and et al. 2019. "Identification of a Novel Adélie Penguin Circovirus at Cape Crozier (Ross Island, Antarctica)" Viruses 11, no. 12: 1088. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121088