Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2


Abstract
:1. Introduction
2. Materials and Methods
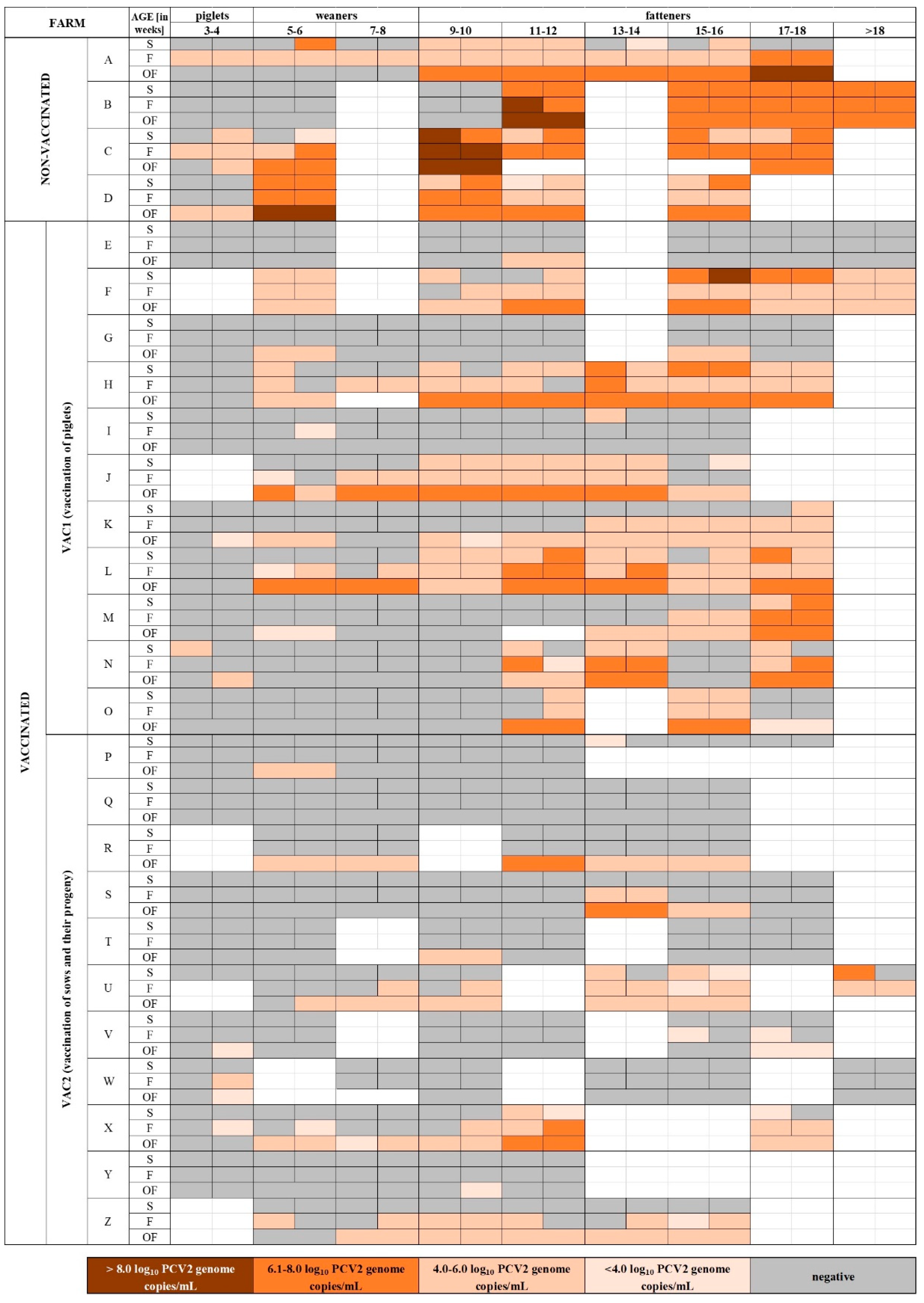
2.1. Study Farms
2.2. Sample Collection and Processing
2.3. Real-Time PCR
2.4. DNA Sequence Analysis
2.5. Statistical Analysis
3. Results
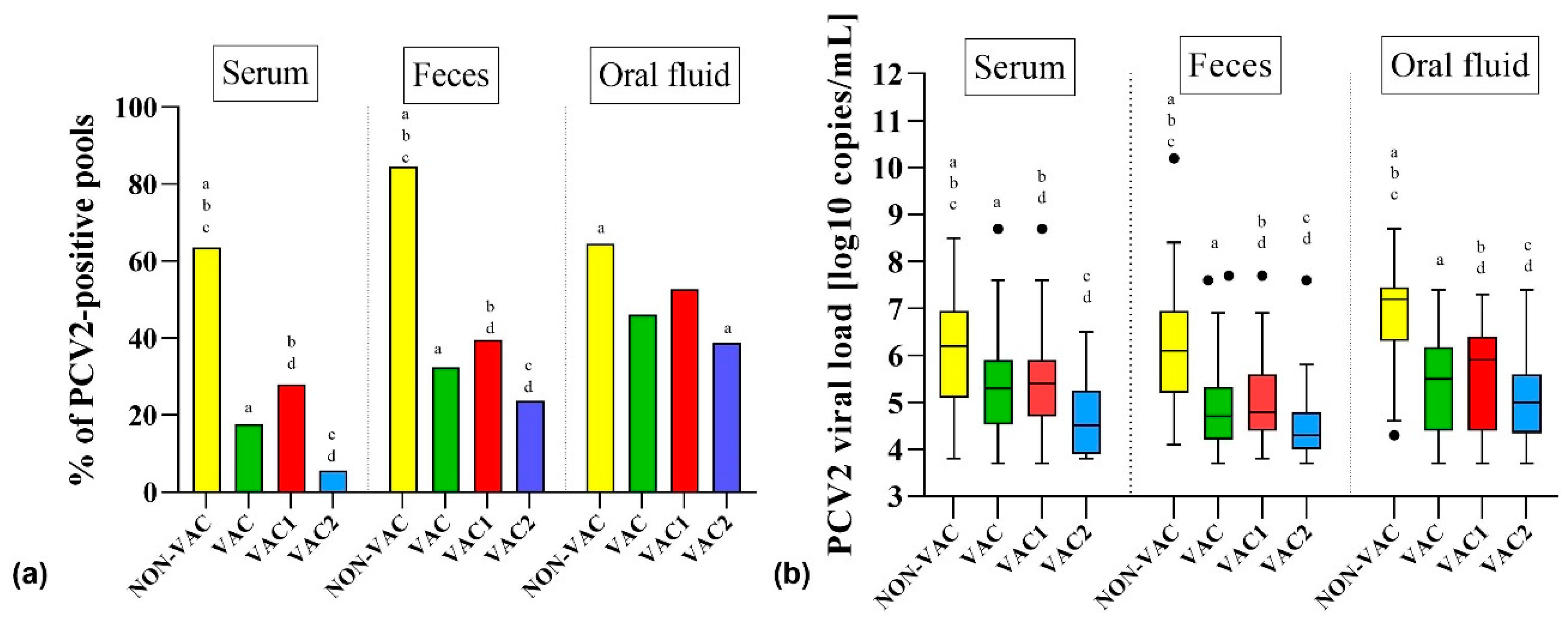
3.1. Detection of PCV2 in Samples from Pigs from Farms that Vaccinate (VAC) and Do Not Vaccinate (NON-VAC) against PCV2
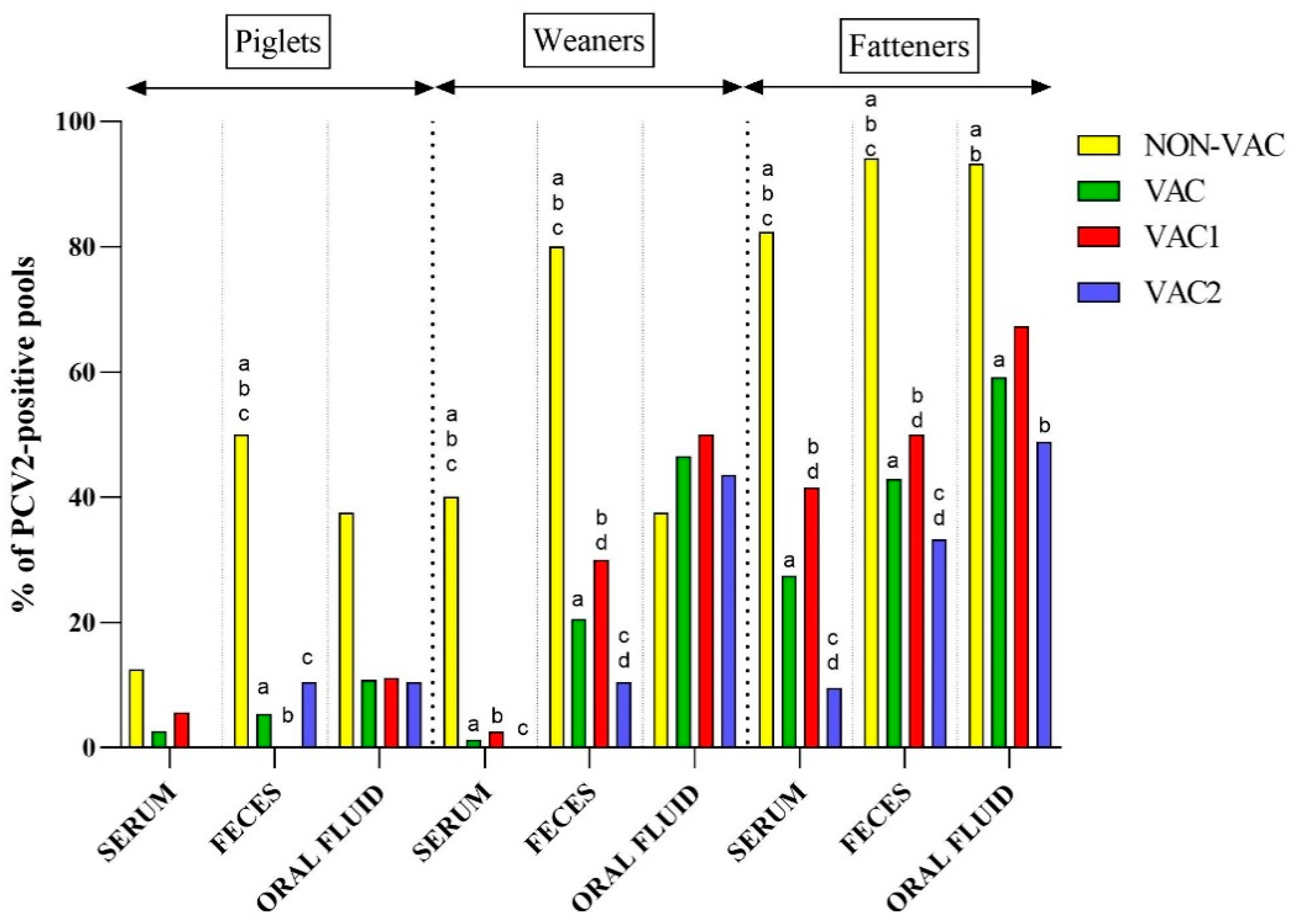
3.2. Detection of PCV2 in Samples from Pigs from Farms Using Two Different PCV2 Vaccination Protocols
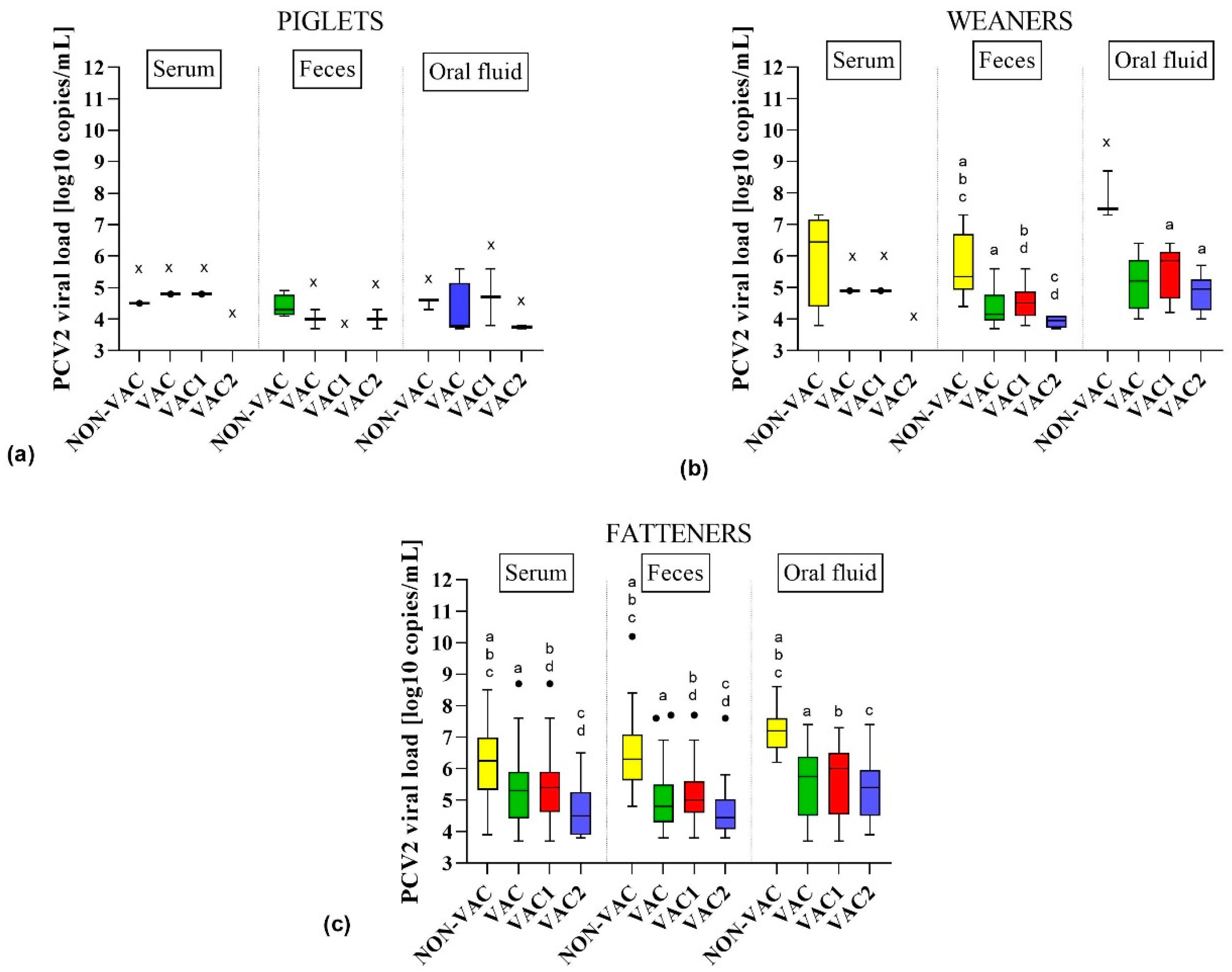
3.3. Detection of PCV2 in Samples from Different Age Groups of Pigs
3.4. Probability of PCV2 Detection in Different Types of Samples, Age Groups, and Vaccination Protocols
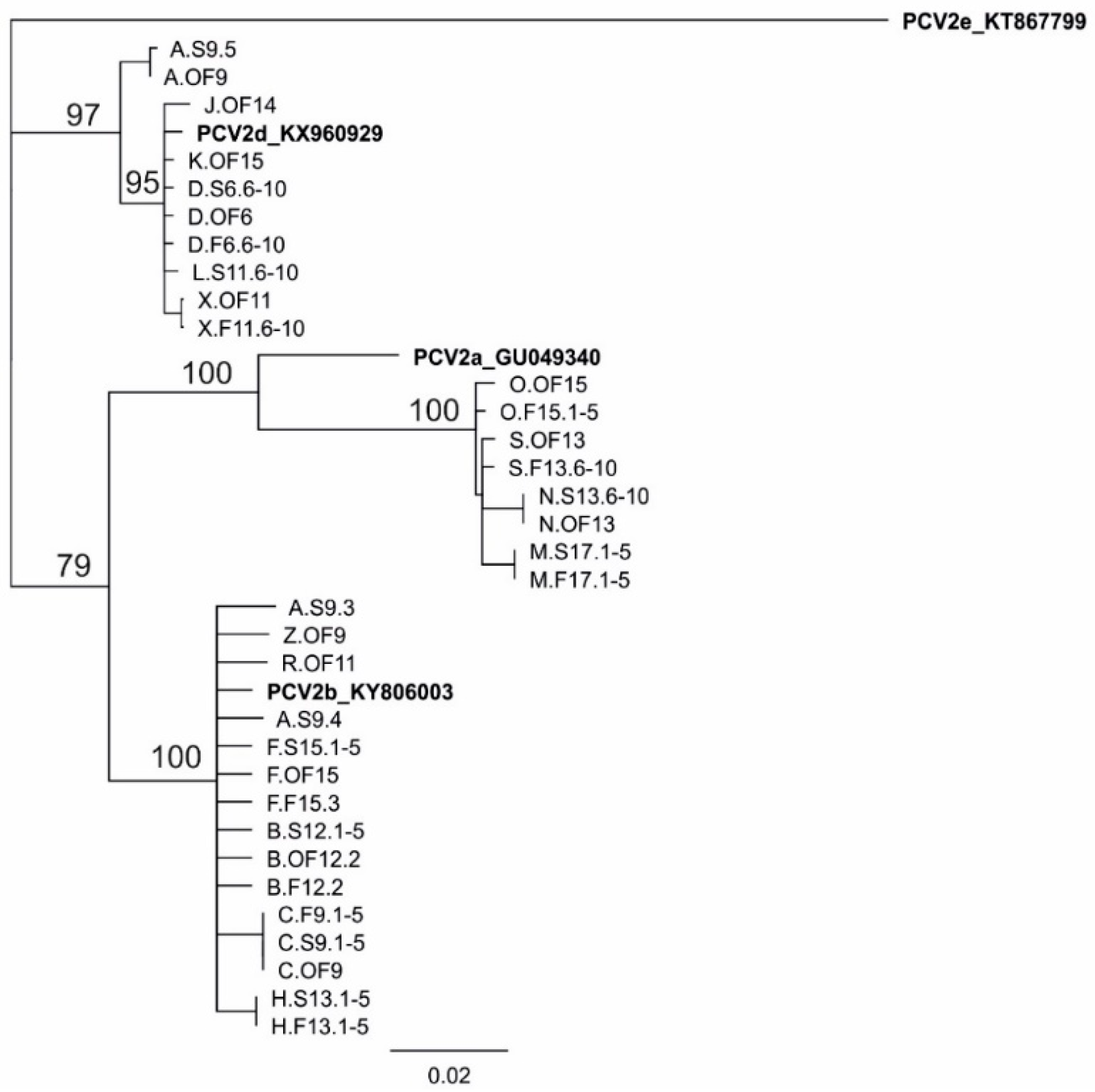
3.5. Genotyping of PCV2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Franzo, G.; Tucciarone, C.M.; Cecchinato, M.; Drigo, M. Porcine circovirus type 2 (PCV2) evolution before and after the vaccination introduction: A large scale epidemiological study. Sci. Rep. 2016, 6, 39458. [Google Scholar] [CrossRef] [PubMed]
- Segales, J. Porcine circovirus type 2 (PCV2) infections: Clinical signs, pathology and laboratory diagnosis. Virus Res. 2012, 164, 10–19. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Meng, X.J.; Halbur, P.G. Porcine circovirus type 2 associated disease: Update on current terminology, clinical manifestations, pathogenesis, diagnosis, and intervention strategies. J. Vet. Diagn. Invest. 2007, 19, 591–615. [Google Scholar] [CrossRef] [PubMed]
- Alarcon, P.; Rushton, J.; Wieland, B. Cost of post-weaning multi-systemic wasting syndrome and porcine circovirus type-2 subclinical infection in England—An economic disease model. Prev. Vet. Med. 2013, 110, 88–102. [Google Scholar] [CrossRef] [Green Version]
- Franzo, G.; Segales, J. Porcine circovirus 2 (PCV-2) genotype update and proposal of a new genotyping methodology. PLoS ONE 2018, 13, e0208585. [Google Scholar] [CrossRef] [Green Version]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Global molecular genetic analysis of porcine circovirus type 2 (PCV2) sequences confirms the presence of four main PCV2 genotypes and reveals a rapid increase of PCV2d. J. Gen. Virol. 2015, 96, 1830–1841. [Google Scholar] [CrossRef]
- Xiao, C.T.; Harmon, K.M.; Halbur, P.G.; Opriessnig, T. PCV2d-2 is the predominant type of PCV2 DNA in pig samples collected in the U.S. during 2014–2016. Vet. Microbiol. 2016, 197, 72–77. [Google Scholar] [CrossRef] [Green Version]
- Fabisiak, M.; Szczotka, A.; Podgórska, K.; Stadejek, T. Prevalence of infection and genetic diversity of porcine circovirus type 2 (PCV2) in wild boar (Sus scrofa) in Poland. J. Wildl. Dis. 2012, 48, 612–618. [Google Scholar] [CrossRef] [Green Version]
- Opriessnig, T.; Xiao, C.T.; Gerber, P.F.; Halbur, P.G. Emergence of a novel mutant PCV2b variant associated with clinical PCVAD in two vaccinated pig farms in the U.S. concurrently infected with PPV2. Vet. Microbiol. 2013, 163, 177–183. [Google Scholar] [CrossRef]
- Opriessnig, T.; Gerber, P.F.; Xiao, C.T.; Halbur, P.G.; Matzinger, S.R.; Meng, X.J. Commercial PCV2a-based vaccines are effective in protecting naturally PCV2b-infected finisher pigs against experimental challenge with a 2012 mutant PCV2. Vaccine 2014, 32, 4342–4348. [Google Scholar] [CrossRef]
- Jeong, J.; Park, C.; Choi, K.; Chae, C. Comparison of three commercial one-dose porcine circovirus type2 (PCV2) vaccines in a herd with concurrent circulation of PCV2b and mutant PCV2b. Vet. Microbiol. 2015, 177, 43–52. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.J.; Fu, Y.J.; Huang, L.P.; Wang, Y.P.; Wei, Y.W.; Wu, H.L.; Liu, C.M. A commercial PCV2a-based vaccine is effective in protection from experimental challenge of PCV2 mutant with two amino acids elongation in capsid protein. Vaccine 2015, 10, e0127859. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, H.G.; Beach, N.M.; Huang, Y.W.; Halbur, P.G.; Meng, M.J.; Opriessnig, T. Comparison of commercial and experimental porcine circovirus type 2 (PCV2) vaccines using a triple challenge with PCV2, porcine reproductive and respiratory syndrome virus (PRRSV), and porcine parvovirus (PPV). Vaccine 2010, 28, 5960–5966. [Google Scholar] [CrossRef] [PubMed]
- Opriessnig, T.; Madson, D.M.; Prickett, J.R.; Kuhar, D.; Lunney, J.K.; Elsener, J.; Halbur, P.G. Effect of porcine circovirus type 2 (PCV2) vaccination on porcine reproductive and respiratory syndrome virus (PRRSV) and PCV2 coinfection. Vet. Microbiol. 2008, 131, 103–114. [Google Scholar] [CrossRef]
- Opriessnig, T.; Patterson, A.R.; Madson, D.M.; Pal, N.; Halbur, P.G. Comparison of efficacy of commercial one dose and two dose PCV2 vaccines using a mixed PRRSV–PCV2–SIV clinical infection model 2–3-months post vaccination. Vaccine 2009, 27, 1002–1007. [Google Scholar] [CrossRef] [PubMed]
- Kristensen, C.S.; Baadsgaard, N.P.; Toft, N. A meta-analysis comparing the effect of PCV2 vaccines on average daily weight gain and mortality rate in pigs from weaning to slaughter. Prev. Vet. Med. 2011, 98, 250–258. [Google Scholar] [CrossRef] [PubMed]
- da Silva, N.; Carriquiry, A.; O’Neill, K.; Opriessnig, T.; O’Connor, A.M. Mixed treatment comparison meta-analysis of porcine circovirus type 2 (PCV2) vaccines used in piglets. Prev. Vet. Med. 2014, 117, 413–424. [Google Scholar] [CrossRef]
- Kixmöller, M.; Ritzmann, M.; Eddicks, M.; Saalmüller, A.; Elbers, K.; Fachinger, V. Reduction of PMWS-associated clinical signs and co-infections by vaccination against PCV2. Vaccine 2008, 26, 3443–3451. [Google Scholar]
- Fachinger, V.; Bischoff, R.; Jedidia, S.B.; Saalmüller, A.; Elbers, K. The effect of vaccination against porcine circovirus type 2 in pigs suffering from porcine respiratory disease complex. Vaccine 2008, 26, 1488–1499. [Google Scholar] [CrossRef]
- Segales, J.; Urniza, A.; Alegre, A.; Bru, T.; Crisci, E.; Nofrarias, M.; Lopez-Soria, S.; Balasch, M.; Sibilia, M.; Xu, Z.; et al. A genetically engineered chimeric vaccine against porcine circovirus type 2 (PCV2) improves clinical, pathological and virological outcomes in postweaning multisystemic wasting syndrome affected farms. Vaccine 2009, 27, 7313–7321. [Google Scholar] [CrossRef]
- Beach, N.M.; Meng, X.J. Efficacy and future prospects of commer- cially available and experimental vaccines against porcine circovirus type 2 (PCV2). Virus Res. 2012, 164, 33–42. [Google Scholar] [CrossRef] [PubMed]
- Gerber, P.F.; Johnson, J.; Shen, H.; Striegel, D.; Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Association of concurrent porcine circovirus (PCV) 2a and 2b infection with PCV associated disease in vaccinated pigs. Res. Vet. Sci. 2013, 95, 775–781. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Oh, T.; Yang, S.; Cho, H.; Kang, I.; Chae, C. Evaluation of a porcine circovirus type 2a (PCV2a) vaccine efficacy against experimental PCV2a, PCV2b, and PCV2d challenge. Vet. Microbiol. 2019, 231, 87–92. [Google Scholar] [CrossRef] [PubMed]
- Ellis, J. Porcine circovirus: A historical perspective. Vet. Pathol. 2014, 51, 315–327. [Google Scholar] [CrossRef]
- Pejsak, Z.; Podgórska, K.; Truszczyński, M.; Karbowiak, P.; Stadejek, T. Efficacy of different protocols of vaccination against porcine circovirus type 2 (PCV2) in a farm affected by postweaning multisystemic wasting disease (PMWS). Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, e1–e5. [Google Scholar] [CrossRef]
- Czyzewska-Dors, E.; Dors, A.; Pomorska-Mol, M.; Podgórska, K.; Pejsak, Z. Efficacy of the Porcine circovirus 2 (PCV2) vaccination under field conditions. Vet. Ital. 2018, 54, 219–224. [Google Scholar] [CrossRef]
- Guo, L.; Fu, Y.; Wang, Y.; Lu, Y.; Wei, Y.; Tang, Q.; Fan, P.; Liu, J.; Zhang, L.; Zhang, F.; et al. A porcie circovirus type 2 (PCV2) mutant with 234 amino acids in Capsid protein showed more virulence in vivo, compared with classical PCV2a/b strain. PLoS ONE 2012, 7, e41463. [Google Scholar]
- Eddicks, M.; Fux, R.; Szikora, F.; Eddicks, L.; Majzoub-Altweck, M.; Hermanns, W.; Sutter, G.; Palzer, A.; Banholzer, E.; Ritzmann, M. Detection of a new cluster of porcine circovirus type 2b strains in domestic pigs in Germany. Vet. Microbiol. 2015, 176, 337–343. [Google Scholar] [CrossRef]
- Seo, H.W.; Park, C.; Kang, I.; Choi, K.; Jeong, J.; Park, S.J.; Chae, C. Genetic and antigenic characterization of a newly emerging porcine circovirus type 2b mutant first isolated in cases of vaccine failure in Korea. Arch. Virol. 2014, 159, 3107–3111. [Google Scholar] [CrossRef]
- Xiao, C.T.; Halbur, P.G.; Opriessnig, T. Complete genome sequence of a novel porcine circovirus type 2b variant present in cases of vaccine failures in the United States. J. Virol. 2012, 86, 12469–12473. [Google Scholar] [CrossRef] [Green Version]
- Sorden, S. Update on porcine circovirus and postweaning multisystemic wasting syndrome (PMWS). Swine Health Prod. 2000, 8, 133–136. [Google Scholar]
- Opriessning, T.; Prickett, J.R.; Madson, D.M.; Shen, H.; Juhan, N.M.; Pogranichniy, R.M.; Meng, X.; Halbur, P.G. Porcine circovirus type 2 (PCV2)-infection and re-inoculation with homologous or heterologous strains: Virological, serological, pathological and clinical effects in growing pigs. Vet. Res. 2010, 41, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wozniak, A.; Milek, D.; Baska, P.; Stadejek, T. Does porcine circovirus type 3 (PCV3) interfere with porcine circovirus type 2 (PCV2) vaccine efficacy? Transbound. Emerg. Dis. 2019, 66, 1454–1461. [Google Scholar] [CrossRef] [PubMed]
- Oliver-Ferrando, S.; Segalés, J.; López-Soria, S.; Callén, A.; Merdy, O.; Joisel, F.; Sibila, M. Evaluation of natural porcine circovirus type 2 (PCV2) subclinical infection and seroconversion dynamics in piglets vaccinated at different ages. Vet. Res. 2016, 47, 121. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing: Vienna, Austria, 2018. Available online: https://www/R-project.org/ (accessed on 1 September 2019).
- Harrell, F.E. rms: Regression Modeling Strategies. R Package Version 5.1-3. 2019. Available online: https://CRAN.R-project.org/package=rms (accessed on 1 September 2019).
- Afghah, Z.; Webb, B.; Meng, X.; Ramamoorthy, S. Ten years of PCV2 vaccines and vaccination: Is eradication a possibility? Vet. Microbiol. 2017, 206, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Rodrigez, A.; Dewulf, J.; Meyns, T.; Del-Pozo-Sacristán, R.; Andreoni, C.; Goubier, A.; Chapat, L.; Charreyre, C.; Joisel, F.; Maes, D. Effect of sow vaccination against porcine circovirus type 2 (PCV2) on virological profiles in herds with or without PCV2 systemic disease. Can. Vet. J. 2016, 57, 619–628. [Google Scholar]
- Yan, M.; Zhu, L.; Yang, Q. Infection of porcine circovirus 2 (PCV2) in intestinal porcine epithelial cell line (IPEC-J2) and interaction between PCV2 and IPEC-J2 microfilaments. Virol. J. 2014, 11, 193. [Google Scholar] [CrossRef] [Green Version]
- Prickett, J.R.; Zimmerman, J.J. The development of oral fluid-based diagnostics and applications in veterinary medicine. Anim. Health Rev. 2010, 11, 201–216. [Google Scholar] [CrossRef]
- Ramirez, A.; Wang, C.; Prickett, J.R.; Pogranichniy, R.; Yoon, K.J.; Main, R.; Johnson, J.K.; Rademacher, C.; Hoogland, M.; Hoffmann, P.; et al. Efficient surveillance of pig populations using oral fluids’. Prev. Vet. Med. 2012, 104, 292–300. [Google Scholar] [CrossRef]
- Hernandez-Garcia, J.; Robben, N.; Magnée, D.; Eley, T.; Dennis, I.; Kayes, S.M.; Thomson, J.R.; Tucker, A.W. The use of oral fluids to monitor key pathogens in porcine respiratory disease complex. Porc. Health Manag. 2017, 3, 7. [Google Scholar] [CrossRef] [Green Version]
- Fraile, L.; Sibila, M.; Nofrarías, M.; López-Jimenez, R.; Huerta, E.; Llorens, A.; López-Soria, S.; Pérez, D.; Segalés, J. Effect of sow and piglet porcine circovirus type 2 (PCV2) vaccination on piglet mortality, viraemia, antibody titre and production parameters. Vet. Microbiol. 2012, 161, 229–234. [Google Scholar] [CrossRef] [PubMed]
- Martelli, P.; Saleri, R.; Ferrarini, G.; De Angelis, E.; Cavalli, V.; Benetti, M.; Ferrari, L.; Canelli, E.; Bonilauri, P.; Arioli, E.; et al. Impact of maternally derived immunity on piglets’ immune response and protection against porcine circovirus type 2 (PCV2) after vaccination against PCV2 at different age. BMC Vet. Res. 2016, 12, 77. [Google Scholar] [CrossRef] [Green Version]
- Eddicks, M.; Koeppen, M.; Willi, S.; Fux, R.; Reese, S.; Sutter, G.; Stadler, J.; Ritzmann, M. Low prevalence of porcine circovirus type 2 infections in farrowing sows and corresponding pre-suckling piglets in southern German pig farms. Vet. Microbiol. 2016, 187, 70–74. [Google Scholar] [CrossRef] [PubMed]
- Dvorak, C.M.T.; Yang, Y.; Haley, C.; Sharma, N.; Murtaugh, M.P. National reduction in porcine circovirus type 2 prevalence following introduction of vaccination. Vet. Microbiol. 2016, 189, 86–90. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, H.; Blanco, G.; Segales, J.; Sibila, M. Can Porcine circovirus type 2 (PCV2) infection be eradicated by mass vaccination? Vet. Microbiol. 2014, 172, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Fenaux, M.; Halbur, P.G.; Gill, M.; Toth, T.E.; Meng, X.J. Genetic characterization of type 2 porcine circovirus (PCV2) from pigs with postweaning multisystemic wasting syndrome in different geographic regions of North America and development of a differential PCR restriction fragment length polymorphism assay to detect and differentiate between infections with PCV1 and PCV2. J. Clin. Microbiol. 2010, 38, 2494–2503. [Google Scholar]
- Patterson, A.R.; Opriessnig, T. Epidemiology and horizontal transmission of porcine circovirus type 2 (PCV2). Anim. Health Res. Rev. 2010, 11, 217–234. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Vaccination Against PCV2 Protocol | Farm ID | Health Status and Clinical Signs | % of PCV2-Positive Pools | PCV2 Viral Load | % of PCV2a-Positive Pools | % of PCV2b/d-Positive Pools | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| (Positive/All Tested) | (Minimum–Maximum, Median) (log10 copies/mL) | (Positive/PCV2-Positive) | (Positive/PCV2-Positive) | ||||||||||||
| S | F | OF | S | F | OF | S | F | OF | S | F | OF | ||||
| NON-VACCINATED | A | poor | 43.8 | 100 | 45.5 | 3.9-6.7, | 4.1–7.1, | 4.3–5.9, | neg | 7.3 | 20.0 | 100 | 100 | 100 | |
| PCVD | (7/16) | (16/16) ** | (5/11) ** | 5.3 | 5.2 | 5.4 | (1/16) | (1/5) | (7/7) | (16/16) | (5/5) | ||||
| B | poor | 57.1 | 57.1 | 50.0 | 6.2–7.6, | 6.3–8.1, | 5.1–8.2, | neg | neg | neg | 100 | 100 | 100 | ||
| PCVD | (8/14) | (8/14) | (4/8) | 6.9 | 6.9 | 6.5 | (8/8) | (8/8) | (4/4) | ||||||
| C | poor | 83.3 | 100 | 83.3 | 3.8–8.5, | 4.4–10, | 7.1–8.3, | neg | neg | neg | 100 | 100 | 100 | ||
| PCVD | (10/12) | (12/12) | (5/6) | 6.2 | 6.8 | 7.2 | (10/10) | (12/12) | (5/5) | ||||||
| D | poor | 80.0 | 80.0 | 100 | 3.9–7.4, | 4.8–7.3, | 4.3–8.6, | neg | neg | neg | 100 | 100 | 100 | ||
| PCVD | (8/10) | (8/10) | (6/6) | 6.0 | 6.1 | 7.2 | (8/8) | (8/8) | (6/6) | ||||||
| VACCINATED | VAC1 (vaccination of piglets) | E | very good | neg | neg | 12.5 (1/8) | neg | neg | 5.1 | neg | neg | neg | neg | neg | 100 (1/1) |
| F | average | 66.7 | 91.7 | 100 | 4.2–8.7, | 4.1–5.7, | 4.7–6.8, | neg | neg | neg | 100 (8/8) | 100 (11/11) | 100 (6/6) | ||
| PCVD | (8/12) | (11/12) | (6/6) | 6.0 | 5.1 | 5.9 | |||||||||
| G | average | neg | neg | 22.2 | neg | neg | 4.2–4.4, | neg | neg | neg | neg | neg | 100 | ||
| PCVD | (2/9) | 4.3 | (2/2) | ||||||||||||
| H | average | 62.5 | 75.0 | 75.0 | 4.3–7.6, | 4.0–6.3, | 5.8–7.0, | neg | neg | neg | 100 | 100 | 100 | ||
| PCVD | (10/16) | (12/16) | (6/8) | 5.3 | 4.6 | 6.4 | (10/10) | (12/12) | (6/6) | ||||||
| I | good | 7.1 | 7.1 | neg | 4.2 | 4.1 | neg | neg | 100 | neg | 100 | neg | neg | ||
| (1/14) | (1/14) | (1/1) | (1/1) | ||||||||||||
| J | average | 58.3 | 75.0 | 100 | 3.7–5.6, | 3.9–5.6, | 5.5–6.8, | neg | neg | neg | 100 | 100 | 100 | ||
| (7/12) | (9/12) | (7/7) | 5.2 | 4.8 | 6.1 | (7/7) | (9/9) | (7/7) | |||||||
| K | average | 6.3 | 37.5 | 80.0 | 5.2 | 4.0–5.0, | 3.8–6.0, | neg | neg | 25.0 | 100 | 100 | 87.5 | ||
| (1/16) | (6/16) | (8/10) ** | 4.5 | 4.8 | (2/8) | (1/1) | (6/6) | (7/8) | |||||||
| L | average | 56.3 | 81.3 | 80.0 | 3.9–7.4, | 3.8–6.3, | 4.3–7.0, | 44.4 | 61.5 | 12.5 | 77.8 | 76.9 | 100 | ||
| (9/16) ** | (13/16) ** | (8/10) ** | 5.8 | 4.9 | 6.3 | (4/9) | (8/13) | (1/8) | (7/9) | (10/13) | (8/8) | ||||
| M | average | 12.5 | 25.0 | 50.0 | 5.9–6.0, | 4.3–6.1, | 4.2-7.3, | 100 | 100 | 100 | neg | neg | neg | ||
| (2/16) | (4/16) | (4/8) | 5.9 | 5.3 | 5.1 | (2/2) | (4/4) | (4/4) | |||||||
| N | average | 31.3 | 37.5 | 44.4 | 4.6–5.8, | 3.8–7.7, | 4.4–6.3, | 80.0 | 100 | 75.0 | 20 | neg | 25.0 | ||
| (5/16) * | (6/16) | (4/9) * | 4.8 | 6.3 | 5.9 | (4/5) | (6/6) | (3/4) | (1/5) | (1/4) | |||||
| O | average | 18.8 | 18.8 | 33.3 | 4.5–5.3, | 5.2–5.4, | 3.7–7.3, | 100 | 100 | 100 | neg | neg | neg | ||
| (3/16) | (3/16) | (3/9) | 4.7 | 5.3 | 6.1 | (3/3) | (3/3) | (3/3) | |||||||
| VAC2 (vaccination of sows and their progeny) | P | very good | 6.3 | neg | 16.7 | 3.8 | neg | neg | neg | neg | 100 | 100 | neg | neg | |
| (1/16) | (1/6) | (1/1) | (1/1) | ||||||||||||
| Q | average | neg | neg | neg | neg | neg | neg | neg | neg | neg | neg | neg | neg | ||
| PCVD | |||||||||||||||
| R | average | neg | 40.0 | 100 | neg | 3.8–4.8, | 5.0–6.1, | neg | neg | neg | neg | 100 | 100 | ||
| PCVD | (4/10) | (5/5) | 4.2 | 5.2 | (4/4) | (5/5) | |||||||||
| S | average | neg | 12.5 | 22.2 | neg | 5.7–5.8, | 4.4–7.4, | neg | 100 | 50 | neg | neg | 50 | ||
| (2/2) | (1/2) | (1/2) | |||||||||||||
| (2/16) | (2/9) * | 5.8 | 5.9 | ||||||||||||
| T | average | neg | neg | 14.3 | neg | neg | 4.5 | neg | neg | neg | neg | neg | 100 | ||
| (1/7) | (1/1) | ||||||||||||||
| U | average | 28.6 | 66.7 | 83.3 | 3.9–6.5, | 3.8–5.0, | 4.3–4.9, | neg | neg | neg | 100 | 100 | 100 | ||
| (4/14) | (8/12) | (5/6) | 4.8 | 4.1 | 4.5 | (4/4) | (8/8) | (5/5) | |||||||
| V | average | neg | 16.7 | 28.6 | neg | 3.8–3.9, | 3.7–3.9, | neg | neg | neg | neg | 100 | 100 (2/2) | ||
| (2/12) | (2/7) | 3.9 | 3.8 | ||||||||||||
| W | average | neg | 8.3 | 16.7 | neg | 4.3 | 3.8 | neg | neg | neg | neg | 100 | 100 | ||
| (1/12) | (1/6) | (1/1) | (1/1) | ||||||||||||
| X | average | 20.0 | 46.7 | 64.3 | 3.9–5.1, | 3.7–7.6, | 4.0–7.1, | neg | neg | neg | 100 | 100 | 100 | ||
| PCVD | (3/15) | (7/15) | (9/14) | 4.8 | 5.1 | 5.7 | (3/3) | (7/7) | (9/9) | ||||||
| Y | average | neg | neg | 11.1 | neg | neg | 3.9 | neg | neg | 100 | neg | neg | neg | ||
| (1/11) | (1/1) | ||||||||||||||
| Z | average | neg | 66.7 | 85.7 | neg | 3.8–4.8, | 4.3–5.9, | neg | neg | neg | neg | 100 | 100 | ||
| (8/8) | (6/6) | ||||||||||||||
| PCVD | (8/12) | (6/7) | 4.4 | 5.4 | |||||||||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Woźniak, A.; Miłek, D.; Matyba, P.; Stadejek, T. Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2. Viruses 2019, 11, 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121135
Woźniak A, Miłek D, Matyba P, Stadejek T. Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2. Viruses. 2019; 11(12):1135. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121135
Chicago/Turabian StyleWoźniak, Aleksandra, Dagmara Miłek, Piotr Matyba, and Tomasz Stadejek. 2019. "Real-Time PCR Detection Patterns of Porcine Circovirus Type 2 (PCV2) in Polish Farms with Different Statuses of Vaccination against PCV2" Viruses 11, no. 12: 1135. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121135