Feline Infectious Peritonitis as a Systemic Inflammatory Disease: Contribution of Liver and Heart to the Pathogenesis

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Tissue Processing
2.1.1. Liver Study (Groups 1.1–1.3)
2.1.2. Heart Study (Groups 2.1–2.3)
2.2. Assessment of Cytokine Transcription
2.2.1. Reverse Transcription and Quantitative Polymerase Chain Reaction (RT-qPCR) for Feline Cytokines
2.2.2. Laser Capture Microdissection (LCM)
2.2.3. Relative Quantification of Cytokine Transcripts and Statistical Analysis
2.3. Assessment of Cytokine Protein Expression
2.3.1. Antibodies Specific for Feline Cytokines
2.3.2. Immunohistology for Feline Cytokines
3. Results
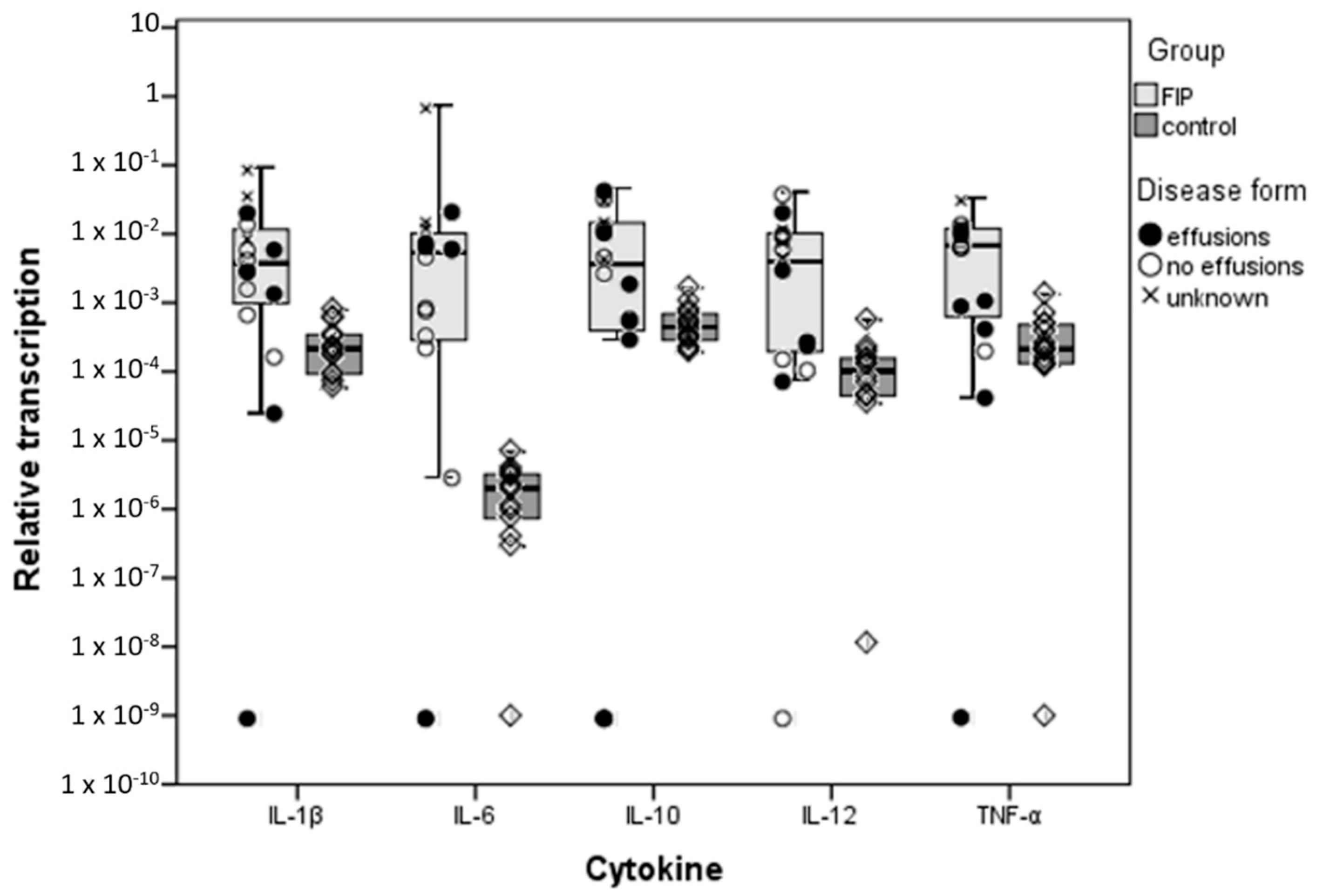
3.1. Inflammatory Cytokine Transcription in the Liver
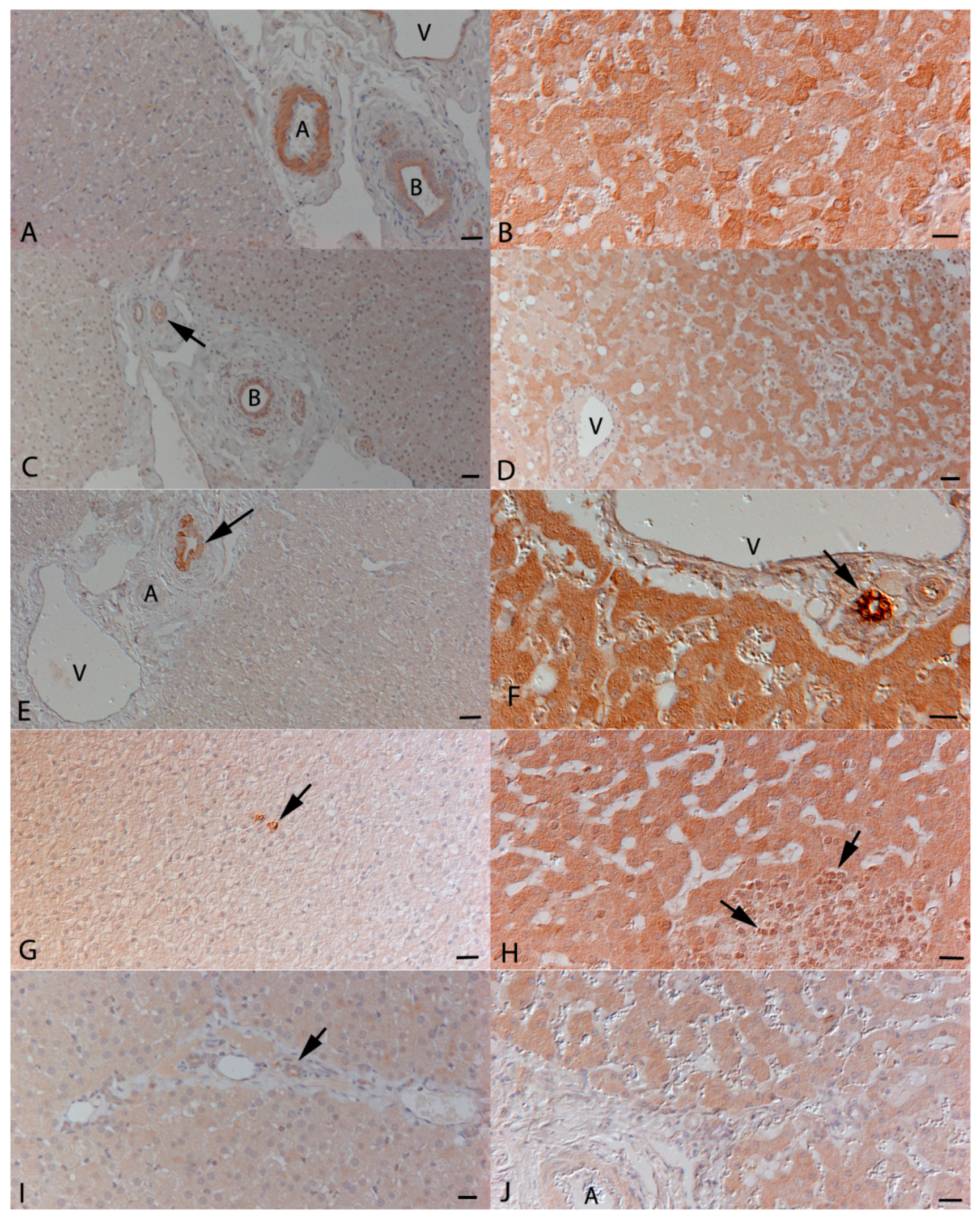
3.2. Hepatocytes Are a Source of Inflammatory Cytokines in FIP
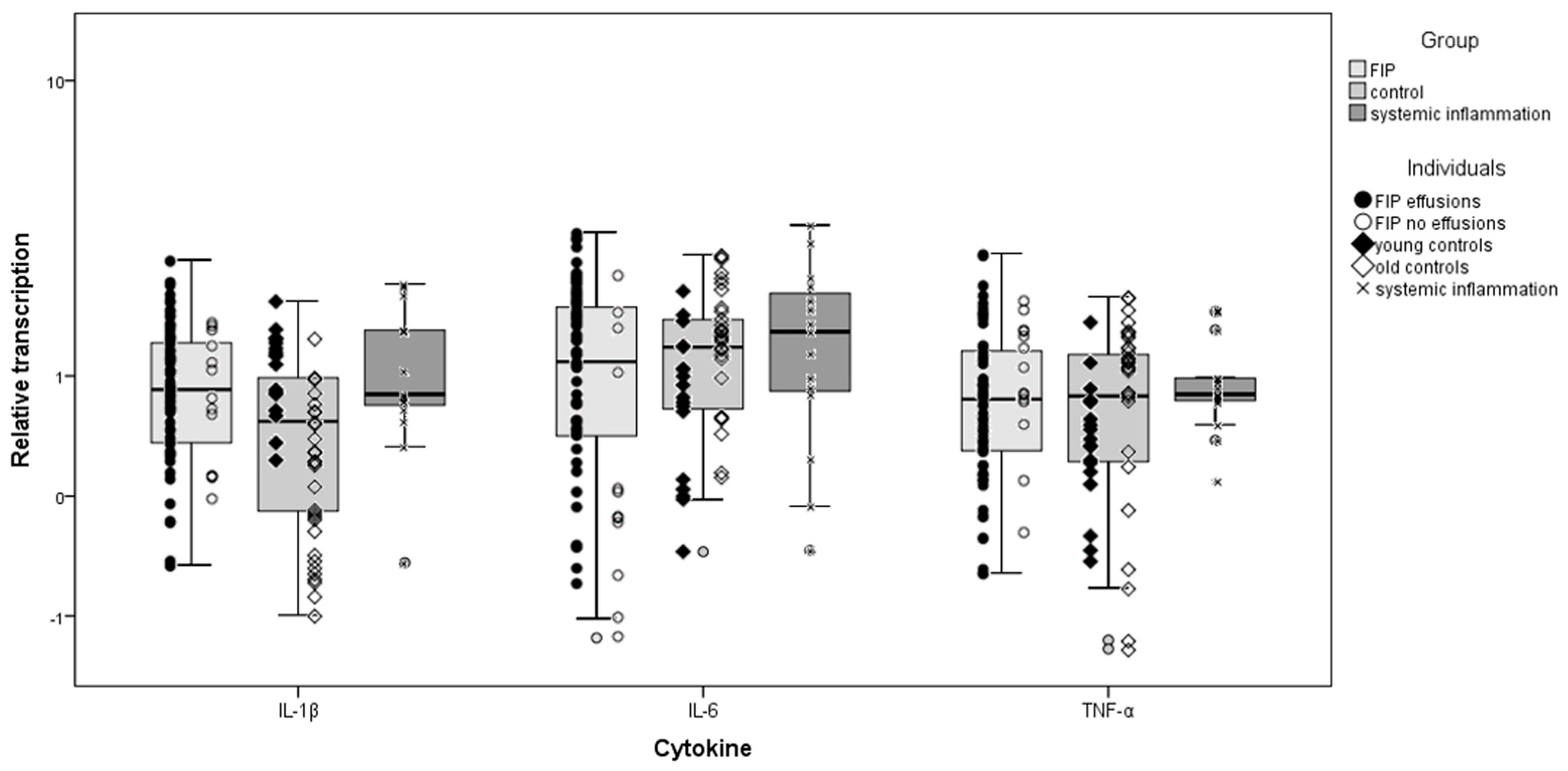
3.3. Inflammatory Cytokine Transcription in the Heart
3.4. Cardiomyocytes Themselves Contribute to Cytokine Expression
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hayashi, T.; Goto, N.; Takahashi, R.; Fujiwara, K. Systemic vascular lesions in feline infectious peritonitis. Jpn. J. Vet. Sci. 1977, 39, 365–377. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, R.C.; Scott, F.W. Pathogenesis of feline infectious peritonitis: Pathologic changes and immunofluorescence. Am. J. Vet. Res. 1981, 42, 2036–2048. [Google Scholar] [PubMed]
- Kipar, A.; Bellmann, S.; Kremendahl, J.; Köhler, K.; Reinacher, M. Cellular composition, coronavirus antigen expression and production of specific antibodies in lesions in feline infectious peritonitis. Vet. Immunol. Immunopathol. 1998, 65, 243–257. [Google Scholar] [CrossRef]
- Kipar, A.; May, H.; Menger, S.; Weber, M.; Leukert, W.; Reinacher, M. Morphologic features and development of granulomatous vasculitis in feline infectious peritonitis. Vet. Pathol. 2005, 42, 321–330. [Google Scholar] [CrossRef] [PubMed]
- Acar, D.D.; Olyslaegers, D.A.J.; Dedeurwaerder, A.; Roukaerts, I.D.M.; Baetens, W.; Van Bockstael, S.; De Gryse, G.M.A.; Desmarets, L.M.B.; Nauwynck, H.J. Upregulation of endothelial cell adhesion molecules characterizes veins close to granulomatous infiltrates in the renal cortex of cats with feline infectious peritonitis and is indirectly triggered by feline infectious peritonitis virus-infected monocytes. J. Gen. Virol. 2016, 97, 2633–2642. [Google Scholar] [CrossRef] [PubMed]
- Dinarello, C.A. Historical Review of Cytokines. Eur. J. Immunol. 2007, 37, S34–S45. [Google Scholar] [CrossRef] [Green Version]
- Akdis, M.; Burgler, S.; Crameri, R.; Eiwegger, T.; Fujita, H.; Gomez, E.; Klunker, S.; Meyer, N.; O’Mahony, L.; Palomares, O.; et al. Interleukins, from 1 to 37, and interferon-γ: Receptors, functions, and roles in diseases. J. Allergy Clin. Immunol. 2011, 127, 701–721. [Google Scholar] [CrossRef]
- Paltrinieri, S. The feline acute phase reaction. Vet. J. 2008, 177, 26–35. [Google Scholar] [CrossRef]
- Goitsuka, R.; Ohashi, T.; Ono, K.; Yasukawa, K.; Koishibara, Y.; Fukui, H.; Ohsugi, Y.; Hasegawa, A. IL-6 activity in feline infectious peritonitis. J. Immunol. 1990, 144, 2599–2603. [Google Scholar]
- Goitsuka, R.; Onda, C.; Hirota, Y.; Hasegawa, A.; Tomoda, I. Feline interleukin 1 production induced by feline infectious peritonitis virus. Jpn. J. Vet. Sci. 1988, 50, 209–214. [Google Scholar] [CrossRef]
- Gunn-Moore, D.A.; Caney, S.M.; Gruffydd-Jones, T.J.; Helps, C.R.; Harbour, D.A. Antibody and cytokine responses in kittens during the development of feline infectious peritonitis (FIP). Vet. Immunol. Immunopathol. 1998, 65, 221–242. [Google Scholar] [CrossRef]
- Kiss, I.; Poland, A.M.; Pedersen, N.C. Disease outcome and cytokine responses in cats immunized with an avirulent feline infectious peritonitis virus (FIPV)-UCD1 and challenge-exposed with virulent FIPV-UCD8. J. Feline Med. Surg. 2004, 6, 89–97. [Google Scholar] [CrossRef] [PubMed]
- Gelain, M.E.; Meli, M.; Paltrinieri, S. Whole blood cytokine profiles in cats infected by feline coronavirus and healthy non-FCoV infected specific pathogen-free cats. J. Feline Med. Surg. 2006, 8, 389–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehrbod, P.; Harun, M.S.R.; Shuid, A.N.; Omar, A.R. Transcriptome analysis of feline infectious peritonitis virus infection. Coronaviruses Methods Protoc. Methods Mol. Biol. 2015, 1282, 241–250. [Google Scholar]
- Harun, M.S.R.; Kuan, C.O.; Selvarajah, G.T.; Wei, T.S.; Arshad, S.S.; Hair Bejo, M.; Omar, A.R. Transcriptional profiling of feline infectious peritonitis virus infection in CRFK cells and in PBMCs from FIP diagnosed cats. Virol. J. 2013, 10, 329. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Hohdatsu, T.; Toda, A.; Tanabe, M.; Koyama, H. TNF-alpha, produced by feline infectious peritonitis virus (FIPV)-infected macrophages, upregulates expression of type II FIPV receptor feline aminopeptidase N in feline macrophages. Virology 2007, 364, 64–72. [Google Scholar] [CrossRef] [Green Version]
- Takano, T.; Hohdatsu, T.; Hashida, Y.; Kaneko, Y.; Tanabe, M.; Koyama, H. A “possible” involvement of TNF-alpha in apoptosis induction in peripheral blood lymphocytes of cats with feline infectious peritonitis. Vet. Microbiol. 2007, 119, 121–131. [Google Scholar] [CrossRef]
- Haagmans, B.L.; Egberink, H.F.; Horzinek, M.C. Apoptosis and T-cell depletion during feline infectious peritonitis. J. Virol. 1996, 70, 8977–8983. [Google Scholar]
- Kipar, A.; Köhler, K.; Leukert, W.; Reinacher, M. A comparison of lymphatic tissues from cats with spontaneous feline infectious peritonitis (FIP), cats with FIP virus infection but no FIP, and cats with no infection. J. Comp. Pathol. 2001, 125, 182–191. [Google Scholar] [CrossRef]
- Malbon, A.J.; Meli, M.L.; Barker, E.N.; Davidson, A.D.; Tasker, S.; Kipar, A. Inflammatory mediators in the mesenteric lymph nodes, site of a possible intermediate phase in the immune response to feline coronavirus and the pathogenesis of feline infectious peritonitis? J. Comp. Pathol. 2019, 166, 69–86. [Google Scholar] [CrossRef]
- Kipar, A.; Meli, M.L.; Failing, K.; Euler, T.; Gomes-Keller, M.A.; Schwartz, D.; Lutz, H.; Reinacher, M. Natural feline coronavirus infection: Differences in cytokine patterns in association with the outcome of infection. Vet. Immunol. Immunopathol. 2006, 112, 141–155. [Google Scholar] [CrossRef] [PubMed]
- Duthie, S.; Eckersall, P.D.; Addie, D.D.; Lawrence, C.E.; Jarrett, O. Value of alpha 1-acid glycoprotein in the diagnosis of feline infectious peritonitis. Vet. Rec. 1997, 141, 299–303. [Google Scholar] [CrossRef] [PubMed]
- Giordano, A.; Spagnolo, V.; Colombo, A.; Paltrinieri, S. Changes in some acute phase protein and immunoglobulin concentrations in cats affected by feline infectious peritonitis or exposed to feline coronavirus infection. Vet. J. 2004, 167, 38–44. [Google Scholar] [CrossRef]
- Giori, L.; Giordano, A.; Giudice, C.; Grieco, V.; Paltrinieri, S. Performances of different diagnostic tests for feline infectious peritonitis in challenging clinical cases. J. Small Anim. Pract. 2011, 52, 152–157. [Google Scholar] [CrossRef] [PubMed]
- González-Amaro, R.; García-Monzón, C.; García-Buey, L.; Moreno-Otero, R.; Alonso, J.L.; Yagüe, E.; Pivel, J.P.; López-Cabrera, M.; Fernández-Ruiz, E.; Sánchez-Madrid, F. Induction of tumor necrosis factor alpha production by human hepatocytes in chronic viral hepatitis. J. Exp. Med. 1994, 179, 841–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alfrey, E.J.; Most, D.; Wang, X.; Lee, L.K.; Holm, B.; Krieger, N.R.; Sibley, R.K.; Huie, P.; Dafoe, D.C. Interferon-gamma and interleukin-10 messenger RNA are up-regulated after orthotopic liver transplantation in tolerant rats: Evidence for cytokine-mediated immune dysregulation. Surgery 1995, 118, 399–405. [Google Scholar] [CrossRef]
- Saad, B.; Frei, K.; Scholl, F.A.; Fontana, A.; Maier, P. Hepatocyte-derived interleukin-6 and tumor-necrosis factor alpha mediate the lipopolysaccharide-induced acute-phase response and nitric oxide release by cultured rat hepatocytes. Eur. J. Biochem. 1995, 229, 349–355. [Google Scholar] [CrossRef]
- Panesar, N.; Tolman, K.; Mazuski, J.E. Endotoxin stimulates hepatocyte interleukin-6 production. J. Surg. Res. 1999, 85, 251–258. [Google Scholar] [CrossRef]
- Stonāns, I.; Stonāne, E.; Russwurm, S.; Deigner, H.P.; Böhm, K.J.; Wiederhold, M.; Jäger, L.; Reinhart, K. HepG2 human hepatoma cells express multiple cytokine genes. Cytokine 1999, 11, 151–156. [Google Scholar] [CrossRef]
- Dikopoulos, N.; Wegenka, U.; Kröger, A.; Hauser, H.; Schirmbeck, R.; Reimann, J. Recently Primed CD8+ T Cells Entering the Liver Induce Hepatocytes to Interact with Naïve CD8+ T Cells in the Mouse. Hepatology 2004, 39, 1256–1266. [Google Scholar] [CrossRef]
- Fonfara, S.; Kitz, S.; Hahn, S.; Kipar, A. The influence of age and disease presentation on myocardial transcription in cats with hypertrophic cardiomyopathy. Res. Vet. Sci. in preparation.
- Levy, J.K.; Ritchey, J.W.; Rottman, J.B.; Davidson, M.G.; Liang, Y.-H.; Jordan, H.L.; Tompkins, W.A.; Tompkins, M.B. Elevated interleukin-10–to–interleukin-12 ratio in feline immunodeficiency virus–infected cats predicts loss of type 1 immunity to Toxoplasma gondii. J. Infect. Dis. 1998, 178, 503–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Hemert, S.; Meijerink, M.; Molenaar, D.; Bron, P.A.; De Vos, P.; Kleerebezem, M.; Wells, J.M.; Marco, M.L. Identification of Lactobacillus plantarum genes modulating the cytokine response of human peripheral blood mononuclear cells. BMC Microbiol. 2010, 10, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meli, M.L.; Kipar, A.; Müller, C.; Jenal, K.; Gönczi, E.; Borel, N.; Gunn-Moore, D.; Chalmers, S.; Lin, F.; Reinacher, M.; et al. High viral loads despite absence of clinical and pathological findings in cats experimentally infected with feline coronavirus (FCoV) type I and in naturally FCoV-infected cats. J. Feline Med. Surg. 2004, 6, 69–81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fonfara, S.; Hetzel, U.; Hahn, S.; Kipar, A. Age- and gender-dependent myocardial transcription patterns of cytokines and extracellular matrix remodelling enzymes in cats with non-cardiac diseases. Exp. Gerontol. 2015, 72, 117–123. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leutenegger, C.M.; Mislin, C.N.; Sigrist, B.; Ehrengruber, M.U.; Hofmann-Lehmann, R.; Lutz, H. Quantitative real-time PCR for the measurement of feline cytokine mRNA. Vet. Immunol. Immunopathol. 1999, 71, 291–305. [Google Scholar] [CrossRef] [Green Version]
- Kipar, A.; Leutenegger, C.M.; Hetzel, U.; Akens, M.K.; Mislin, C.N.; Reinacher, M.; Lutz, H. Cytokine mRNA levels in isolated feline monocytes. Vet. Immunol. Immunopathol. 2001, 78, 305–315. [Google Scholar] [CrossRef]
- Cummings, M.; McGinley, C.V.; Wilkinson, N.; Field, S.L.; Duffy, S.R.; Orsi, N.M. A robust RNA integrity-preserving staining protocol for laser capture microdissection of endometrial cancer tissue. Anal. Biochem. 2011, 416, 123–125. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Frontera-Acevedo, K.; Sakamoto, K. Local pulmonary immune responses in domestic cats naturally infected with Cytauxzoon felis. Vet. Immunol. Immunopathol. 2015, 163, 1–7. [Google Scholar] [CrossRef]
- Kipar, A.; Meli, M.L. Feline infectious peritonitis: Still an enigma? Vet. Pathol. 2014, 51, 505–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giambelluca, M.S.; Laflamme, C.; Pouliot, M. Post-transcriptional regulation of tumour necrosis factor alpha biosynthesis: Relevance to the pathophysiology of rheumatoid arthritis. OA Inflamm. 2013, 1, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Kipar, A.; Meli, M.L.; Baptiste, K.E.; Bowker, L.J.; Lutz, H. Sites of feline coronavirus persistence in healthy cats. J. Gen. Virol. 2010, 91, 1698–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Groot-Mijnes, J.D.F.; Van Dun, J.M.; Van Der Most, G.; De Groot, R.J. Natural History of a Recurrent Feline Coronavirus Infection and the Role of Cellular Immunity in Survival and Disease. J. Virol. 2005, 79, 1036–1044. [Google Scholar] [CrossRef] [Green Version]
- Abbas, A.K.; Lichtman, A.H.; Pillai, S. Cellular and Molecular Immunology, 9th ed.; Saunders Elsevier: Amsterdam, The Netherlands, 2017; ISBN 9780323479783. [Google Scholar]
- Olyslaegers, D.A.J.; Dedeurwaerder, A.; Desmarets, L.M.B.; Vermeulen, B.L.; Dewerchin, H.L.; Nauwynck, H.J. Altered expression of adhesion molecules on peripheral blood leukocytes in feline infectious peritonitis. Vet. Microbiol. 2013, 166, 438–449. [Google Scholar] [CrossRef]
- Watanabe, H.; Nakanishi, I.; Yamashita, K.; Hayakawa, T.; Okada, Y. Matrix metalloproteinase-9 (92 kDa gelatinase/type IV collagenase) from U937 monoblastoid cells: Correlation with cellular invasion. J. Cell Sci. 1993, 104 Pt 4, 991–999. [Google Scholar]
- Sarén, P.; Welgus, H.G.; Kovanen, P.T. TNF-alpha and IL-1beta selectively induce expression of 92-kDa gelatinase by human macrophages. J. Immunol. 1996, 157, 4159–4165. [Google Scholar]
- Robinson, S.C.; Scott, K.A.; Balkwill, F.R. Chemokine stimulation of monocyte matrix metalloproteinase-9 requires endogenous TNF-alpha. Eur. J. Immunol. 2002, 32, 404–412. [Google Scholar] [CrossRef]
- Kent, L.W.; Rahemtulla, F.; Hockett, R.D.; Gilleland, R.C.; Michalek, S.M. Effect of lipopolysaccharide and inflammatory cytokines on interleukin-6 production by healthy human gingival fibroblasts. Infect. Immun. 1998, 66, 608–614. [Google Scholar]
- Takano, T.; Ohyama, T.; Kokumoto, A.; Satoh, R.; Hohdatsu, T. Vascular endothelial growth factor (VEGF), produced by feline infectious peritonitis (FIP) virus-infected monocytes and macrophages, induces vascular permeability and effusion in cats with FIP. Virus Res. 2011, 158, 161–168. [Google Scholar] [CrossRef]
- Pedersen, N.C. An update on feline infectious peritonitis: Virology and immunopathogenesis. Vet. J. 2014, 201, 123–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loeffler, S.; Fayard, B.; Weis, J.; Weissenberger, J. Interleukin-6 induces transcriptional activation of vascular endothelial growth factor (VEGF) in astrocytes in vivo and regulates VEGF promoter activity in glioblastoma cells via direct interaction between STAT3 and Sp1. Int. J. Cancer 2005, 115, 202–213. [Google Scholar] [CrossRef] [PubMed]
- Feurino, L.W.; Zhang, Y.; Bharadwaj, U.; Zhang, R.; Li, F.; Fisher, W.E.; Brunicardi, F.C.; Chen, C.; Yao, Q.; Min, L. IL-6 stimulates Th2 type cytokine secretion and upregulates VEGF and NRP-1 expression in pancreatic cancer cells. Cancer Biol. Ther. 2007, 6, 1096–1100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurkan, O.U.; He, C.; Zielinski, R.; Rabb, H.; King, L.S.; Dodd-o, J.M.; D’Alessio, F.R.; Aggarwal, N.; Pearse, D.; Becker, P.M. Interleukin-6 mediates pulmonary vascular permeability in a two-hit model of ventilator-associated lung injury. Exp. Lung Res. 2011, 37, 575–584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kishimoto, T. The biology of interleukin-6. Blood 1989, 74, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwasaki, H.; Takeuchi, O.; Teraguchi, S.; Matsushita, K.; Uehata, T.; Kuniyoshi, K.; Satoh, T.; Saitoh, T.; Matsushita, M.; Standley, D.M.; et al. The IκB kinase complex regulates the stability of cytokine-encoding mRNA induced by TLR-IL-1R by controlling degradation of regnase-1. Nat. Immunol. 2011, 12, 1167–1175. [Google Scholar] [CrossRef]
- Buchwald, U.K.; Geerdes-Fenge, H.F.; Vöckler, J.; Ziege, S.; Lode, H. Interleukin-10: Effects on phagocytosis and adhesion molecule expression of granulocytes and monocytes in a comparison with prednisolone. Eur. J. Med. Res. 1999, 4, 85–94. [Google Scholar]
- de Waal Malefyt, R.; Abrams, J.; Bennett, B.; Figdor, C.G.; de Vries, J.E. Interleukin 10 (IL-10) inhibits cytokine synthesis by human monocytes: An autoregulatory role of IL-10 produced by monocytes. J. Exp. Med. 1991, 174, 1209–1220. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Liu, J.; Chesi, M.; Bergsagel, P.L.; Ho, I.-C.; Donnelly, R.P.; Ma, X. Differential regulation of IL-12 and IL-10 gene expression in macrophages by the basic leucine zipper transcription factor c-Maf fibrosarcoma. J. Immunol. 2002, 169, 5715–5725. [Google Scholar] [CrossRef] [Green Version]
- Watson, D.C.; Sargianou, M.; Panos, G. Interleukin-12 (IL-12)/IL-10 ratio as a marker of disease severity in Crimean-Congo hemorrhagic fever. Clin. Vaccine Immunol. 2012, 19, 823–824. [Google Scholar] [CrossRef] [Green Version]
- Lasarte, J.J.; Corrales, F.J.; Casares, N.; López-Díaz de Cerio, A.; Qian, C.; Xie, X.; Borrás-Cuesta, F.; Prieto, J. Different doses of adenoviral vector expressing IL-12 enhance or depress the immune response to a coadministered antigen: The role of nitric oxide. J. Immunol. 1999, 162, 5270–5277. [Google Scholar] [PubMed]
- Lee, K.; Overwijk, W.W.; O’Toole, M.; Swiniarski, H.; Restifo, N.P.; Dorner, A.J.; Wolf, S.F.; Sturmhoefel, K. Dose-dependent and schedule-dependent effects of interleukin-12 on antigen-specific CD8 responses. J. Interf. Cytokine Res. 2000, 20, 589–596. [Google Scholar]
- Glansbeek, H.L.; Haagmans, B.L.; te Lintelo, E.G.; Egberink, H.F.; Duquesne, V.; Aubert, A.; Horzinek, M.C.; Rottier, P.J.M. Adverse effects of feline IL-12 during DNA vaccination against feline infectious peritonitis virus. J. Gen. Virol. 2002, 83, 1–10. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Cat | Group | Signalment | Lesion Distribution | Presence of Effusions |
|---|---|---|---|---|
| 1 | 1.1a | 6 m, MN, DSH | Peritoneum | Y |
| 2 | 1.1a | 6 m, FE, DSH | Peritoneum | Y |
| 3 | 1.1a | 1.5 y, FN, DLH | Kidney, lung | Y |
| 4 | 1.1a | 5 m, ME, DSH | Kidney, eye, brain | N |
| 5 | 1.1a | Juvenile, FE, BSH | Kidneys, liver, spleen, lung, CNS | N |
| 6 | 1.1a | Juvenile, MN, DSH | Liver, spleen, lungs, peritoneum, pleura | N |
| 7 | 1.1a | 8 m, MN, Burmese | Lung, kidney, brain, eye | N |
| 8 | 1.1a | 1 y, FN, DSH | Lung, brain | N |
| 9 | 1.1a | 2 y, MN, DSH | Lung, kidney, liver, peritoneum | N |
| 10 | 1.1a | 2 y, FE, DSH | NR | NR |
| 11 | 1.1a | 3 y, FN, DLH | NR | NR |
| 12 | 1.1a | 4 y, MN, DSH | NR | NR |
| 13 | 1.1b | 14–16 w, FE, DSH | Heart, lungs, spleen | N |
| 14 | 1.1b | 14–16 w, FE, DSH | Peritoneum, liver, kidney, omentum, spleen | Y |
| 15 | 1.1b | 14–16 w, FE, DSH | Peritoneum, liver, kidney, omentum, spleen | Y |
| 16 | 1.1b | 14–16 w, FE, DSH | Peritoneum, liver, omentum, spleen | Y |
| Cat | Group | Signalment | Lesion Distribution */Disease † | Presence of Effusions |
|---|---|---|---|---|
| 1 | 2.1 | 4 m, ME, Birman | Pleuritis, peritonitis, med ln | Y |
| 2 | 2.1 | 7 m, ME, Devon Rex | Peritoneum, mes ln | Y |
| 3 | 2.1 | 7 m, MN, BSH | Peritoneum, lung, liver, kidney, mes ln | Y |
| 4 | 2.1 | 9 m, FN, DSH | Peritoneum, liver, mes ln, kidney | Y |
| 5 | 2.1 | 1 y, MN, BSH | Pleura, liver, kidney, lung, mes ln | Y |
| 6 | 2.1 | 11 m, ME, Birman | Pleura, peritoneum, liver, mes ln | Y |
| 7 | 2.1 | 11 m, FN, Ragdoll | Peritoneum, intestinal wall, mes ln | Y |
| 8 | 2.1 | 1 y, FN, DSH | Pleura, peritoneum, leptomeninx, lung, liver, kidneys | Y |
| 9 | 2.1 | 1 y, MN, DLH | Peritoneum, liver, spleen, mes ln | Y |
| 10 | 2.1 | 1 y, MN, Birman | Kidney, colon (BALT), liver, ln | Y |
| 11 | 2.1 | 2 y, MN, DSH | Peritoneum, mes ln | Y |
| 12 | 2.1 | 2 y, ME, British Blue | Widespread visceral lesions | Y |
| 13 | 2.1 | 3 y, MN, Siamese | Pleuritis, peritoneum | Y |
| 14 | 2.1 | 3 y, ME, Birman | Pleura | Y |
| 15 | 2.1 | 4 m, ME, DSH | Liver, kidney, mes ln | N |
| 16 | 2.1 | 8 m, MN, Ragdoll | Kidney, liver | N |
| 17 | 2.1 | 9 m, ME, Birman | Brain | N |
| 18 | 2.1 | 1 y, FN, DLH | NR | NR |
| 19 | 2.2a | 2 y, FN, DSH | Nasal polyp | |
| 20 | 2.2a | 2 y, FN, DSH | Oesophageal stricture | |
| 21 | 2.2a | 3 y, FN, DSH | Vertebral disc prolapse | |
| 22 | 2.2a | 3 y, MN, DLH | Lymphocytic cholangiohepatitis | |
| 23 | 2.2b | 9 y, FN, DSH | Unknown, no lesions | |
| 24 | 2.2b | 10 y, FN, DLH | Behavioural | |
| 25 | 2.2b | 10 y, FN, DSH | Behavioural | |
| 26 | 2.2b | 14 y, ME, DSH | Age-related | |
| 27 | 2.2b | 14 y, FN, DSH | Nasal osteosarcoma | |
| 28 | 2.2b | 19 y, FN, DSH | Gastrointestinal stromal tumour | |
| 29 | 2.3 | 5 y, FN, Oriental Shorthair | Chlamydial infection | |
| 30 | 2.3 | 5 y, MN, Oriental Shorthair | Chlamydial infection | |
| 31 | 2.3 | 11 y, FN, DLH | Septic peritonitis |
| p Values for Each Cytokine; * = Significant at 95% CI | |||||
|---|---|---|---|---|---|
| Group Comparison | IL-1β | IL-6 | IL-10 | IL-12p40 | TNF-α |
| 1.1a vs. 1.1b | 0.103 | 0.770 | 0.170 | 0.133 | 0.078 |
| 1.1 vs. 1.2 | 0.002 * | 0.001 * | 0.034 * | 0.001 * | 0.017 * |
| 1.1a vs. 1.2 | 0.000 * | 0.003 * | 0.031 * | 0.002 * | 0.004 * |
| 1.1b vs.1. 2 | 0.721 | 0.012 * | 0.382 | 0.061 | 0.878 |
| 1.1 eff vs. 1.1 no eff | 0.628 | 0.035 * | 1.000 | 0.836 | 0.234 |
| 1.1 eff vs. 1.2 | 0.274 | 0.020 * | 0.312 | 0.009 * | 0.494 |
| 1.1 no eff vs. 1.2 | 0.003 * | 0.025 * | 0.224 | 0.067 | 0.046 * |
| p Values for Each Cytokine; * = Significant at 95% CI | |||
|---|---|---|---|
| Group Comparison | IL-1β | IL-6 | TNF-α |
| 2.1 vs. 2.2 | 0.008 * | 0.000 * | 0.000 * |
| 2.1 vs. 2.2a | 0.165 | 0.122 | 0.000 * |
| 2.1 vs. 2.2b | 0.000 * | 0.000 * | 0.000 * |
| 2.1 vs. 2.3 | 0.474 | 0.355 | 0.614 |
| 2.1 eff vs. 2.1 no eff | 0.256 | 0.885 | 0.155 |
| 2.1 eff vs. 2.2 | 0.001 * | 0.000 * | 0.000 * |
| 2.1 eff vs. 2.2a | 0.410 | 0.093 | 0.000 * |
| 2.1 eff vs. 2.2b | 0.000 * | 0.000 * | 0.000 * |
| 2.1 eff vs. 2.3 | 0.886 | 0.255 | 0.437 |
| 2.1 no eff vs. 2.2 | 0.331 | 0.018 * | 0.001 * |
| 2.1 no eff vs. 2.2a | 0.159 | 0.332 | 0.025 * |
| 2.1 no eff vs. 2.2b | 0.013 * | 0.002 * | 0.000 * |
| 2.1 no eff vs. 2.3 | 0.217 | 0.529 | 0.894 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Malbon, A.J.; Fonfara, S.; Meli, M.L.; Hahn, S.; Egberink, H.; Kipar, A. Feline Infectious Peritonitis as a Systemic Inflammatory Disease: Contribution of Liver and Heart to the Pathogenesis. Viruses 2019, 11, 1144. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121144
Malbon AJ, Fonfara S, Meli ML, Hahn S, Egberink H, Kipar A. Feline Infectious Peritonitis as a Systemic Inflammatory Disease: Contribution of Liver and Heart to the Pathogenesis. Viruses. 2019; 11(12):1144. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121144
Chicago/Turabian StyleMalbon, Alexandra J, Sonja Fonfara, Marina L Meli, Shelley Hahn, Herman Egberink, and Anja Kipar. 2019. "Feline Infectious Peritonitis as a Systemic Inflammatory Disease: Contribution of Liver and Heart to the Pathogenesis" Viruses 11, no. 12: 1144. https://0-doi-org.brum.beds.ac.uk/10.3390/v11121144